
A two-pronged anti-leukemic agent based on a hyaluronic acid–green tea catechin conjugate for inducing targeted cell death and terminal differentiation†

Abstract
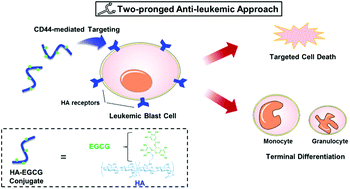
Acute myeloid leukemia (AML) is an aggressive malignancy that leads to a poor prognosis even with intensive chemotherapy. As the key feature of AML is the blockade of hematopoietic cell maturation, considerable attention has been paid to ‘differentiation therapy’ aimed at transforming AML cells into more mature, benign phenotypes using pharmacological agents. Here we report a hyaluronic acid–(−)-epigallocatechin-3-O-gallate (HA–EGCG) conjugate as a unique anti-leukemic agent, capable of selectively killing AML cells as well as promoting their terminal differentiation into monocytes and granulocytes. This ‘two-pronged’ effect of the HA–EGCG conjugate was demonstrated in two different AML cell lines (NB4 and HL60), but absent in a physical mixture (HA + EGCG), highlighting the importance of HA conjugation for targeting of EGCG moieties to AML cells. Moreover, administration of the HA–EGCG conjugate not only suppressed AML progression, but also prolonged survival in the HL60 xenograft mouse model. Our study suggests new opportunities for designing two-pronged anti-leukemic agents for more effective AML treatment.
 Please wait while we load your content...
Please wait while we load your content...

