
Inhibition of SARS coronavirus helicase by bismuth complexes†
Abstract
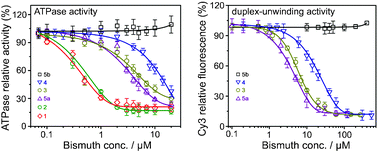
A series of bismuth complexes were synthesized and characterized, and most of them exhibited inhibition against the SARS coronavirus helicase ATPase and duplex-unwinding activities at micromolar concentrations.
- This article is part of the themed collection: Coronavirus articles - free to access collection
 Please wait while we load your content...
Please wait while we load your content...

