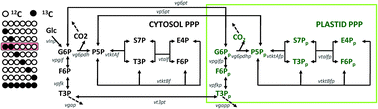
Metabolic fluxes are powerful indicators of cell physiology and can be estimated by isotope-assisted metabolic flux analysis (MFA). The complexity of the compartmented metabolic networks of plants has constrained the application of isotope-assisted MFA to them, principally because of poor identifiability of fluxes from the measured isotope labeling patterns. However, flux identifiability can be significantly improved by a priori design of isotope labeling experiments (ILEs). This computational design involves evaluating the effect of different isotope label and isotopomer measurement combinations on flux identifiability, and thereby identifying optimal labels and measurements toward evaluating the fluxes of interest with the highest confidence. This article reports ILE designs for two major, compartmented plant metabolic pathways – the pentose phosphate pathway (PPP) and γ-aminobutyric acid (GABA) shunt. Together, these pathways represent common motifs in plant metabolism including duplication of pathways in different subcellular compartments, reversible reactions and cyclic carbon flow. To compare various ILE designs, we employed statistical A- and D-optimality criteria. Our computations showed that 1,2-13C Glc is a powerful and robust label for the plant PPPs, given currently popular isotopomer measurement techniques (single quadrupole mass spectrometry [MS] and 2-D nuclear magnetic resonance [NMR]). Further analysis revealed that this label can estimate several PPP fluxes better than the popular label 1-13C Glc. Furthermore, the concurrent measurement of the isotopomers of hexose and pentose moieties synthesized exclusively in the cytosol or the plastid compartments (measurable through intracellular glucose or sucrose, starch, RNA ribose and histidine) considerably improves the identifiability of PPP fluxes in the individual compartments. Additionally, MS-derived isotopomer measurements outperform NMR-derived measurements in identifying PPP fluxes. The potency of 1,2-13C Glc can be improved substantially by combining it with other labels (e.g. 3-13C Glc, 1-13C Glc and U-13C Glc) in parallel ILEs. For the GABA shunt, we calculated that 100% 2-13C Ala and 100% U-13C Gln constitute the best labels. We anticipate that the ILE designs presented in this article can enhance the quality of flux estimates in these two complex plant pathways. In the future, these ILE designs can be further improved by leveraging recent analytical and computational developments in isotope-assisted MFA.

 Please wait while we load your content...
Please wait while we load your content...