
In situ formation of peptidic nanofibers can fundamentally optimize the quality of immune responses against HIV vaccine†
Abstract
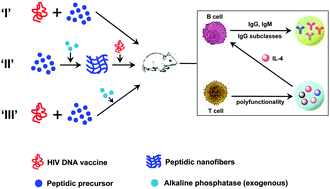
Herein, we report that the in situ formed peptidic nanofibers facilitate the induction of multiple crucial immunities against HIV DNA vaccine, including polyfunctional T cell response, broad IgG subclasses response, and V1/V2 loop-specific antibody response, all of which can hardly be triggered by HIV DNA vaccine alone. Such novel in situ formation fundamentally overcomes the big hurdle for the applications of such nanofibers, which previously can only trigger these crucial immune responses via adding exogenous alkaline phosphatase. Such robustness of peptidic nanofibers for inducing crucial immune responses may allow better inhibition against HIV than reported materials.
- This article is part of the themed collections: Nanoscale Horizons 10th anniversary cover articles, Nanoscale Horizons 10th anniversary regional spotlight collection: China and Top articles from NCNST
 Please wait while we load your content...
Please wait while we load your content...

