
Targeting Acr3 from Ensifer medicae to the plasma membrane or to the tonoplast of tobacco hairy roots allows arsenic extrusion or improved accumulation. Effect of acr3 expression on the root transcriptome†

Abstract
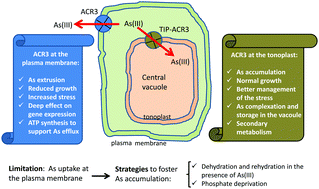
Transgenic tobacco hairy roots expressing the bacterial arsenite efflux pump Acr3 from Ensifer medicae were generated. The gene product was targeted either to the plasma membrane (ACR3 lines) or to the tonoplast by fusing the ACR3 protein to the tonoplast integral protein TIP1.1 (TIP-ACR3 lines). Roots expressing Acr3 at the tonoplast showed greater biomass than those expressing Acr3 at the plasma membrane. Furthermore, higher contents of malondialdehyde (MDA) and RNA degradation in ACR3 lines were indicative of higher oxidative stress. The determination of ROS-scavenging enzymes depicted the transient role of peroxidases in ROS detoxification, followed by the action of superoxide dismutase during both short- and medium-term exposure periods. Regarding As accumulation, ACR3 lines accumulated up to 20–30% less As, whereas TIP-ACR3 achieved a 2-fold increase in As accumulation in comparison to control hairy roots. Strategies that presumably induce As uptake, such as phosphate deprivation or dehydration followed by rehydration in the presence of As, fostered As accumulation up to 10 800 μg g−1. Finally, the effects of the heterologous expression of acr3 on the root transcriptome were assessed. Expression at the plasma membrane induced drastic changes in gene expression, with outstanding overexpression of genes related to electron transport, ATP synthesis and ATPases, suggesting that As efflux is the main detoxification mechanism in these lines. In addition, genes encoding heat shock proteins and those related to proline synthesis and drought tolerance were activated. On the other hand, TIP-ACR3 lines showed a similar gene expression profile to that of control roots, with overexpression of the glutathione and phytochelatin synthesis pathways, together with secondary metabolism pathways as the most important resistance mechanisms in TIP-ACR3, for which As allocation into the vacuole allowed better growth and stress management. Our results suggest that modulation of As accumulation can be achieved by subcellular targeting of Acr3: expression at the tonoplast enhances As accumulation in roots, whereas expression at the plasma membrane could promote As efflux. Thus, both approaches open the possibilities for developing safer crops when grown on As-polluted paddy soils, but expression at the tonoplast leads to better growth and less stressed roots, since the high energy cost of As efflux likely compromises growth in ACR3 lines.
 Please wait while we load your content...
Please wait while we load your content...