
Controlled assembly of heterotypic cells in a core–shell scaffold: organ in a droplet†
Dong
Chen,  b
b
b
Abstract
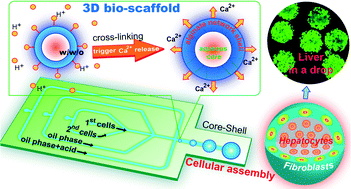
This paper reports a droplet-based microfluidic approach to fabricate a large number of monodisperse, portable microtissues, each in an individual drop. We use water–water–oil double emulsions as templates and spatially assemble hepatocytes in the core and fibroblasts in the shell, forming a 3D liver model in a drop.
 Please wait while we load your content...
Please wait while we load your content...

