
Early gene regulation of osteogenesis in embryonic stem cells†
Abstract
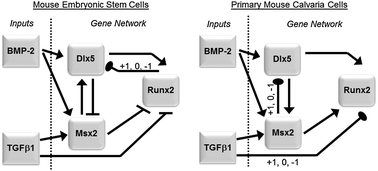
The early gene regulatory networks (GRNs) that mediate stem cell differentiation are complex, and the underlying regulatory associations can be difficult to map accurately. In this study, the expression profiles of the genes Dlx5, Msx2 and Runx2 in mouse embryonic stem cells were monitored over a 48 hour period after exposure to the growth factors BMP2 and TGFβ1. Candidate GRNs of early osteogenesis were constructed based on published experimental findings and simulation results of Boolean and ordinary differential equation models were compared with our
 Please wait while we load your content...
Please wait while we load your content...