
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceBiological production and downstream separation of xylitol using Candida tropicalis
Sarvada Chipkar ,
Claire E. Richter,
Charlotte A. Tonelli,
Luke W. Baxter,
Melanie H. Cotta,
Isabella C. Doyle,
William P. Rice,
Gillian I. Mohr,
Avery W. Suza and
Emma C. Brace*
,
Claire E. Richter,
Charlotte A. Tonelli,
Luke W. Baxter,
Melanie H. Cotta,
Isabella C. Doyle,
William P. Rice,
Gillian I. Mohr,
Avery W. Suza and
Emma C. Brace*
Department of Engineering, Morrissey College of Arts and Sciences, Boston College, 140 Commonwealth Avenue, Chestnut Hill, MA 02467, USA. E-mail: emma.brace@bc.edu; Tel: +1-617-552-6153
First published on 26th May 2026
Abstract
Xylitol is one of the platform chemicals identified by the U.S. Department of Energy as a promising candidate for production from biomass. It is an important chemical building block with applications in both the food and pharmaceutical industries. Xylitol is a pentose sugar alcohol with low caloric value, anti-cariogenic properties, and some ability to inhibit microbial growth. The objective of this work is to identify new routes for production of xylitol from lignocellulosic biomass with a focus on post-fermentation downstream processing and separation. Fermentation (via C. tropicalis) conditions including varied initial xylose concentrations (10 and 30 g L−1) and different pH control methodologies (with sodium hydroxide and PBS buffer or without adjustment) were evaluated. An initial xylose concentration of 10 g L−1 with pH control using sodium hydroxide resulted in the highest xylitol yield of 75%. Amberlite IRC 748, a commonly used cationic resin for wastewater treatment, was used to develop an ion-exchange chromatographic separation for the xylitol from fermentation hydrolysates, with the intention to then concentrate and crystallize the xylitol. It was determined that three consecutive water washes were able to selectively recover xylitol from the ion-exchange column with 45% of the recovered xylitol concentrated in the second water wash at even low retention times of 1 minute. Future work may include comprehensive techno-economic analyses, comparisons of different feedstocks, and a life cycle assessment of the xylitol biorefinery.
1. Introduction
In 2004, the U.S. Department of Energy (DOE) identified 12 platform molecules as high-priority targets for development from lignocellulosic biomass.1 Platform molecules are valuable chemical building blocks with a variety of applications, including conversion to commodity chemicals and high-value products. Xylitol is one such platform molecule of interest in food, pharmaceuticals, and consumer products industries.2,3 Xylitol, a pentose sugar alcohol, is a low calorie, anti-cariogenic, and anti-microbial compound.4Conventional xylitol production methods use feedstocks such as birchwood chips and sugarcane bagasse,5 and involve chemical or thermochemical processes that require high pressures and temperatures, resulting in increased manufacturing costs and increased energy consumption.6,7 There are four fundamental stages involved in conventional xylitol manufacturing processes, which produce final yields up to 99.9%: (1) converting xylan to xylose by acid hydrolysis; (2) purifying the hydrolysate to remove hydrolysis byproducts; (3) catalytic hydrogenation of xylose to obtain xylitol solution using metal catalysts; (4) purification of xylitol broth followed by crystallization.2 The exorbitant operating costs for these processes are due to high energy consumption in the processing and use of raw materials such as purified xylan and metal-catalysts, which incur adverse environmental impacts and contribute to higher xylitol market prices.5
Agricultural waste, such as corn stover residue, are a largely untapped source of xylan-rich hemicellulose, representing the opportunity for a sustainably fed, comparatively energy efficient xylitol production process.8 Hemicellulose is a major component of lignocellulosic biomass and is high in xylan, which can be used to create a hydrolysate for the biological production of xylitol.9 However, industrial-scale, cost-effective xylitol biorefineries have not yet been realized. Biological production of xylitol from lignocellulosic biomass waste is promising because it lacks both the high costs of raw materials and the energy intensive operating factors associated with the conventional methods of xylitol production.7,10 Fermentation conducted at near-ambient temperatures and pressures could mitigate the adverse environmental impacts of xylitol production by reducing energy consumption and by utilizing inexpensive, relatively abundant biomass waste products, such as corn cobs, corn stover, or sugarcane bagasse. Many researchers have explored xylitol production through various combinations of pH and temperature conditions for C. tropicalis fermentation with varying amounts of initial xylose loading. Various research studies reported xylitol yield between 0.45–0.87 g g−1 xylose at pH ranges of 4–5.5 at 35 °C at an industrial scale of 60–120 g L−1 initial xylose loading with wild type or genetically modified C. tropicalis.5,11–15 Mohamad et al. reported 0.82 g g−1 xylitol yield with a starting pH and xylose loading of 4 and 23 g L−1 respectively.14 Xu et al. found that 100 g L−1 of initial xylose loading gave maximum xylitol yield of 65 g L−1 (0.65 g g−1) at an initial media pH 4.5.15 Tamburini et al. used a genetically modified C. tropicalis strain which reached a xylitol yield of 87% (w/w) with initial pH and xylose loading of 2.5 and 80 g L−1 respectively.12 However, Tamburini et al. demonstrated that maximum specific yeast growth is more viable at higher pH when fermentation was tested at different pH levels between 2.5 to 6.15.12 Near-neutral pH (∼7) than acidic operating pH, and higher specific cell growth that contributes to recycling the yeast cells are crucial operational aspects for an economically profitable biorefinery.
A major driver of costs for biological production pathways is the downstream separations and purification. There are two main challenges: to remove compounds generated during the pretreatment process and to selectively separate xylitol from competitive carbohydrate molecules present in the fermentation hydrolysate. Researchers have employed activated charcoal or aluminum-salts as a clarification step to remove colored compounds or pretreatment inhibitors.16,17 Others have used membrane filtration including nano- and micro-porous membranes to remove salts formed either as fermentation byproducts or as precipitates from pH neutralization.18,19 Subsequent separation and purification techniques, including ion-exchange resins, liquid–liquid extraction, or membrane nanofiltration, are then employed to achieve high purity xylitol crystals.19–21
The objectives of this work are to: (1) investigate the use of Candida tropicalis to produce xylitol from lignocellulosic biomass-derived xylose; (2) explore the impact of fermentation parameters such as pH on xylitol titer and yield; (3) develop a cationic exchange process to separate and recover xylitol from the fermentation hydrolysate. In this work, C. tropicalis was used in shake-flask fermentation experiments to produce xylitol under varied xylose feed concentrations and pH control methods. Additionally, an ion-exchange chromatography system with a cationic resin bed was developed as a novel downstream processing step to purify xylitol from the fermentation hydrolysate. The impacts on titers and yields of xylitol were quantitatively measured.
2. Materials and methods
2.1. C. tropicalis culture and fermentation
An initial culture of C. tropicalis ATCC 750 was obtained (Kwik-Stik, Microbiologics, Inc.) and cultured using a yeast extract-malt extract-peptone (YMP) media. The C. tropicalis strain was chosen based on a review of yeasts with a metabolic pathway for converting xylose to xylitol.22,23 The C. tropicalis YMP growth media was prepared with 5 g L−1 yeast extract, 5 g L−1 malt extract, and 10 g L−1 peptone. Fermentation was performed in 250 mL shake-flasks with a working volume of 200 mL in an incubator-shaker (Innova 42R, Eppendorf) at 30 °C and 200 rpm. Xylose was added to growth media to achieve concentrations of either 10 g L−1 or 30 g L−1. 199 mL of this was added to an Erlenmeyer flask, followed by inoculation with 1 mL of C. tropicalis seed culture. Each fermentation condition (initial xylose concentration of 10 g L−1 or 30 g L−1; with or without pH adjustments as described in Section 2.2) was completed in triplicate over 96 hours. Aliquots (1 mL) were removed at regular time intervals (hourly from 0–16 hours; every 8 hours from 16–96 hours) to characterize the C. tropicalis growth rate. These aliquots were refrigerated and later used for quantifying xylose and xylitol concentration using high performance liquid chromatography (HPLC). The basis for the focus on 10 and 30 g L−1 xylose feed concentrations comes from literature and knowledge of reasonable sugar feed concentrations for various strains of C. tropicalis. These concentrations were also chosen to be consistent with and/or reasonable for comparison with sugar concentrations in liquefied biomass, which would be the ultimate source if this work were expanded on and/or scaled-up to use actual lignocellulosic biomass as the feedstock. The authors did initially test a 50 g L−1 feed concentration, but crude analysis indicated it led to poor results (growth and xylitol production), and thus was excluded from any further study at this time. Ion-exchange time optimization fermentation experiments were performed with six biological replicates for 10 g L−1 initial xylose loading with and without pH control to account for a robust biological variability.2.2. pH control methods
A benchtop meter (Thermo Scientific Orion Star A211) was used to measure pH. The effect of pH on fermentation was investigated using three pH control methods: (1) no pH adjustment or control; (2) addition of 3 M sodium hydroxide (NaOH) throughout fermentation to maintain a pH above 6.5; (3) addition of phosphate-buffered saline (PBS) solution (10× PBS solution, pH 6.5 at 1×, Teknova) at a ratio of 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :9 (v/v) with the YMP growth media before beginning fermentation. These choices were made based on preliminary experiments which indicated that without any controls or interventions, the pH would start between 6.5 and 7.0, drop, and then increase during fermentation; this is reflected in the results for set (1) with no pH control. Literature would suggest that the acidic environment in the no pH control group can be attributed to the C. tropicalis metabolic pathway producing acidic products such as acetic acid and glycolic acid, while the xylitol producing pathway may require a more basic pH.24,25 As pH was measured at regular intervals as described in Section 2.1, whenever the pH of an individual flask for pH set (2) dropped below 6.4, 5–30 µL of 3 M sodium hydroxide were added to increase pH above 6.5, thus attempting to maintain a pH near 6.5 throughout all 96 hours of fermentation. For sets (1) and (3), pH was only measured and recorded throughout fermentation.
:9 (v/v) with the YMP growth media before beginning fermentation. These choices were made based on preliminary experiments which indicated that without any controls or interventions, the pH would start between 6.5 and 7.0, drop, and then increase during fermentation; this is reflected in the results for set (1) with no pH control. Literature would suggest that the acidic environment in the no pH control group can be attributed to the C. tropicalis metabolic pathway producing acidic products such as acetic acid and glycolic acid, while the xylitol producing pathway may require a more basic pH.24,25 As pH was measured at regular intervals as described in Section 2.1, whenever the pH of an individual flask for pH set (2) dropped below 6.4, 5–30 µL of 3 M sodium hydroxide were added to increase pH above 6.5, thus attempting to maintain a pH near 6.5 throughout all 96 hours of fermentation. For sets (1) and (3), pH was only measured and recorded throughout fermentation.
2.3. Cell growth measurements
C. tropicalis growth curves were developed by taking optical density measurements at a wavelength of 600 nm (OD600) on a UV-VIS spectrophotometer (Genesys 30, Thermo Scientific) at each sampling timepoint. A sample taken immediately pre-incubation was used as the blank. Samples were diluted as necessary using a 10× dilution factor in order to read an absorbance value lower than 1.00 on the spectrophotometer. The absorbance reading of each sample was recorded and multiplied by the corresponding dilution factor, if used. Because each fermentation condition was conducted using three biological replicates for initial xylose loading optimization fermentation and six biological replicates for ion-exchange time optimization fermentation experiments, the average values were calculated and used to construct a logarithmic growth curve for the C. tropicalis.2.4. Xylose and xylitol concentrations
2.5. Ion-exchange chromatography
The fermentation hydrolysate was passed through commercially available cationic ion-exchange resin Amberlite IRC-748 (Thermo Scientific) to obtain a purified xylitol broth solution. A resin bed of 31 cm in height was hand-packed inside a glass column with 2 cm column diameter with between 3 and 5 cm of glass wool at the bottom of the column. An additional 2 cm of glass wool was added to the top surface of the resin bed. The column was conditioned using deionized water. The column was also disinfected after use with 30% (by volume) ethanol solution, followed by subsequent washes with deionized water to remove any ethanol residue. Hydrolysate from the six replicates of the second set of fermentation (10 g L−1 initial xylose loading with and without pH control) was added to the column, followed by an equal volume (30 mL) of water washes (WW). To characterize the xylitol elution from the column, eluents for the initial sample eluent (SE) and each of the water washes were collected subsequently in two separate tubes in ∼14 mL increments and were tagged accordingly (Eg, 1–1, 1–2, 2–1, 2–2; the first number represents the water wash cycle while the second number represents the two collection tubes in chronological order). Since the six hydrolysate replicates had a similar trend, three fermentation hydrolysates from the second set of fermentation were chosen as technical replicates for the time retention optimization experiments. The eluents were collected in single collection tubes for each of the water washes and the sample eluent. The unaccounted xylitol was calculated for each samples by subtracting the xylitol present in the three water washes from the total xylitol present in the fermentation hydrolysate that was added to the column. Error propagation math was used to calculate standard deviations for the averaged data points.3. Results and discussion
3.1. pH monitoring
The 10 g L−1 and the 30 g L−1 initial xylose feed groups followed similar pH trends between 0–16 hours across each of the three pH control methods: (1) no pH adjustment or control; (2) addition of 3 M sodium hydroxide throughout fermentation to maintain a pH above 6.5; (3) addition of phosphate-buffered saline (PBS) solution with a pH of 6.5 (Teknova) to the growth media before beginning fermentation (as described in Section 2.2). The pH consistently decreased between 0 – 16 hours across all samples, independent of xylose concentrations and pH control type, as shown in Fig. 1. | ||
| Fig. 1 pH measurements as fermentation progresses. Initial xylose concentrations were 10g L−1 (purple) and 30 g L−1 (green). pH was measured for three biological replicates at various timepoints from 0–96 hours as described in the methods, with the shading representing standard deviations. | ||
3.2. Growth of C. tropicalis
The experimentally determined cell growth follows a typical logarithmic growth pattern, with a lag phase, exponential growth phase, and stationary phase (Fig. 2). The three sets of pH conditions displayed similar growth curves, with only slight differences in the initial lag phase.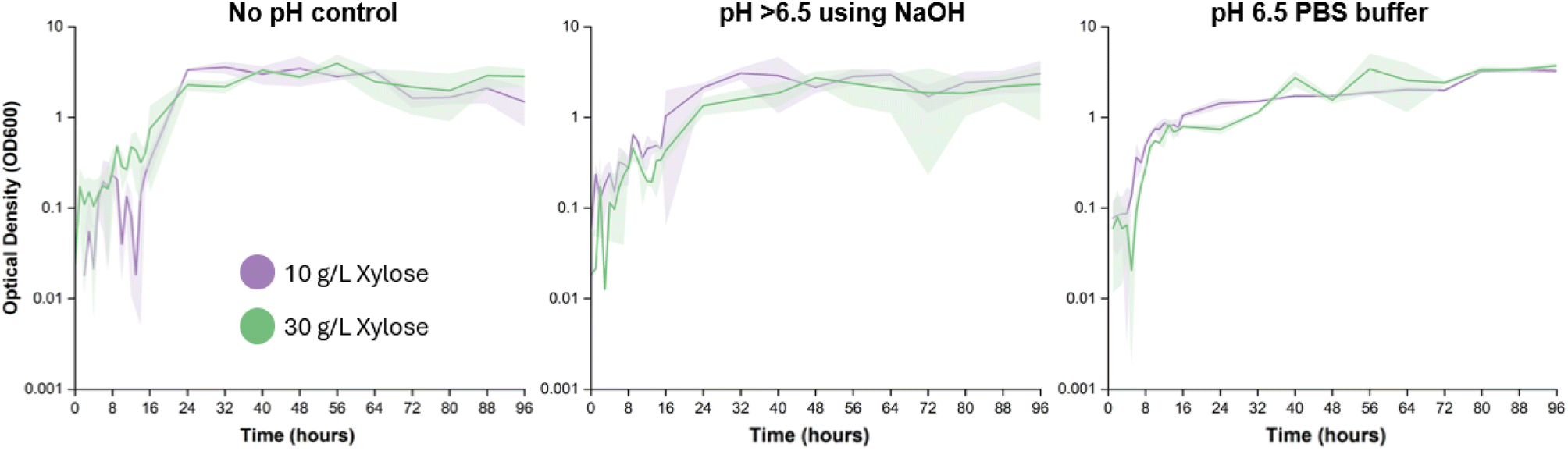 | ||
| Fig. 2 The optical density measurements at 600 nm (OD600) as fermentation progresses over 96 hours show a typical logarithmic growth pattern of the C. tropicalis, with minimal impact on the growth phase from using different pH control methods. Each point is an average of three biological replicates with the shaded region representing the standard deviation. | ||
The primary difference is in the time it takes to reach the stationary phase; this occurred at approximately 24 hours for the flasks with no pH control, as opposed to between 16 and 24 hours for flasks with some sort of pH adjustment (NaOH added periodically, or PBS buffer added initially, as described in Section 2.2). The no pH control group offered relatively higher acidic environment to the yeast during the first 24 hours as opposed to the pH adjusted environment (Fig. 1) that led to faster acclimatization of the yeast and hence relatively shorter lag phase. The acidic environment in the no pH control group can be attributed to the C. tropicalis metabolic pathway producing acidic products such as acetic acid and glycolic acid.24,25 Xylose is metabolized by C. tropicalis to form xylitol using two different pathways: (1) reduction of xylose to d-xylitol through xylose reductase and NADPH as co-enzymes, (2) oxidation of xylose to D-xylulose through xylose dehydrogenase and NAD as co-enzymes.29 However, xylitol is further metabolized by C. tropicalis by the second pathway of xylulose kinase to form xylulose-5-phosphate,30 which is then converted into pyruvate via the phosphate pentose pathway.31 The phosphate pentose pathway eventually leads to generation of lactate and acetic acid thus reducing the pH of the fermentation hydrolysate. The type of enzymes dominating in the procreating yeast determines the pH of the fermentation hydrolysate.
3.3. Analysis of xylitol titer, yield, and xylose depletion
Initially, xylitol assays were performed (Section 2.4) to measure the concentration of xylitol in fermentation samples taken at various timepoints. The final xylitol titer and yield for the two initial xylose concentrations and three pH control methods are shown in Table 1.| Initial xylose concentration (g L−1) | pH control method | Final xylitol titer (g L−1) | Xylitol yield (g xylitol/g xylose) | Percent yield |
|---|---|---|---|---|
| 10 | No pH control | 4.72 ± 1.96 | 0.47 ± 0.16 | 46.60% ± 0.16 |
| Maintained > 6.5 using NaOH | 7.59 ± 4.62 | 0.76 ± 0.38 | 74.90% ± 0.37 | |
| Initial pH of 6.5 using PBS buffer | 3.06 ± 0.47 | 0.31 ± 0.04 | 30.20% ± 0.04 | |
| 30 | No pH control | 12.90 ± 4.70 | 0.43 ± 0.13 | 42.40% ± 0.13 |
| Maintained > 6.5 using NaOH | 17.10 ± 6.60 | 0.57 ± 0.18 | 56.30% ± 0.18 | |
| Initial pH of 6.5 using PBS buffer | 8.79 ± 2.73 | 0.29 ± 0.07 | 28.90% ± 0.07 |
The fermentation condition resulting in the highest xylitol yield was 10 g L−1 initial xylose concentration with pH maintained above 6.5 using NaOH, at 74.9%, as shown in Table 1 and Fig. 3. However, the highest titer came from the 30 g L−1 initial xylose concentration, also with pH maintained above 6.5 using NaOH.
 | ||
| Fig. 3 Xylitol concentrations under different fermentation conditions measured using a D-sorbitol/D-xylitol assay kit (Megazyme/Neogen). Each bar represents the average of biological triplicates, with error bars representing standard deviations. Samples that do not share a letter on top of the bars resemble significant difference based on Tukey's pairwise statistical comparison (α = 0.05) (SI-Table 1A). | ||
Fermentation flasks in which pH was adjusted through addition of NaOH produced the highest xylitol titers for both 10 g L−1 and 30 g L−1 xylose loading which was followed by the samples without pH control (Table 1 and Fig. 3). Samples with PBS buffer-control had no significant trend over the course of fermentation for 10 g L−1 xylose loading while samples experienced an increasing trend for 30 g L−1 xylose loading group. Overall, 30 g L−1 xylose loading group gave higher xylitol titers than 10 g L−1 groups as expected. The xylitol producing yeast favor basic pH levels for efficient xylitol production. However, the 30 g L−1 initial xylose concentrations result in lower yields (per mass basis of initial xylose feedstock). Hence, only two pH conditions – no pH control and pH adjustment with NaOH – with 10 g L−1 xylose loading were chosen for further study. We hypothesize that xylitol may switch from playing the role of product to that of substrate in the C. tropicalis metabolism when all xylose has been depleted, which could contribute to the interesting trends in the 30 g L−1 xylose loaded fermentations.
3.4. Analysis of xylose and xylitol concentrations using HPLC
Ultimately, experiments were repeated using the advantageous 10 g L−1 initial xylose concentrations, with either no pH adjustment or pH adjustments made to maintain pH greater than 6.5 through addition of NaOH. Xylitol concentrations described in the previous section were determined using assay kits due to logistical constraints. High-Performance Liquid Chromatography (HPLC) was performed for these repeated experiments to measure xylose and xylitol concentrations in aliquots collected from fermentation experiments to achieve greater quantitative rigor than the xylitol assay kits; OD600 was measured in the same way as before, using UV-VIS spectrophotometry at 600 nm. The results are shown in Fig. 4. | ||
| Fig. 4 Concentration (g L−1) of xylose and xylitol at fermentation timepoints align with the left y-axis; optical density measurements at 600 nm (OD600) align with the right y-axis. Each point is the average of six biological replicates; the error bars represent standard deviation. | ||
The trends in the results of cell growth (OD600) and xylitol concentration measurements were largely consistent with the previous set of experiments, although xylitol concentrations were significantly lower than what was measured by the assay kits. This could be due to error and overestimation of xylitol in the assay kits, including the presence of other compounds being misconstrued as xylitol. The overall trend, however, remained the same, in which xylitol concentration peaked at 24 hours for both pH control methods used. Use of HPLC allowed for more accurate measurement of xylitol concentration, and for analysis of the rate of xylose depletion. Interestingly, there is no major difference in the cell growth rates or the xylose depletion rates under the different initial xylose concentrations or under the different pH conditions; the only difference seems to be in the final xylitol concentration, which is marginally higher for the samples without any pH control. This trend is consistent with other findings on effect of pH on xylitol production from lignocellulosic xylose by Tamburini et al. and Singh et al.11,12 The final titers and yields are shown in Table 2.
| Initial xylose concentration (g L−1) | pH control method | Final xylitol titer (g L−1) | Xylitol yield (g xylitol/g xylose) | Percent yield (%) |
|---|---|---|---|---|
| 10 | No pH control | 4.55 ± 0.26 | 0.56 ± 0.04 | 55.10 ± 0.04 |
| Maintained > 6.5 using NaOH | 3.84 ± 0.34 | 0.49 ± 0.04 | 47.88 ± 0.04 |
The fermentation conditions without pH adjustment reach higher titers and yields than those with pH adjustment; this is highly advantageous in that a lack of need for pH control simplifies the process and reduces resources required and associated costs. During this repeated set of experiments, pH was also monitored and is shown in Fig. 5.
 | ||
| Fig. 5 pH monitoring for 10 g L−1 initial xylose concentration under two different pH adjustment methods: no adjustment, and pH maintained near 6.5 through addition of NaOH. Each point is an average of three biological replicates, with error bars representing standard deviations. | ||
The results of pH monitoring in these set of experiments were consistent with the previous set of experiments. The pH of samples with NaOH addition was lower than in previous fermentations, which could be due to more conservative addition of NaOH. There are additional reasons as to why no pH control would be advantageous, such as reduced cost (not needing to add any reagents to control or change pH) and reduced environmental impacts from using fewer chemical reagents in the process.
3.5. Ion-exchange separation
The fermentation hydrolysates from the aforementioned experiments (10 g L−1 initial xylose concentration, with no pH control or adjustment using NaOH) were used for ion-exchange chromatography experiments (methods described in Section 2.6). The concentration of xylitol in the fermentation hydrolysate (“initial”), and in each subsequent elution (elution 1 through water wash 3–2) were measured using HPLC. Xylitol was either not detected or was present in minimal amounts in initial eluents when the hydrolysates were added to the column, as expected. The xylitol concentrations in eluents from the ion-exchange column are shown in Fig. 6. Amberlite IRC 748 cationic resin, typically used for brine purification in chlor-alkali process, was repurposed to achieve efficient separation of xylitol. Other researchers have used similar cationic ion-exchange resins and hence we repurposed Amberlite IRC 748 resin to test for xylitol separation.32–34 Due to its chelating nature that usually removes metal contaminants,32–34 this resin has also proven to be effective in selectively separating the xylitol in this fermentation hydrolysate. | ||
| Fig. 6 Xylitol concentration in ion-exchange eluents as measured by HPLC. Each bar represents an average xylitol concentration in the eluents from six replicates of hydrolysates obtained through fermentation with no pH control using C. tropicalis, with error bars representing standard deviations. Samples that do not share a letter on top of the bars resemble significant difference based on Tukey's pairwise statistical comparison (α = 0.05) (SI-Table 2A). | ||
The xylitol presumably holds on to the cationic molecules of the ion-exchange bed during this stage. In subsequent wash cycles with deionized water, xylitol is eluted from the bed with the highest xylitol amount eluted in the second wash cycle for both pH conditions. However, adhesion of the xylitol molecule is stronger, for pH-adjusted samples with NaOH than without pH control samples, especially in the first wash cycle possibly due to the chelating nature of the repurposed ion-exchange resin towards metallic compounds such as Na+. Amberlite IRC 748 resin was chosen for its weak chelating resin characteristic in the Na+ form with the active iminodiacetic acid functional group.35 Xylitol acts as a chelating ligand due to availability of three neighboring hydroxyl groups that are arranged in an alternating plane arrangement, forming strong coordinate complexes with sodium (Hämäläinen and Lönnberg, 1991) when the fermentation hydrolysate is passed through the resin column. The actual distribution coefficient of xylitol depends on the localized temperature, time, bed void, concentration of xylitol around the resin particle and superficial velocity (Watanabe and Adachi, 2023). However, as the water is introduced in the subsequent water washes the available hydrogen molecule poses slightly greater affinity to the xylitol hydroxyl molecule that helps with subsequent separation from other elements in the fermentation hydrolysate (Hämäläinen and Lönnberg, 1991).13–15,29–31,35–37
To understand the xylitol separation trends more thoroughly for pH control hydrolysates, three different retention times of 1, 3 and 10 minutes were chosen for xylitol separation using the ion-exchange method on fermentation hydrolysates with and without pH control. A mass balance was performed to account where all the xylitol ended up; this is shown in Fig. 7.
 | ||
| Fig. 7 Xylitol mass balance (%mass) over the ion-exchange separation experiments at different retention times (1, 3, and 10 minutes) for 10g L−1 initial xylose loading with and without pH control with sodium hydroxide. Each mass percent is an average of three replicates of hydrolysates obtained through fermentation using C. tropicalis, with error bars representing standard deviations. Three water washes (WW1–WW3) were used to desorb and recover the xylitol. Unaccounted xylitol was calculated by subtracting total xylitol present in the eluents (WW1–WW3) within a replicate from the xylitol present in the fermentation hydrolysate added to the ion-exchange column. Samples that do not share a letter within each pH control group resemble significant difference based on Tukey's pairwise statistical comparison (α = 0.05) (SI-Table 3A & B). | ||
Consistent trends of xylitol mass recovery (%) for samples were observed across wash cycles irrespective of retention times within each of the pH control groups (Fig. 7). Xylitol was not present in the initial collected sample eluent (represented in orange), given its affinity towards cationic ion-exchange resin. Xylitol was also minimally present in the first water wash for the no pH control group and third water wash of the pH control group. The majority of the xylitol ended up in the second water wash: for retention times of 1, 3, and 10 minutes, xylitol mass percent in the second water wash was found to be 44%, 41%, and 46% respectively for no pH control group and 45%, 48% and 50% respectively for the pH control group. The remaining xylitol eluted in the third water wash. However, the difference in trend is notable: within no pH control group xylitol favors eluting in second and third washes whereas within the pH control group it favors the first and second washes. This can be rooted in the fact that in the absence of additional sodium atoms (no pH control group) xylitol favors a strong complex with Na+ form of the resin while the presence of Na+ from sodium hydroxide favors the eluting with the water washes over binding with Na+ resin form (Hämäläinen and Lönnberg, 1991). The xylitol that was unaccounted for could be retained on the column, or could simply be from experimental error, given the size of the error bars. Water washes are preferable to avoid costs and environmental impacts associated with other reagents. This study indicated a ten minute retention time is sufficient, if not optimal, for xylitol recovery above 90%.
4. Conclusions
The objectives of this work were to evaluate xylitol production under different fermentation conditions (initial xylose loadings, and pH adjustment/control methods), and to develop novel ion-exchange separation methods using repurposed Amberlite IRC 748 resin. With a strain of C. tropicalis for fermentation, we found that using a feed xylose concentration of 10 g L−1 resulted in the most favorable xylitol yields (75%) with no significant improvement in yield or titer by maintaining a pH near 6.5 vs. having no pH control or adjustment throughout fermentation. The ion-exchange chromatography experiments demonstrated the ability of the Amberlite resin to first retain xylitol and separate it from the fermentation hydrolysate, and subsequently simply wash it off the ion-exchange column using deionized water, resulting in 90% recovery of the xylitol produced from fermentation. This results in a dissolved, aqueous xylitol solution, ready for crystallization or conversion to other chemicals or products. Ongoing and future work includes process modeling, techno-economic analysis, and life cycle assessment to evaluate the economic viability and potential environmental impacts of the scale-up of this novel laboratory process.Author contributions
SC contributed to conceptualization, data curation, formal analysis, investigation, methodology, resources, supervision, validation, visualization, and writing (original draft, review, and editing). CER, CAT, LWB, MHC, ICD, WPR, and GIM contributed to data curation, formal analysis, investigation, resources, visualization, and writing (original draft, review, and editing). AWS contributed to investigation, resources, and visualization. ECB contributed to conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, resources, supervision, validation, visualization, and writing (original draft, review, and editing).Conflicts of interest
There are no conflicts to declare.Data availability
Some data is available in the supplementary information (SI) file attached to this article. The rest of the experimental data is available in the Boston College Dataverse at https://libguides.bc.edu/dataverse.Acknowledgements
The authors thank the Morrissey College of Arts and Sciences at Boston College for supporting the work of CER, CAT, LWB, MHC, ICD, WPR, GIM, and AWS with Undergraduate Research Fellowships during some semesters. The authors also thank the Department of Engineering at Boston College for supporting MHC as a summer undergraduate research assistant in 2024. Finally, the authors thank Antonio Carlos Freitas dos Santos, Catherine Hoar, and Marek Domin for their collaboration and assistance with various laboratory equipment.References
- T. Werpy, G. Petersen. Top Value Added Chemicals from biomass: Volume I – Results of screening for potential candidates from sugars and synthesis gas, No. DOE/GO-102004-1992, 15008859, doi: DOI:10.2172/15008859.
- Y. Delgado Arcaño, O. D. Valmaña García, D. Mandelli, W. A. Carvalho and L. A. Magalhães Pontes, Xylitol: A review on the progress and challenges of its production by chemical route, Catal. Today, 2020, 344, 2–14, DOI:10.1016/j.cattod.2018.07.060.
- S. O. Ramchuran, F. O'Brien, N. Dube and V. Ramdas, An overview of green processes and technologies, biobased chemicals and products for industrial applications, Curr. Opin. Green Sustainable Chem., 2023, 41, 100832, DOI:10.1016/j.cogsc.2023.100832.
- R. S. Prakasham, R. S. Rao and P. J. Hobbs, Current trends in biotechnological production of xylitol and future prospects, Curr. Trends Biotechnol. Pharm., 2009, 3, 8–36 CAS.
- R. S. Rao, C. P. Jyothi, R. S. Prakasham, P. N. Sarma and L. V. Rao, Xylitol production from corn fiber and sugarcane bagasse hydrolysates by Candida tropicalis, Bioresour. Technol., 2006, 97, 1974–1978, DOI:10.1016/j.biortech.2005.08.015.
- S. Mathur, D. Kumar, V. Kumar, A. Dantas, R. Verma and K. Kuca, Xylitol: Production strategies with emphasis on biotechnological approach, scale up, and market trends, Sustainable Chem. Pharm., 2023, 35, 101203, DOI:10.1016/j.scp.2023.101203.
- L. Venkateswar Rao, J. K. Goli, J. Gentela and S. Koti, Bioconversion of lignocellulosic biomass to xylitol: an overview, Bioresour. Technol., 2016, International Conference on New Horizons in Biotechnology (NHBT-2015), vol. 213, pp. 299–310, DOI:10.1016/j.biortech.2016.04.092.
- S. Irmak, H. Canisag, C. Vokoun and B. Meryemoglu, Xylitol production from lignocellulosics: are corn biomass residues good candidates?, Biocatal. Agric. Biotechnol., 2017, 11, 220–223, DOI:10.1016/j.bcab.2017.07.010.
- M. Besson, P. Gallezot and C. Pinel, Conversion of biomass into chemicals over metal catalysts, Chem. Rev., 2014, 114, 1827–1870, DOI:10.1021/cr4002269.
- N. L. Mohamad, S. M. Mustapa Kamal and M. N. Mokhtar, Xylitol Biological Production: A Review of Recent Studies, Food Rev. Int., 2015, 31, 74–89, DOI:10.1080/87559129.2014.961077.
- A. K. Singh, A. K. Pandey, M. Kumar, T. Paul and N. A. Gaur, Improved xylitol production by the novel inhibitor-tolerant yeast Candida tropicalis K2, Environ. Technol., 2024, 45, 1–15, DOI:10.1080/09593330.2022.2095227.
- E. Tamburini, S. Costa, M. G. Marchetti and P. Pedrini, Optimized Production of Xylitol from Xylose Using a Hyper-Acidophilic Candida tropicalis, Biomolecules, 2015, 5, 1979–1989, DOI:10.3390/biom5031979.
- L.-F. Chen and C.-S. Gong, Fermentation of Sugarcane Bagasse Hemicellulose Hydrolysate to Xylitol by a Hydrolysate-Acclimatized Yeast, J. Food Sci., 1985, 50, 226–228, DOI:10.1111/j.1365-2621.1985.tb13315.x.
- N. L. Mohamad, S. M. Mustapa Kamal, N. Abdullah and I. Ismail, Evaluation of Fermentation Conditions by Candida tropicalis for Xylitol Production from Sago Trunk Cortex, BioResources, 2013, 8, 2499–2509, DOI:10.15376/biores.8.2.2499-2509.
- L. Xu, L. Liu, S. Li, W. Zheng, Y. Cui, R. Liu and W. Sun, Xylitol Production by Candida tropicalis 31949 from Sugarcane Bagasse Hydrolysate, Sugar Tech, 2019, 21, 341–347, DOI:10.1007/s12355-018-0650-y.
- S. S. Silva, R. M. Ramos, C. G. A. Rodrigues and I. M. Mancilha, Downstream processing for xylitol recovery from fermented sugar cane bagasse hydrolysate using aluminium polychloride, Z. Für Naturforschung C, 2000, 55, 10–15, DOI:10.1515/znc-2000-1-204.
- J. Wei, Q. Yuan, T. Wang and L. Wang, Purification and crystallization of xylitol from fermentation broth of corncob hydrolysates, Front. Chem. Eng. China, 2010, 4, 57–64, DOI:10.1007/s11705-009-0295-1.
- V. Ahuja, D. Dasgupta, S. Kshirsagar, P. Ghosh, S. More, P. Gupta, B. Behera and T. Bhaskar, Crystalline xylitol production from corncob biomass with oral toxicity analysis, Ind. Crops Prod., 2022, 187, 115407, DOI:10.1016/j.indcrop.2022.115407.
- S. S. Queiroz, F. M. Jofre, S. I. Mussatto, M. das and G. A. Felipe, Scaling up xylitol bioproduction: Challenges to achieve a profitable bioprocess, Renewable Sustainable Energy Rev., 2022, 154, 111789, DOI:10.1016/j.rser.2021.111789.
- Y. P. Cerceau-Alves, F. A. Fernandes Antunes, S. Silverio da Silva and M. B. S. Forte, From by- to bioproducts: selection of a nanofiltration membrane for biotechnological xylitol purification and process optimization, Food Bioprod. Process., 2021, 125, 79–90, DOI:10.1016/j.fbp.2020.10.005.
- R. Desiriani, M. T. A. P. Kresnowati and I. G. Wenten, Membrane-based downstream processing of microbial xylitol production, Int. J. Technol., 2017, 8, 1393, DOI:10.14716/ijtech.v8i8.726.
- R. M. Cadete, M. A. Melo, K. J. Dussán, R. C. L. B. Rodrigues, S. S. Silva, J. E. Zilli, M. J. S. Vital, F. C. O. Gomes, M.-A. Lachance and C. A. Rosa, Diversity and physiological characterization of d-xylose-fermenting yeasts isolated from the brazilian amazonian forest, PLoS One, 2012, 7, e43135, DOI:10.1371/journal.pone.0043135.
- C.-H. Ko, P.-C. Chiu, C.-L. Yang and K.-H. Chang, Xylitol conversion by fermentation using five yeast strains and polyelectrolyte-assisted ultrafiltration, Biotechnol. Lett., 2008, 30, 81–86, DOI:10.1007/s10529-007-9507-2.
- E. A. Martínez, J. B. de Almeida e Silva, M. Giulietti and A. I. N. Solenzal, Downstream process for xylitol produced from fermented hydrolysate, Enzyme Microb. Technol., 2007, 40, 1193–1198, DOI:10.1016/j.enzmictec.2006.09.003.
- A. K. Singh, F. Deeba, M. Kumar, S. Kumari, S. A. Wani, T. Paul and N. A. Gaur, Development of engineered Candida tropicalis strain for efficient corncob-based xylitol-ethanol biorefinery, Microb. Cell Factories, 2023, 22, 201, DOI:10.1186/s12934-023-02190-3.
- Y. Lu, R. Warner, M. Sedlak, N. Ho and N. S. Mosier, Comparison of glucose/xylose cofermentation of poplar hydrolysates processed by different pretreatment technologies, Biotechnol. Prog., 2009, 25, 349–356, DOI:10.1002/btpr.158.
- E. Winkelhausen and S. Kuzmanova, Microbial conversion of d-xylose to xylitol, J. Ferment. Bioeng., 1998, 86, 1–14, DOI:10.1016/S0922-338X(98)80026-3.
- J. Wu, J. Hu, S. Zhao, M. He, G. Hu, X. Ge and N. Peng, Single-cell Protein and Xylitol Production by a Novel Yeast Strain Candida intermedia FL023 from Lignocellulosic Hydrolysates and Xylose, Appl. Biochem. Biotechnol., 2018, 185, 163–178, DOI:10.1007/s12010-017-2644-8.
- T.-B. Kim and D.-K. Oh, Xylitol production by Candida tropicalis in a chemically defined medium, Biotechnol. Lett., 2003, 25, 2085–2088, DOI:10.1023/B:BILE.0000007069.74876.0c.
- B. S. Ko, J. Kim and J. H. Kim, Production of Xylitol from d-Xylose by a Xylitol Dehydrogenase Gene-Disrupted Mutant of Candida tropicalis, Appl. Environ. Microbiol., 2006, 72, 4207–4213, DOI:10.1128/AEM.02699-05.
- L. Zhang, Z. Chen, J. Wang, W. Shen, Q. Li and X. Chen, Stepwise metabolic engineering of Candida tropicalis for efficient xylitol production from xylose mother liquor, Microb. Cell Factories, 2021, 20, 105, DOI:10.1186/s12934-021-01596-1.
- M. G. C. da Silva, R. L. S. Canevesi, R. A. Welter, M. G. A. Vieria and E. A. da Silva, ., Chemical equilibrium of ion exchange in the binary mixture Cu2+ and Ca2+ in calcium alginate, Adsorption, 2015, 21, 445–458, DOI:10.1007/s10450-015-9682-8.
- R. K. Misra, S. K. Jain and P. K. Khatri, Iminodiacetic acid functionalized cation exchange resin for adsorptive removal of Cr(VI), Cd(II), Ni(II) and Pb(II) from their aqueous solutions, J. Hazard. Mater., 2011, 185, 1508–1512, DOI:10.1016/j.jhazmat.2010.10.077.
- Z. Yu, T. Qi, J. Qu and Y. Guo, Application of mathematical models for ion-exchange removal of calcium ions from potassium chromate solutions by Amberlite IRC 748 resin in a continuous fixed bed column, Hydrometallurgy, 2015, 158, 165–171, DOI:10.1016/j.hydromet.2015.10.015.
- A. A. M. Abusultan, J. A. Wood, T. Sainio, A. J. B. Kemperman and W. G. J. van der Meer, Ion exchange resin – Bipolar membrane electrodialysis hybrid process for reverse osmosis permeate remineralization: Cation exchange resins equilibria and kinetics, Sep. Purif. Technol., 2023, 317, 123798, DOI:10.1016/j.seppur.2023.123798.
- M. M. Hämäläinen and H. Lönnberg, Complexing of sugars and sugar alcohols with metal ions: a comparative study by ion-exchange chromatography, Carbohydr. Res., 1991, 215, 357–360, DOI:10.1016/0008-6215(91)84035-D.
- Y. Watanabe and S. Adachi, Estimation of the Binding Constant of Sugar Alcohol to Sodium Ion by Using Its Apparent Distribution Coefficients to Cation-exchange Resins with Different Cross-linkages, Jpn. J. Food Eng., 2023, 24, 77–82, DOI:10.11301/jsfe.23631.
| This journal is © The Royal Society of Chemistry 2026 |