
Shiga toxin binding alters lipid packing and the domain structure of Gb3-containing membranes: a solid-state NMR study†

Abstract
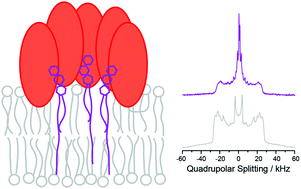
We studied the influence of globotriaosylceramide (Gb3) lipid molecules on the properties of phospholipid membranes composed of a liquid ordered (lo)/liquid disordered (ld) phase separated 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC)/N-palmitoyl-D-erythro-sphingosylphosphorylcholine (PSM)/cholesterol mixture (40/35/20, mol/mol/mol) supplemented with 5 mol% of either short acyl chain palmitoyl-Gb3 or long acyl chain lignoceryl-Gb3 using 2H solid-state NMR spectroscopy. To this end, both globotriaosylceramides were chemically synthesized featuring a perdeuterated lipid acyl chain. The solid-state 2H NMR spectra support the phase separation into a POPC-rich ld phase and a PSM/cholesterol-rich lo phase. The long chain lignoceryl-Gb3 showed a rather unusual order parameter profile of the acyl chain, which flattens out for the last ∼6 methylene segments. Such an odd chain conformation can be explained by partial chain interdigitation and/or a very fluid midplane region of the membrane. Possibly, the Gb3 molecules may thus preferentially be localized at the lo/ld phase boundary. In contrast, the short chain palmitoyl-Gb3 was well associated with the PSM/cholesterol-rich lo phase. Gb3 molecules act as membrane receptors for the Shiga toxin (STx) produced by Shigella dysenteriae and by enterohemorrhagic strains of Escherichia coli (EHEC). The B-subunits of STx (STxB) forming a pentameric structure were produced recombinantly and incubated with the membrane mixtures leading to alterations in the lipid packing properties and lateral organization of the membranes. Typically, STxB binding led to a decrease in lipid chain order in agreement with partial immersion of protein segments into the lipid–water interface of the membrane. In the presence of STxB, Gb3 preferentially partitioned into the lo membrane phase. In particular the short acyl chain palmitoyl-Gb3 showed very similar chain order parameters to PSM. In the presence of STxB, all lipid species showed isotropic contributions to the 2H NMR powder spectra; this was most pronounced for the Gb3 molecules. Such isotropic contributions are caused by highly curved membrane structures, which have previously been detected as membrane invaginations in fluorescence microscopy. Our analysis estimated that STxB induced highly curved membrane structures with a curvature radius of less than ∼10 nm likely related to the insertion of STxB segments into the lipid–water interface of the membrane.
 Please wait while we load your content...
Please wait while we load your content...

