
Size-dependent enrichment of leukocytes from undiluted whole blood using shear-induced diffusion†
 ab
and
Ian
Papautsky
*ab
ab
and
Ian
Papautsky
*ab
Abstract
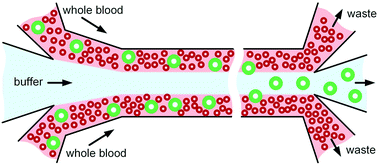
Little work has been done in microfluidics with separation of cells directly from whole blood, and the handful of microfluidic systems reported the literature offer only limited throughput. Yet high throughput is highly desirable to avoid degradation of samples, which can result in loss of information critical to disease diagnosis or monitoring. In this work, we investigated particle migration dynamics in whole blood flow at a single-particle level and subsequently successfully demonstrated the preferential enrichment of white blood cells (WBCs) in unprocessed whole blood flows flanking a buffer flow. Our in-depth investigation reveals a counter-intuitive, size-based migration of cells in whole blood flow and their tendency to accumulate in the regions near flow interfaces, which is employed for inherent enrichment of WBCs. More importantly, we found the strong size-dependent migration in blood flow stemming from the differentiated downstream velocity of particles, which inversely scales with particle size. Our new insights improve understanding of this counterintuitive microfluidics field, offering guidance for new device design to directly handle whole blood and to expand the applications to meet the real-world need for ultra-fast cell separation.
 Please wait while we load your content...
Please wait while we load your content...

