
LC-MS/MS suggests that hole hopping in cytochrome c peroxidase protects its heme from oxidative modification by excess H2O2†

Abstract
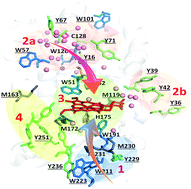
We recently reported that cytochrome c peroxidase (Ccp1) functions as a H2O2 sensor protein when H2O2 levels rise in respiring yeast. The availability of its reducing substrate, ferrocytochrome c (CycII), determines whether Ccp1 acts as a H2O2 sensor or peroxidase. For H2O2 to serve as a signal it must modify its receptor so we employed high-performance LC-MS/MS to investigate in detail the oxidation of Ccp1 by 1, 5 and 10 M eq. of H2O2 in the absence of CycII to prevent peroxidase activity. We observe strictly heme-mediated oxidation, implicating sequential cycles of binding and reduction of H2O2 at Ccp1's heme. This results in the incorporation of ∼20 oxygen atoms predominantly at methionine and tryptophan residues. Extensive intramolecular dityrosine crosslinking involving neighboring residues was uncovered by LC-MS/MS sequencing of the crosslinked peptides. The proximal heme ligand, H175, is converted to oxo-histidine, which labilizes the heme but irreversible heme oxidation is avoided by hole hopping to the polypeptide until oxidation of the catalytic distal H52 in Ccp1 treated with 10 M eq. of H2O2 shuts down heterolytic cleavage of H2O2 at the heme. Mapping of the 24 oxidized residues in Ccp1 reveals that hole hopping from the heme is directed to three polypeptide zones rich in redox-active residues. This unprecedented analysis unveils the remarkable capacity of a polypeptide to direct hole hopping away from its active site, consistent with heme labilization being a key outcome of Ccp1-mediated H2O2 signaling. LC-MS/MS identification of the oxidized residues also exposes the bias of electron paramagnetic resonance (EPR) detection toward transient radicals with low O2 reactivity.
 Please wait while we load your content...
Please wait while we load your content...

