a
CNRS, LCC (Laboratoire de Chimie de Coordination), 205, route de Narbonne, F-31077 Toulouse, France
E-mail:peter.faller@lcc-toulouse.fr Fax: 0033/561553003
b
Université de Toulouse, UPS, INPT, LCC, F-31077 Toulouse, France
c
Department of Biochemistry, University of Zürich, Winterthurerstrasse 190, CH-8057 Zürich, Switzerland
Abstract
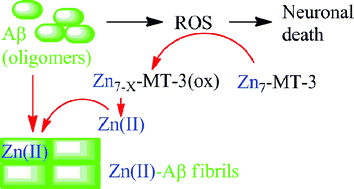
The reactive oxygen species H2O2 promotes the Zn7-metallothionein-3 induced Aβ40 aggregation of fibrillar type structures via slow cysteineoxidation and Zn2+ release, whereas amorphous aggregates are formed by addition of Zn2+ to Aβ40.
You have access to this article
Please wait while we load your content...
Something went wrong. Try again?

 Please wait while we load your content...
Please wait while we load your content...