
Hybrid DNA/RNA nanostructures with 2′-5′ linkages†

Abstract
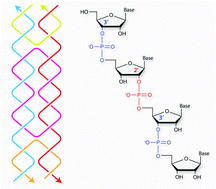
Nucleic acid nanostructures with different chemical compositions have shown utility in biological applications as they provide additional assembly parameters and enhanced stability. The naturally occurring 2′-5′ linkage in RNA is thought to be a prebiotic analogue and has potential use in antisense therapeutics. Here, we report the first instance of DNA/RNA motifs containing 2′-5′ linkages. We synthesized and incorporated RNA strands with 2′-5′ linkages into different DNA motifs with varying number of branch points (a duplex, four arm junction, double crossover motif and tensegrity triangle motif). Using experimental characterization and molecular dynamics simulations, we show that hybrid DNA/RNA nanostructures can accommodate interspersed 2′-5′ linkages with relatively minor effect on the formation of these structures. Further, the modified nanostructures showed improved resistance to ribonuclease cleavage, indicating their potential use in the construction of robust drug delivery vehicles with prolonged stability in physiological conditions.
- This article is part of the themed collection: Nanoscale 2021 Emerging Investigators
 Please wait while we load your content...
Please wait while we load your content...

