
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceSustainable valorization of food waste for microalgal production of value-added compounds, bio-oil, and biochar
Himanshu Srivastavaa,
Shivani Dimria,
Bhawna Bishta,
Dhruva,
Rahini Parsoyaa,
Krishna Kumar Jaiswalb,
Mikhail S. Vlaskin c,
Manisha Nandad,
Sanjay Kumara,
Harish Chandra Joshie and
Vinod Kumar*af
c,
Manisha Nandad,
Sanjay Kumara,
Harish Chandra Joshie and
Vinod Kumar*af
aAlgal Research and Bioenergy Laboratory, Department of Food Science & Technology, Graphic Era (Deemed to be University), Dehradun 248002, India. E-mail: vinodkdhatwalia@gmail.com
bBioprocess Engineering Laboratory, Department of Green Energy Technology, Pondicherry University, Puducherry 605014, India
cJoint Institute for High Temperatures of the Russian Academy of Sciences, Moscow 125412, Russian Federation
dDepartment of Microbiology, Graphic Era (Deemed to be University), Dehradun 248002, India
eDepartment of Chemistry, Graphic Era (Deemed to be University), Dehradun, Uttarakhand 248002, India
fPeoples' Friendship University of Russia (RUDN University), 6 Miklukho-Maklaya Street, Moscow, 117198, Russian Federation
First published on 1st June 2026
Abstract
The environmental burden of food waste (FW), a major source of greenhouse gases, necessitates advanced upcycling strategies. This study presents an integrated biorefinery to sustainably valorize FW into a suite of valuable products. Physically pretreated FW was utilized as an alternative culture medium at varying concentrations (10–50%) for microalgae cultivation. This approach successfully recycled waste nutrients and significantly enhanced the microalgae's bioproduct profile. Among saturated fatty acids (SFAs), palmitic acid (C16:0) was the dominant component in both groups and increased significantly from 42.60 ± 0.40% in the control to 47.40 ± 0.52% in FWCM. Similarly, arachidic acid (C20:0) increased from 1.67 ± 0.49% to 3.03 ± 0.15% following food waste culture medium (FWCM). The antioxidant capacity was also modulated, as evidenced by changes in DPPH scavenging activity, total phenolic (TPC) and flavonoid (TFC) content, and reactive oxygen species (ROS) levels. Food waste solid residues were subsequently processed via hydrothermal liquefaction (HTL) at 250 °C and 350 °C to produce bio-oil and hydrochar, while the hydrochar was further pyrolyzed at 550, 650, and 750 °C to produce biochar. The biochar was thoroughly characterized using FTIR, SEM, and XRD. This work demonstrates a circular economy model that mitigates FW by generating a nutrient-rich growth medium and sequentially converting the biomass into energy (bio-oil) and biochar.
Sustainability spotlightThis research presents a transformative circular bioeconomy model that addresses two critical environmental challenges simultaneously: food waste management and sustainable bioproduct generation. We demonstrate that food waste, a major source of greenhouse gases, can be efficiently upcycled into a nutrient-rich culture medium for microalgae. This process not only diverts waste from landfills but also eliminates the need for synthetic culture media for algae cultivation. The resulting S. abundans biomass shows enhanced growth and a superior biochemical profile, rich in lipids and antioxidants for nutraceuticals. Furthermore, the biomass is valorized through hydrothermal liquefaction, co-producing bio-oil and biochar. By converting a major environmental burden into high-value bioproducts such as bio-oil, functional metabolites, and biochar, this work advances circular bioeconomy principles and reduces resource consumption, waste generation, and greenhouse gas emissions. |
1 Introduction
FW has grown substantially on a global scale in recent years, raising serious environmental and socioeconomic issues.1 Food that is appropriate for human utilization but is discarded after spoiling or expiration is referred to as FW.2 Every year, around 1.3 billion tonnes of food is thought to be wasted along the whole supply chain from farming to consumption.3 The disposal of FW has severe environmental implications. FW is a major source of methane, a potent greenhouse gas (GHG) that contributes substantially to global warming.4 When not properly managed, it leads to soil, air and water contamination, posing significant threats to public health.5 Additionally, FW represents a loss of valuable resources such as water, land, energy, and labour that were utilized during food production. Landfilling remains the most common method of FW disposal; however, it consumes valuable land resources and releases harmful chemicals and gases, including pesticides and leachates, into the environment.6Large-scale sustainable valorisation of food waste (FW) is essential for reducing the environmental and economic burdens of FW.7,8 Biochemical techniques (fermentation, composting and anaerobic digestion) and thermochemical techniques (hydrothermal carbonisation (HTC), pyrolysis and gasification) are commonly used to uphold FW into valuable bioenergy.9 However, there are three major problems associated with biochemical techniques, namely that (1) they are inherently slow and the conversion efficiency is low owing to the complexity of food waste; (2) they depend on microorganisms, which are susceptible to variation in operating conditions; (3) they require additional rebuilding treatments to transform main products with high molecular weight variants.10 The valorisation of FW into functional food ingredients is a transformative approach toward achieving sustainability in the food industry. While challenges persist, lack of integrated and circular systems capable of simultaneously recovering energy, nutrients, and high-value products.11
To address these issues, the valorisation of FW transforms waste into value-added products which has arisen as a sustainable approach. Valorisation not only reduces waste generation but also offers economic opportunities through the recovery of valuable bioactive compounds.12 Various extraction techniques can be employed to obtain phenols, polysaccharides, aromatic oils, colorants, proteins, flavour compounds, enzymes and fibres from FW. Additionally, FW can serve as a substrate for producing biofuels and bioenergy products such as butanol, ethanol, biogas and biodiesel.13
The high content of energy-rich molecules in FW are lipids (70–300 mg g−1), proteins (60–100 mg g−1), and carbohydrates (300–600 mg g−1) which makes it an ideal candidate for bioenergy generation.14 These components can be transformed into useful forms of energy through thermochemical methods such as gasification, pyrolysis and hydrothermal treatment. Among these, hydrothermal liquefaction (HTL) is particularly appropriate for high-moisture feedstocks like FW as it allows direct processing without prior drying. Under high temperature and pressure, HTL converts wet biomass into a high-energy biocrude oil and highly nutritious aqueous phase along with minor solid and gaseous by-products.15 Earlier research has demonstrated that HTL is best method for conversion of biofuel and biochar. Bayat et al.16 stated that HTL was used for conversion of FW into bio-crude oil at moderate operating conditions, including temperatures of 240–295 °C, reaction durations of 0–60 min, and 15 wt% solids loading. The highest yield of bio-crude oil (27.5 wt%) were obtained at 240 °C for 30 minutes. Aierzhati et al.17 uses HTL for transforming food residues into biocrude oil with high temperatures (280–380 °C) and time (10–60 min). Analysis of different feedstocks represented various biochemical profiles (protein, carbohydrate and lipid) and generated about 2–79% biocrude oil for the respective optimized HTL time and temperature. Saengsuriwong et al.18 determined HTL for production of biocrude oil using a simplified high-through reactor. A highest biocrude oil of approximately 40% w/w dry basis and energy recovery greater than 70% were produced.
Furthermore, implementing systematic management practices across the food production and consumption chain can significantly reduce FW generation at its source. Therefore, a dual strategy emphasizing both the prevention and valorisation of FW is vital for achieving sustainable resource management, reducing environmental impact and fostering long-term ecological resilience.7
Most of the current research ignores other categories of waste, such as dairy or meat by-products which are suitable for different processing techniques is in favour of concentrating on certain waste kinds, such as fruit and vegetable residues. Moreover, standardised approaches for assessing the effectiveness of valorisation strategies across various waste categories are lacking. This discrepancy restricts the scalability of effective valorisation techniques and makes it difficult to compare findings from different investigations. Although advanced technologies like artificial intelligence (AI) and omics techniques show the potential in maximising the value of FW is still understudied.19
The aim of this study is to investigate the potential of food waste (FW) as a sustainable feedstock for hydrothermal liquefaction (HTL)-based bioresource recovery and biochar production. This study involves the collection, preparation, and characterization of FW based on its proximate and physicochemical properties to evaluate its suitability for thermochemical conversion. Furthermore, FW was processed into a food waste culture medium (FWCM) for microalgae cultivation and hydrochar at different HTL operating temperatures and determine the influence of temperature variations on product distribution, particularly the yield of hydrochar and bio-oil. The obtained hydrochar was further pyrolyzed to generate biochar. Comprehensive characterization of the produced biochar was carried out using analytical techniques such as scanning electron microscopy (SEM), X-ray diffraction (XRD), and other physicochemical analyses to better understand its structural and surface properties.
2 Materials and methods
For preparation of inoculum culture of microalgae, Scenedesmus abundans (SA) was cultivated in Blue-Green 11 medium (BG-11) under continuous illumination with white LED light sources at an intensity of approximately 300 µmol m−2 s−1 with a photoperiod of 16![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :8 (light:dark) and temperature 25 ± 1 °C. The culture was shaken manually after 6 h intervals, and its growth was assessed every day by recording the optical density (OD) at 680 nm until it reached the log phase.
:8 (light:dark) and temperature 25 ± 1 °C. The culture was shaken manually after 6 h intervals, and its growth was assessed every day by recording the optical density (OD) at 680 nm until it reached the log phase.
2.1 Collection of FW and preparation of FWCM
FW (fruit and vegetable peels, rice, bread, Indian flatbread) was collected from university mess. Solid materials such as non-edible components and bone sludge were manually separated during the collection of FW. The proximate analysis like moisture content (%), ash content (%), fat content (%) and fiber content (%) of raw FW was determined according to the AOAC protocol.20 Carbohydrate content was estimated by using the difference method as shown in eqn (1).| Carbohydrates (%) = {100 − (moisture content + ash content + protein content + fat content + fibre content)} | (1) |
The amount of nitrogen was determined by utilizing CHNS elemental analyzer (Elementar vario EL cube, 19151007, Central Instrumentation Facility, Pondicherry University) and then the crude protein was determined by using the following formula.21
| Crude protein (% w/w) = nitrogen (%) × 6.25 | (2) |
The FW was subjected to physical pretreatment using a grinder (500 g + 1 L water). The pre-treated FW mixture was then left overnight for sedimentation process to separate the remaining solid and liquid homogeneous mixture. The liquid FW portion was further filtered through a 4 layers muslin fabric to remove any excess solid and tiny suspended particles. The residue and liquid portion of the slurry was subjected to centrifugation at 5000 rpm for 5 minutes for separation. The liquid FW was then autoclaved and allowed to cool down at room temperature. The pH was maintained between 7 and 8 throughout the experiment. The liquid FW culture medium was marked as FWCM and then stored overnight at 4 °C in a refrigerator prior for the cultivating microalgae to prevent inhibition of microbial degradation (Fig. 1).
 | ||
| Fig. 1 Schematic diagram of the integrated biorefinery process for the valorization of FW into a FWCM medium for microalgae, bio-oil, and biochar. | ||
2.2 Characterization of FWCM
FWCM medium was then used for determining the chemical oxygen demand (COD), nitrogen content (NC), total chlorine (TC), hardness (ppm), calcium carbonate (ppm), pH, elemental analysis etc.22 COD of the sample was determined using the standard dichromate oxidation followed by titration with ammonium iron(II) sulphate. COD values were determined based on difference in titrant volume consumed by the sample and the blank.23 Estimation of physiochemical parameters including total hardness, calcium carbonate (CaCO3) content, pH and total chlorine were checked using a quick multi parameter test strip. The strip was dipped into the sample for 1–2 seconds, allowing the reactive pads to develop a color change. The color changes on the strip were immediately matched with the comparative color cart provided with the strips, and the corresponding values were noted.2.3 Experimental setup
Food Waste Culture Medium (FWCM) was mixed with inoculum culture (containing BG-11) of microalgae OD (0.6) at ratio (90:10; 80:20; 70:30 and 60:40) in 250 mL flask. Cultures were grown under continuous illumination provided by white LED lamps at a light intensity of 300 µmol m−2 s−1, with a 16:8 h light: dark photoperiod and a constant temperature of 25 ± 1 °C. The cultures were manually agitated at 6 h intervals. Growth was monitored daily by measuring the optical density at 680 nm until the culture reached the stationary phase (14th days). The growth curve was prepared using OD values. Out of all the experiments, the optimum concentration was (BG11 80 mL:FWCM 20 mL) used to cultivate microalgae as it shows the maximum growth in a 5 L pond with 2.5 L working volume. The microalgae grown in BG11 media is only considered as control. After 14 days, the cultivated microalgae biomass was centrifuged at 5000 rpm for 15 min for harvesting. The collected biomass was then lyophilized, weighed, stored, and utilized for biochemical analysis, total phenolic content (TPC), total flavonoid content (TFC), and reactive oxygen species (ROS). Functional groups of the sample and control biomass were analyzed using a FTIR spectrophotometer (Bruker Invenio-S).
2.4 Biochemical analysis
Effect of FWCM on microalgae pigment content was estimated on 10th day according to Lichtenthaler.24 2 mL of algal culture was centrifuged at 5500 rpm for 5 min. The harvested biomass pellets were mixed with 2 mL of methanol (99%) and incubated for 30 min at 45 °C. The absorbance of supernatant obtained after centrifugation at 5500 rpm for 5 minutes was measured at wavelength 665.2 nm, 652.4 nm, 470 nm and 750 nm. Turbidity from absorbencies at 665.2 nm, 652.4 nm, and 470 nm were corrected by subtracting them at 750 nm. The following formulas were utilized to determine different pigment levels in µg mL−1.| Chlorophyll a (Chl a) = 16.72OD665.2 − 9.16OD652.4 | (3) |
| Chlorophyll b (Chl b) = 34.09OD652.4 − 15.28OD665.2 | (4) |
| Carotenoid (Car) (µg mL−1) = (1000OD470 − 1.63 Chl a − 104.9 Chl b)/221 | (5) |
Carbohydrate analysis was measured on 14th day by mixing dry microalgae biomass with 5% H2SO4 and autoclaving for 1 h at 121 °C and 15 psi. pH was calibrated to 7 using 10 M NaOH after cooling. After centrifuging the sample at 8000 rpm for 10 min, the resulting supernatant was used for quantification of carbohydrates using the phenol-sulfuric acid method.25
Total lipids were estimated by the modified Bligh–Dyer method.26,27
The following equation was applied to estimate total lipid content.
 | (6) |
The extracted lipids were trans-esterified into fatty acid methyl esters (FAMEs) following the method of Arora et al.26 Briefly, lipids were treated with 6% methanolic sulfuric acid (H2SO4) at 90 °C for 1 h. The resulting FAMEs were then extracted by adding hexane, and the mixture was washed with distilled water. After centrifugation at 5000 rpm for 5 minutes, the upper hexane layer containing the purified FAMEs was collected for analysis. Fatty acid methyl esters (FAMEs) were analyzed using an Agilent GC system equipped with a flame ionization detector (FID). Separation was performed on an Agilent HP-88 capillary column (100 m × 0.250 mm i.d., 0.20 µm film thickness). The injector temperature was maintained at 250 °C, and the detector temperature was set at 280 °C. Helium was used as the carrier gas with a column flow rate of 0.9 mL min−1. The FID gas flows were set as follows: air flow 400 mL min−1, hydrogen fuel flow 40 mL min−1, and helium makeup flow 25 mL min−1.
2.5 Estimation of DPPH, TPC, TFC and ROS
The extract was prepared by mixing 1 g of sample with 10 mL of 80% ethanol. The mixture was shaken overnight at room temperature to enhance extraction efficiency. The sample was then centrifuged at 8000 rpm for 5 min, and the supernatant was collected. The supernatant was dried using a water bath, and the dried extract was reconstituted in ethanol to obtain a final concentration of 1 mg mL−1. This extract was subsequently used for the determination of DPPH radical scavenging activity, total phenolic content (TPC), and total flavonoid content (TFC).A stock solution of DPPH (12 mg/50 mL) in 100% ethanol was prepared and kept overnight in dark. The working solution of DPPH was prepared by diluting the stock solution in the ratio 1:6 (v/v), 10 mL of stock solution was dissolved in 60 mL of 100% ethanol. Subsequently, 2.9 mL of DPPH working solution was combined with 100 µL of each microalgal extract in separate test tubes and incubated for 120 minutes in the dark. Finally, the absorbance was recorded at 515 nm, and the percentage of radical-scavenging activity was computed using the following equation.
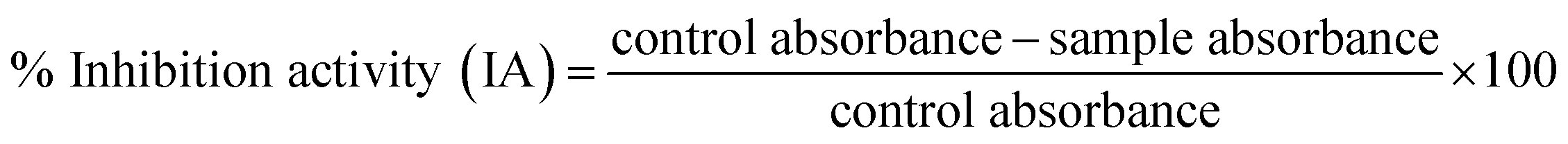 | (7) |
The TPC of control algal biomass SA and biomass grown in FWCM was estimated by using the Folin–Ciocalteu reagent method.28 The calibration curve for TPC was prepared by using gallic acid as standard. 5 mL of gallic acid was prepared with a concentration of 1 mg mL−1 in distilled water. Varying concentration of gallic acid was taken in test tubes ranging from (20–120 µL) and distilled water was added to bring the volume up to 1 mL. Following this, 500 µL of 10% Folin–Ciocalteu reagent was added and incubated for 3 minutes. Subsequently, 1.5 mL of 20% Na2CO3 was added, followed by 15 minutes of mixing. Finally, 7 mL of distilled water was poured to each test tube and incubated for 45 minutes. The total phenolic content (TPC) of the microalgal extracts was determined spectrophotometrically by measuring the absorbance at 760 nm.
The total flavonoid content (TFC) for both the control (SA) and FWCM biomass was quantified using the aluminum chloride colorimetric method, with quercetin serving as the standard ref. 29. A quercetin stock solution (1 mg mL−1) was prepared in distilled water, from which a series of working standards (20–120 µL) were transferred to test tubes and adjusted to a final volume of 1 mL with ethanol.
The reaction was initiated by adding 75 µL of 5% NaNO2 followed by a 6 min incubation. Subsequently, 150 µL of 10% AlCl3 was added and allowed to react for 5 min. The mixture was then stabilized by adding 500 µL of 1 M NaOH and 2.5 mL distilled water. The TFC in the biomass extracts was determined by measuring the absorbance at 510 nm, with results expressed as quercetin equivalents.
The intracellular reactive oxygen species (ROS) scavenging potential of the extracts was evaluated using a hydrogen peroxide (H2O2) radical scavenging assay.28 Briefly, 1.5 mL of the microalgal extract was mixed with, 2.5 mL of 10 mM solution. The mixture was homogenized and incubated at room temperature for 30 minutes. Following incubation, the absorbance was recorded at 230 nm. The percentage of ROS inhibition was calculated using the following equation:
 | (8) |
2.6 Hydrothermal liquefaction of FW residue
Food waste was physically treated to produce a food waste culture (FWC) medium after filtration. The remaining solid residues were collected, dried at 70 °C, and subsequently utilized as feedstock for hydrothermal liquefaction (HTL) experiments. Hydrothermal liquefaction (HTL) of physically pretreated FW solid residues was carried out in a custom-designed 40 mL stainless steel reactor. A slurry containing 20 g of dry FW solid residues (12 wt%) and 146.7 mL distilled water was prepared and loaded into 5 reactors and the experiment was repeated three times. The HTL experiments were conducted at 250 °C and 350 °C with a retention time of 45 min. After the completion of the reaction, both the solid and liquid phases were separated by filtration. Dichloromethane was added to the liquid phase to extract the light oil. The solid residue washed three to four times with acetone to extract heavy oil. To obtain the biochar, the washed solid residue was dried at 50 ± 5 °C and subsequently weighed. Light and heavy oil were mixed and weighted. Gas phases were produced during the process and were not considered in this study.The hydrochar (solid residues) obtained from hydrothermal liquefaction at 350 °C were combined and subsequently pyrolyzed at 550 °C, 650 °C, and 750 °C for 6 hours to produce biochar. The resulting biochar was characterized by scanning electron microscopy (SEM), and X-ray diffraction (XRD).
2.7 Statistical analysis
All experiments were conducted in triplicate (n = 3). Data are presented as mean ± standard deviations (SD). MS excel was used for statistical analysis. To assess statistically significant differences, one-way and two-way ANOVA with replications was utilized (SI). A p-value of < 0.05 was regarded as significant.3 Result and discussion
3.1 Proximate composition of raw FW
The initial characterization of the raw FW revealed a high moisture content of 84.19 ± 1.78% which is characteristic of organic waste fractions. The organic composition was primarily dominated by proteins (8.98%), followed by carbohydrates (3.37 ± 1.29%) and fats (1.65 ± 1.00%). In contrast, the inorganic and fibrous fractions were relatively low, with ash and fibre contents recorded at 1.11 ± 0.86% and, 0.68 ± 0.07% respectively (Table S1).3.2 Selection of optimized microalgae FWCM
Based on the preliminary trails, the SA inoculum and FWCM were selected at different concentrations in the ratio (90:10; 80:20; 70:30 and 60:40) as mentioned in SI Table 2. SA was taken as a control for the experiment. Out of all the experiments, the optimum concentration was (FWCM 80:20). Fig. 1 shows a different concentration of FWCM medium in SA microalgae. SI Fig. 1 shows the optimized FWCM grown in SA microalgae (Fig. 2).
 | ||
| Fig. 2 Cultivation of microalgae in FWCM at varying concentrations (10%, 20%, 30%, 40%) with standard BG-11. | ||
3.3 Physicochemical characterization of FWCM before and after microalgae cultivation
The Food Waste Culture Medium (FWCM) medium initially exhibited a very high chemical oxygen demand (COD) of 1520 ± 3.5 mg L−1, indicating a substantial organic load. Upon dilution, the COD decreased markedly 340 ± 1.2 mg L−1 upon dilution. Following microalgal cultivation, COD further declined to 196 ± 5.2 mg L−1, demonstrating efficient biodegradation and nutrient uptake by microalgae. It is important to note that COD represents the oxygen demand of both biodegradable and non-biodegradable fractions, and its variability is strongly influenced by food waste composition and environmental conditions, with reported values showing wide fluctuations in different systems.30Total nitrogen (TN) in the diluted FWCM appeared higher, likely due to improved solubilization and homogenization of nitrogenous compounds and addition algae media (BG11); however, it was significantly reduced to 45.63 mg L−1 after algal cultivation, confirming efficient nitrogen assimilation.
Previous studies have reported ammonium concentrations ranging from 985 to 134 mg L−1 and total nitrogen levels between 3.973 and 267 mg L−1, depending on substrate characteristics and treatment conditions.31
Similarly, total phosphorus (TP) decreased sharply from 12.08 mg L−1 in the diluted FW medium to 1.08 mg L−1, indicating effective phosphorus removal by microalgae. Hardness decreased from 425 ± 1 ppm in FWCM to 120 ± 0.5 ppm in diluted and algae-treated water. Additionally, the pH increased from acidic values (6.0) to alkaline (8.4) after cultivation, indicating active photosynthesis and CO2 uptake by microalgae (Table 1). Overall, these results highlight the strong potential of microalgal systems for simultaneous organic load reduction and nutrient recovery from FW derived media.
:20), and microalgae treated food waste culture medium (FWCM)
| Parameters | Raw FW | Diluted FW (80:20) |
Microalgae treated FWCM |
|---|---|---|---|
| Values are expressed as mean ± standard deviation (n = 3). Different superscript letters within the same row indicate statistically significant differences among groups according to one-way ANOVA at p < 0.05. | |||
| COD (mg L−1) | 1520 ± 3.5a | 340 ± 1.5b | 196 ± 5.2c |
| TN (mg L−1) | 1.58 ± 0.15b | 190.07 ± 1.02a | 45.63 ± 0.5c |
| TP (mg L−1) | 27.88 ± 0.05a | 12.84 ± 0.02b | 1.084 ± 0.03c |
| Total chlorine (ppm) | 0a | 0a | 0a |
| Hardness (ppm) | 425 ± 1a | 120 ± 0.5b | 120 ± 1.2b |
| pH | 6.0 ± 0.2b | 6.2 ± 0.5b | 8.4 ± 1.2a |
According to the one-way ANOVA analysis (SI Table 4), there is a statistically significant difference between treatments. Treatments, especially dilution and microalgae treatment, significantly change water quality. This analysis revealed that both treatment type and parameter significantly affect the measured values (p < 0.05). Additionally, a significant interaction effect (p < 0.05) indicated that the impact of treatment varies across different water quality parameters.
3.4 Growth and biomass content of microalgae cultivated in FWCM
The growth curve of microalgae cultivated in FWCM at a concentration ratio of 80:20, along with the control medium, is presented in Fig. 3. The graph exhibited the growth pattern of SA as OD at 680 nm over 14 days under two conditions: control and FWCM. The FWCM microalgae shows a consistent OD throughout the cultivation period, indicating rapid growth and faster biomass accumulation as compared to the control. A sharp increase in OD is observed between day 1 and day 3 in the FWCM sample, suggesting that the additional organic nutrients accelerate early growth. Both samples reached stationary phase around day 12–14 but the final OD of the FWCM microalgae remains maximum, showing FWCM addition enhances overall microalgae productivity.
 | ||
| Fig. 3 Growth curve of SA with FWCM supplementation. | ||
Biomass content (g L−1) of microalgal cells with FWCM were analyzed and compared with the control medium. The highest biomass was 1.14 g L−1 in the optimum concentration of (BG11:FWCM-80:20) whereas in control (SA), the biomass content was 0.75 g L−1. The observed improvement aligns with previous studies utilizing FW hydrolysates. For instance, Ramandani et al.32 reported that a 10% FWCM medium generated a biomass concentration of 2.869 g L−1 for Chlorella vulgaris.
3.5 FTIR spectroscopy of microalgae biomass SA and FWCM
FTIR spectroscopy was performed to identify the functional groups present in control biomass (SA) and FWCM biomass. The observed characteristic wavenumbers and their respective vibrational mode assignments are depicted in Fig. 4, highlighting compositional differences between the samples. A composition with relative similarity was observed in the FTIR spectra of the SA (control) and FWCM. | ||
| Fig. 4 FTIR spectra of microalgae biomass SA and FWCM. | ||
The FTIR spectra of the microalgal biomass exhibited a series of distinct vibrational bands that serve as molecular fingerprints for its biochemical constituents. Aliphatic components were characterized by C–C skeletal stretch vibrations at 448 and 543 cm−1, complemented by aromatic C–H stretch vibrations observed at 693 and 867 cm−1. The carbohydrate and polysaccharide fractions were defined by C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O and C–O–C glycosidic stretch vibrations at 1033 and 1246 cm−1, respectively. Protein-related signals were particularly diverse, involving CO vibration at 1538 cm−1, corresponding to the amide II band associated with N–H bending and C–N stretching vibrations. Notably, the peak at 1640 cm−1 was assigned to the CO stretching of esters (RCOOR) or the characteristic amide-I region, confirming the presence of complex protein structures. Finally, the band at 1396 cm−1 was indicative of C–H and O–H functional groups associated with the amino acid profiles of the microalgal biomass.
O and C–O–C glycosidic stretch vibrations at 1033 and 1246 cm−1, respectively. Protein-related signals were particularly diverse, involving CO vibration at 1538 cm−1, corresponding to the amide II band associated with N–H bending and C–N stretching vibrations. Notably, the peak at 1640 cm−1 was assigned to the CO stretching of esters (RCOOR) or the characteristic amide-I region, confirming the presence of complex protein structures. Finally, the band at 1396 cm−1 was indicative of C–H and O–H functional groups associated with the amino acid profiles of the microalgal biomass.
In microalgae biomass FWCM, a decrease in spectral intensity was seen at bands 1246 cm−1 and 1538 cm−1 indicating reduced protein amino acid synthesis and stimulating lipid induction in biomass of SA linked to the treatment of FWCM components. Differences in peak intensity are indicative of compositional shifts in the biomass, primarily associated with variations in lipid accumulation and carbohydrate content.33
3.6 Biochemical estimation
In algae, chlorophyll is considered a crucial pigment involved in photosynthetic process which absorbs light energy and the shift in photosynthesis process of algal cell is benefited by an uptake in Chl content.34 The FWCM biomass sample had chlorophyll a, chlorophyll b and carotenoids contents of 2.84 ± 0.03, 1.33 ± 0.19 and 1.54 ± 0.06 µg mL−1, while the control sample had 0.90 ± 0.08, 0.36 ± 0.03 and 0.20 ± 0.01 µg mL−1. Hence, as compared to control, the concentration of Chl a, Chl b and Car content was subsequently increased after addition of FW at the 10th day. The calculated values are shown in Table 2. This result indicated that increasing FWCM can enhance the Chl and Car content of microalgal cells. The elevated carotenoid content further indicates a protective physiological response against oxidative stress generated during intensified metabolic activity under mixotrophic growth conditions. Similar observations have been reported by Giwa et al.,35 who demonstrated that Chlorella vulgaris cultivated using food waste-derived media maintained substantial chlorophyll a production despite variations in protein synthesis.:20)a
| Parameter | Control (SA) | FWCM (80:20) |
||||
|---|---|---|---|---|---|---|
| a Data are shown as mean ± standard deviation of three replicates. Different superscript letters (a, b) within the same row indicate statistically significant differences between control and treated samples at p < 0.05, based on two-way ANOVA with replication. | ||||||
| Pigment | Chl a (µg mL−1) | Chl b (µg mL−1) | Car (µg mL−1) | Chl a (µg mL−1) | Chl b (µg mL−1) | Car (µg mL−1) |
| 0.90 ± 0.08a | 0.36 ± 0.03a | 0.20 ± 0.01a | 2.84 ± 0.03b | 1.33 ± 0.19b | 1.54 ± 0.06b | |
| Carbohydrate (%) | 14.37 ± 0.5a | 17.36 ± 0.8b | ||||
| Protein (%) | 30 ± 0.08a | 21.75 ± 0.08b | ||||
| Lipid (%) | 22 ± 0.5a | 35.93 ± 0.98b | ||||
The carbohydrate content of the FWCM was 17.36 ± 0.8% and for the control it was 14.37 ± 0.5% as mentioned in Table 2. The study revealed a significant increase in carbohydrate content when FWCM was added into the microalgal culture. Incorporation of FWCM into the SA medium may provide an extra source of organic carbon C:N balance, promoting mixotrophic carbon uptake and diverting surplus carbon into storage carbohydrates. Such an increase in carbon metabolism together with nutrient regulated pathways that reduce protein synthesis under C:N conditions, leads to greater carbohydrate accumulation in microalgae cultivated with organic food waste.36
The protein content of control (SA) was 30 ± 0.08% and for the FWCM sample it was 21.75 ± 0.08%. The result showed that the FWCM biomass significantly reduced compared to the control sample. Hence, addition of FWCM reduces the protein content in the microalgae culture medium.
This reduction may be associated with variations in nitrogen availability and metabolic redistribution toward lipid and carbohydrate biosynthesis under mixotrophic conditions. Since nitrogen is a key component required for amino acid and protein synthesis, reduced nitrogen availability in food waste-derived media can limit protein accumulation and alter cellular metabolism. Similar observations have been reported in previous studies. Chew et al.36 demonstrated that microalgae cultivated in food waste-derived media (50C and 25C mixtures) showed slightly lower protein content compared with the control medium, although the reduction was less than 10%. Likewise, Zhang et al.37 reported reduced protein accumulation in microalgae grown using kitchen waste effluents relative to BG11 medium, primarily due to lower nitrogen concentrations in the diluted effluent. The authors suggested that insufficient nitrogen availability directly affected protein biosynthesis. This enhancement may be attributed to the presence of readily available organic carbon and nutrients in FWCM, which can stimulate carbon assimilation and redirect cellular metabolism toward storage lipid accumulation.38 Chew et al.36 observed that microalgae cultivated in compost-based media exhibited higher lipid content compared with the control, which was attributed to the availability of organic carbon that accelerated carbon assimilation and lipid accumulation.38 Likewise, Zeng et al.39 reported lipid contents ranging from 16.8–25.5% in mixed cultures of Chlorella vulgaris and yeast cultivated using food waste hydrolysate.
A two-way ANOVA with replication (SI Table 6) revealed that both treatment and biological parameter had a significant effect (p < 0.05). A significant interaction between treatment and parameter type was also observed indicating that the effect of treated FW sample varied across different biochemical components. The treated FW sample causes increase in Chl a, Chl b, Car, lipids but decrease in protein.
The fatty acid composition of control and FWCM microalgae biomass showed substantial alterations under FWCM supplementation (SI Table 11). Among saturated fatty acids (SFAs), palmitic acid (C16:0) was the dominant component in both groups and increased significantly from 42.60 ± 0.40% in the control to 47.40 ± 0.52% in FWCM biomass (p < 0.05). Similarly, arachidic acid (C20:0) increased from 1.67 ± 0.49% to 3.03 ± 0.15% following FWCM treatment. In contrast, monounsaturated fatty acid (MUFA) palmitoleic acid (C16:1) exhibited a remarkable enhancement, increasing nearly six-fold from 2.57 ± 0.45% in the control to 15.13 ± 0.12% in the FWCM grown biomass. However, oleic acid (C18:1) decreased significantly from 8.37 ± 0.60% to 4.60 ± 0.20%.
Notable changes were also observed in polyunsaturated fatty acids (PUFAs). Linoleic acid (C18:2) decreased from 8.63 ± 0.35% in the control to 3.97 ± 0.85% in FWCM biomass, while linolenic acid (C18:3), which was abundant in the control (18.03 ± 0.15%), was completely absent after FWCM treatment. Additionally, the emergence of eicosenoic acid (C20:1) in the FWCM biomass (2.60 ± 0.44%) indicates metabolic restructuring induced by food waste derived nutrients. Similar trends have been reported in previous studies. For instance, Nannochloropsis oceanica cultivated under mixotrophic conditions using hydrolyzed canteen food waste demonstrated a substantial increase in polyunsaturated fatty acids (PUFAs), with eicosapentaenoic acid (EPA) accounting for up to 32% of total fatty acid methyl esters (FAMEs).40 Furthermore, Almutairi et al.41 reported that pretreatment of lipid-rich food waste followed by microalgal cultivation achieved a FAME recovery efficiency of 92.6% based on total lipid content (Fig. 5).
 | ||
| Fig. 5 FAMES analysis of SA and FWCM. | ||
3.7 Total phenolic and flavonoid content, DPPH and ROS
Connan et al. (2011) reported that phenolic compounds are important constituents of microalgal cell walls and play a protective role against both biotic stresses, such as bacterial fouling and grazing, and abiotic stresses. The phenolic and flavonoid compounds in SA contribute to its antioxidant activity. Table 3 summarizes and compares the phenolic and flavonoid levels in the control culture and in algae cultivated in FWCM. The flavonoid content was calculated using the standard calibration curve of quercetin. An increase in flavonoid content was noticed in FWCM. The flavonoid content in control algae biomass was 5.57 ± 3.15 mg QE per g DW and in algae cultivated in FWCM was 11 ± 0.78 mg QE per g DW whereas the phenolic content was calculated using the standard calibration curve of gallic acid and a similar increase in phenolic content was noticed in FWCM. The control algae biomass had 5.80 ± 2.63 mg GAE per g DW phenolic content and algae cultivated in FWCM had 12.07 ± 2.63 mg GAE per g DW phenolic content. This shows that the FWCM substantially enhances the production of flavonoids and phenols in algae. FWCM contains extra nutrients and organic compounds that may induce mild stress for the algae. Such conditions often stimulate algae to produce more protective compounds like flavonoids and phenols.| Sample | TPC (mg GAE per g DW) | TFC (mg QE per g DW) | DPPH scavenging (%) | ROS scavenging (%) |
|---|---|---|---|---|
| a Data are shown as mean ± standard deviation of three replicates. Different superscript letters (a, b) within the same row indicate statistically significant differences between treatments for each parameter as determined by one-way ANOVA (p < 0.05), where values sharing the same letter are not significantly different and values with different letters are significantly different. | ||||
| Control | 5.80 ± 2.63a | 5.57 ± 3.15a | 12.45 ± 0.80a | 20.95 ± 1.80a |
| FWCM | 12.07 ± 2.63b | 11.00 ± 0.78b | 11.05 ± 0.04a | 24.59 ± 2.42b |
The impact of free radicals was assessed using DPPH antioxidant assays. In the DPPH radical scavenging test, the algal biomass exhibited a higher scavenging activity of 12.45 ± 0.80% while the culture grown in FW water exhibited slightly less scavenging activity of 11.05 ± 0.04%. Although both samples exhibited the ability to neutralize free radicals, the reduction observed in the FWCM suggests that the change in growth medium may have affected the production of certain antioxidant compounds. It is possible that while flavonoid production increased in the food-waste medium, other antioxidant metabolites that contribute to DPPH activity were either lower or substantially not enhanced. Overall, both samples showed moderate antioxidant capacity, but the control biomass showed a slight stronger free-radical-scavenging effect.
When environmental or hazardous substances causes stress in algal cells, they produce ROS like singlet oxygen, superoxide and hydroxyl radicals, which can lead to oxidative damage and even algal cell death. In the present work, SA showed a slight increase in the ROS level in terms of H2O2 from 20.95 ± 1.80% (control) to 24.59 ± 2.42% (SA + FWCM) respectively. FW affects cellular homeostasis, resulting in elevated ROS. This shift may be explained by an increase in its metabolic regime. The added FW likely increases the availability of organic carbon. This increment elevates electron flux through respiratory and photosynthetic pathways, which can increase electron leakage to oxygen and thereby generate more ROS such as H2O2.
Similar variations in antioxidant metabolites and pigment accumulation under different nutrient conditions have been reported previously. Mtaki et al.42 observed that microalgae cultivated in 5% banana stem compost medium (BCM) exhibited the highest phenolic content (8.53 ± 0.10 mg per g GAE) and antioxidant scavenging activity (97.87 ± 0.088%). These findings indicate that nutrient-rich waste-derived media can significantly influence the biosynthesis of antioxidant compounds and pigments in microalgae.
One-way ANOVA was performed individually for each parameter as mentioned in SI Tables 7–10 to evaluate the effect of treated FW sample on phytochemical parameters of algae as compared to the control. The result showed phenolics and flavonoid contents increases significantly in treated FW sample compared to the control (p < 0.05). DPPH scavenging activity also differed significantly between groups, with slightly higher activity observed in the control. However, no significant difference was observed in ROS (p < 0.05). Hence, treated samples influenced phenolic and flavonoid accumulation more strongly than antioxidant activity.
3.8 HTL of FW residues
The results show that temperature has a significant effect on product yields of food waste residues. At 250 °C, the total yield was low (9.03 ± 0.44 g). The bio-oil produced was only around (3.87 ± 0.58 g), and solid residue (5.15 ± 0.77 g). This means that at lower temperatures, the biomass does not break down completely. At 350 °C, the total yield increased to (15.88 ± 0.54 g). However, significant increase was seen only in hydrochar (solid residue) (11.76 ± 0.65 g), while the bio-oil increased slightly to (4.11 ± 0.68 g). This suggests that at higher temperatures, breakdown of biomass increased, but some of the liquid products convert to solid char instead of oil. This occurs due to secondary cracking and repolymerization reactions at high temperatures, which has been also reported by ref. 43. Firstly, very high temperatures cause the reaction intermediates to break down and join back together, which further leads to secondary reactions. These secondary reactions convert many of the liquid intermediates into hydrocarbon gases. Secondly, when the temperature reaches near the critical point of water, the properties of water change significantly, leading to production of many free radicals. Further, these free radicals promote polymerization, condensation, and ring-forming reactions. As a result, more gases and solid hydrochar are formed, while the amount of hydrochar.443.9 Pyrolysis of HTL hydrochar
The hydrochar obtained from HTL at 350 °C was further subjected to pyrolysis at 550 °C, 650 °C, and 750 °C. The effect of pyrolysis temperature on the physicochemical transformation and conversion behavior of the HTL-derived hydrochar was systematically investigated. The mass yield of the resulting biochar, from a consistent 2 g hydrochar, demonstrated that temperature had a notable effect. The yields were quantified as 0.48 ± 0.03 g, 0.45 ± 0.02 g, and 0.49 ± 0.00 g at 550 °C, 650 °C, and 750 °C, respectively, indicating relative stability in solid residue across this temperature range. | ||
| Fig. 6 XRD pattern of the biochar of FW hydrochar pyrolysis at different temperatures. | ||
 | ||
| Fig. 7 FESEM micrograph of FW biochar at 550 °C (a and b), 650 °C (c and d), and 750 °C (e and f). | ||
4 Conclusion
This study successfully demonstrates a sustainable and efficient strategy for the valorisation of FW by using it as a substitute culture medium for the cultivation of SA. The optimal culture medium, with a ratio of 80:20 (BG11:FWCM), proved highly effective, generating a significantly higher biomass yield (1.14 g L−1) compared to the control (0.75 g L−1). This enhanced growth was supported by a substantial increase in photosynthetic pigments Chl a, Chl b, and Car contents indicating superior photosynthetic health and activity in the FWCM. Furthermore, cultivation in FWCM led to a remarkable enhancement in the biochemical profile of SA. We observed a significant increase in valuable compounds, with lipid content rising to 35.93% and carbohydrate content to 17.36%. The synthesis of bioactive compounds was also increased, with flavonoid and phenolic content nearly doubling, which enhances the potential nutraceutical value of the algae. Comprehensive techno-economic analysis (TEA) and life cycle assessment (LCA) studies are required to evaluate the overall process economics, energy efficiency, environmental sustainability, and industrial scalability. Future studies should focus on large-scale optimization and integrated TEA/LCA approaches to assess the commercial applicability of the proposed biorefinery system for sustainable food industry applications.
Author contributions
Himanshu Srivastava and Shivani Dimari: methodology; data curation; writing – original draft. Bhawna Bisht: data curation; writing – review & editing. Dhruv: data curation; writing – review & editing. Rahini Parsoya: data curation; writing – review & editing. Krishna Kumar Jaiswal: data curation; writing – review & editing. Mikhail S. Vlaskin: data curation; writing – review & editing. Manisha Nanda: data curation, writing – review & editing. Sanjay Kumar: data curation, writing – review & editing. Harish Chandra Joshi: writing – review & editing, analysis. Vinod Kumar: conceptualization; supervision; writing – review & editing.Conflicts of interest
The authors declare no conflict of interest.Data availability
Data will be made available on request.Supplementary information (SI) is available. See DOI: https://doi.org/10.1039/d5fb00925a.
Acknowledgements
This paper has been supported by the grant provided by DST, Govt. of India under the project (DST/INT/RUS/RSF/P-60/2021). M. S. V. would like to thank Ministry of Science and Higher Education of the Russian Federation (State Assignment No. 075-00270-26-00). This paper has also been supported by the RUDN University Strategic Academic Leadership Program.References
- P. F. Albizzati, D. Tonini and T. F. Astrup, A quantitative sustainability assessment of food waste management in the European Union, Environ. Sci. Technol., 2021, 55(23), 16099–16109 CrossRef CAS PubMed.
- T. I. Assis and R. F. Gonçalves, Valorization of food waste by anaerobic digestion: a bibliometric and systematic review focusing on optimization, J. Environ. Manage., 2022, 320, 115763 Search PubMed.
- V. Amicarelli, G. Lagioia and C. Bux, Global warming potential of food waste through the life cycle assessment: an analytical review, Environ. Impact Assess. Rev., 2021, 91, 106677 CrossRef.
- J. Zhu, Y. Liu, Z. Luo, C. Ling, K. Yin and H. Tong, Methane mitigation strategy for food waste management: balancing socio-economic acceptance and environmental impacts, Sustain. Prod. Consum., 2023, 37, 389–397 Search PubMed.
- J. Jeevahan, A. Anderson, V. Sriram, R. B. Durairaj, G. Britto Joseph and G. Mageshwaran, Waste into energy conversion technologies and conversion of food wastes into the potential products: a review, Int. J. Ambient Energy, 2021, 42(9), 1083–1101 CrossRef.
- K. Slopiecka, F. Liberti, S. Massoli, P. Bartocci and F. Fantozzi, Chemical and physical characterization of food waste to improve its use in anaerobic digestion plants, Energy Nexus, 2022, 5, 100049 CrossRef CAS.
- P. Roy, A. K. Mohanty, P. Dick and M. Misra, A review on the challenges and choices for food waste valorization: environmental and economic impacts, ACS Environ. Au, 2023, 3(2), 58–75 CrossRef CAS PubMed.
- J. Aschemann-Witzel, D. Asioli, M. Banovic, M. A. Perito, A. O. Peschel and V. Stancu, Defining upcycled food: The dual role of upcycling in reducing food loss and waste, Trends Food Sci. Technol., 2023, 132, 132–137 CrossRef CAS.
- P. Murugesan, V. Raja, S. Dutta, J. A. Moses and C. Anandharamakrishnan, Food waste valorisation via gasification–a review on emerging concepts, prospects and challenges, Sci. Total Environ., 2022, 851, 157955 CrossRef CAS PubMed.
- E. O. Ebikade, S. Sadula, Y. Gupta and D. G. Vlachos, A review of thermal and thermocatalytic valorization of food waste, Green Chem., 2021, 23(8), 2806–2833 RSC.
- M. S. Mia, M. M. Ahmed and W. Zzaman, Valorization of food waste into functional ingredients supports a sustainable strategy for the food industry, Discover Food, 2025, 5(1), 275 CrossRef.
- Z. Liu, T. S. de Souza, B. Holland, F. Dunshea, C. Barrow and H. A. Suleria, Valorization of food waste to produce value-added products based on its bioactive compounds, Processes, 2023, 11(3), 840 CrossRef CAS.
- A. Talan, B. Tiwari, B. Yadav, R. D. Tyagi, J. W. C. Wong and P. Drogui, Food waste valorization: energy production using novel integrated systems, Bioresour. Technol., 2021, 322, 124538 CrossRef CAS PubMed.
- V. Raja, S. Dutta, P. Murugesan, J. A. Moses and C. Anandharamakrishnan, Electricity production using food waste: a review, Environ. Chem. Lett., 2023, 21(2), 839–864 CrossRef CAS.
- B. Motavaf and P. E. Savage, Effect of process variables on food waste valorization via hydrothermal liquefaction, ACS ES&T Eng., 2021, 1(3), 363–374 Search PubMed.
- H. Bayat, M. Dehghanizadeh, J. M. Jarvis, C. E. Brewer and U. Jena, Hydrothermal liquefaction of food waste: effect of process parameters on product yields and chemistry, Front. Sustain. Food Syst., 2021, 5, 658592 CrossRef.
- A. Aierzhati, M. J. Stablein, N. E. Wu, C. T. Kuo, B. Si, X. Kang and Y. Zhang, Experimental and model enhancement of food waste hydrothermal liquefaction with combined effects of biochemical composition and reaction conditions, Bioresour. Technol., 2019, 284, 139–147 CrossRef CAS PubMed.
- R. Saengsuriwong, T. Onsree, S. Phromphithak and N. Tippayawong, Biocrude oil production via hydrothermal liquefaction of food waste in a simplified high-throughput reactor, Bioresour. Technol., 2021, 341, 125750 CrossRef CAS PubMed.
- S. Dhiman, B. Thakur, S. Kaur, M. Ahuja, S. Gantayat, S. Sarkar and M. Tripathi, Closing the loop: technological innovations in food waste valorisation for global sustainability, Discov. Sustain., 2025, 6(1), 1–35 Search PubMed.
- Association of Official Analytical Chemists, Official Methods of Analysis of the Association of Official Analytical Chemists, The Association, 2000, vol. 11 Search PubMed.
- N. Arora, A. Patel, K. Sartaj, P. A. Pruthi and V. Pruthi, Bioremediation of domestic and industrial wastewaters integrated with enhanced biodiesel production using novel oleaginous microalgae, Environ. Sci. Pollut. Res., 2016, 23(20), 20997–21007 CrossRef CAS PubMed.
- APHA, in Standard Methods for the Examination of Water and Wastewater, ed. E. W. Rice, R. B. Baird, A. D. Eaton and L. S. Clesceri, American Public Health Association (APHA), American Water Works Association (AWWA) and Water Environment Federation (WEF), Washington, D.C., USA, 22nd edn, 2012 Search PubMed.
- A. Gnanavelu, T. S. Shanmuganathan, V. Deepesh and S. Suresh, Validation of a Modified Procedure for the Determination of Chemical Oxygen Demand using Standard Dichromate Method in Industrial Wastewater Samples with High Calcium Chloride Content, Indian J. Sci. Technol., 2021, 14(29), 2391–2399 CrossRef CAS.
- H. K. Lichtenthaler, [34] Chlorophylls and carotenoids: pigments of photosynthetic biomembranes, Methods Enzymol., 1987, 148, 350–382 CAS.
- M. DuBois, K. A. Gilles, J. K. Hamilton, P. T. Rebers and F. Smith, Colorimetric method for determination of sugars and related substances, Anal. Chem., 1956, 28(3), 350–356 CrossRef CAS.
- N. Arora, K. Gulati, A. Patel, P. A. Pruthi, K. M. Poluri and V. Pruthi, A hybrid approach integrating arsenic detoxification with biodiesel production using oleaginous microalgae, Algal Res., 2017, 24, 29–39 Search PubMed.
- E. G. Bligh and W. J. Dyer, A rapid method of total lipid extraction and purification, Can. J. Biochem. Physiol., 1959, 37(8), 911–917 Search PubMed.
- V. L. Singleton, R. Orthofer and R. M. Lamuela-Raventós, [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent, Methods Enzymol., 1999, 299, 152–178 CAS.
- B. Bisht, U. C. Lohani, M. Nanda, S. Kumar, M. Vlaskin, A. Dsouza and V. Kumar, Synergistic interaction between hydrocolloids and kinnow peel biowaste for the development of edible fillers using response surface methodology, Environ. Sci. Pollut. Res., 2024, 31(2), 2595–2613 CrossRef CAS PubMed.
- S. Q. Aziz, H. A. Aziz, M. S. Yusoff, M. J. Bashir and M. Umar, Leachate characterization in semi-aerobic and anaerobic sanitary landfills: a comparative study, J. Environ. Manag., 2010, 91(12), 2608–2614 CrossRef CAS PubMed.
- I. Kontodimos, C. Ketikidis and P. Grammelis, Valorization of food waste leachates through anaerobic digestion, Eng. Proc., 2022, 31(1), 25 Search PubMed.
- A. A. Ramandani, Y. M. Sun, J. C. W. Lan, J. W. Lim, J. S. Chang, S. Srinuanpan and K. S. Khoo, Upcycling food waste as a low-cost cultivation medium for Chlorella sp. microalgae, J. Sci. Food Agric., 2025, 105(9), 4715–4727 CrossRef CAS PubMed.
- L. Dao, J. Beardall and P. Heraud, Characterisation of Pb-induced changes and prediction of Pb exposure in microalgae using infrared spectroscopy, Aquat. Toxicol., 2017, 188, 33–42 CrossRef CAS PubMed.
- X. Li, X. Zhang, Y. Zhao and X. Yu, Cross-talk between gama-aminobutyric acid and calcium ion regulates lipid biosynthesis in Monoraphidium sp. QLY-1 in response to combined treatment of fulvic acid and salinity stress, Bioresour. Technol., 2020, 315, 123833 CrossRef CAS PubMed.
- A. Giwa, B. Chalermthai, N. Moheimani and H. Taher, Effective nutrient removal and metabolite accumulation by C. vulgaris cultivated using digested food waste and brine, Environ. Technol. Innovation, 2021, 24, 101935 CrossRef CAS.
- K. W. Chew, S. R. Chia, P. L. Show, T. C. Ling, S. S. Arya and J. S. Chang, Food waste compost as an organic nutrient source for the cultivation of Chlorella vulgaris, Bioresour. Technol., 2018, 267, 356–362 CrossRef CAS PubMed.
- L. Zhang, J. Cheng, H. Pei, J. Pan, L. Jiang, Q. Hou and F. Han, Cultivation of microalgae using anaerobically digested effluent from kitchen waste as a nutrient source for biodiesel production, Renew. Energy, 2018, 115, 276–287 CrossRef CAS.
- T. S. Lin and J. Y. Wu, Effect of carbon sources on growth and lipid accumulation of newly isolated microalgae cultured under mixotrophic condition, Bioresour. Technol., 2015, 184, 100–107 CrossRef CAS PubMed.
- Y. Zeng, T. Xie, P. Li, B. Jian, X. Li, Y. Xie and Y. Zhang, Enhanced lipid production and nutrient utilization of food waste hydrolysate by mixed culture of oleaginous yeast Rhodosporidium toruloides and oleaginous microalgae Chlorella vulgaris, Renew. Energy, 2018, 126, 915–923 CrossRef CAS.
- A. Pugazhendhi and A. Sharma, Food waste upcycling via microalgal adaptive evolution for lipids and fatty acids enhancement towards circular bioeconomy, Chem. Eng. J., 2024, 484, 149593 CrossRef CAS.
- A. W. Almutairi, Z. M. Al-Hasawi and A. E. F. Abomohra, Valorization of lipidic food waste for enhanced biodiesel recovery through two-step conversion: a novel microalgae-integrated approach, Bioresour. Technol., 2021, 342, 125966 CrossRef CAS PubMed.
- K. Mtaki, M. S. Kyewalyanga and M. S. Mtolera, Assessment of antioxidant contents and free radical-scavenging capacity of chlorella vulgaris cultivated in low cost media, Appl. Sci., 2020, 10(23), 8611 CrossRef CAS.
- J. Cheikhwafa, K. Glińska, E. Torrens and C. Bengoa, Effect of temperature on hydrothermal liquefaction of high lipids and carbohydrates content municipal primary sludge, Heliyon, 2024, 10(3), e24731 CrossRef CAS PubMed.
- B. Bisht, P. Gururani, S. Pandey, K. K. Jaiswal, S. Kumar, M. S. Vlaskin, M. Verma, H. Kim and V. Kumar, Multi-stage hydrothermal liquefaction modeling of sludge and microalgae biomass to increase bio-oil yield, Fuel, 2022, 328, 125253 CrossRef CAS.
- K. K. Jaiswal, V. Kumar, R. Verma, M. Verma, A. Kumar, M. S. Vlaskin and H. Kim, Graphitic bio-char and bio-oil synthesis via hydrothermal carbonization-co-liquefaction of microalgae biomass (oiled/de-oiled) and multiple heavy metals remediations, J. Hazard. Mater., 2021, 409, 124987 Search PubMed.
- S. M. Chistie, S. U. Naik, P. Rajendra, Apeksha, R. K. Mishra, G. Albasher and J. Hameed, Production and characterization of magnetic Biochar derived from pyrolysis of waste areca nut husk for removal of methylene blue dye from wastewater, Sci. Rep., 2025, 15(1), 23209 CrossRef CAS PubMed.
- C. Ma, Y. Zhao, H. Chen, Y. Liu, R. Huang and J. Pan, Biochars derived from by-products of microalgae pyrolysis for sorption of gaseous H2S, J. Environ. Chem. Eng., 2022, 10(3), 107370 CrossRef CAS.
- M. Makowska and K. Dziosa, Influence of different pyrolysis temperatures on chemical composition and graphite-like structure of biochar produced from biomass of green microalgae Chlorella sp, Environ. Technol. Innovation, 2024, 35, 103667 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2026 |