
Calcium ion-assisted lipid tubule formation†
 *a
*a
Abstract
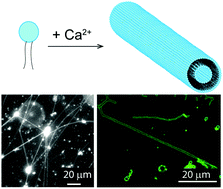
Self-assembled lipid tubules are unique supramolecular structures in cell functions. Lipid tubules that are engineered in vitro are of great interest for technological applications ranging from the templated synthesis of nanomaterials to drug delivery. Herein, we report a study to create long lipid tubules from a mono-unsaturated lipid, 1-stearoyl-2-oleoyl-sn-glycero-3-phosphocholine (SOPC), due to the effect of calcium ions. We found that calcium ions at mM concentrations promote the self-assembly of SOPC lipids into inter-connected hollow lipid tubes that are μm thick and as long as a few millimeters. Higher calcium concentration leads to an increase in the numbers of lipid tubules formed, but has little effect on tubule diameter. Calcium ions also stabilize lipid tubules, which break up upon the removal of ions. We showed that the lipid tubule-promoting effect is general for divalent ions. We were able to vary the morphology of lipid tubules from thin tube to “strings of pearls” structures or increase the tubule thickness by mixing SOPC with other lipids of different spontaneous curvature effects. Our results reveal that the divalent charges of calcium ions and the asymmetric mono-unsaturated structure of SOPC acyl chains act in combination to cause the formation of lipid tubules.
- This article is part of the themed collection: 2017 Emerging Investigators by MCF
 Please wait while we load your content...
Please wait while we load your content...

