
Nanosilicon-based recovery of barley (Hordeum vulgare) plants subjected to drought stress†

Abstract
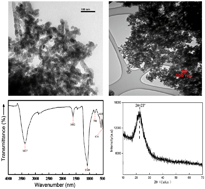
The present study explores the potential impact of silicon nanoparticles (Si NPs), in comparison with their bulk counterpart (silicate), on post-stress recovery performance of barley (Hordeum vulgare) seedlings under different drought stress intensities during vegetative growth. Barley plants were grown under 100% field capacity (FC), or mild (75% FC), moderate (50% FC) and severe (25% FC) drought stress levels, and were subsequently recovered by different treatments including soil application of 150 mL of Si NPs and silicate (at 125 and 250 mg Si L−1), and water. Si NPs application at 250 mg L−1 led to formation of Si NP aggregates in plant tissues, large pores in roots, and also rapid stomata closure in leaves. However, the lower Si NPs dose (125 mg Si L−1) was accompanied by a wider distribution of Si NPs in cells, and formation of a regular porosity pattern in roots i.e. more frequent pores of a smaller size. Upon recovery from all the drought stress levels, shoot biomass increased significantly in recovered plants compared to the respective non-recovered controls, and the maximum shoot biomass increase (27.3%) belonged to the moderate-stressed plants treated with Si NPs at 125 mg L−1. Exposure to Si NPs and silicate (at both doses) after all drought stress intensities caused a significant increase in total chlorophyll (up to 17.1%) and carotenoid (up to 24.1%) content of leaves except for the carotenoid content under severe drought stress. Post-drought recovery with Si NPs and silicate was linked to alterations in the plant osmolyte and metabolite profile, cellular injury and membrane stability indices, and the activity of antioxidant enzymes. Soil application of Si NPs (at a low dose of 125 mg Si L−1) showed a promising potential for post-drought recovery of barley plants via modifying plant morpho-physiological and antioxidative attributes and synthesis of specific metabolites.
 Please wait while we load your content...
Please wait while we load your content...

