
The mechanism of lowering cholesterol absorption by calcium studied by using an in vitro digestion model†
Abstract
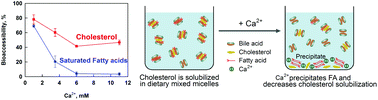
Studies in humans show that a calcium-enriched diet leads to lower cholesterol in blood serum. This phenomenon is usually explained in the literature with a reduced cholesterol absorption in the small intestine. Our study aims to clarify the effect of calcium on the solubilisation of cholesterol and fatty acid in the dietary mixed micelles (DMM), viz. on the bioaccessibility of these lipophilic substances in the gut. We use an in vitro digestion model which mimics very closely the intestinal pH-profile and the composition of the intestinal fluids. We quantified the effects of Ca2+ concentration on the lipid solubilization for fats and oils with different saturated/unsaturated fatty acid (FA) contents. We found that the increase of calcium significantly decreases the solubilization of cholesterol, FA and MG. Most importantly, we observe a clear positive correlation between the amounts of solubilized cholesterol, on one side, and solubilized free fatty acids and monoglycerides, on the other side. The main conclusion is that Ca2+ ions strongly affect the bioaccessibility of both cholesterol and saturated FA. Therefore, calcium may decrease the serum cholesterol via two complementary mechanisms: (1) fatty acid precipitation by calcium ions reduces the solubilisation capacity of the DMM, thus decreasing the levels of solubilised (bioaccessible) cholesterol; (2) the observed strong decrease of the bioaccessible saturated FA, in its own turn, may suppress the cholesterol synthesis in the liver.
 Please wait while we load your content...
Please wait while we load your content...

