
Effects of temperature and soil moisture on methyl halide and chloroform fluxes from drained peatland pasture soils
Abstract
A series of laboratory-based incubations using a stable isotope
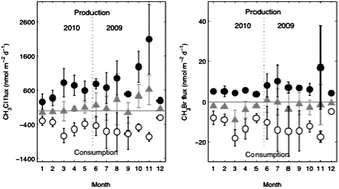
A series of laboratory-based incubations using a stable isotope
M. A. H. Khan, M. E. Whelan and R. C. Rhew, J. Environ. Monit., 2012, 14, 241 DOI: 10.1039/C1EM10639B
To request permission to reproduce material from this article, please go to the Copyright Clearance Center request page.
If you are an author contributing to an RSC publication, you do not need to request permission provided correct acknowledgement is given.
If you are the author of this article, you do not need to request permission to reproduce figures and diagrams provided correct acknowledgement is given. If you want to reproduce the whole article in a third-party publication (excluding your thesis/dissertation for which permission is not required) please go to the Copyright Clearance Center request page.
Read more about how to correctly acknowledge RSC content.
 Please wait while we load your content...
Please wait while we load your content...