
Trophic transfer and biomagnification of fullerenol nanoparticles in an aquatic food chain†

Abstract
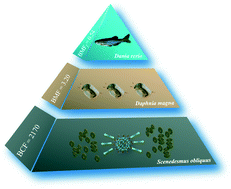
Understanding the trophic transfer and biomagnification potential of nanomaterials in aquatic food chains is crucial for assessing the environmental risks of such materials. To analyse the fate of fullerenols in an aquatic food chain, we quantitatively investigated the bioaccumulation, tissue-specific distribution, depuration, trophic transfer and biomagnification potential of 13C-labelled fullerenols in a three-level aquatic food chain. The fullerenol nanoparticles accumulated in Scenedesmus obliquus through water exposure, and the latter were ingested by Daphnia magna before being transferred to Danio rerio. The tissues of D. rerio were ranked from highest to lowest fullerenol concentration as follows: intestine > liver > muscle > gills > brain. The biomagnification factor (BMF) value of fullerenols from S. obliquus to D. magna was 3.20, while the calculated fitted BMF from D. magna to D. rerio was less than 1 (BMFf = 0.54). Thus, fullerenols were significantly biomagnified from the first to the second trophic level but not from the second to the third trophic level. Although fullerenols eventually enter fish tissues via the food chain, a lack of biomagnification greatly reduce the potential risk to high-trophic-level aquatic organisms. These results will be useful for environmental and ecological risk assessments of carbon nanomaterials.
- This article is part of the themed collection: Celebrating our 2021 Prizewinners
 Please wait while we load your content...
Please wait while we load your content...

