
Gastrointestinal interactions, absorption, splanchnic metabolism and pharmacokinetics of orally ingested phenolic compounds

Abstract
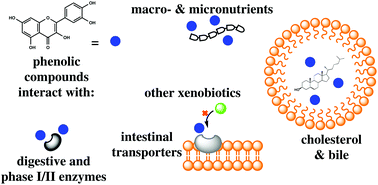
The positive health effects of phenolic compounds (PCs) have been extensively reported in the literature. An understanding of their bioaccessibility and bioavailability is essential for the elucidation of their health benefits. Before reaching circulation and exerting bioactions in target tissues, numerous interactions take place before and during digestion with either the plant or host's macromolecules that directly impact the organism and modulate their own bioaccessibility and bioavailability. The present work is focused on the gastrointestinal (GI) interactions that are relevant to the absorption and metabolism of PCs and how these interactions impact their pharmacokinetic profiles. Non-digestible cell wall components (fiber) interact intimately with PCs and delay their absorption in the small intestine, instead carrying them to the large intestine. PCs not bound to fiber interact with digestible nutrients in the bolus where they interfere with the digestion and absorption of proteins, carbohydrates, lipids, cholesterol, bile salts and micronutrients through the inhibition of digestive enzymes and enterocyte transporters and the disruption of micelle formation. PCs internalized by enterocytes may reach circulation (through transcellular or paracellular transport), be effluxed back into the lumen (P-glycoprotein, P-gp) or be metabolized by phase I and phase II enzymes. Some PCs can inhibit P-gp or phase I/II enzymes, which can potentially lead to drug–nutrient interactions. The absorption and pharmacokinetic parameters are modified by all of the interactions within the digestive tract and by the presence of other PCs. Undesirable interactions have promoted the development of nanotechnological approaches to promote the bioaccessibility, bioavailability, and bioefficacy of PCs.
- This article is part of the themed collection: Food & Function 2017 Most Downloaded Articles
 Please wait while we load your content...
Please wait while we load your content...

