
Fabrication of phospho-phytase/heteroatomic hierarchical Fe-ZSM-5 zeolite (HHFeZ) bio-conjugates for eco-sustainable utilization of phytate-phosphorus†
Abstract
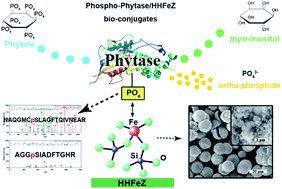
The originally fabricated phytase/HHFeZ bio-conjugates could appropriately enhance enzyme stabilization after heteroatomic hierarchical Fe-ZSM-5 zeolite (HHFeZ) was developed as a novel matrix for affinitive immobilization of Escherichia coli (E. coli) phytase. The immobilization strategy was primarily dependent on the affinitive coordination between heteroatomic iron atoms anchored into the HHFeZ frameworks with multiple sites of phosphorylation in phytase. In addition, the ordered multiporous structures of HHFeZ contributed to phytase immobilization and stabilization. The conditions for phytase immobilization were optimized under a solution pH of 4.5 in a batch mode, and the maximum phytase immobilization capacity was determined to be 2.01 mg mg−1. Compared to those of free phytase, the thermal and proteolytic resistance of phytase/HHFeZ bio-conjugates were effectively improved, and the enzymatic activity was maintained over a broad pH range. The phytase/HHFeZ bio-conjugates would be an innovative choice for eco-sustainable exploitation and utilization of phytate-phosphorus, especially with respect to eutrophication control.
 Please wait while we load your content...
Please wait while we load your content...

