
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceEmerging porous materials for cell encapsulation
Francesco Carraro†
 a,
Miriam de J. Velásquez-Hernández†‡
a,
Anita Emmerstorfer-Augustinb,
Daniel Kracherb,
Qilu Wuc,
Robert Kouristb,
Lien-Yang Choud,
Ju Gec,
Fa-Kuen Shiehe,
Christian Doonanf and
Paolo Falcaro*a
a,
Miriam de J. Velásquez-Hernández†‡
a,
Anita Emmerstorfer-Augustinb,
Daniel Kracherb,
Qilu Wuc,
Robert Kouristb,
Lien-Yang Choud,
Ju Gec,
Fa-Kuen Shiehe,
Christian Doonanf and
Paolo Falcaro*a
aInstitute of Physical and Theoretical Chemistry, Graz University of Technology, Stremayrgasse 9, Graz, 8010, Austria. E-mail: paolo.falcaro@tugaz.at
bInstitute of Molecular Biotechnology, Graz University of Technology, Petersgasse 14, Graz, 8010, Austria
cKey Lab of Industrial Biocatalysis, Ministry of Education, Department of Chemical Engineering, Tsinghua University, Beijing 100084, China
dSchool of Physical Science and Technology, ShanghaiTech University, Shanghai, 201210, P.R. China
eDepartment of Chemistry, National Central University, Taoyuan, 32001, Taiwan
fDepartment of Chemistry and Centre for Advanced Nanomaterials, University of Adelaide, Adelaide, SA 5005, Australia
First published on 13th May 2026
Abstract
This review summarizes recent progress in cell@MOF, cell@COF, and cell@HOF composites from a synthetic biology and materials science perspective. It outlines key synthetic strategies for the synthesis of porous abiotic exoskeletons, focusing on framework-based materials. Additionally, it discusses the cell surface chemistry and current methods for assessing cell viability. Major applications, including cell therapy, biocatalysis, biosensing, and CO2 mitigation, are examined alongside approaches for composite preparation and characterization. This review concludes with prospects and challenges for using framework materials to engineer synthetic cells and enhance cellular functions.
 Francesco Carraro | Francesco Carraro received his PhD in Science and Engineering of Materials and Nanostructures from the University of Padova (Italy) in 2018. After postdoctoral work in the Falcaro group at the Graz University of Technology (TU Graz, Austria) on metal–organic framework films and biomolecule encapsulation, he became Assistant Professor at TU Graz in 2022, focusing on the crystal engineering and biotechnological applications of metal–organic and hydrogen-bonded organic framework biocomposites. |
 Miriam de J. Velásquez-Hernández | Miriam de J. Velásquez-Hernández received her PhD degree from the Universidad Nacional Autónoma de México (UNAM). She subsequently joined Paolo Falcaro's group at the Graz University of Technology (TU Graz, Austria) as a CONACyT postdoctoral fellow, focusing on the synthesis of metal–organic framework based biocomposites for drug delivery applications. She is currently an MSCA postdoctoral fellow in Rob Ameloot's group at KU Leuven. |
Introduction
In nature, the survival of biological species relies on their ability to cope with diverse environmental conditions. To adapt to environmental stress, some microorganisms have developed the ability to construct abiotic coatings via the self-assembly of molecular precursors.1,2 These coatings are purposely designed to confer various functional properties to the enclosed cells, such as mechanical strength, thermal resistance, and physical protection, that ensure cell survival under inhospitable conditions.1–3 For example, some bacteria produce an organic polymeric network that provides mechanical robustness to withstand external pressures and enhances their desiccation tolerance in water-deficient environments.4 This survival mechanism is called sporulation.5 Specifically, the cell divides asymmetrically, forming a new compartment: the forespore.6–8 In the forespore, the cell encapsulates precious biological materials for the strain's survival (e.g., deoxyribonucleic acid (DNA), ribonucleic acid (RNA), and enzymes). Then, the forespore undergoes a maturation process in which a cortex and a multi-layer coating surround the forespore's core, forming a protective shield (Fig. 1a). Subsequently, metabolic inactivation or dormancy is achieved by gradually dehydrating the core by replacing water with dipicolinic acid (DPA) and calcium ions, leading to endospore formation within the mother cell (Fig. 1a). Finally, the mother cell undergoes programmed cell death (apoptosis§), accompanied by release of the endospore into the environment (i.e., spore). The secreted spore can remain dormant until it senses that harsh conditions are alleviated (Fig. 1a). Details of this process can be found in dedicated review papers.6–8 The natural formation of protective coating materials is not restricted to organic biopolymers. Some cells possess a metabolic machinery to assimilate minerals from the environment and synthesize an inorganic material-based coating.1,2 This process is called biomineralization and yields rigid and protective exoskeletons for different biological systems.1,2 Biosilicification is the archetype of individual cell biomineralization, wherein diatoms convert soluble silicic acid into an amorphous silica shell.1,2 This inorganic coating enhances cell viability when exposed to environmental stresses, such as heat, desiccation, microbial attack, and lytic enzymatic degradation. Simultaneously, the inorganic coating affords optical transparency and enables transport of nutrients necessary for cellular functions. Other typical examples of inorganic cell coatings are calcium carbonate, which can be found in mollusk shells and pearls,9 and calcium phosphate that is present in bone tissue.10 Inspired by these natural coatings, multidisciplinary research investigates synthetic methods for the development of abiotic protective exoskeletons on cells that lack natural biomineralization capability. These synthetic cell@shell systems are known as “artificial spores” (Fig. 1b)11 and can possess three main advantages: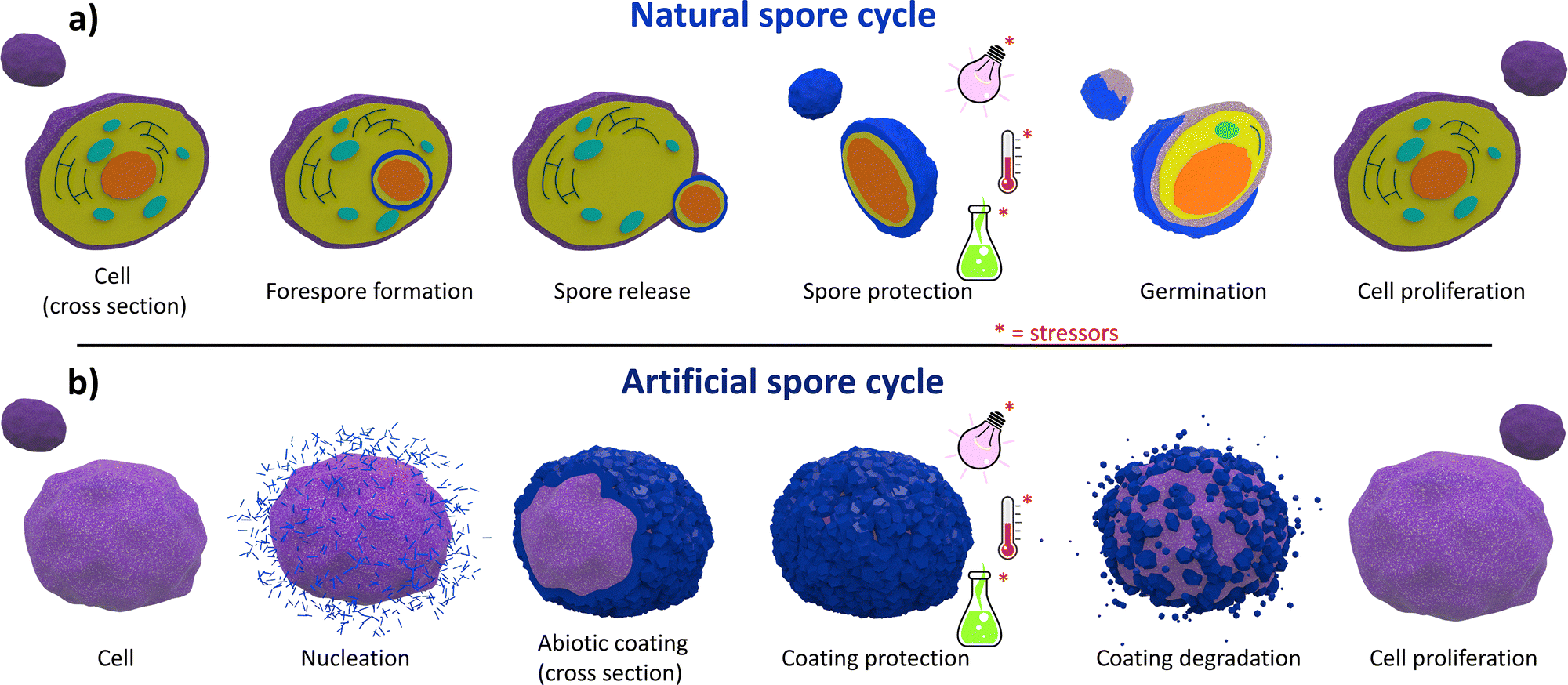 | ||
| Fig. 1 Schematic of the natural (a) and artificial (b) sporulation. The natural process begins with asymmetric cell division: the mother cell (purple) forms a forespore (orange). The mother cell engulfs the forespore, leading to a mature spore (blue) that is resistant to multiple stressors (UV, chemicals, and heat). The spore can remain dormant until nutrient-rich conditions trigger germination and cell growth. Artificial sporulation similarly employs protective coatings (blue crystals) around individual cells (purple), suppressing proliferation and enhancing resistance. On-demand dissolution of this abiotic coating then restores normal growth and metabolism. | ||
I. Enhanced cell resistance to chemical and physical stressors: unlike naked cells, artificial spores exhibit enhanced tolerance to unfavourable environmental conditions such as enzymatic degradation,12 changes in the osmotic pressure,13 high temperatures,14 and UV radiation (Fig. 1b);15
II. On-demand suppression and reactivation of cell division: the formation of rigid artificial shells around living cells hinders cell division, inducing a state of dormancy akin to spores. The cell proliferation and natural metabolic functions can be restored by on-demand shell degradation (Fig. 1b);11
III. Tailored exogenous biochemical properties: by designing coatings with specific chemical and biochemical properties, cells can be engineered with abiotic exoskeletons with exogenous chemical functionalities: the new cell@shell systems possess functionalities that are not present in the original naked cells. For example, this material design strategy has enhanced cell adaptability to nutrient-deficient and protease-rich environments.12,16
We note that abiotic exoskeletons should satisfy requirements such as (i) perm-selectivity, (ii) durability, (iii) degradability on demand, and (iv) functionalizability.11,17,18 These properties are described as follows:
I. Perm-selectivity: preserving cell viability in a cell@shell system depends on the continuous supply of nutrients to the cytosol. Ideally, artificial cell coatings should act as a molecular sieve allowing the free transport of biologically relevant molecules (e.g. see Video S1), such as cell nutrients, oxygen, and metabolites, while preventing the diffusion of cytotoxic macromolecules.11,17,18
II. Durability: emulating spore-like features requires the fabrication of artificial coatings sufficiently robust to withstand the mechanical stress caused by changes in the osmotic pressure and dehydration. Additionally, rigid artificial shells retard or suppress cell division, mimicking the spore-like dormancy.11,17,18
III. Degradability on demand: achieving programmable recovery of original metabolic cell functions, the artificial shell must be able to degrade upon applying external stimuli. The on-demand removal of the artificial coating grants control over the dormant and active state transition.19
IV. Functionalizability: imparting exogenous functional properties artificially enables the fabrication of systems with non-natural functions. This approach can be used to adapt cells to hostile habitats (e.g., nutrient-deficient or cytotoxic environments).12,16
Inspired by the potential to design distinctive functional properties, researchers have focused on coating living cells with various inorganic (e.g., SiO2, CaCO3, and MnO2), organic (e.g., alginate, polyethylene, chitosan, and cell membranes), and hybrid (e.g. metal-phenolic networks and framework materials) materials (Fig. 2a–c).9,18,20–30 In this review, we focus on framework materials for encasing individual living cells (Fig. 2c).31 A framework material can be defined as an extended crystalline network comprised of molecular building blocks interconnected via directional bonding interactions.32 The most common framework materials employed to encapsulate living cells are metal–organic frameworks (MOFs). Recently, the encapsulation of living cells within covalent organic frameworks (COFs) and hydrogen-bonded organic frameworks (HOFs) has been reported.31,33,34 All three framework materials are assembled via bottom-up synthetic approaches. MOFs consist of inorganic clusters linked together via multitopic organic linkers,35,36 whereas COFs and HOFs are constructed exclusively from organic building blocks interconnected through covalent and hydrogen-bonding interactions, respectively.37,38 A common feature of these materials is their bottom-up synthesis, which allows for the chemical and structural properties of the abiotic coatings to be precisely tailored.32,37 For example, pore sizes can be adjusted to modulate the diffusion of essential cell nutrients while preventing the cytotoxic effects of proteolytic agents.31
 | ||
| Fig. 2 Schematic representation (top) of the inorganic (a), organic (b), and hybrid materials (c) used as abiotic cell coatings, together with SEM (middle) and TEM (bottom) micrographs of selected examples. (a) SEM micrographs of yeast@SiO2 at different magnifications. The TEM images of microtome-sliced yeast@SiO2 indicate silica shells with a thickness above 50 nm (adapted with permission from ref. 22 Copyright 2009, Wiley-VCH). (b) SEM and TEM micrographs of organic polymeric materials for multilayer cell coatings (adapted with permission from ref. 39). (c) Illustration of single cells encapsulated within a metal–polyphenol nanoshell (adapted with permission from ref. 19 Copyright 2014, Wiley-VCH). | ||
In this review, we provide a general overview of the emerging research on cell@MOF, cell@COF, and cell@HOF composites from a synthetic biology and materials science perspective. First, we discuss the principles behind the two synthetic strategies used to grow framework materials for abiotic exoskeletons (i.e., one-pot and multi-step processes), the cell surface chemistry, and the best practices for evaluating the viability of the coated cells. Next, we explore the applications of cell@MOF, cell@COF, and cell@HOF composites, including cell adaptability,11,17 cell therapy,21,33 biocatalysis,39 biosensing,40 and CO2 mitigation,41 with a focus on the preparation and characterization of these biocomposites. Then, we discuss the potential of MOF-, COF-, and HOF-based abiotic coatings for the fabrication of synthetic cells. Finally, we provide brief insights into the future opportunities and challenges of using framework materials as exoskeletons to enhance cell functionality, with specific attention to Escherichia coli (E. coli), or genetically engineered bacteria for targeted therapeutic delivery. Such microorganisms can be encapsulated within MOF materials to enhance their therapeutic performance against cancer. This approach aims to develop a diverse range of bacteria@MOF biocomposites and explore their potential applications in cancer immunotherapy, specifically through bacterial-mediated cancer therapy (BMCT).42,43
Coating approaches for living cells
Two main synthetic approaches have been used for the fabrication of cell@MOF, cell@COF, and cell@HOF composites: (i) the one-pot cell coating (Fig. 3a) and (ii) the multi-step cell cytoprotective encapsulation strategy (Fig. 3b). In this section, we introduce aspects that are at the foundation of these two synthetic approaches and underpin all the examples of artificial spores discussed in this review.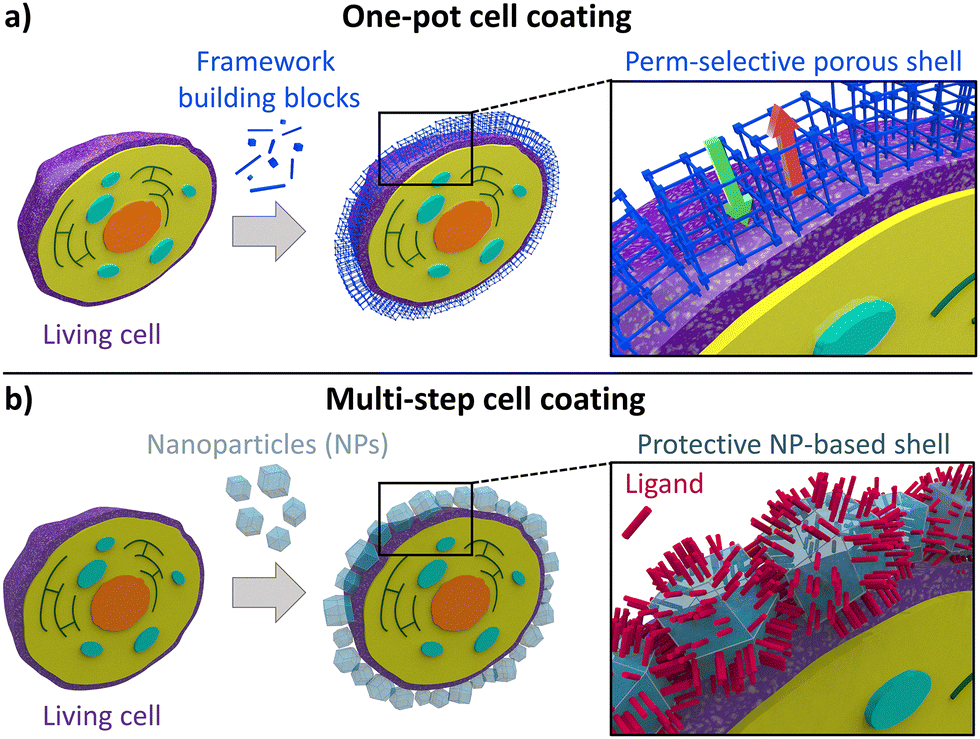 | ||
| Fig. 3 Schematic representation of (a) the one-pot encapsulation process and (b) the “SupraCell64 structure” formed by depositing pre-formed abiotic nanoparticles onto the cell membrane. | ||
One-pot encapsulation strategy
The one-pot approach requires biological entities regulating the on-site formation of abiotic coatings (Fig. 3a), mimicking natural biomineralization processes. Such a one-pot cell coating method is carried out by mixing a cell suspension with an aqueous solution of the MOF, COF, or HOF precursors. Similar to how biological materials can trigger natural biomineralization, leading to the deposition of inorganic nanoparticles on their surfaces, living organisms can induce the self-assembly of MOF, COF, or HOF materials on their surfaces.31,33,34,44 At the bio-interface, two main driving forces promote the growth of the porous coatings: (i) the free energy of nucleation and (ii) the electrostatic intermolecular interactions. The first can be explained by a heterogeneous nucleation effect induced by cells: as observed for inorganic nanoparticles,45,46 these nucleation seeds lower the energy barrier for the nucleation of extended network materials. According to the classical crystal nucleation theory, the free energy associated with the formation of a spherical crystal has a negative bulk contribution and a positive surface contribution, the latter opposing the crystal growth.47 The presence of cells provides the system with a scaffold for the crystal growth, thus reducing the need to create a new surface and lowering the crystallization energy barrier.48 Although classical nucleation theory refers to crystalline materials, experimental evidence in biomineralization, especially in biological and biomimetic systems, shows that crystal formation may pass through an amorphous phase that serves as a precursor to the subsequent crystalline structure.49 The second driving force arises from electrostatic interactions between the cell surface and the MOF/COF/HOF precursors. In this context, the isoelectric point (i.e., the pH at which the cell carries no net electrical charge50) and the synthesis environment (e.g., pH, ionic strength) together determine the net cell surface charge during synthesis, which is commonly described by the zeta potential (ζ-potential).50 A large positive or negative value of the cell ζ-potential (i.e., >+ 30 mV or <−30 mV) will therefore indicate that the cell carries a large positive or negative net charge in the synthetic environment.51 Similarly, the net charge of the MOF/COF/HOF precursors in the synthetic environment is determined by their chemical nature (e.g., charge of the metal cations and pKa of an organic ligand). For MOFs, the role of electrostatic interactions between biomacromolecules at the cell surface and framework precursors in porous framework nucleation and growth has been extensively discussed based on ex situ observations, whereas for HOFs and COFs the corresponding mechanistic details require further understanding. In fact, research on MOF biomimetic mineralization and biomacromolecule encapsulation has revealed that the presence of surface-exposed charged functional groups reduces the ζ-potential, thus inducing local supersaturation of oppositely charged precursors and ultimately triggering MOF nucleation.34,52–54 Specific cell membrane components, such as negatively charged glycoproteins, peptidoglycans, and carbohydrates, promote electrostatic interaction between the cell and the MOF metal precursors.34,52–55 These interactions are recognized as the main variable driving the rapid formation of amorphous MOF layers on biomacromolecules as well as on the outer surfaces of cells and viruses.34,52–54 This amorphous phase can subsequently transform into crystalline materials.16,56It is important to note that while electrostatic interactions and heterogeneous nucleation are widely suggested to be the primary driving forces for the framework shell formation, direct in situ experimental data elucidating the kinetics of framework growth on living cell surfaces are currently lacking. Nevertheless, there are observations supporting the assumption that mechanistic insights derived from protein systems may be extended to cells. In particular, the ability of charged proteins to promote the framework nucleation has been reported as a size-independent phenomenon, consistent with a mechanism governed primarily by interfacial charge density and local coordination chemistry rather than by the dimensions of the biomolecule used as the nucleation seed.52–54,57 Conceptually, this mechanism is consistent with nucleation strategies developed for synthetic surfaces, on which charged, self-assembled layers are engineered to control the growth of framework-based films; for example, in layer-by-layer (LbL) approaches, self-assembled monolayers provide carboxylate interfaces that enable the formation of a homogeneous MOF film.58,59 Systems that are conceptually closer to cellular interfaces are protein and fatty acid films: these have been shown to act as effective nucleating systems for different MOF coatings.60,61 These observations collectively support the notion that charged biomacromolecules, either as individual entities or as densely packed biomolecular interfaces, can induce framework nucleation and growth. However, the direct experimental validation that these molecular-level models accurately describe the formation of abiotic shells on living interfaces remains a significant challenge, primarily due to dimensional constraints. For instance, while time-resolved small angle X-ray scattering (SAXS) is an advanced technique to investigate the nucleation and early growth of frameworks around proteins, the micrometric size of cells falls outside the observable scattering range of conventional SAXS setups.54,57 Consequently, the development of adaptable in situ characterization techniques capable of bridging the length scale between molecular nucleation and cellular dimensions is urgently needed to move beyond extrapolated models and fully clarify the mechanistic details of cell encapsulation.
When the target cell exhibits a low surface charge density, the spontaneous and rapid formation of a continuous coating is more challenging. This limitation can be overcome by electrostatically adsorbing a charged capping agent onto the cell surface (e.g., PDADMAC: poly(dimethyl diallyl ammonium chloride) (+)/PAA: polyacrylic acid (−)),62 to synthetically modify the ζ-potential of the cell surface. The abundance of charged groups on the precoated cells, in the presence of the MOF precursors, accelerates the deposition of the abiotic shell around the cell surface.62
We note that rapid formation of the cell coating is often a crucial condition in maximizing cell viability during encapsulation processes: upon the rapid shell formation, it is possible to minimize cell exposure to metal cations, toxic linkers, and non-physiological pH conditions. In contrast, unoptimized coating protocols that prolong cell exposure to such non-physiological environments typically result in cell damage and loss of viability.23,63
An additional limitation of the one-pot encapsulation strategy is its limited capability to produce shells with controlled thickness and homogeneity across different cell types. For example, the reported cells and zeolitic imidazolate framework-8 (ZIF-8) composites (cell@ZIF-8) show different ZIF shell thicknesses. In general, customized protocols are needed for specific cell batches; this includes optimization depending on cell types, cell density, and the specific media used.
Multi-step encapsulation strategy
Brinker et al.64 have developed a versatile nanoparticle-based cell coating technique, which can be applied to a variety of abiotic materials, including ceramics (e.g., SiO2 and Fe3O4) and MOFs (e.g., MIL-100 and ZIF-8). This approach relies on the electrostatic interactions between proteins on the surface of cells and the nanoparticles (NPs). Typically, NP–cell interactions lead to the accumulation of NPs around the living cell, followed by particle internalization through phagocytosis or micropinocytosis processes (Fig. 3b).64 Such interfacial interactions between NPs and cells suggest that NPs could be used for cellular encapsulation if the internalization mechanisms of NPs are suppressed. Internalization pathways can be suppressed by promoting inter-particle binding through supramolecular interactions. Therefore, unlike the biomimetic mineralization approach, in the multi-step strategy, the MOF precursors are premixed in the absence of the targeted cell, yielding a colloidal solution of MOF NPs. Then, the cells are added and incubated in a colloidal suspension of pre-synthesized MOF NPs with the addition of tannic acid (an additive that promotes interparticle binding). Non-covalent interactions between the cellular membrane and NPs (e.g., metal–phosphate) promote the accumulation of those particles around the cell wall. Tannic acid enables a strong multivalent metal–phenolic complexation process, causing interparticle binding. This process yields a continuous coating of NPs around individual cells. By engineering the process of NP adsorption onto cells and selecting the appropriate additive for interparticle binding, internalization of MOF nanoparticles can be successfully inhibited, enabling the formation of a protective abiotic coating based on MOF NPs.We have thus far described the fundamental criteria and general approaches to cell coating. In the following sections, we will further explore how different cell surface properties govern the affinity between cells and abiotic coatings.
Type of cells
All living cells encode their genetic information with DNA and are encased by a semipermeable lipid bilayer – termed the cytosolic membrane in prokaryotes and the plasma membrane in eukaryotes. Prokaryotes and eukaryotes differ in several fundamental aspects. For example, eukaryotic cells have a nucleus in which the DNA is separated from the cytoplasm, whereas prokaryotic cells lack a nuclear envelope. Prokaryotic cells are typically smaller and simpler than eukaryotic cells (e.g., prokaryotic cells do not contain cytoplasmic organelles or a sophisticated cytoskeleton), and their genomes are smaller and less complex.Cells can further be classified depending on whether a cell can exist in its single-celled form, like microorganisms, or only in tissues of multicellular organisms, like most higher eukaryotes. Microorganisms, including bacteria, archaea, protozoa, algae, or fungi, produce cell walls in addition to their cell membranes. Cell walls serve as outer protective layers for many microorganisms and some multicellular organisms (e.g., plant cells). Beyond its protective role, the cell wall – composed of diverse biopolymers – offers structural cohesion. The specific composition and architecture of the cell wall vary between species and will be discussed in more detail below. Regarding synthetic encapsulation methods, these outermost cell components act as the primary interface for the MOF, HOF, or COF precursor/particle crystallization/accumulation, guiding the formation of the external abiotic layer at the bio-interface.
Bacteria and their cell surface
Bacteria are single-celled prokaryotic microorganisms. Depending on the composition and structure of their membranes and cell walls, bacteria are differentiated between Gram-negative and Gram-positive bacteria.65Gram-negative bacteria are surrounded by two membrane bilayers, the inner (cytoplasmic) and the outer membrane, separated by a space termed the periplasm. The periplasm consists of a thin peptidoglycan layer and provides a distinct reducing environment, which allows more efficient and diverse mechanisms of protein oxidation, folding, and quality control.66 The outer membrane of Gram-negative bacteria is an asymmetric bilayer with an inner leaflet consisting of phospholipids and an outer leaflet consisting of lipopolysaccharides (LPSs).67 Since LPSs are partially phosphorylated, the phosphate group confers a net negative charge.68 The most prominent example of a Gram-negative bacterium is Escherichia coli (E. coli). E. coli colonizes the human and mammalian intestinal tract, and it is used as a model organism of choice when it comes to DNA cloning and expression of recombinant genes in the field of molecular biology and biotechnology.69 Core advantages of this bacterium are simple and cheap cultivation conditions, fast growth, and well-established genetic engineering tools. For these reasons, E. coli is frequently chosen as the benchmark for novel technologies and was also among the first bacteria used in MOF encapsulation experiments.70 Another relevant example of Gram-negative bacterium is Pseudomonas putida (P. putida), a solvent-tolerant bacterium that can be used as a biocatalyst in two-phase fermentation systems for the synthesis of fine chemicals.71 Both examples show negatively charged surfaces under neutral or slightly acidic conditions. For example, different E. coli strains exhibit varying ζ-potentials ranging from −4.9 to −33.9 mV in 150 mM phosphate-buffered saline (PBS) buffer at pH 7.4.72 For P. putida, ζ-potentials were found to be typically close to −30 mV under slightly acidic conditions (e.g., −27.4 mV in 1 mM NaCl);73 −30 mV in 10 mM KNO3 at pH 6.2.71
In contrast to Gram-negative bacteria, the cell wall of Gram-positive bacteria consists of a thick peptidoglycan layer that surrounds the cytoplasmic membrane, and the peptidoglycan is decorated with teichoic acids, polysaccharides, and proteins.74 Since teichoic acid is negatively charged, the cell surfaces of Gram-positive bacteria also exhibit a negative charge.75 Examples of Gram-positive bacteria are Lactobacillus acidophilus (L. acidophilus), Staphylococcus aureus (S. aureus), and Moorella thermoacetica (M. thermoacetica). L. acidophilus CRL 640, a Gram-positive bacterium, exhibits a ζ-potential of approximately −45 mV.76 A direct comparison of the ζ-potential of E. coli and S. aureus was given, for example, by Oh et al., and it was calculated to be −37.1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) mV and −12.7mV, respectively.77
mV and −12.7mV, respectively.77
In conclusion, we note that despite the differences in structure and composition of the cell walls of Gram-negative and Gram-positive bacteria, bacteria typically display a negative surface charge under physiological conditions.78
Eukaryotes and their cell surface
Eukaryotic cells display a diverse range of surface structures, which serve critical roles in maintaining cell integrity, regulating interactions with the environment, and mediating biochemical processes. This section will explore the distinct characteristics of fungal, mammalian, and plant cell surfaces, illustrating their varied compositions and functions.Fungal cells have cell walls made up of glucans, chitin, and glycoproteins. In most fungal species, cell walls are layered. The innermost layer typically consists of a core of covalently attached, branched (1,3)-β glucan with 3 to 4% interchain and chitin, and these components assemble into fibrous microfibrils, which provide the cell with the strength required to withstand the substantial internal pressure exerted by the cytoplasm and membrane.79 The outer layers of the wall tend to be more heterogeneous and tailored to the physiology of particular fungi. In Saccharomyces cerevisiae (S. cerevisiae), (1,3)-β glucan and (1,6)-β glucan are linked to mannoprotein in the outermost parts of the cell wall, which is thought to control porosity and thus mass transfer across the cell wall.80 The yeast surface is charged negatively due to the presence of phosphates in mannoproteins.81 ζ-Potential values of the S. cerevisiae cell surface depend on the growth phase and on aerobic or anaerobic cultivation conditions: ζ-potentials dropped at later growth phases (from −10 to −20 mV) and also under anaerobic conditions (from −18 to −26 mV).82 Single-cell measurements revealed a correlation between the presence of dead cells and reduced ζ-potentials, likely resulting from cell wall damage. Additionally, S. cerevisiae was observed to release significant amounts of acids into the culture supernatant, which further contributed to the decrease in ζ-potentials. Similar findings were reported by Rogowska et al.83 While the ζ-potential value decreased from −3 to −18 mV in the 2–6 pH interval, values ranging from −19 to −20 mV were measured for pH > 7.83
Plant cells have walls that are arranged in layers and contain cellulose microfibrils, hemicellulose, pectin, lignin, and soluble protein, whereas the exact composition strongly depends on the cell type.84 In general, these components are organized into three major layers: the innermost secondary cell wall (formed only in specialized, differentiated plant cells), the primary cell wall, and the outermost middle lamella. The secondary cell wall is built from three layers, typically referred to as S1, S2, and S3, and mostly contains cellulose, hemicellulose, and lignin. The primary cell wall is the thickest layer and contains similar components, but more pectin, than the secondary cell wall. The middle lamella is mainly composed of pectic polysaccharides, lignin, and a small amount of proteins and serves as a cementing layer between the primary walls of adjacent cells.85 For research purposes, so-called protoplasts are often used, which are spherical cells whose cell wall has been removed by mechanical means or digestive enzymes.86 Removal of the cell wall leaves the protoplast surrounded and protected by the plasma membrane only. Even though protoplasts are usually more sensitive to extracellular stresses than their native counterparts, they exhibit diverse advantages, e.g., simplified handling of single cells, ease of genetic manipulation, and use in screening experiments and single-cell microscopy. ζ-Potentials have also been measured for diverse plant cell protoplasts, e.g., from barley leaf, tobacco leaf, and Rauwolfia serpentina cultured cell protoplasts,87 and their surfaces typically exhibited a negative ζ-potential ranging from −6 to −28 mV.
Mammalian cells, unlike fungi and plant cells, lack cell walls. Instead, mammalian cells are protected by a dense gel-like meshwork abundant in carbohydrates, known as the glycocalyx, which overlays their plasma membrane.84 The glycocalyx constitutes a physical barrier for nanoparticles like pathogens to enter the cell, and it consists of various proteoglycans, glycosaminoglycans, glycolipids, and plasma proteins, which are important for cellular adhesion and signaling. The absence of cell walls makes mammalian cells more sensitive to changes in turgor pressure and shear forces.88 The surface charge of mammalian cells is typically negative at physiological pH. At pH 7.4, the ζ-potential for different types of cells showed variations over a wide range and was equal to −19.4 ± 0.8 mV for HeLa cells and −31.8 ± 1.1 mV for erythrocytes.89 The difference could presumably be attributed to the differences in the biochemical composition of the cell plasma. Exposure to 45 °C for 30 min induced apoptosis§ and necrosis¶ in 65% of the cells and decreased the ζ-potential from −19 mV to −25 mV. The authors argue that this can be attributed to the presence of larger amounts of the phospholipid phosphatidylserine on the cell surface, which is considered to be an early marker of apoptosis. In another study, ζ-potentials of different fixed cells were measured. Cell fixation is achieved by treating cells with fixatives (e.g., paraformaldehyde), which quickly kill the cell, prevent autolysis, and preserve the cell structure as faithfully as possible compared to the living state.90 Fixed cells can then be applied to different staining and microscopy analyses. The cell surface charges of CytoRich Red-fixed cells were found to be lower (−30 mV to −50 mV) than those reported for living cells (summarized in Table 1).90
| Types of cells | ζ | Exp. conditions | Ref. |
|---|---|---|---|
| E. coli | −4.9 to −33.9 mV | 1 mM NaCl | 91 |
| P. putida | −74.8 to −27.4 mV | 1 mM NaCl | 73 |
| S. aureus | −37.1 mV | 1 mM KCl | 77 |
| L. acidophilus | −45 mV | 1 mM NaCl, pH 7.4 | 76 |
| S. cerevisiae | −3 to − 26 mV | 5 mM NaNO3 at pH range 2–11 | 83 |
| Tobacco leaf protoplasts | −25 mV | 0.01 M KCl, 0.6 M sucrose, 6.7 mM sodium phosphate buffer (pH 5.8) | 87 |
| Barley leaf protoplasts | −18 mV | 0.6 M sorbitol, sodium phosphate buffer (pH 5.6) | 92 |
| HeLa | −19.4 ± 0.8 mV | PBS (1.7 mM KH2PO4, 5.2 mM Na2HPO4, 150 mM NaCl) | 89 |
| Erythrocytes | −31.8 ± 1.1 mV | PBS (1.7 mM KH2PO4, 5.2 mM Na2HPO4, 150 mM NaCl) | 89 |
| Diverse fixed human cell lines | −50 to −30 mV | Fixed cells resuspended in ultrapure water | 90 |
Having disclosed different classes of cells and examined their surface composition and charge, we will next discuss various methods to study their viability. This toolkit of knowledge is crucial for assessing abiotic coatings for cells and guiding the future development of this research field.
Cell viability methods
The viability of cells reflects their ability to sustain metabolic activity and structural integrity over a certain time frame. Typically, cell viability is defined as the number of actively proliferating or dividing cells in a sample or population;93 it is therefore an important parameter in many biological and biomedical settings. In the context of cell encapsulation, viability assays are commonly used to determine cell survival after encapsulation, or to assess the resistance of encapsulated cells to toxic chemicals (e.g., antibiotics) or physical stressors such as elevated temperature, extremes of pH, or radiation.17,18,22Cell viability assays can be classified into direct measurements that quantify the number of dividing cells (e.g., plating assays) or indirect assays, which measure various parameters as a proxy of cell viability (e.g., conversion of dyes or quantification of metabolic key intermediates). Due to the complexity of cells and their underlying metabolism, it may not always be straightforward to distinctly quantify cell viability, and the outcome likely depends on the assay that is used. The “culturability||” of cells remains the preferred definition of cell viability.94 Numerous techniques are available to assess cell viability (summarized in Table 2), including (i) cell counting of colony-forming units, (ii) membrane permeability assays, (iii) metabolic activity tests, (iv) luminometric adenosine triphosphate (ATP) measurements, (v) mitochondrial function assays, and (vi) inclusion dye evaluations.43
| Assay | Principle | Advantages | Disadvantages | Types of cells | Framework | Ref. |
|---|---|---|---|---|---|---|
| Agar plating | Detects viable cells based on colony formation on solid media | – Simple, rapid, and low-cost | – Limited to organisms that form isolated colonies | Bacteria (L. acidophilus; B. infantis) | ZIF-8 | 95 |
| – Applicable to a broad range of microorganisms | – Time-consuming | Bacteria (B. breve) | ZIF-8 | 21 | ||
| – Provides direct quantification of viable cells in a sample | – Not suitable for multicellular cell types | Bacteria (E. coli) | ZIF-8 | 96 | ||
| – Many cells may be viable but non-culturable | ||||||
| Membrane permeability assays | Assesses the permeability of a substance or substrate across the cell membrane | – Rapid and highly sensitive | – Matrix components may affect enzyme activity | Bacteria (P. putida) | MIL-100(Fe) | 108 |
| – High specificity of dyes for nucleic acids | – Apoptotic cells can retain membrane integrity | Bacteria (B. breve) | ZIF-8 | 21 | ||
| – Compatible with flow cytometry and fluorescence microscopy | – Membrane integrity can be influenced by growth conditions | Yeast (S. cerevisiae) | Cu-MOP | 109 | ||
| – Limited penetration of SYTO9 into Gram-negative bacteria | ||||||
| Luminometric ATP assays | Measures intracellular ATP levels | – Rapid with sensitive and robust signal output | – ATP levels can vary between different cell types or microorganisms | Mammalian cells (HeLa, A549, MCF-7, HLF, MCF-10A, RAW264.7, and B16 cells) | ZIF-8 | 123 |
| – High specificity | – ATP concentrations are dependent on growth conditions | Mammalian cells (human embryonic kidney, lung bronchial epithelial, and lung carcinoma epithelial cells) | MOF-801 | 124 | ||
| – Enables real-time analysis | Mammalian cells (HeLa) | ZIF-8, MIL-100(Fe), UiO-66-NH2, MET-3-Fe | 64 | |||
| – Suitable for high-throughput screening | ||||||
| Metabolic assays | Measures the activity of cellular metabolic pathways | – Simple, rapid, and cost-effective | – Results can be influenced by the physiological state of cells (e.g., dormancy) | Mammalian cells (neural stem cells) | HOF | 33 |
| – Applicable to a wide range of cell types | – Susceptible to interference from reducing agents and ROS scavengers | Mammalian cells (sperm cells) | ZIF-8 | 114 | ||
| – Cu(II)-containing complexes may affect absorption readings | Mammalian cells (CHO-K1) | Mn-based MOF | 115 | |||
| Mammalian cells (breast cancer cells MDA-MB-231) | ZIF-8 | 116 | ||||
| Bacteria (E. coli) | ZIF-8 | 70 | ||||
| Yeast (S. cerevisiae) | ZIF-8 | 12 | ||||
| Bacteria (Micrococcus luteus) | ZIF-8 | 12 | ||||
| Bacteria (E. coli) | ZIF-90 | 97 | ||||
| Flow cytometry assays | Detects changes or alterations in the cell membrane | – High-throughput compatible | – Requires expensive equipment | No examples yet | ||
| – Does not require staining | – Some stains are sensitive to EDTA | |||||
| – Sensitive ratiometric probes available | – Light-sensitive reagents | |||||
| – Compatible with a wide range of fluorescent dyes | – Time-consuming procedure | |||||
| Mitochondrial assays | Measures mitochondrial membrane integrity or membrane potential | – Provides insights into the cellular energy status | – Requires live cells (fixation is not possible) | No examples yet | ||
| Dye exclusion assays | Assesses cellular membrane integrity | – Simple and rapid | – Prone to over- or underestimation of cell numbers | No examples yet | ||
| – Compatible with a variety of dyes | – Requires preparation of cell suspensions | |||||
| – Results can be affected by the physiological state of the cells | ||||||
Cell counting methods
Counting of viable cells is a classical technique in microbiology and relies on the ability of cells to grow and divide on a nutrient-rich medium. When a cell suspension is spread onto a solid surface, only proliferating cells undergo binary fission and eventually form colonies of visible size referred to as colony-forming units (CFUs). CFUs therefore provide a direct readout of actively dividing cells in a sample. However, it should be noted that a variety of Gram-positive and Gram-negative bacteria, including E. coli (EHEC strains), can enter a dormant state in which cells remain alive and show metabolic activity, but do not show significant growth. Such a state was observed in response to starvation, at extremes of temperature and non-physiological oxygen concentrations. Wei et al.95 and Yuan et al.21 determined the viability of bacterial strains (L. acidophilus and Bifidobacterium longum subspecies infantis (B. infantis)95 and Bifidobacterium breve (B. breve))21 encapsulated in ZIF-8 biomineralized compartments by viable cell counting after removal of the MOF shell using ethylenediaminetetraacetic acid (EDTA). The strains showed diverse viabilities: while CFUs of L. acidophilus and B. breve were only slightly reduced compared to unencapsulated cells, the viability of B. infantis decreased by 2–4 orders of magnitude. For B. breve, CFU values after the dissolution of the MOF shell were comparable to a control using free cells.Luzuriaga et al.96 investigated the viability of E. coli cells encapsulated in a polycrystalline ZIF-8 shell. To determine viability, the ZIF-8 shell was removed by treatment with 500 mM sodium acetate buffer, pH 5, and the cells were spread on agar plates to test their ability to form visible colonies. However, no growth was observed, indicating that the cells were deactivated during the encapsulation and/or immobilization with the ZIF-8 shell.
Optical density (OD) measurements provide an alternative, inexpensive, and rapid method for qualitatively and quantitatively measuring positive variation in the cell number (cell growth) in a liquid medium. This method is based on the principle that cells scatter visible light; thus, a UV-vis spectrophotometer is typically used with a monochromatic wavelength at 600 nm. The extent of the scattered light, and thus the measured variation in the transmittance, is proportional to the density of cells (the number of cells per unit of volume) in a sample. This method was used by Gan et al.16 to assess the growth of S. cerevisiae cells released from ZIF-8- and ZIF-C-based shells with a thickness of 60 ± 20 nm. The released cells were inoculated into a liquid, nutrient-rich growth medium, and their proliferation was tracked by measuring the optical density at 600 nm. Although this technique does not provide a direct measure of cell viability in the ZIF composite, it revealed that coated cells exposed to external stressors had a shorter lag phase before the onset of exponential growth compared with uncoated cells.
Li et al.97 showed that E. coli cells encapsulated in ZIF-90 can be released and maintain viability. After removal of the ZIF shell and transfer to a nutrient-rich LB medium, bacterial growth was restored, although a delay in the onset of exponential growth was observed.
Ji et al.98 used cell counting to determine the growth of the photosynthetic anaerobic bacterium Moorella thermoacetica enclosed in MOF-shells with a thickness of 1–2 nm. The cells maintained full viability, with growth curves for both encapsulated and free cells being identical. The authors used super-resolution 3D-structured illumination microscopy (3D SIM) to directly visualize cell division of MOF-enclosed cells. The growth of encapsulated bacteria in an oxygen-containing atmosphere was found to be faster than that of free cells.
Flow cytometry is a technique that allows for the simultaneous multi-parametric analysis of the physical and chemical characteristics of single cells suspended in a liquid medium. Cells are first singularized before being subjected to a laser beam. As these cells pass through a laser beam, they scatter light and, if labeled with fluorescent markers, emit a fluorescence signal.99 Scattered and emitted light is detected and analyzed for various properties, such as size, granularity, and the presence of specific molecules, providing detailed insights into individual cellular characteristics. These parameters offer insights into the cell distribution and viability within the analyzed cell population.
In addition to scattering-based measurements, labeling of cells with a variety of fluorescent dyes that can be excited by the laser greatly extends the utility of flow cytometry. For example, the ratiometric membrane probe F2N12S** produces a green fluorescence (λexcitation = 405 nm; λemission = 530 nm) when bound to the membrane of healthy cells. When cells undergo apoptosis, a change in the membrane potential (i.e., the difference in the electric potential between the inside and outside of the cell) results in a red shift of the emission wavelength to 585 nm. The ratio of the two emission maxima allows for a quantitative and qualitative estimation of the cell viability. Flow cytometry can be combined with various dyes and staining techniques, as elaborated in detail by Kessel et al.100 In the following section, we discuss frequently employed methods in the context of cell encapsulation and coating.
Membrane permeability assays
Membrane permeability assays are based on alterations in membrane integrity that occur in dying or dead cells. Cell viability can be assessed by measuring the ability of certain molecules to penetrate the cell membrane.101 This method involves fluorescent or colorimetric dyes that interact with intracellular components inside the cells. Typically, these assays are carried out as live/dead staining by combining exclusion dyes, which penetrate cells with compromised membranes, with inclusion dyes that can pass through intact membranes. However, membrane permeability does not necessarily reflect cellular metabolic activity, as membrane integrity can also be affected by growth conditions, such as growth phase or environmental stress, potentially leading to under- or overestimation of cell viability in some cases.102 In mammalian cells, membrane permeability assays are often complemented by additional methods to distinguish between necrosis and apoptosis, as these processes play distinct roles in cellular responses. Apoptosis is a highly conserved, controlled, and programmed cell death.103 In contrast, necrosis is an uncontrolled process that results from external damage, such as toxins or environmental stress, and is associated with pathological responses.104 Importantly, the plasma membrane integrity is lost in necrotic cells, but is generally preserved during early stages of apoptosis; consequently, apoptotic cells may not be detected by many exclusion dyes.105 This distinction is crucial in the choice of staining methods.105Propidium iodide (PI) is a widely used membrane permeability probe that functions as an exclusion dye to stain dead cells. PI cannot enter living cells with intact plasma membranes, but readily penetrates dead or dying cells with compromised membrane integrity. Once inside the cells, the positively charged PI stoichiometrically intercalates with double-stranded nucleic acids. Upon excitation λexcitation = 488 nm, the PI-bound DNA complex exhibits fluorescence at λemission = 550 nm, enabling the quantification of inactive cells via fluorescence microscopy or flow cytometry. However, depending on the growth state, this method may yield a high fraction (up to 40%) of false positives, particularly during the early exponential growth phase of the cells. This increased uptake of PI by viable cells was linked to a temporary instability of the cell membrane due to cell wall reconstruction during cell division and growth, which may allow the dye to penetrate viable cells.106
Exclusion dyes are commonly used in conjunction with inclusive counterstains that have non-overlapping fluorescence spectra and can penetrate intact membranes of living cells. A frequently employed stain is SYTO9, which can enter both living and dead cells and exhibits enhanced fluorescence when bound to DNA (λexcitation = 485 nm; λemission = 498 nm) or RNA (λexcitation = 486 nm; λemission = 501 nm).102,107 SYTO9 is frequently used in combination with PI for live/dead staining since both dyes have distinct fluorescence profiles. Furthermore, PI has a higher affinity for DNA than SYTO9; thus, in situations where both dyes are present inside a cell, SYTO9 will be displaced.102 One known constraint of SYTO9 dyes is their limited ability to penetrate the cell walls of Gram-negative bacteria, which depends on their composition or active export from the cell.102
Permyakova et al.108 used fluorescence staining with SYTO9 and PI to discriminate living and dead P. putida cells encapsulated in MIL-100(Fe). This staining method allowed the qualitative evaluation of the living/dead cell ratio when coated with a MIL-100(Fe) exoskeleton. Under optimized conditions, the large majority of the encapsulated cells displayed intact cell membranes.
Using cell counting, Yuan et al.21 showed that the ZIF-8 coating moderately reduced the viability of B. breve cells. However, live/dead staining with PI and SYTO9 indicated pronounced damage to the cell walls after removal of the ZIF-8 shell. This damage was also evident from growth curves recorded by optical density: following inoculation with these cells of growth medium, the onset of exponential growth was significantly delayed when compared to an untreated control sample of B. breve cells. This study highlights the importance of employing a combination of different viability methods to obtain a more comprehensive understanding of the effect of encapsulation methods on cell viability.
An alternative counterstain that detects living cells or early apoptotic cells is Acridine Orange, which enters intact membranes and causes green fluorescence upon binding to DNA (λexcitation = 502 nm, λemission = 525 nm). A drawback of this dye is the necessity of washing steps to remove the unbound dye, since the fluorescence intensity is not notably enhanced upon binding to DNA. Qin et al.109 used a combined ethidium bromide/acridine orange stain to assess the effect of heat, reactive oxygen species, UV-radiation, and proteases on S. cerevisiae cells encapsulated in a copper metal–organic polyhedron (MOP) hydrogel. In this study, the authors showed by fluorescence microscopy that encapsulation decreased the percentage of dead cells after exposing the cell@MOP composite to the aforementioned physical, chemical, and biological stressors.
Metabolic assays
Metabolic assays are widely used to assess cell viability. Using these methods, viable cells with intact metabolism produce a measurable fluorimetric or colorimetric signal upon the addition of specific substrates, which can be correlated to the number of living cells. Conversely, dead or dying cells with compromised metabolism exhibit decreased or no conversion rates at all. Such assays can be conducted in conventional spectrophotometers or plate readers that are routinely available in biochemical or biological laboratories and thus can also be used in a high-throughput format.110An excellent indicator for cell viability is the presence of reducing nicotinamide cofactors (NADH or NADPH), which are metabolic key components. Several selective tetrazolium dyes are available to indirectly assess the concentration of these cofactors through the activity of intracellular, NAD(P)H-dependent redox enzymes. MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) is a widely used positively charged substrate that can easily penetrate through cell walls.110 It is converted through an unknown NAD(P)H-dependent metabolic process to insoluble formazans, which can be quantified spectrophotometrically at 570 nm after a solubilization step, providing a quantifiable measure of cell viability. Derivatives of MTT, such as MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium), XTT (2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide) or WTS (2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium), have been developed with negative sulfone groups and release soluble formazan derivatives.111 However, the dyes cannot cross the cell membrane due to their net negative charge. To overcome this, electron carriers such as PMS (5-methyl-phenazinium methyl sulfate) or PES (phenazine ethyl sulfate) are added to the assay. These carriers facilitate formazan reduction by shuttling electrons between the cytoplasm and the dye, producing a soluble formazan product that can be measured via spectroscopy.112
We note that under certain growth conditions, cells may undergo a state of dormancy in which they show greatly reduced metabolic activity but maintain viability. Therefore, assays require diligent control of the reaction conditions, including the concentration of the dye and the incubation time. The outcome of the assay can be influenced by the physiological state or the microbial strain being used. Tetrazolium-based assays are prone to a variety of interferences, as reviewed by Grabowiecka and coworkers.113 For example, unspecific reduction of MTT in the growth medium and the presence of radical scavengers can interfere with the assay and affect the result. Also, the presence of copper(II)-containing complexes can influence the original absorption of formazan.113 We note that shifts induced by the presence of cations may be of particular relevance in experiments with cell encapsulation into MOFs and other coordination compounds (e.g., MOPs). Yu et al. used the commercially available CCK-8 (cell counting kit-8) viability assay to assess cell viability of HOF-encapsulated neural stem cells.33 This colorimetric assay is based on the reduction of the tetrazolium salt WST-8 (2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt), which is converted to a water-soluble formazan derivative. In this case, encapsulation showed little effect on the biological activity of cells. The same assay was also used to determine the viability of “ZIFSpermbots”, consisting of spermatozoa encapsulated in a ZIF-8 framework.114
Ohtani et al.115 used the cell counting kit-8 to assess the viability of Chinese hamster ovary K1 cells (CHO-K1) in response to cyanide-bridged 2D coordination polymers (CPs) consisting of metal ions and networking metal complex lipids. The cells maintained more than 90% viability when challenged with 40 µM NiCl2 but lost 50% viability in the presence of the metal complex lipid (i.e. 10 µM (dabco-(CH2)15-CH3)2[MnN-(CN)4]).
The tetrazolium derivative MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) has been used to determine the viability of ZIF-8-encapsulated breast cancer cell line (MDA-MB-231 cell), showing that the viability of coated cells was ∼75% after incubation for 6 h.116
Calcein acetoxymethyl ester (calcein-AM) is another frequently used non-fluorescent metabolic marker that can passively cross the membranes of intact cells. Inside the cell, it is enzymatically hydrolyzed by unspecific esterases into the acidic, cell impermeable calcein (λexcitation = 494 nm; λemission = 517 nm), resulting in a strong, green fluorescence.117 Calcein-AM is frequently used as an indicator of metabolic activity and finds widespread application as an inclusion dye to visualize viable cells in live/dead fluorescence staining. Frequently, it is used in combination with PI. For example, Yu et al.33 used a differential staining with the dyes PI and calcein-AM to visualize the viability of HOF-encapsulated neural cells. Similarly, a dual calcein-AM/PI live/dead staining was used to assess the viability of MOF-encapsulated cancer cells.116 Chen et al. used a combination of the WST-8 dye and calcein-AM staining to show that “ZIFSpermbots”, consisting of spermatozoa encapsulated in a ZIF-8 framework, remained largely viable inside the framework, while the cell growth was arrested.114
Yan et al.70 determined the viability of E. coli cells encapsulated in ZIF-8 with the colorless probe fluorescein acetate (FDA). Upon metabolization, the chemical is cleaved by hydrolytic enzymes, thus releasing the highly fluorescent fluorescein. Enzymes facilitating this cleavage are unspecific esterases, lipases, or proteases.118 In this study, the authors did not observe viability differences in encapsulated E. coli cells. Similarly, Chen et al.119 used the FDA method in combination with CFU counting and growth curves to assess the viability of E. coli and S. cerevisiae following encapsulation by different ZIF-8 shells. Also, Falcaro and co-workers12,120 used FDA to monitor the time-dependent viability of S. cerevisiae cells encapsulated in a β-galactosidase/ZIF-8 shell. In some cases, fluorescent proteins produced by the cells themselves have been utilized to assess cell activity in ZIF-90-coated cells. Li et al.97 encapsulated E. coli cells that recombinantly produced the fluorescent reporter protein mCherry. However, protein expression had to be induced prior to MOF encapsulation, as the inducer (isopropyl β-D-1-thiogalactopyranoside, IPTG) could not permeate the molecular-sieving ZIF-90 shell. As a consequence, both viable and non-viable cells may exhibit fluorescence, and the fraction of encapsulated viable cells could not be determined.
A different viability assay is based on the quantification of intracellular ATP, the primary carrier of chemical energy in living cells. ATP is continuously synthesized and consumed as a result of various anabolic and catabolic processes, and its concentration is an indicator of intact cellular metabolism and physiology. At the onset of cell death, ATP levels typically decrease because cells lose their ability to replenish ATP. ATP levels can be quantified using commercial kits containing the enzyme firefly luciferase.121 In these assays, cells are first lysed to release intracellular ATP, and the resulting lysate is mixed with a luciferase solution. Luciferase catalyzes the ATP- and O2-dependent conversion of luciferin to oxyluciferin, producing a luminescent signal that correlates with the ATP concentration in the sample and thus overall cell viability. To ensure accurate ATP quantification, cells are lysed using detergents in the presence of ATPase inhibitors to prevent enzymatic ATP depletion. Several commercially available kits employ engineered, robust luciferase systems yielding luminescent signals stable for several hours. ATP assays are typically fast, with a workup procedure of a few minutes, and can be implemented in a high-throughput format, including 1536-well plate configurations.110 Additionally, the ATP assay can detect as few as 20 cells, while the MTT assay requires the presence of a minimum of ∼25000 cells.97 It is important to note that ATP concentrations can vary between different cell types and microbial strains and may also be influenced by the physiological state of the cell. Because ATP assays rely on enzymatic activity, careful consideration of media composition is required to avoid inhibition of luciferase. Overall, ATP assays offer a reliable and sensitive method for assessing cell viability.
A recently introduced variation of the ATP assay allows real-time assessment of viability. Here, a membrane-permeable pro-substrate of luciferin is added to the sample. Upon uptake, the pro-substrate is enzymatically converted to luciferin, which diffuses into the culture supernatant and is converted by luciferase to yield a luminescence signal.122
Previously, several commercially available ATP-quantification kits (CellTiter-Lumi Plus Luminescent Cell Viability Assay Kit and the CellTiter-Glo cell viability kit) have been used to monitor the viability of MOF-encapsulated mammalian cells125 and to test the tolerance of various cell lines including the human cervical carcinoma cell line (HeLa), human lung adenocarcinoma cell line (A549), human breast cancer cell line (MCF-7), and mouse melanoma cell line (B16) in ZIF-8, which has been proposed for cryoprotective applications.124 For example, ATP-based viability measurements indicated ∼90% viability for mammalian cell lines such as HeLa, A549, human promyelocytic leukemia (HL-60), and mouse macrophage Raw 264.7 cells encapsulated in ZIF-8 and were also used to determine the pH- and UV-tolerance of the cells.64
Another class of assays used to investigate cell viability are mitochondrial assays. Mitochondria are eukaryotic organelles central to cellular energy metabolism. Through oxidative phosphorylation, mitochondria consume oxygen while producing ATP as the main metabolic energy carrier. The functional state of mitochondria can therefore serve as an indicator of cellular viability and can be assessed using dyes that specifically target mitochondria.122 Commercially available mitochondrial membrane potential kits use cationic, lipophilic dyes (e.g., JC-10), which accumulate in the mitochondria and form aggregates. In this state, JC-10 produces red fluorescence (λexcitation = 570–590 nm).126 During apoptotic events, the dye diffuses into the cytoplasm, where it becomes monomeric, resulting in a shift in emission to 520–540 nm and the appearance of green fluorescence.126 Alternatively, calcein-AM in combination with CoCl2 has been used to assess the integrity of mitochondrial membranes. Calcein-AM is readily cleaved by intracellular esterases, producing fluorescence that is readily quenched by CoCl2 in the cytoplasm. In intact cells, the fluorescence is retained within mitochondria, allowing selective evaluation of mitochondrial membrane integrity. Upon membrane damage, the dye diffuses into the cytoplasm, resulting in a loss of mitochondria-localized fluorescence.126 Thus far, only a few studies applied mitochondrial assays to assess the viability of MOF-encapsulated cells. For example, Wang et al. demonstrated that the mitochondrial dye JC-1 can be used to quantify the mitochondrial membrane potential of mitochondria (Mito) encapsulated in ZIF-8 (Mito@ZIF-8).127 While the membrane potential of the free mitochondria rapidly decreased after isolation, the Mito@ZIF-8 samples maintained a relatively stable membrane potential and sustained ATP production for 48 hours. These results show the potential of the JC-1 assay for viability assessment in encapsulated cells, offering a sensitive and quantitative approach that could expand the current tools for assessing eukaryotic cell viability. In particular, mitochondrial dyes like JC-1 can detect the early onset of cell death, often before the cell membrane is compromised. This assay is less influenced by factors such as the shape, size, or density of mitochondria, which can alter the fluorescence intensity in single-component assays.128 However, researchers should carefully assess the potential barriers to implementation: these might primarily stem from the specific porous properties of the framework shells, which could restrict the diffusion of assay reagents. Additionally, the chemical instability of many frameworks (e.g., ZIF-8) when exposed to acidic or phosphate-rich environments could be incompatible with typical standard metabolic assay protocols. Furthermore, the potential light scattering or background autofluorescence introduced by the porous shell could interfere with the precise ratiometric readings required for probes like JC-1.
Other exclusion dyes
Trypan blue staining is a commonly used technique to assess the live-to-dead cell ratio.129 Addition of the trypan blue dye to a cell suspension results in the accumulation of the dye inside dead or dying cells with compromised cell membranes. Conversely, the dye cannot penetrate the cell membrane of intact cells, which remain unstained. The stained and unstained cells can be counted under a microscope using a cell counting chamber. Trypan blue staining is a simple and inexpensive technique that is commonly used in cell biology and immunology.129 Other commonly used stains that target cytoplasmic structures include eosin, Congo red, and erythrosine B. Erythrosine B staining is based on the ability of the dye to diffuse into the cytoplasm of cells with compromised membranes, where it binds to basic proteins. Viable cells remain unstained, whereas dead or dying cells appear pink. Congo red is a sulfonated azo dye that binds to amyloid proteins present in the cytoplasm.130 Eosin, a derivative of fluorescein, is an acidic dye that binds to basic cellular components, primarily cytoplasmic proteins.130 Upon binding, it retains a pink color. To date, none of these dyes has been applied to assess the viability of MOF-encapsulated cells.The following section builds upon the basic principles of artificial spores, cell types, and viability assessment methods and summarizes current progress in materials and coating strategies, providing detailed insights into reported protocols.
Materials and coating methods
Initial studies on cell encapsulation focused on using sol–gel methods to prepare rigid cell exoskeletons of porous SiO2 or TiO2.4,17,22,123 These archetypal oxide-based shells protect cells from mechanical stressors while allowing mass transfer between the environment and the encased cells. Although these oxides fulfilled key requirements such as permselectivity and durability, their chemical stability made controlled degradation challenging, especially without affecting cell viability. Interesting developments in engineering other inorganic nanoparticles, such as manganese dioxide, could be translated into protective shells, as MnO2 NPs can be degraded via glutathione (GSH) exposure. However, further progress in this field is required to examine the pros and cons of MnO2 for cell coating.131 Thus, the intrinsic chemical robustness of inorganic NPs could significantly limit their use in specific biotechnological applications aiming at the controlled release of cells, such as cell therapy.To address these challenges, current research focuses on the development of artificial coatings that meet all four criteria necessary for artificial spore formation (vide supra). A variety of natural and synthetic materials are under intense research for the fabrication of degradable exoskeletons under conditions that maintain cell compatibility. Examples include polysaccharide-based coatings such as starch, chitosan, and alginate, which can be enzymatically degraded,132 making them strong candidates for cell therapy. In addition, recent studies have merged the properties of organic and inorganic materials in hybrid coatings. Caruso et al.133 demonstrated the self-assembly of metal–organic coatings, utilizing Fe3+ and tannic acid, on S. cerevisiae cells. These metal–organic coatings are mechanically stable yet degradable on demand, thus meeting the criteria for artificial spore formation. The selection of appropriate building blocks allows control over chemical properties such as shell functionalization, self-assembly conditions, and degradability.
Building on these developments, three notable classes of microporous materials have emerged as promising options for encapsulating living cells and fragile biomolecules: MOFs, COFs, and HOFs.31,33 MOFs consist of inorganic clusters linked by multitopic organic linkers,35,36 while COFs and HOFs are assembled from organic compounds connected through covalent and hydrogen-bonding interactions, respectively.93 By carefully selecting the molecular building blocks, the stability, porosity, crystalline phase, and chemical and structural properties of these materials can be fine-tuned.
The microporous nature of the coatings provides permselective barriers that allow for the transport of small molecules such as glucose and oxygen, while preventing contact between the cell membrane and cytotoxic macromolecules like enzymes (e.g., trypsin and lyticase).16 These coatings can also be degraded on demand using chemical stimuli, such as chelating agents or pH changes for MOFs,31,56,134 or physical stimuli, such as light for HOF composites.33
Though post-synthetic modification methods have not yet been widely tested in this context, they have the potential to further expand the chemical versatility of these coatings.114 Given the properties of MOFs, COFs, and HOFs, they represent promising materials for the development of artificial spore-like systems. Two main approaches have been used for fabricating cell@MOF and cell@HOF composites: (i) the one-pot coating strategy and (ii) the multi-step cytoprotective encapsulation strategy.16
Cells@shell: one-pot encapsulation
In 2016, Falcaro and co-workers reported the one-pot encapsulation of S. cerevisiae and Micrococcus luteus (M. luteus) within a ZIF-8 exoskeleton (Fig. 4a).120 The cell coating was formed by mixing an aqueous zinc acetate solution with a premixed aqueous dispersion of cells and 2-methylimidazole (HmIM). After 10 min, the coated cells were recovered by centrifugation and washed with deionized water (DI water). X-ray diffraction (XRD) analysis confirmed the formation of ZIF-8 with sodalite topology (sod ZIF-8), and scanning electron microscopy (SEM) images showed individual cells encased in a continuous ZIF-8 exoskeleton with an average shell thickness of 100 ± 10 nm (Fig. 4c–e). Confocal scanning laser microscopy (CLSM) was also employed to assess the homogeneity of the ZIF-8 coating (Fig. 4f–h). The permselectivity of the cells@ZIF-8 composites was tested by incubating the coated and uncoated cells in a medium containing glucose (used as a nutrient) and lyticase, a cytotoxic biomacromolecule (molecular weight = 54.6 kDa). According to the viability test assay, the coated cells displayed a decrease of 19% in cell viability after 24 hours of exposure to lyticase. On the other hand, the control sample shows that non-coated cells experience a reduction of 95% in viability after only three hours of exposure (Fig. 4b). The bioprotection capabilities of the sod ZIF-8 coating were further validated by exposing the cells@ZIF-8 composites to an antifungal agent called filipin (molecular weight = 655 Da).140 The cell viability assay indicates that 90% of the cells surviving the coating process remain metabolically active, even after 24 h of exposure to filipin. In contrast, the control sample (naked yeast cells) showed nearly 100% mortality.
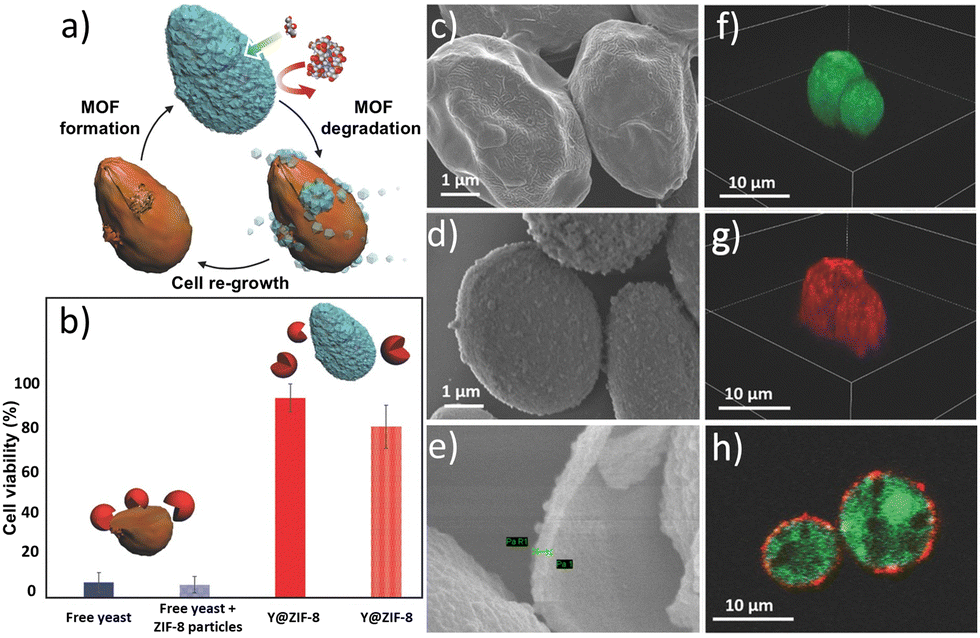 | ||
| Fig. 4 Biomimetic mineralization of yeast cells within a ZIF-8 coating. The schematic of the encapsulation and release processes is shown in (a). The cell viability (b) of yeast (blue) and yeast plus free ZIF-8 particles (patterned blue) after exposure to lyticase for 3 h and of yeast released from yeast@ZIF-8 previously exposed to lyticase for 3 h (red) and 24 h (patterned red) demonstrates the protection offered by the MOF shell. The homogeneity of the ZIF-8 coating was demonstrated by SEM (see images of native yeast (c) and ZIF-8 coated yeast (d) and of the calcined yeast@ZIF-8 sample (e)) and by labelling the living yeast cells with FDA (green), and the ZIF-8 coatings with Alexa Fluor 647 fluorescent dye (red) (see the 3D cellular reconstruction of CLSM images (f) and (g) and a cross-section CLSM image (h) of yeast@ZIF-8). Adapted with permission from ref. 120 Copyright 2016, Wiley-VCH. | ||
Finally, the optical density measurements at λ = 600 nm (OD600) obtained from the coated and uncoated cells incubated in a rich-nutrient medium demonstrated that the MOF shell formation inhibits cell division; this configuration mimics a spore-induced state. Upon exposure of cells@ZIF-8 to an EDTA solution, the decomposition of the MOF shell allows the resumption of cell division in those cells that remain viable after the coating/de-coating process.
In a following study, the authors used S. cerevisiae as model cells to fabricate a bioactive MOF exoskeleton; such an exoskeleton was designed to impart cell adaptability in nutrient-deficient environments.12 The yeast cells were first coated with β-galactosidase (β-gal),141 an exogenous enzyme adsorbed onto the cell wall due to electrostatic interaction between the positively charged enzyme and the negatively charged cell wall. The protein-decorated cell system was then resuspended in an aqueous solution of HmIM, followed by the rapid addition of aqueous zinc acetate to induce the spontaneous formation of the ZIF-8 exoskeleton. CLSM was used to demonstrate the co-localization of the β-gal (labeled with purple Alexa Fluor 568) and the MOF coating (infiltrated with red Alexa Fluor 647). The SEM images of yeast@β-gal@ZIF-8 composites confirm the formation of a continuous ZIF coating with an average thickness of 100 nm ± 10 nm. The yeast@β-gal@ZIF-8 composite was incubated in an aqueous solution of lactose, a sugar that cannot be metabolized by S. cerevisiae cells. The immobilization of β-gal in the ZIF shell allowed the hydrolysis of lactose to glucose and galactose, two sugars that can be used by the cell as nutrients. Thus, the immobilization of a non-native enzyme (β-gal) between the cell wall and the MOF coating conferred adaptability to nutrient-depleted environments. In the same study, naked yeast (control) and the coated cells (yeast@β-gal@ZIF-8) were incubated in a nutrient-deficient medium containing biomacromolecules that are detrimental to both yeast (e.g., lyticase) and β-gal (e.g., proteases). The cell viability tests indicate that after seven days, ∼70% of the coated cells that were initially viable after the coating process remained alive, while the viability of naked yeast rapidly decreased to 10% within the first days of incubation. Overall, this study showed that the ZIF-8 coating acts as a semipermeable barrier (i.e., molecular sieve) allowing the diffusion of non-nutrients and their conversion into nutrients via the immobilized β-gal and preventing contact between cytotoxic lyticase and cells. Lastly, on-demand release of the protective coating was demonstrated by exposing the enzyme-functionalized ZIF-coated cells to EDTA.
The potential of abiotic ZIF shells was further expanded by Gan et al.,16 who reported the synthesis of bioactive multi-layered ZIF coatings on yeast cells (Y). Specifically, the authors immobilized a protease inhibitor, alpha-1-antitrypsin (AAT), between the two MOF layers constituting the abiotic exoskeleton to impart cell adaptability against protease-rich environments (Fig. 5a). The synthesis of a multilayered ZIF-8 coating was achieved by inducing a sod ZIF-8 layer via the biomimetic mineralization strategy to afford the Y@ZIF-8 biocomposite. Then, Y@ZIF-8 was exposed to AAT, which was adsorbed onto the outer surface of the ZIF-8 exoskeleton to yield Y@ZIF-8@AAT. Finally, Y@ZIF-8@AAT was exposed to a fresh solution of ZIF precursors, in which the pre-adsorbed AAT triggered the on-site formation of a second ZIF layer. This work showed that the bio-replication approach previously applied to synthetic substrates (e.g., silicon, glass, polystyrene, and polypropylene)61,120 to facilitate the MOF growth can be successfully implemented in biological systems. We note that the crystalline phase and composition of the second yeast ZIF coating could be tuned by varying the Zn2+:HmIM ratio, i.e. the outer MOF layer could be either sod ZIF-8 or ZIF-C (Fig. 5b). This allows for the preparation of two different systems: (i) Y@ZIF-8@AAT@ZIF-8 and (ii) Y@ZIF-8@AAT@ZIF-C. It should be noticed that sod ZIF-8 and ZIF-C display differences in their chemical compositions (Zn(mIM)2 for sod ZIF-8 and Zn2(mIM)2CO3 for ZIF-C), crystalline structures, and porosity. The different MOF outer layers can imbue the MOF biocomposites with unique properties. For example, in terms of porosity, sod ZIF-8 is microporous, and ZIF-C is nonporous.56 The release profile for ZIF-C is faster for encapsulated biomolecules at pH = 6.5,56 and ZIF-C has lower cytotoxicity in specific cancer cells (e.g. human prostate cancer cells (PC-3)).142 To investigate the biopreservation performance of both materials (i.e., sod ZIF-8 and ZIF-C), the authors incubated both composites, Y@ZIF-8@AAT@ZIF-8 and Y@ZIF-8@AAT@ZIF-C, in a protease-rich medium. The latter was prepared by dissolving trypsin in a phosphate-buffered solution (pH = 6.5); the presence of phosphate anions in the incubation media triggered the slow degradation of the ZIF shell,139 followed by the controlled release of the AAT in solution (Fig. 5c). The release profiles recorded for AAT show that Y@ZIF-8@AAT@ZIF-C releases 50% of the biomacromolecule within the first 2 h. In contrast, the Y@ZIF-8@AAT@ZIF-8 composite requires about 18 h to release 50% of AAT. These results indicate that the crystalline phase of the outer shell directly affects the MOF degradation and, thereby, the release kinetics of AAT. Finally, a trypsin activity assay was used to assess protease inhibitor efficiency. This test revealed that the trypsin becomes completely inhibited once the AAT is fully released from the abiotic coating. Subsequently, the released cells were incubated in a yeast growth medium (yeast extract-peptone-dextrose, YPD) to evaluate the cell proliferation by OD600 measurements. This experiment demonstrated that the released yeast cells exhibit exponential proliferation when placed in nutrient-rich media. A similar concept, combining yeast and enzyme by utilizing MOFs to enhance enzyme activity, was reported by Zhan and co-workers in 2023.143 These studies indicate that yeast@ZIF-8 composites can serve as a platform for biocompatible immobilization materials and effective biocatalysts. Wang and co-workers reported a vaccine adjuvant application from the yeast-derived MOF composite named yeast@Mn-MOF-74@ZIF-8.144 Yeast and MOFs can serve as antigen display carriers, but they cause different immune responses. Yeast can activate the adjuvant properties of cellular immunity, while MOFs can induce strong humoral immune responses. The yeast@Mn-MOF-74@ZIF-8 composite can not only be used as a delivery system for subunit vaccine antigens but also as an immunostimulant in subunit vaccine and inactivated virus vaccine preparations. In this study, the yeast@Mn-MOF-74@ZIF-8 composite demonstrated promising application potential.
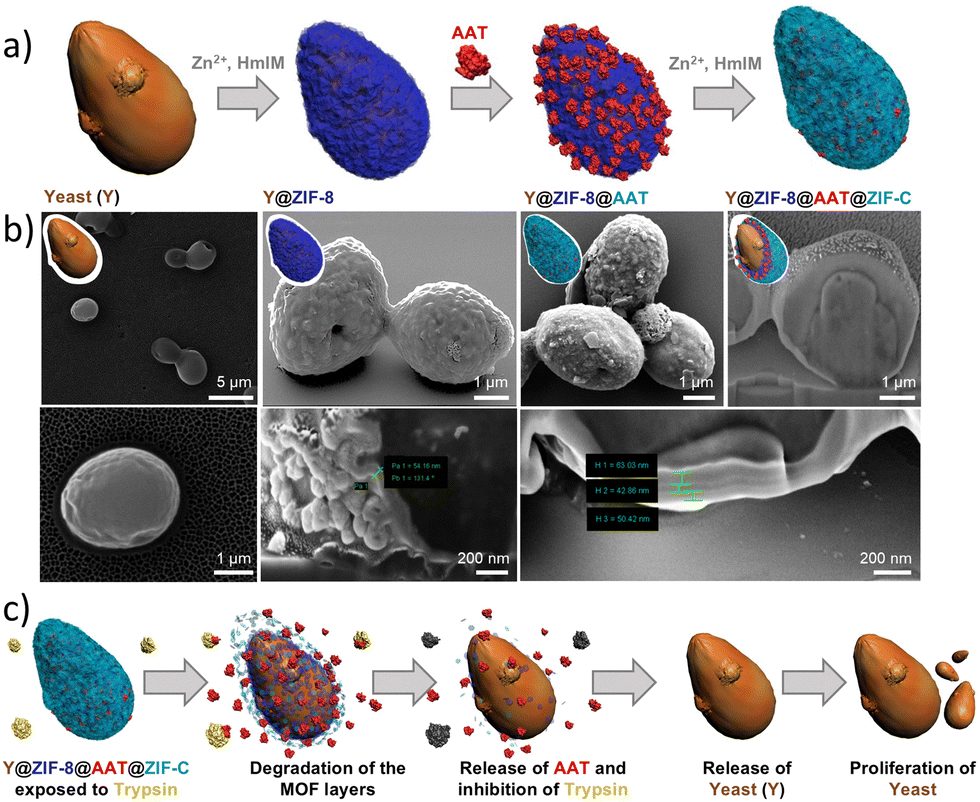 | ||
| Fig. 5 Multi-layered ZIF-coated cells. Schematic of the encapsulation process (a). SEM images and cross-section analysis of Y@ZIF-8, Y@ZIF-8@BSA@ZIF-8, and Y@ZIF-8@BSA@ZIF-C composites (b). Schematic of yeast cell proliferation upon release of yeast cells in the presence of trypsin (c). Adapted with permission from ref. 16 Royal Society of Chemistry. | ||
The progress of ZIF-based abiotic shells showcases that protective exoskeleton functions of biomineralized MOFs can be engineered with biomacromolecules; by selecting the MOF coating property (e.g., phase and thickness) and the biomolecule property (e.g., enzymes and enzyme inhibitors), the coating enables functional adaptability, converting hostile environments (e.g., nutrient-deficient and lyase-rich media) into biocompatible ones. Subsequent programmable release of cells from their ZIF coating can enable the restoration of the proliferation functions of the cells surviving the coating/de-coating procedure.
More recently, Luo et al.148 reported the cascade biosynthesis of D-phenyllactic acid (D-PLA) from L-phenylalanine using two E. coli strains (pET28a-lrldh and pET28a-ladd2) immobilized in an amorphous ZIF-90 (aZIF-90) exoskeleton. The encapsulation was achieved via a one-pot method by mixing Zn(NO3)2 with imidazole-2-carboxaldehyde (HICA) in the presence of the E. coli strains in a 1:1 mixture. The resulting amorphous E. coli@aZIF-90 biocomposites were characterized via SEM, infrared spectroscopy, thermogravimetric analysis (TGA), XRD, X-ray photoelectron spectroscopy (XPS), and CLSM, confirming successful cell encapsulation and a mesoporous structure conducive to substrate diffusion. Catalytic performance assays revealed that aZIF-90 encapsulation enhanced the cells’ thermal and pH stability, tolerance to metal ions and organic solvents, and retained >75% of activity over four reuse cycles. In a fed-batch system, the immobilized biocatalyst reached a peak yield of 9.00 g L−1 D-PLA with an 89.4% conversion rate after 12 hours. Compared to free cells, E. coli@aZIF-90 exhibited a 1.15-fold increase in space–time yield and superior resistance to harsh environmental conditions. These results underscore the potential of aZIF-90 as a robust platform for whole-cell catalysis in industrial bioproduction of D-PLA.
One-pot encapsulation of living cells within abiotic MOF coatings is not restricted to Zn-based MOFs. Recent studies suggest that Fe-based MOFs are suitable candidates for one-pot encapsulation of living microorganisms. Lee et al.63 reported the encapsulation of the model cell S. cerevisiae within a MOF coating made of Fe3+ ions and benzene-1,3,5-tricarboxylate (BTC) linkers. This abiotic coating was formed by mixing an aqueous solution of FeCl3 with a premixed aqueous dispersion of cells and BTC. After 1 min of stirring, the resulting yeast@Fe-BTC composites were recovered by centrifugation and washed with DI water. This MOF coating process was repeated three times to obtain a robust MOF coating. The ζ-potential for uncoated S. cerevisiae cells is ζ = −37.8, while for the resulting yeast@Fe-BTC composite, it is ζ = −13. Cell viability was assessed by the live/dead assay, which monitors cell membrane integrity and internal esterase activity. This test indicates that 95% of yeast cells remained metabolically active after the encapsulation process, confirming the cytocompatibility of the MOF coating process. Then, to impart cell adaptability against cytotoxic agents like octyl-β-D-glucopyranoside, the MOF shell was functionalized with a set of exogenous enzymes, including β-glucosidase (β-glu), glucose oxidase (GOx), and horseradish peroxidase (HRP), yielding a yeast@β-glu&GOx&HRP@Fe-BTC composite with a bioactive MOF shell. This bioactive shell was designed to enable an enzymatic cascade reaction that starts with β-glu, which cleaves the O-glycosidic bond at C1 of octyl-β-D-glucopyranoside to yield 1-octanol and D-glucose. Then, GOx transforms D-glucose and O2 to D-gluconic acid and H2O2. Finally, HRP uses H2O2 as a co-substrate for the catalytic oxidation of 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) into its radical cation (ABTS+˙). The radical cation, which has a distinct blue color, was used to monitor the feasibility of the enzymatic cascade reaction when exposing the encased cells to an octyl-β-D-glucopyranoside-rich medium.
Then, to evaluate the bioprotection capabilities of the bioactive coating to the encased cells, the authors compared the viability of the yeast@β-glu&GOx&HRP@Fe-BTC and the free yeast cells after being exposed to lethal concentrations of octyl-β-D-glucopyranoside (20 × 10−3, 50 × 10−3, and 100 × 10−3 M) for 48 h. Cell viability measured upon exposing yeast@β-glu&GOx&HRP@Fe-BTC at each lethal concentration was 47.7%, 47.3%, and 42.7%, respectively. In contrast, for the free cells exposed to the same cytotoxic conditions, the viability values obtained were 17.4%, 16.2%, and 14.6%, respectively. These observations support the integration of multiple exogenous enzymes into the MOF coating without compromising their catalytic activity. This study further expands the versatility of MOF coatings for designing bioactive shells capable of mitigating the adverse effects of specific cytotoxic agents.
More recently, Sicard and co-workers reported the one-pot encapsulation of P. putida CFBP 5039 within a Fe-based MOF known as MIL-100(Fe) (MIL = Matériaux Institut Lavoisier).108 MIL-100(Fe) is a mesoporous crystalline material comprised of Fe-oxo trimers interconnected by BTC linkers.149 The synthesis of MIL-100 around living cells required carefully optimized conditions to form a crystalline MOF coating without compromising cell viability. To determine the concentration threshold at which each molecular precursor begins to drive cell stress, the authors exposed the P. putida cells to various concentrations of MOF precursors. Then, the bacteria's integrity was assessed by a live/dead assay using propidium iodide (PI), a fluorescent dye that only permeates damaged cell membranes, and SYTO9, a fluorescent probe that permeates healthy membranes. The study indicates that when using diluted concentrations of BTC (8.5 mM) and iron(III) nitrate (13 mM), the cell membrane remains intact, suggesting that these are the optimal concentrations to prepare a MIL-100 MOF coating. The one-pot synthesis was performed by mixing aqueous solutions of MOF precursors in the presence of the cells; this mixture was kept at 30 °C for 21 h. The XRD pattern of the resultant biocomposite indicates the successful formation of crystalline MIL-100(Fe). Furthermore, the TEM micrographs (Fig. 6a–c) and elemental mappings (Fig. 6d) indicate the formation of a continuous MOF coating with a thickness of 30–60 nm. Interestingly, TEM images reveal that the MOF particles are not directly attached to the cell wall; instead, a gap of ∼60–80 nm is observed between the MOF coating and the cell. The authors suggested that this separation might be caused by an exopolysaccharide (EPS) network surrounding the bacterium. The EPS secretion is typically observed in many Gram-negative bacterial strains during their stationary phase. The author claimed that the functional groups of the EPS network (–OH, –COOH, –NH2) might enhance local interactions between the cell and Fe ions, leading to the heterogeneous nucleation of MOF crystals at the cell's surroundings, mimicking the natural mineralization process.
 | ||
| Fig. 6 One-pot encapsulation of P. putida within MIL-100. TEM images (a) and (b) and STEM-HAADF images (c) of P. putida@MIL-100(Fe). The STEM-HAADF image and STEM-XEDS elemental maps of the biohybrid material (d) demonstrate the localization of the MOF shell on the bacterial surface. Adapted with permission from ref. 108 Copyright 2022, American Chemical Society. | ||
In 2024, the one-pot encapsulation of living cells within abiotic coatings was expanded from MOFs to COFs. J. Liang and co-workers coated S. cerevisiae cells with a COF based on p-phenylenediamine and benzene-1,3,5-tricarboxaldehyde (COF-LZU1).150 After suspending the cells in a solution containing the two COF precursors, COF formation was induced by subsequent addition of acetic acid and sodium hydroxide, for a reaction time of 15 min. Microscopy studies showed that the COF shell was a uniform thin film of approximately 40 nm firmly adhering to the cell wall and particle aggregates. The authors attributed the film homogeneity to the formation of covalent bonds between the COF material and the amine or thiol groups of (glyco) proteins in the yeast cell wall during the one-pot reaction. Notably, upon encapsulation within COFs, 72.5% of cellular activity was retained (resazurin assay results), suggesting that the rapid formation of the COF coating minimizes the harmful effects on yeast cells induced by exposure to acetic acid and sodium hydroxide. The COF coating provided a high degree of protection against different stressors (high temperatures, pH fluctuations, oxidative stress, high metal ion contents, bisphenol A, and UV radiation). Furthermore, by incorporating exogenous enzymes (i.e., catalase) into the COF coating, stable yeast fermentation and ethanol production were achieved. Building on this approach, in 2025, P. thermoglucosidasius Gt-08, a Gram-positive bacterium genetically optimized for high riboflavin production, was encapsulated into COF-42.151 Upon encapsulation, the authors showed that the production of riboflavin was preserved, demonstrating that the COF shell effectively safeguards cells. The one-pot encapsulation is based on the exposure of the bacteria to an acetic acid and water solution of one of the COF-42 precursors (1,3,5-triformylbenzene). Subsequent addition of the second COF precursor (2,5-diethoxyterephthalohydrazide) induced immediate COF formation and precipitation. Microscopy and structural investigations confirmed the growth of a COF thin film (35–80 nm) around the cells. ζ-Potential measurements demonstrated the interaction between the cell surface and the acid-activated 1,3,5-triformylbenzene: bare cells showed a ζ-potential of −12.4 mV, whereas the exposure to the COF ligand increased the ζ-potential to +15.5 mV. Based on these data, the authors hypothesized that the enrichment of the cell surface with 1,3,5-triformylbenzene promoted the COF nucleation directly at the cell surface, leading to the homogeneous coating observed via electron microscopy. Overall, these findings indicate that COF precursor–cell surface interactions follow principles analogous to MOFs on cell surfaces, where electrostatic interactions between the cell and precursors play a crucial role, highlighting mechanistic similarities in the growth of different extended framework materials on living cells.
Yang et al. encapsulated cancer cells using ZIF-8125 to apply these cells as whole-cell cancer vaccines. Here, a biocompatible and selective cell encapsulation strategy based on a precursor-functionalized nucleolin aptamer and in situ MOF mineralization on the aptamer-identified cancer cell surface was developed. After MOF coating, the encapsulated cancer cells (HeLa, A549, MCF-7, and B16 cells) underwent immunogenic cell death, which was associated with variations in cell stiffness. As aspired, immunogenic dead cancer cells efficiently exposed calreticulin, a hallmark of efficient whole-cell cancer vaccines, on their cell surface, and were able to release antigens and induce in vivo antitumor T-cell immune responses.
An alternative strategy was shown for neural stem cells, which were encapsulated with biocompatible HOFs.33 These composites formed at the cytomembrane of neural stem cells via electrostatic interaction, as well as hydrogen-bonding interactions between protein residues on the cytomembrane and the HOF building blocks. Additionally, porous carbon nanosphere nanozymes (PCNs) were doped into the HOF shells to endow the cellular exoskeletons with hierarchical hydrogen bonds, NIR-II triggered degradation, and antioxidant activity. Neither the biological activity nor the cell viability of neural stem cells was influenced by the encapsulation process. Neural stem cells are of great interest for the treatment of neurodegenerative diseases, but successful transplantations are often impeded due to loss of ‘stemness’, cytomembrane damage, and apoptosis resulting from the oxidative stress in the adverse pathological microenvironment. To prove that HOF-encapsulated cells can be transplanted, they were injected into the brains of neurodegenerative disease mouse models. These mice indeed exhibited ameliorated neurogenesis.
Even though the number of these pioneering examples is still limited, they suggest that this could be a promising approach for creating high-performing carriers for biotherapeutics and vaccines in the near future.
Cells@shell: multi-step encapsulation
Jeon et al. applied zirconium (Zr)-based MOF-801 NPs as a cryoprotectant for human embryonic kidney (293T), nontumorigenic lung bronchial epithelial (BEAS-2B), and lung carcinoma epithelial (A549) cell lines.124 MOF NPs with diameters of 10, 35, 100, and 250 nm were prepared. In this approach, the amino acids valine and threonine were introduced into the MOF NP-surface through the acrylate-based functionalization to mimic ice-binding proteins and provide surfaces with hydrophilic and hydrophobic dualities. The MOF-801 NPs were biocompatible regardless of concentration or NP surface-functionalization, whereas the smaller-sized surface-functionalized NPs showed a good cell recovery rate after freezing/thawing by inhibiting ice recrystallization.
So-called ZIFSpermbots were created by encapsulating sperm cells using pre-synthetized ZIF-8 NPs.114 Coating of sperm membranes was facilitated through complexation with tannic acid, resulting in selectively permeable, porous ZIF-8 wrappings. This cell surface engineering had a negligible impact on sperm motility under optimized conditions, whereas it efficiently blocked the binding of antisperm antibodies. These so-called “ZIFSpermbots” may be used as active drug delivery systems by making use of the drug-loading capacity of ZIF-8 NPs.
MOF cell–surface coatings can also be applied to create microdomains on the cell membrane of mammalian cells like Chinese hamster ovary K1 (CHO-K1).115 For this approach, a metal complex lipid (dabco-(CH2)15-CH3)2[MnN-(CN)4] was inserted into mammalian cell membranes through simple incubation and cross-linked by adding Ni cations, forming stable MOF microdomains and leading to phase separation. The induced phase separation systems remain stable even in the absence of the actin cytoskeleton. Moreover, these cells showed enhanced cellular calcium response to ATP due to the activation of P2 purinoceptors.
Emerging applications of framework-encapsulated cells
Immunotherapy
The preservation of cell viability during cell@shell formation opens the possibility of using these composites in novel applications. For instance, degradable abiotic shells to encase living bacteria may significantly improve bacterial-mediated antitumor therapy (BMAT).27,43,158–160 Although the use of live bacteria for cancer treatment has been known since the 19th century, the risks of uncontrolled bacterial infection have hindered the clinical application of BMAT.161 Nevertheless, with the advent of new antibiotics and genetic engineering techniques, researchers have developed safe strategies for using attenuated bacteria in cancer therapy.158 When compared to traditional chemotherapy, BMAT provides several advantages: (1) tumor targeting: the hypoxic tumor microenvironment promotes bacterial colonization,162 (2) intratumoral penetration: the bacterial flagella enhance the bacteria's ability to penetrate within the tumor mass,163 (3) tumor colonization: the immunosuppression of the tumor microenvironment promotes the preferential accumulation of bacteria in the cancer tissue;43,158 and (4) development of bacteria-based delivery systems, where by genetic engineering and bacterial surface modification, it is possible to create delivery systems that use living cells as carriers.43,158 Therefore, these systems are ideal for delivering antitumoral therapeutics with high spatial and temporal precision.14,158,161One effective way to enhance the efficacy of bacteria-based therapy is encapsulation of living microorganisms within an abiotic shell. This strategy enables co-delivery of therapeutic agents to cancer cells, prolongs bacterial circulation lifespan, and improves regulation of bacterial proliferation at the treatment site.43,158 A recent study by Yan and co-workers showcased the synergistic potential between the abiotic MOF coating and bacterial-based cancer therapy.70 In this work, the authors reported the one-pot encapsulation of E. coli (ζ = −23) within a bioactive ZIF-8 coating to yield E. coli@ZIF-8 composites (ζ = 7.5).70 The ZIF-8 coating preserved the viability of the encased cells while preventing uncontrolled bacterial replication in healthy tissue. To further improve the therapeutic efficacy of the E. coli@ZIF-8 composites, the researchers used the MOF shell for the co-immobilization of two therapeutic agents, doxorubicin (DOX) and chlorin e6 (Ce6), resulting in an E. coli@DOX&Ce6@ZIF-8 composite (ζ = −14). DOX is an anthracycline antibiotic commonly used in chemotherapy,164 and Ce6 is an FDA-approved photosensitizer utilized in cancer photodynamic therapy (PDT).165 Ce6 is known for its high reactive oxygen species (ROS) generation efficiency upon exposure to mild near-infrared (NIR) irradiation. The NIR radiation, which ranges from 800 to 2500 nm, is less phototoxic than UV or high-energy visible light. In addition, in mammalian tissues, NIR light penetrates more deeply than visible light, making it more suitable for treating deeper-seated wounds, infections, and cancers.166
The chemo-photodynamic therapeutic efficacy of the E. coli@DOX&Ce6@ZIF-8 composite was evaluated through both in vitro and in vivo experiments. The in vitro tests were performed by evaluating the cell viability of mouse breast tumor (4T1) cells after different treatments with and without laser exposure (L). These treatments included: (1) free E. coli, (2) E. coli@ZIF-8, (3) E. coli@DOX&Ce6@ZIF-8, (4) E. coli@Ce6@ZIF-8 + L, and (5) E. coli@DOX&Ce6@ZIF-8 + L. The results showed that 4T1 cells retained over 80% viability after being exposed to E. coli@ZIF-8. This observation confirms the biocompatibility of the E. coli@ZIF-8 composite. However, the cell viability dropped to ∼60% after exposure to the E. coli@DOX&Ce6@ZIF-8 composite. The moderate toxicity of the E. coli@DOX&Ce6@ZIF-8 composite was attributed to the release of the chemotherapeutic drug DOX. Nevertheless, the therapeutic efficacy of the E. coli@DOX&Ce6@ZIF-8 composite could be intensified with NIR laser exposure. Thus, the dual chemo-photodynamic therapy provided by the E. coli@DOX&Ce6@ZIF-8 + L treatment resulted in a cell viability of ∼25%. These observations indicate that the strong synergy between Ce6 and DOX leads to higher therapeutic efficacy in in vitro tests. To explore the efficacy of the dual chemo-photodynamic therapy, the authors investigated the biodistribution of E. coli@DOX&Ce6@ZIF-8 in mice. E. coli@DOX&Ce6@ZIF-8 was injected into tumor-bearing mice, and the biodistribution analysis after 24 h revealed that the accumulation efficacy of E. coli@DOX&Ce6@ZIF-8 was ∼6.1%. This value was obtained from the ratio of E. coli colonies in tumor sites to the injected number of E. coli@DOX&Ce6@ZIF-8. Then, by tracking the tumor size and weight, the authors demonstrated that E. coli@DOX&Ce6@ZIF-8 inhibits tumor growth after a one-time NIR laser treatment (10 min, λ = 600 nm). This study highlights the versatility of using bacteria@MOF coatings as a cell-based delivery platform for biotherapeutic applications.
The MOF bioprotection can also be extended to cellular organelles. This was recently demonstrated by Zhou and co-workers, who reported a novel strategy to biomineralize isolated mitochondria within ZIF-8,167 to preserve their bioactivity and enhance their transplantation efficiency into cancer cells for therapeutic purposes. The mitochondria were isolated from non-tumorigenic mammary epithelial cells (MCF-10A). The encapsulation was achieved through a one-pot synthesis method by mixing the freshly isolated mitochondria with Zn2+ and HmIM in a 0.9% NaCl solution. To improve intracellular delivery, the resulting MIT@ZIF-8 nanostructures were further surface-functionalized by incorporating polyethyleneimine (PEI) and the cell-penetrating peptide TAT during the synthesis process. TEM images confirmed the formation of a distinct ZIF-8 layer around the mitochondria, and cut-section analysis revealed the inclusion structures. Fluorescence lifetime measurements of a mitochondrial-binding dye (MVG) further confirmed the full encapsulation of mitochondria by the ZIF-8 shell. This study indicates that the encapsulated mitochondria effectively maintained their bioactivity, assessed by mitochondrial membrane potential and ATP synthesis capability, for at least 4 weeks at room temperature. In contrast, non-encapsulated controls, even when stored on ice, lost a significant portion of their membrane potential within 6 hours. The MIT@ZIF-8 nanostructures were shown to release the encapsulated mitochondria in response to an acidic environment, with approximately 70% release at pH 5.0 after 6 hours, compared to minimal release at pH 7.4. The surface modification of MIT@ZIF-8 with PEI and TAT resulted in improved aqueous dispersion, which enhanced cellular uptake. The modified MIT@ZIF-8 was successfully delivered into breast cancer cell lines (BT-549 and MDA-MB-231) with uptake efficiencies of 11.8% and 17.2%, respectively, after 4 days of incubation. CLSM imaging showed the presence of these exogenous mitochondria within the recipient cancer cells, and instances of fusion between the transplanted and endogenous mitochondria were observed. Functional analysis via Seahorse assays revealed an increased oxygen consumption rate (OCR) and an extracellular acidification rate (ECAR) in cancer cells that received MIT@ZIF-8, indicating improved mitochondrial function. Therapeutically, the transplantation of these non-tumorigenic mitochondria into cancer cells resulted in significant inhibition of cancer cell proliferation, reduced the cancer stem cell population in MDA-MB-231 cells from 95.5% to 76.5%, and suppressed the epithelial–mesenchymal transition (EMT) process in these cells. The authors concluded that this MOF-based biomineralization technique represents an advancement for mitochondrial research and transplantation, offering a robust method to preserve mitochondrial activity and enhance mitochondrial delivery for potential cancer therapy.
Probiotic bacteria and enzyme delivery
A different research direction to explore the potential of cell@ZIF-8 composites targets the encapsulation of probiotics. Probiotics are microorganisms that confer health benefits when administered in adequate amounts to humans or animals, as they maintain, restore, and balance intestinal microflora.168,169 The latter is especially crucial for the clinical control of intestinal tract infections.170,171 However, the beneficial effects of the probiotic bacteria can only be achieved if the microorganism reaches the intestinal target in a viable state.168,169 Nevertheless, from a practical point of view, this represents an important challenge, since orally administered probiotics are subjected to strong acidic conditions during their journey through the gastrointestinal tract.169 Such acidic172 conditions can severely hamper the probiotic viability, limiting their efficacy in restoring gut flora. On the other hand, the clinical treatment of intestinal infections typically requires the co-administration of antibiotics and probiotics. Therefore, to ensure the effectiveness of the treatment, it is crucial to develop strategies that provide probiotics with protection against antibiotics, such that antibiotics only kill infectious pathogens without altering the viability and functionality of probiotics.173Microencapsulation technology has emerged as a promising solution to tackle these issues and boost probiotic viability when deployed alongside antibiotics for treating intestinal infections. Traditional microencapsulation techniques involve immobilizing probiotics within organic matrices.174,175 One crucial aspect of microencapsulation is ensuring that the encapsulating material can release the probiotic cells at the site of action. Advances in nanotechnology have led to the development of novel abiotic coatings that enable targeted delivery of probiotics upon exposure to an external stimulus while protecting them against the harsh conditions of the gastrointestinal tract. In this regard, Busscher and co-workers explored the encapsulation of probiotic bacteria such as B. breve,21 and L. acidophilus76 within various abiotic coatings, including (i) protamine-assisted SiO2 nanoparticle yolk–shell coating, (ii) alginate hydrogel, and (iii) sod ZIF-8. Then the authors compared the bioprotection properties of B. breve against simulated gastric fluid (SGF††) and a model antibiotic (i.e., tetracycline).21 The SiO2 yolk–shell encapsulation was obtained through the pre-adsorption of a protamine film onto a bacterial cell, followed by exposure to a colloidal suspension of SiO2 NPs. The SiO2 NPs get assembled onto the protamine film, leading to the formation of a silica shell. The protamine film was subsequently internalized into the bacterium to create a void between the bacterial cell surface and the nanoparticle shell. The alginate hydrogel shell was prepared by adding dropwise a PBS suspension of either B. Breve or L. acidophilus and alginate into a CaCl2 solution. The electrostatic interaction between Ca2+ ions and the alginate chains triggers the formation of a three-dimensional hydrogel around the microorganisms. Finally, the ZIF-8 coating was deposited by mixing an aqueous solution of zinc acetate with a premixed aqueous dispersion of probiotics and HmIM. The authors demonstrated that the ζ-potential of non-coated B. Breve cells remained negative across the pH range from 2 (ζ = −0.1 mV) to 9 (ζ = −0.2 mV). However, protamine-assisted SiO2 encapsulation increased the ζ-potential over the entire pH range. For example, at pH = 2, the reported ζ-potential was ζ = 8 mV, whereas at pH = 9, the ζ-potential −18 mV. Interestingly, when using ZIF-8 as a cell exoskeleton, the B. breve ζ-potential only changed in the pH range from 7 to 9 (see Table 1). The ζ-potentials B. Breve cells encased within the alginate-hydrogel were not reported. The elemental surface composition of the cell@shell systems was analysed by XPS. The marked differences in the elemental composition of free probiotics and the cell@shell composites indicated the successful encapsulation of B. Breve and L. acidophilus within the SiO2, alginate, and ZIF-8 abiotic coatings. Then, the authors evaluated, for each abiotic coating, how the encapsulation process affected the viability of the encapsulated cells (B. breve and L. acidophilus). The number of viable cells before and after the encapsulation was determined by the colony-forming unit technique (CFU). The cell viability assay showed that, on agar plates, the growth of B. breve cells coated with SiO2 and alginate hydrogel shells had a cell death comparable to the non-coated cells (control). However, on agar plates, the cells coated within ZIF-8 showed slightly decreased bacterial viability, indicating an enhanced cytotoxic effect of the MOF coating. To assess potential cell wall damage produced by the different abiotic coatings, the authors used the SYTO9/propidium iodide staining test (BacLight Bacterial Viability Kit). In this experiment, microorganisms with cell wall damage present red fluorescence, while those without cell wall damage exhibit green fluorescence. This study concluded that neither the protamine-assisted SiO2 coating nor the alginate hydrogel damaged the cell wall. However, the formation of a ZIF-8 coating induced cell wall damage. The authors hypothesized that this might be attributed to the strong interaction between the bacterial surface proteins and the zinc cations. Subsequently, the authors determined the protection offered by the three different abiotic coatings against SGF and the antibiotic tetracycline. Thus, to mimic the conditions encountered by the probiotics on their way to an intestinal infection site, the same number of uncoated cells and the three different cell@shell composites were suspended in SGF medium at pH = 2. Then, the exposed microorganisms were collected by centrifugation and analyzed by the CFU test. L. acidophilus encapsulated within ZIF-8 and alginate-based shells exhibited around 75% viability upon being exposed to SGF. By contrast, the cell viability dropped to 50% for the cells encased within the SiO2 yolk–shell. The results obtained from B. breve composites indicated that only the alginate-based coating provided effective protection to the cells against SGF. Similarly, the coated and non-coated cells were cultured in a modified medium supplemented with a model antibiotic (i.e., tetracycline), and then the L. acidophilus and B. breve cells were plated on agar for CFU counting. The results obtained from L. acidophilus@shell composites indicated that none of the abiotic coatings protected against negatively charged tetracycline. Nevertheless, cell viability assay of B. breve composites exposed to the model antibiotic indicates that only the alginate-based coating provides full protection against tetracycline. Finally, the authors performed in vitro experiments to determine the therapeutic effect of the coated probiotics against pathogenic E. coli adhered to intestinal epithelial layers. This experiment indicated that B. breve@alginate operated synergistically with tetracycline in protecting intestinal epithelial layers against tetracycline-resistant E. coli.
In combination with probiotic bacteria, researchers are investigating the delivery of enzymes (e.g. lipase) as a therapeutic strategy designed to enhance digestive efficiency, boost the survival and efficacy of the probiotics, and improve metabolic health. In 2024, Qi and co-workers reported the engineering of E. coli with a ZIF-8 exoskeleton for long-term oral delivery of lipase.176 The ZIF-8 coating was formed by first mixing an aqueous dispersion of E. coli with lipase, followed by sequential addition of HmIM and Zn(NO3)2 under stirring for 10 min; the resulting E. coli@ZIF-8 composites were then recovered by centrifugation and washed with deionized water. Transmission electron microscopy (TEM) confirmed the formation of a continuous ZIF-8 shell surrounding individual E. coli cells without altering their rod-shaped morphology. Confocal laser scanning fluorescence microscopy, combining DAPI/SYTOX Green live–dead staining with Cy5-lipase imaging, showed homogeneous distribution of lipase within the ZIF-8 exoskeleton and demonstrated that ∼10.3% of bacteria remained viable after coating. IR spectroscopy revealed a P–O stretching band shift from 1080 cm−1 to 1146 cm−1, consistent with coordination between Zn2+ and phosphates on the bacterial surface, while PXRD patterns matched the sod ZIF-8 topology. In simulated gastric fluid (SGF, pH ∼ 2.0) and simulated intestinal fluid (SIF), the encapsulated lipase retained 8.5% and 23.4% of its activity, respectively, whereas free lipase activity dropped to <5% in SGF and was severely impaired in SIF, indicating that the ZIF-8 shell conferred protection against harsh gastrointestinal conditions. Upon oral gavage of Cy5-labeled E. coli@ZIF-8 in BALB/c mice, in vivo fluorescence imaging demonstrated pronounced retention in the gastrointestinal tract, as lipase@ZIF-8 persisted for ∼4 h, and E. coli&lipase@ZIF-8 was detectable up to 48 h post-administration. Finally, histological analysis of major organs (heart, liver, spleen, lung, and kidney) as well as stomach, small intestine, and large intestine after 48 h of exposure showed intact tissue architecture and no signs of inflammation or damage, confirming the biocompatibility of the ZIF-8-engineered E. coli platform.
Bioremediation
E. coli@ZIF-8 composites can also be included in bio-remediation applications. For instance, in 2023, Ghasemi et al.177 synthesized rod-shaped E. coli@ZIF-8 via biomimetic mineralization, mixing an aqueous suspension of E. coli with HmIM and Zn(OAc)2·2H2O followed by centrifugation and drying, which yielded ∼3 µm rods mirroring the bacterial template. SEM and PXRD analyses confirmed the homogeneous coating with the sod ZIF-8 topology. These E. coli@ZIF-8 particles were then incorporated (40 wt%) into polyacrylonitrile (PAN) nanofibers via electrospinning (10 wt% PAN in DMF, 20 kV, 15 cm), producing E. coli@ZIF-8@PAN composites. The resultant material was then employed for thin-film microextraction (TFME) of benzoylurea insecticides (hexaflumuron and teflubenzuron). The performance of the synthesized E. coli@ZIF-8@PAN composite mat was compared with bare PAN and ZIF-8@PAN fiber mats by using HPLC-UV to evaluate their extraction efficiency. The results indicated that E. coli@ZIF-8@PAN fibers showed better extraction than the other fiber mats. This result was attributed to the increased surface area and the variety of functional groups obtained from the inclusion of the biocomposite into PAN fibers.More recently, Liu et al.178 reported the biomimetic mineralization of B. subtilis ZL09–26 using ZIF-8 and the green modifier citric acid (CA), forming a protective shell (ZIF-8-CA) to enhance phenanthrene (PHE) biodegradation. The mineralization was achieved by co-incubating B. subtilis with HmIM and zinc acetate, followed by surface modification with CA. SEM and EDS analyses confirmed the formation of uniform ZIF-8 and ZIF-8-CA coatings, with the latter exhibiting a more compact and negatively charged shell.
The encapsulated bacteria demonstrated a significantly improved PHE removal efficiency of 94.1% within 6 days, which was 1.9 times that of non-encapsulated cells. Proteomic and enzymatic analyses revealed that the MOF coating reduced oxidative stress and upregulated key metabolic pathways, including central carbon metabolism and oxidative phosphorylation. Additionally, encapsulated cells retained over 83.31% degradation efficiency after five cycles and showed superior viability and storage stability. These results underscore the utility of ZIF-8 coatings in enhancing microbial resilience and bioremediation efficacy under environmental stress.
Whole-cell vaccines
Recently, studies have demonstrated the applicability of micoorganisms@MOF even though the encased cells were inactivated during the coating process. For instance, Luzuriaga et al.96 reported the encapsulation of E. coli (CFT073) within ZIF-8 to treat urinary tract infections. Developing immunity against bacterial infection can be achieved through the administration of inactivated bacteria or a lysed fraction of dead cells. The whole-cell formulations contain strain-specific proteins that trigger antibody production against pathogenic bacteria. However, whole-cell vaccines often fail to provide long-term immune protection, because they elicit weak immune responses, resulting from the short half-life of whole-cell formulations in the body and surface antigen degradation caused by harsh fixation methods.179 Thus, to enhance the bioprotection of the cell-surface antigens, the authors prepared E. coli@ZIF-8 composites, obtained by mixing a zinc acetate aqueous solution with a premixed aqueous dispersion of cells and HmIM. Both solutions were prepared in saline media (NaCl, 100 mM) to keep bacteria near isotonic conditions. After 20 min of stirring, the resulting composite E. coli@ZIF-8 was recovered by centrifugation. Then, the authors employed the CFU assay to evaluate the viability of the encapsulated cells. This experiment revealed that the ZIF-8 shell formation process itself inactivated bacteria; this effect was explained by overexposure to Zn2+, causing cell death. Nevertheless, the challenge in whole-cell formulation is inactivating the bacteria with minimal damage to the surface epitopes, including membrane proteins and oligosaccharides. Therefore, to demonstrate that the encapsulation process inactivates the bacteria while preserving the native protein conformation, the authors performed an agglutination assay to monitor the binding affinity of bacterial glycoproteins towards carbohydrates. This test demonstrated that the growth of the MOF coating does not significantly influence the surface epitopes. Preservation of natively folded bacterial proteins might result in a stronger immune response against E. coli than traditional inactivation methods. To test this hypothesis, the authors measured antibody titers in mice injected with a saline solution containing different formulations of inactivated E. coli: (i) uncoated and thermally inactivated E. coli, (ii) formalin-fixed E. coli, and (iii) E. coli@ZIF-8 composite. Mice immunized with the E. coli@ZIF-8 composite produced the highest antibody levels, whereas the uncoated and thermally inactivated E. coli formulation induced the lowest antibody production. The authors explained the difference by suggesting that the high temperatures required to inactivate cells cause denaturation of proteinaceous and sugar-based epitopes. By contrast, in the case of the E. coli@ZIF-8 formulation, the ZIF encapsulation helped to prevent protein denaturation. In addition, it provided a depot effect by prolonging the presence of E. coli@ZIF-8 in the tissue compared to the uncoated bacteria.Energy storage
Another attractive application of cells@ZIF-8 composites is for energy storage. Gassensmith and co-workers reported the application of the E. coli@ZIF-8 composite as a soft template for fabricating hierarchical porous carbons employed in energy storage.180 The fabrication of E. coli@ZIF-8 was performed by using biomimetic mineralization conditions. However, the authors stated that, unlike S. cerevisiae, the encapsulation of E. coli required a higher ligand-to-metal ratio to form a homogenous cell coating. The TEM micrographs of the E. coli@ZIF-8 composite revealed the formation of a MOF shell comprised of closely packed nanocrystals. Interestingly, the nitrogen sorption isotherm of the E. coli@ZIF-8 composite, collected at 77 K, shows a hysteresis loop at a P/P0 of 0.45–1.0. The latter is typically associated with the capillary condensation of the adsorbate in mesopores. Then, pore size distribution analysis confirmed the presence of mesopores of ∼60 nm.Mesopore formation was attributed to crystalline defects arising during the rapid MOF nucleation process. Then, the E. coli@ZIF-8 composite underwent a carbonization process to remove the bacterial matter. The TEM images showed that calcinated samples, calc-E. coli@ZIF-8, retained the rod-like morphology from the starting biocomposite. However, unlike the E. coli@ZIF-8, the calc-E. coli@ZIF-8 exhibited reduced contrast in the inner region of rod-like structures. This observation confirmed successful removal of the biological entity, leading to a material with high graphitic carbon content. The gas sorption isotherm and the pore size distribution analysis indicated that calc-E. coli@ZIF-8 material retains the mesoporosity, confirming the formation of hierarchical porous carbon (HPC). Finally, the authors compared the electrochemical properties of the calc-E. coli@ZIF-8 sample against the calc-ZIF-8. The cyclic voltammetry analysis indicated that calc-E. coli@ZIF-8 presented superior electrochemical capacity compared to calc-ZIF-8. Overall, this study demonstrated the potential of cells as templating agents for the preparation of hierarchical porous carbons for energy storage.
Recently, Teng and colleagues used the E. coli@ZIF-8 system for the fabrication of a biological nanoreactor.181 They achieved this by expressing alcohol dehydrogenase (ADH) and glucose dehydrogenase (GDH) within E. coli and then coating the cells with ZIF-8. The ADH&GDH multi-enzyme cascade catalytic system demonstrated high efficiency in asymmetrically reducing ketones to produce chiral alcohols with high enantioselectivities (<99%). The researchers also investigated the recyclability of the E. coli@ZIF-8 system. They found that although catalytic reactivity decreased gradually, possibly due to hydrolysis of the ZIF-8 shell under acidic conditions, the system still retained 80% after four cycles. Similarly, Wang and co-workers reported the encapsulation of two bacterial strains (E. coli/pET28a-world and E. coli/pET28a-ladd2) within an amorphous ZIF-90 shell for the catalytic synthesis of D-phenylacetic acid from L-phenylalanine through a two-step cascade reaction.148 Using E. coli@ZIF-90 as a bioreactor for D-PLA production resulted in a yield of 9.00 g L−1 with a conversion rate of 89.4%. After 7 cycles, the immobilized material retained 43.8% of its relative activity. After 9 days of storage at 4 °C, the activity of immobilized cells remained above 75%. In contrast, free cells became almost inactive under the same conditions. This demonstrates the successful catalytic conversion of L-phenylalanine to D-PLA without the need for coenzymes or intermediate substances. Furthermore, compared with free cells, the immobilized cells exhibited good stability toward high temperatures, acidity, alkalinity, organic reagents, and metal ions. This work showed that the immobilized cell method has great potential as an industrial production tool for cost-effective D-PLA production.
Biosensing
The fabrication of cell@MOF composites obtained from inactivated microorganisms has been recently studied for biosensing applications. As an example, Xiang and co-workers reported the use of microorganism@UiO-66-NH2 as a diagnostic probe for the detection of colorectal cancer.182 Specifically, three different microorganisms were used for the cell@shell preparation, including (i) S. cerevisiae, (ii) E. coli (DH5a), and (iii) Synechocystis sp. PCC 6803 (PCC 6803). These microorganisms were inactivated using formaldehyde treatment before the MOF coating. Then, the dormant cells were suspended in a solution of the MOF precursors (Zr(OnPr)4 and H2BDC-NH2) for 18 h. The coated cells were collected by centrifugation and dried in an oven at 60 °C for 12 h. The SEM images collected from the three different cell@MOF composites showed the presence of a uniform abiotic coating. The structural characterization of this artificial coating confirmed the formation of porous UiO-66-NH2. The high surface area of this MOF resulted in advantages for immobilizing biologically relevant molecules, as the Zr4+ ions located on the outer surface of the MOF shell acted as multiple binding sites for protein immobilization. Following this approach, the authors adsorbed streptavidin (SA) onto the outer surface of the MOF shell. Then, the biotinylated-capture probes were immobilized through the strong SA–biotin non-covalent interaction. Using cytometry-based analysis technology, the functionalized cell@MOF composites were used to detect three different miRNA biomarkers (miRNA-21, miRNA-17, and miRNA-182) for colorectal cancer. This study indicated that due to the large amount of protein immobilization on cell@MOF composites, miRNA detection sensitivity could be enhanced to the picomolar range. The reported limit of detection (LOD) values for miRNA-21, miRNA-182, and miRNA-17 biomarkers are 0.75, 0.30, and 0.25 pM, respectively.Biocatalysis
Chen and co-workers developed protocols to co-immobilize enzymes (i.e., inulinase and lipase) and cells (e.g., S. cerevisiae) or bacteria (e.g., E. coli) into a series of parent COFs (i.e., NKCOF-98, COF-42-B, and NKCOF-141) to produce cascade biocatalysts with high efficiency, stability, and recyclability (Fig. 7a and b).44 The COFs investigated in this manuscript are based on 1,3,5-triformylbenzene and derivatives of terephthalohydrazide that differ in the presence of hydrophilic (NKCOF-98), hydrophobic (COF-42-B), or amphiphilic (NKCOF-141) functionalization. The authors showed that coating with NKCOF-98 did not result in effective surface coverage, coating with COF-42-B was non-uniform, and only NKCOF-141 produced homogeneously coated cells. The authors hypothesized that, since cell walls usually exhibit amphiphilic features, the amphiphilic nature of the NKCOF-141 ligand facilitated the homogenous growth of the COF on the cell surface. The authors focused their attention on NKCOF-141 for further biocatalytic tests. In particular, by integrating inulinase in E. coli cells expressing D-allulose 3-epimerase coated with NKCOF-141, the authors developed a flow reaction apparatus for the continuous production of D-allulose. The performance of these innovative biocatalysts (e.g., inulinase/E. coli@NKCOF-141: productivity of D-allulose of 161.3 g L−1 day−1, with >76% initial catalytic efficiency retained after 2 weeks of continuous reaction) showed that the co-immobilization of enzymes and cells into COFs could lead to innovative and customizable platforms for biocatalysis, favoring the development of enzyme–cell cascade production of high value products.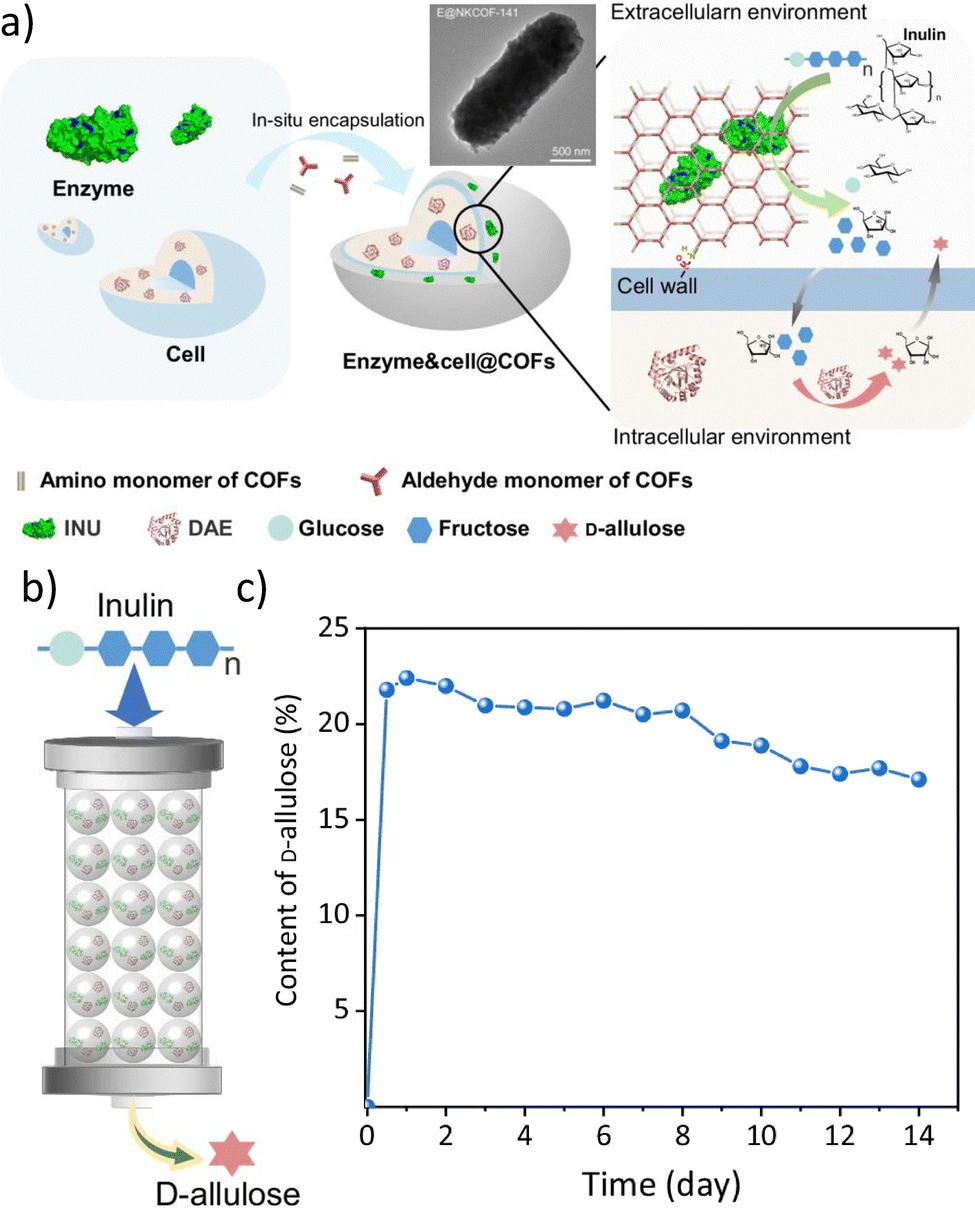 | ||
| Fig. 7 Pioneering example of the co-encapsulation of cells and enzymes within COFs. Schematic diagram of the in situ assembly approach of enzyme&cell@COFs and SEM micrograph of a representative example (a). Schematic diagram of the continuous-flow reaction for the production of D-allulose from inulin and E. coli/D-allulose 3-epimerase co-encapsulated in NKCOF-141 (b). Time-dependent content D-allulose for the continuous-flow reaction at room temperature (30 °C) and 0.1 mL min−1 (c). Adapted with permission from ref. 44 (licensed under CC BY 4.0, https://creativecommons.org/licenses/by/4.0/). | ||
The cells@MOF composites have been recently investigated to boost the performance of strictly anaerobic bacteria for photocatalysis, in particular for artificial photosynthesis applications. One example is M. thermoacetica: this bacterium employs solar energy and CO2 as the only carbon source to produce acetate, which is attractive for carbon remediation applications. However, M. thermoacetica exhibits high susceptibility to O2 and reactive oxygen species (ROS), reducing its performance as a photocatalytic agent.41,183 To tackle this drawback, Yang, Yaghi, and co-workers reported the use of a pre-synthesized Zr-based MOF (Zr6O4(OH)4(BTB)2(OH)6(H2O)6) to coat M. thermoacetica cells and exploit the ROS-scavenging properties of the Zr-based coating.98 The authors tested the performance of M. thermoacetica and M. thermoacetica@MOF in the photocatalytic conversion of CO2. This study revealed that the bare cells could only fix CO2 within the first day of reaction since the accumulation of ROS and O2 by-products caused cellular damage. By contrast, the M. thermoacetica@MOF system remained photo-catalytically active for 2.5 days. Such results showcase the suitability of cell@MOF composites to enhance the bioproduction of value-added chemicals.
MOF-based coatings for cell-like systems
In addition to investigating the encapsulation of living cells, inspired by nature, researchers are engaged in developing synthetic cell-like structures or systems to achieve analogous biological functions in vitro. The capacity of cells to perform chemical reactions with high efficiency and selectivity is attributed to the spatial organization of cell-restricted reaction media and cellular components, which enables the attainment of high local concentrations and spatially directed transport of cellular components.184 Consequently, the fabrication of micro/nanoreactors (MNRs) that emulate the internal configuration of cells is a highly active area of biomimetic study (Fig. 8). Such reactors can be regarded as highly functional material units that can function as biosensors,185 therapeutics,186 and especially in biocatalysis187 either independently or in conjunction with other material matrices. One of the most crucial elements of the reactor is the shell, which can also be defined as the exoskeleton. The outer coating plays a dual role in the reactor, providing robust protection against external factors while regulating the transport of substances and molecules. This phenomenon enables these reactors to perform specific biological functions.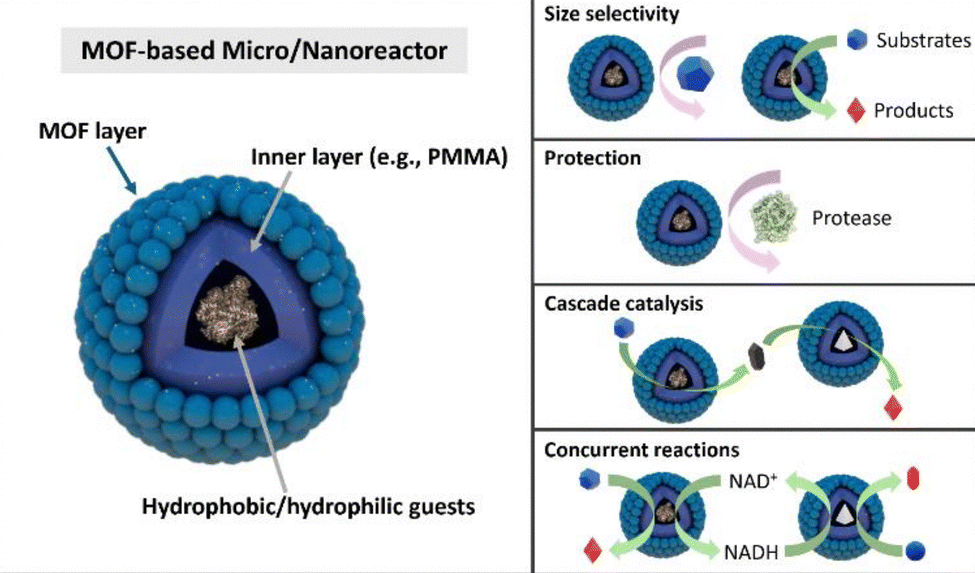 | ||
| Fig. 8 Schematic illustration of MOF-based MNRs. MOF-based MNRs offer several advantages, including size-selection capability for substrates, protection of internal guests from external environmental factors, and the capacity to construct cascade or concurrent catalytic reactions using MNRs with different catalysts as guests. | ||
At the outset, liposomes based on phospholipid bilayer membranes were the most prevalent type of coating used for cell-like reactors.188 This is because liposomes most closely resemble the phospholipid bilayer structure of cell membranes in nature, affording them the potential to exhibit states and functions analogous to those observed in living cells. In addition to liposomes, MNRs based on polymersomes,189,190 colloidosomes,190 and proteinosomes191 have also been developed. Nevertheless, these classes of MNRs continue to exhibit shortcomings, including suboptimal membrane permeability, limitations imposed by osmotic pressure, diminished mechanical stability, and inadequate modulation of small molecules. The advent of organic and hybrid framework materials presents an additional avenue for engineering cell-like MNRs, distinguished by selective molecular diffusion and enhanced protection. As previously stated, MOF materials are notable for their porousness, tunable synthesis, adaptable modification, and extensive diversity, which collectively position them as competitive tools in a multitude of fields. As with the encapsulation of living cells, MOF shells of MNRs can provide effective enhancement of stability, ensure space for molecular reactions, and increase the functional versatility of the reactor (Fig. 8).192 Furthermore, the tunable organic and metal components of the MOF provide a rich chemical microenvironment conducive to reactor functionality.193
Pickering emulsion-based coating strategy
A fundamental attribute of living cells is compartmentalization, which safeguards the integrity of biological processes from external influences. It follows that the majority of cell-like MNRs also exhibit a well-defined compartmentalized structure. To achieve this structural feature, these MNRs are typically encapsulated using a Pickering emulsion-based strategy, which can be described as the self-assembly of MOFs as shells around emulsion droplets.194 Enzymes with dimensions larger than the MOF micropore can be encapsulated, and the dynamic environment facilitates the transport of biomolecules within the compartment.187 In addition, the Pickering emulsion strategy permits the encapsulation of guest molecules with diverse properties in oil-in-water (O/W) systems or water-in-oil (W/O) systems.195 For instance, in W/O systems, the construction of hollow composite microspheres called MOF capsules (MOF-Cs) is achieved through the self-assembly of hydrophobic MOF nanoparticles at the water–oil interface.194 The structural stability of the capsules is enhanced through the precipitation of polymers. Xu et al. demonstrated that such MOF-Cs possess a MOF layer regulating molecular migration, a hollow structure facilitating a confined reaction space, and an encapsulation capability providing catalytically active sites.187 Additionally, these characteristics enable MOF-Cs to encapsulate incompatible substances and facilitate mass diffusion for tandem reactions.187 Therefore, MOF-Cs can be considered analogous to cells in that they can occupy different regions to compartmentalize enzymes or molecules that might otherwise interfere with each other, yet still allow the free diffusion of external molecules necessary for reactions.Despite the exceptional functionality of some hydrophobic molecules, such as organic dyes and organic catalysts, the challenge of dispersing them in aqueous solutions due to their hydrophobic nature can impede their practical applications. Accordingly, researchers introduced hydrophobic molecules (also known as guests) into the oil phase, subsequently forming stable O/W emulsions through the self-assembly of UiO-66-NH2 NPs at the water–oil interface. The hydrophobic guests were encapsulated within the MOF-Cs, followed by PMMA deposition.196 The successful encapsulation of a hydrophobic dye, Nile red, in MOF-Cs resulted in an improvement in energy transfer and promoted size-selective catalysis.196 Therefore, O/W systems are relevant for the utilization of hydrophobic molecules.
Recently, researchers have aimed at expanding single-chambered MOF-Cs into multi-compartmental MOF microreactors, structures much closer to living cells that can catalyze cascade reactions. Tian et al. prepared hierarchically multi-compartmental MOF microreactors through a general Pickering double emulsion-based interfacial synthesis method (Fig. 9a).197 The stabilized Pickering double emulsion (oil-in-water-in-oil double emulsion, O/W/O) can be employed as a growth-oriented template for the formation of a crystalline MOF structure in a large liquid–liquid interfacial region, thereby creating dense MOF layers. The double emulsion system is influenced by many forces, including van der Waals forces between droplet surfaces, intrinsic migratory interactions of droplets generated by interfacial tension, and interactions between metal nodes and organic linkers.198,199 These forces collectively influence the formation of the MOF shell. Accordingly, the interior microstructure of multi-compartmental MOF microreactors can be modified by adjusting the volume fraction of O/W droplets.197 The results demonstrate that the Pickering double emulsion-based synthesis method exhibits robust scalability,197 which will further accelerate rapid development and application of MOF-based MNRs. The structure of MOF-encapsulated MNRs is approaching a configuration that closely resembles the overall structure of living cells. MOFs serve as both a potential outer shell for the reactor and a pivotal element for internal partitioning, offering a broader range of possibilities for MOF-based cell-like reactors and artificial cells.
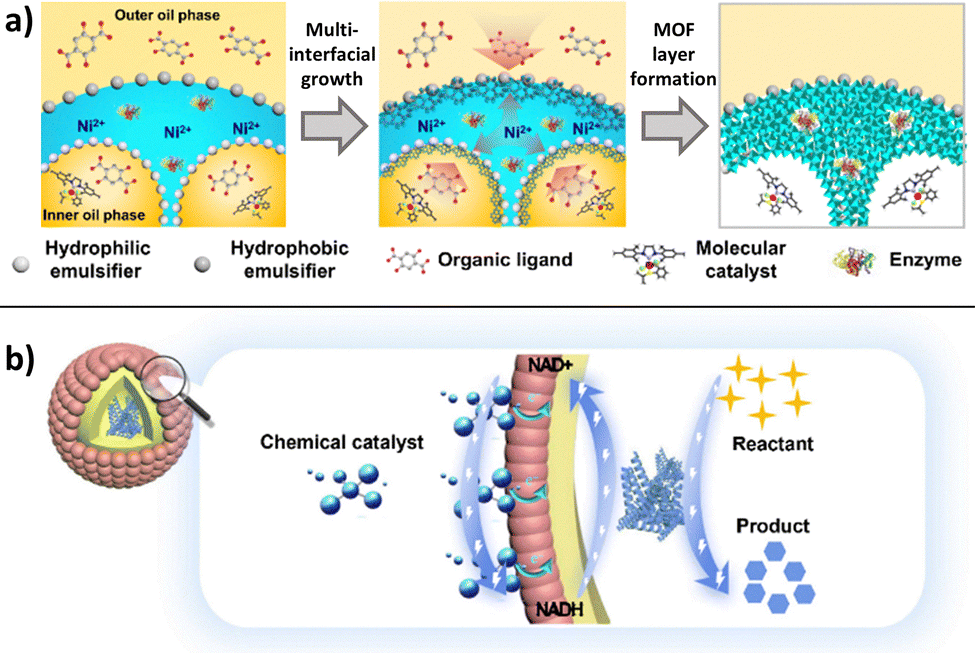 | ||
| Fig. 9 Schematic illustration of (a) the construction of multi-compartmental MOF microreactors via Pickering double emulsions and multi-interfacial growth (adapted with permission from ref. 197, licensed under CC BY 4.0, https://creativecommons.org/licenses/by/4.0/) and (b) and of the electrostatic interaction between MOF-MNRs and chemical catalysts. | ||
Development in cascade catalysis
Cascade catalysis is a pervasive phenomenon in natural systems, whereby multiple enzymes are confined within subcellular compartments. An efficient and highly selective series of catalytic reactions within cells are crucial for maintaining normal metabolic processes and overall organismal functioning. In addition to the requisite reactants and catalysts, the spatial environment in which these biocatalytic reactions occur is particularly important. To be more precise, a confined space can effectively isolate and compartmentalize incompatible substances, while the cascade reactions that take place therein can enhance effective communication between catalysts by facilitating the diffusion of intermediates.200 In light of this natural phenomenon, MOF-Cs have been employed to serve as cell-like structures, encapsulating enzymes within the capsule. The outer MOF shell assumes a cell membrane-like function, providing protection and size-selectivity, thus ensuring that the specific substrate enters the capsule to undergo the reaction. The capacity for protection and size selection is attributable to the hierarchical pore structure of MOF-Cs, which is achieved through the self-assembly of MOF nanoparticles (MOF NPs).187 In particular, the intrinsic microporous structure of MOF NPs regulates molecular transport, while the hollow structure inside the capsule allows for the loading of different enzymes. For example, Xu et al. synthesized MOF-Cs in W/O emulsions comprising UiO-66-NH2 NPs as components, encapsulating GOx within them (GOX@MOF-Cs). Their findings demonstrated that GOx@MOF-Cs exhibited efficient glucose oxidation without being affected by proteases in the external environment.187 Consequently, MOF-Cs exhibit the ability to transport, encapsulate, and compartmentalize.Future opportunities and challenges
Research on living cells encapsulated in MOF, COF, and HOF exoskeletons has made considerable progress, but several key challenges and questions remain unaddressed. As the field of emerging porous materials is more advanced for MOFs, we will predominantly refer to this class. Below, we highlight several promising directions, ranging from advanced therapeutics to synthetic biology, while also highlighting roadblocks that must be overcome to enable wide-scale translation.Biomedical applications
Immunotherapy. Cancer immunotherapy has emerged as a powerful clinical treatment, utilizing specialized cells (e.g., T-lymphocytes, NK cells, and dendritic cells) and bacterial-mediated cancer therapy (BMCT) to target malignancies. While BMCT harnesses the inherent tumor-targeting ability of specific bacterial strains, clinical translation has been historically hindered by risks of systemic toxicity, uncontrolled bacterial proliferation, and insufficient drug loading. In this context, cell encapsulation within porous frameworks (MOFs, COFs, and HOFs) offers a transformative solution. Framework shells, particularly zinc imidazolate frameworks (ZIFs), act as cloaking devices that physically restrict pathogen-associated molecular patterns (PAMPs, e.g. lipopolysaccharides) during transport. This shielding delays immune recognition and reduces off-target inflammation until the vehicle reaches the tumor microenvironment.
Opportunities in synergistic delivery. The most significant opportunity in this domain lies in the transition from simple cell carriers to multifunctional combination therapies. Beyond protecting the cargo, porous frameworks can serve as dual-delivery systems, co-encapsulating small-molecule chemotherapeutics or immunomodulators alongside the cellular payload.203 These systems are designed to degrade in response to tumor microenvironment specific stimuli (e.g., acidic pH and high ATP concentrations),38,56,139 ensuring the simultaneous, localized release of both the therapeutic agent and the biological cargo. Furthermore, integrating genetically engineered bacteria capable of secreting anticancer proteins within these confined frameworks represents a high-potential avenue for increasing drug-loading capacity and advancing toward more potent combination therapies.
Engineered cell interactions. In tissue engineering, controlling spatial organization and communication among cells is crucial.204–206 Porous framework exoskeletons can be designed to promote or inhibit specific cell−cell interactions by selectively allowing growth factors, cytokines, or other signaling molecules to diffuse through the framework material. Such coatings might also help cells endure mechanical stresses or nutrient fluctuations common in early tissue scaffolding stages.
Integration with biopolymer scaffolds. A major advance would be the integration of encapsulated cells into larger 3D scaffolds, where biocompatibility and structural stability must be maintained for long periods (e.g., weeks and months). Future work on hybrid materials, combining organic polymers (e.g., collagen and alginate) with MOF/COF/HOF, could yield composite scaffolds that simultaneously offer mechanical support and molecular sieving. Success in this goal may offer new ways to engineer tissues with higher viability and functionality, yet it will require new characterization methods (e.g., in situ imaging of shell integrity) and standardized viability assays in complex, multi-component systems.
Concerns are also related to the antibiotic resistance of bacteria-based therapeutics. ZIF-encapsulated bacteria often exhibit improved tolerance to antibiotics97 and UV irradiation.109 As a result, the use of antibiotic-resistant plasmids—which raise safety and regulatory issues—may be unnecessary, and UV sterilization could be employed to enhance manufacturing safety without compromising bacterial viability. In immunotherapy applications, this would simplify clinical translation by lowering the risks of antibiotic resistance and facilitating Good Manufacturing Practice (GMP) protocols.
Industrial biotechnology
Porous shell design, scale-up, reproducibility, and manufacturing standards
Conclusions and outlook
In summary, the functional properties of MOF-, COF-, and HOF-based coatings offer unprecedented opportunities for cell applications ranging from separation technology to medical treatments. By engineering framework materials with controlled pore size distribution, pore chemistry, stimulus-responsive behavior, and the ability to host and co-deliver functional agents, researchers can design novel microbial therapies and engineered tissues. Yet, significant scientific and regulatory hurdles remain, including reproducibility challenges, robust assessment of viability, finely tuned “on-demand” degradability, large-scale process optimization, and safety profiling. Through cross-disciplinary collaborations among chemists, materials scientists, bioengineers, and clinical researchers, these challenges could be addressed, enabling cells coated with porous shells to overcome current limitations of cell-based technologies in industrial biocatalysis and biomedicine.Author contributions
FC and MVH: data curation, visualization, writing – original draft; AEM, DK, QW, RK, LYC, JG, FKS, CD: writing – original draft; PF: conceptualization, visualization, funding acquisition, project administration, writing – original draft; FC and PF: writing – review and editing.Conflicts of interest
There are no conflicts to declare.Data availability
No primary research results, software or code have been included and no new data were generated or analysed as part of this review.Supplementary information: video S1 visualizes the role of artificial cell coatings as molecular sieves. See DOI: https://doi.org/10.1039/d4cs00071d.
Acknowledgements
We acknowledge the PMWS Lead Project (LP-03) and TU Graz for support.Notes and references
- J. Guo, Y. Yu, W. Zhu, R. E. Serda, S. Franco, L. Wang, Q. Lei, J. O. Agola, A. Noureddine, E. Ploetz, S. Wuttke and C. J. Brinker, Adv. Funct. Mater., 2021, 31, 2005935 CrossRef CAS.
- S. Yao, B. Jin, Z. Liu, C. Shao, R. Zhao, X. Wang and R. Tang, Adv. Mater., 2017, 29, 1605903 CrossRef PubMed.
- I. Holzmeister, M. Schamel, J. Groll, U. Gbureck and E. Vorndran, Acta Biomater., 2018, 74, 17–35 CrossRef CAS PubMed.
- H. K. Baca, C. Ashley, E. Carnes, D. Lopez, J. Flemming, D. Dunphy, S. Singh, Z. Chen, N. Liu, H. Fan, G. P. López, S. M. Brozik, M. Werner-Washburne and C. J. Brinker, Science, 2006, 313, 337–341 CrossRef CAS PubMed.
- W. A. M. Wolken, J. Tramper and M. J. van der Werf, Trends Biotechnol., 2003, 21, 338–345 CrossRef CAS PubMed.
- E. I. Tocheva, D. R. Ortega and G. J. Jensen, Nat. Rev. Microbiol., 2016, 14, 535–542 CrossRef PubMed.
- M. Y. Galperin, S. L. Mekhedov, P. Puigbo, S. Smirnov, Y. I. Wolf and D. J. Rigden, Environ. Microbiol., 2012, 14, 2870–2890 CrossRef CAS PubMed.
- P. Setlow, Cell, 2008, 135, 410–412 CrossRef CAS PubMed.
- M. Cuéllar-Cruz, Prog. Cryst. Growth Charact. Mater., 2017, 63, 94–103 CrossRef.
- C. S. Kovacs, C. Chaussain, P. Osdoby, M. L. Brandi, B. Clarke and R. V. Thakker, Nat. Rev. Endocrinol., 2021, 17, 336–349 CrossRef CAS PubMed.
- D. Hong, M. Park, S. H. Yang, J. Lee, Y.-G. Kim and I. S. Choi, Trends Biotechnol., 2013, 31, 442–447 CrossRef CAS PubMed.
- K. Liang, J. J. Richardson, C. J. Doonan, X. Mulet, Y. Ju, J. Cui, F. Caruso and P. Falcaro, Angew. Chem., Int. Ed., 2017, 56, 8510–8515 CrossRef CAS PubMed.
- R. Kempaiah, S. Salgado, W. L. Chung and V. Maheshwari, Chem. Commun., 2011, 47, 11480 RSC.
- G. Wang, L. Wang, P. Liu, Y. Yan, X. Xu and R. Tang, ChemBioChem, 2010, 11, 2368–2373 CrossRef CAS PubMed.
- C. Sicard, M. Perullini, C. Spedalieri, T. Coradin, R. Brayner, J. Livage, M. Jobbágy and S. A. Bilmes, Chem. Mater., 2011, 23, 1374–1378 CrossRef CAS.
- L. Gan, M. de J. Velásquez-Hernández, A. Emmerstorfer-Augustin, P. Wied, H. Wolinski, S. D. Zilio, M. Solomon, W. Liang, C. Doonan and P. Falcaro, Chem. Commun., 2022, 58, 10004–10007 RSC.
- S. H. Yang, D. Hong, J. Lee, E. H. Ko and I. S. Choi, Small, 2013, 9, 178–186 CrossRef CAS PubMed.
- W. Youn, J. Y. Kim, J. Park, N. Kim, H. Choi, H. Cho and I. S. Choi, Adv. Mater., 2020, 32, 1907001 Search PubMed.
- J. H. Park, K. Kim, J. Lee, J. Y. Choi, D. Hong, S. H. Yang, F. Caruso, Y. Lee and I. S. Choi, Angew. Chem., Int. Ed., 2014, 53, 12420–12425 CrossRef CAS PubMed.
- Q. Lei, J. Guo, F. Kong, J. Cao, L. Wang, W. Zhu and C. J. Brinker, J. Am. Chem. Soc., 2021, 143, 6305–6322 CrossRef CAS PubMed.
- L. Yuan, H. Wei, X.-Y. Yang, W. Geng, B. W. Peterson, H. C. Van Der Mei and H. J. Busscher, ACS Appl. Mater. Interfaces, 2021, 13, 15973–15982 CrossRef CAS PubMed.
- S. H. Yang, K.-B. Lee, B. Kong, J.-H. Kim, H.-S. Kim and I. Choi, Angew. Chem., Int. Ed., 2009, 48, 9160–9163 Search PubMed.
- S. H. Yang, E. H. Ko and I. S. Choi, Langmuir, 2012, 28, 2151–2155 CrossRef CAS PubMed.
- Z. Cao and J. Liu, Nat. Protoc., 2024, 19, 3162–3190 CrossRef CAS PubMed.
- Z. Cao, S. Cheng, X. Wang, Y. Pang and J. Liu, Nat. Commun., 2019, 10, 3452 CrossRef PubMed.
- Z. Cao and J. Liu, Acc. Mater. Res., 2024, 5, 872–883 CrossRef CAS.
- P. Feng, Z. Cao, X. Wang, J. Li and J. Liu, Adv. Mater., 2020, 32, 2002406 CrossRef CAS PubMed.
- H. Luo, F. Wu, X. Wang, S. Lin, M. Zhang, Z. Cao and J. Liu, Mater. Today, 2023, 62, 98–110 Search PubMed.
- H. Luo, J. Xie, X. Su, P. Wang, H. Chen, X. Kuang and J. Liu, Sci. China Mater., 2024, 67, 3833–3848 CrossRef CAS.
- W. Yu, H. Luo, B. Han, S. Lin, Q. Li, R. Xue, H. Tang, X. Jia, L. Wang and J. Liu, Nat. Commun., 2025, 16, 7691 CrossRef CAS PubMed.
- M. de, J. Velásquez-Hernández, M. Linares-Moreau, E. Astria, F. Carraro, M. Z. Alyami, N. M. Khashab, C. J. Sumby, C. J. Doonan and P. Falcaro, Coord. Chem. Rev., 2021, 429, 213651 CrossRef.
- O. M. Yaghi, Nano Lett., 2020, 20, 8432–8434 CrossRef CAS PubMed.
- D. Yu, H. Zhang, Z. Liu, C. Liu, X. Du, J. Ren and X. Qu, Angew. Chem., Int. Ed., 2022, 61, e202201485 CrossRef CAS PubMed.
- R. Riccò, W. Liang, S. Li, J. J. Gassensmith, F. Caruso, C. Doonan and P. Falcaro, ACS Nano, 2018, 12, 13–23 CrossRef PubMed.
- H. Furukawa, K. E. Cordova, M. O’Keeffe and O. M. Yaghi, Science, 2013, 341, 1230444 CrossRef PubMed.
- X. Song, Y. Wang, C. Wang, D. Wang, G. Zhuang, K. O. Kirlikovali, P. Li and O. K. Farha, J. Am. Chem. Soc., 2022, 144, 10663–10687 CrossRef CAS PubMed.
- P. Li, M. R. Ryder and J. F. Stoddart, Acc. Mater. Res., 2020, 1, 77–87 Search PubMed.
- S. Kandambeth, K. Dey and R. Banerjee, J. Am. Chem. Soc., 2019, 141, 1807–1822 CrossRef CAS PubMed.
- Z. Sun, R. Hübner, J. Li and C. Wu, Nat. Commun., 2022, 13, 3142 Search PubMed.
- M. H. O. Roshid, M. Moraskie, G. O’Connor, E. Dikici, J.-M. Zingg, S. Deo, L. G. Bachas and S. Daunert, Microchem. J., 2023, 193, 109088 CrossRef CAS PubMed.
- K. K. Sakimoto, A. B. Wong and P. Yang, Science, 2016, 351, 74–77 Search PubMed.
- A. K. Singh, R. Awasthi and R. Malviya, J. Controlled Release, 2023, 354, 439–452 CrossRef CAS PubMed.
- X. Lou, Z. Chen, Z. He, M. Sun and J. Sun, Nano-Micro Lett., 2021, 13, 37 CrossRef PubMed.
- D. Zheng, Y. Zheng, J. Tan, Z. Zhang, H. Huang and Y. Chen, Nat. Commun., 2024, 15, 5510 CrossRef CAS PubMed.
- P. Falcaro, F. Normandin, M. Takahashi, P. Scopece, H. Amenitsch, S. Costacurta, C. M. Doherty, J. S. Laird, M. D. H. Lay, F. Lisi, A. J. Hill and D. Buso, Adv. Mater., 2011, 23, 3901–3906 CrossRef CAS PubMed.
- C. M. Doherty, E. Knystautas, D. Buso, L. Villanova, K. Konstas, A. J. Hill, M. Takahashi and P. Falcaro, J. Mater. Chem., 2012, 22, 11470 RSC.
- E. Zanchetta, L. Malfatti, R. Ricco, M. J. Styles, F. Lisi, C. J. Coghlan, C. J. Doonan, A. J. Hill, G. Brusatin and P. Falcaro, Chem. Mater., 2015, 27, 690–699 CrossRef CAS.
- M. J. Van Vleet, T. Weng, X. Li and J. R. Schmidt, Chem. Rev., 2018, 118, 3681–3721 CrossRef CAS PubMed.
- A. Veis and J. R. Dorvee, Calcif. Tissue Int., 2013, 93, 307–315 CrossRef CAS PubMed.
- in The IUPAC Compendium of Chemical Terminology: The Gold Book, ed. V. Gold, International Union of Pure and Applied Chemistry (IUPAC), Research Triangle Park, NC, 4th edn, 2019 Search PubMed.
- J. D. Clogston and A. K. Patri, in Characterization of Nanoparticles Intended for Drug Delivery, ed. S. E. McNeil, Humana Press, Totowa, NJ, 2011, vol. 697, pp. 63–70 Search PubMed.
- N. K. Maddigan, A. Tarzia, D. M. Huang, C. J. Sumby, S. G. Bell, P. Falcaro and C. J. Doonan, Chem. Sci., 2018, 9, 4217–4223 RSC.
- W. Liang, P. Wied, F. Carraro, C. J. Sumby, B. Nidetzky, C.-K. Tsung, P. Falcaro and C. J. Doonan, Chem. Rev., 2021, 121, 1077–1129 Search PubMed.
- K. Alt, F. Carraro, E. Jap, M. Linares-Moreau, R. Riccò, M. Righetto, M. Bogar, H. Amenitsch, R. A. Hashad, C. Doonan, C. E. Hagemeyer and P. Falcaro, Adv. Mater., 2022, 34, 2106607 CrossRef CAS PubMed.
- E. Astria, M. Thonhofer, R. Ricco, W. Liang, A. Chemelli, A. Tarzia, K. Alt, C. E. Hagemeyer, J. Rattenberger, H. Schroettner, T. Wrodnigg, H. Amenitsch, D. M. Huang, C. J. Doonan and P. Falcaro, Mater. Horiz., 2019, 6, 969–977 RSC.
- F. Carraro, M. de, J. Velásquez-Hernández, E. Astria, W. Liang, L. Twight, C. Parise, M. Ge, Z. Huang, R. Ricco, X. Zou, L. Villanova, C. O. Kappe, C. Doonan and P. Falcaro, Chem. Sci., 2020, 11, 3397–3404 RSC.
- F. Carraro, J. D. Williams, M. Linares-Moreau, C. Parise, W. Liang, H. Amenitsch, C. Doonan, C. O. Kappe and P. Falcaro, Angew. Chem., 2020, 132, 8200–8204 CrossRef.
- L. Heinke and C. Wöll, Adv. Mater., 2019, 31, 1806324 CrossRef PubMed.
- P. Falcaro, D. Buso, A. J. Hill and C. M. Doherty, Adv. Mater., 2012, 24, 3153–3168 CrossRef CAS PubMed.
- K. Liang, C. Carbonell, M. J. Styles, R. Ricco, J. Cui, J. J. Richardson, D. Maspoch, F. Caruso and P. Falcaro, Adv. Mater., 2015, 27, 7293–7298 CrossRef CAS PubMed.
- M. R. Hafner, F. Carraro, L. A. Brandner, S. Maniam, G. Grenci, S. Ljubojevic-Holzer, H. Bischof, R. Malli, S. M. Borisov, C. Doonan and P. Falcaro, Chem. Commun., 2020, 56, 12733–12736 RSC.
- R. M. Olabisi, J. Biomed. Mater. Res., 2015, 103, 846–859 CrossRef PubMed.
- J. Lee, J. Choi, J. H. Park, M. Kim, D. Hong, H. Cho, S. H. Yang and I. S. Choi, Angew. Chem., Int. Ed., 2014, 53, 8056–8059 Search PubMed.
- W. Zhu, J. Guo, S. Amini, Y. Ju, J. O. Agola, A. Zimpel, J. Shang, A. Noureddine, F. Caruso, S. Wuttke, J. G. Croissant and C. J. Brinker, Adv. Mater., 2019, 31, 1900545 CrossRef PubMed.
- T. J. Silhavy, D. Kahne and S. Walker, Cold Spring Harbor Perspect. Biol., 2010, 2, a000414 Search PubMed.
- S. I. Miller and N. R. Salama, PLoS Biol., 2018, 16, e2004935 CrossRef PubMed.
- G. Zhang, T. C. Meredith and D. Kahne, Curr. Opin. Microbiol., 2013, 16, 779–785 CrossRef CAS PubMed.
- I. Bononi, V. Balatti, S. Gaeta and M. Tognon, Appl. Environ. Microbiol., 2008, 74, 6470–6472 CrossRef CAS PubMed.
- N. Ruiz and T. J. Silhavy, J. Bacteriol., 2022, 204, e00230 CrossRef PubMed.
- S. Yan, X. Zeng, Y. Wang and B. Liu, Adv. Healthcare Mater., 2020, 9, 2000046 CrossRef CAS PubMed.
- G. Neumann, S. Cornelissen, F. Van Breukelen, S. Hunger, H. Lippold, N. Loffhagen, L. Y. Wick and H. J. Heipieper, Appl. Environ. Microbiol., 2006, 72, 4232–4238 CrossRef CAS PubMed.
- J. Li and L. A. McLandsborough, Int. J. Food Microbiol., 1999, 53, 185–193 CrossRef CAS PubMed.
- D. Montag, M. Frant, H. Horn and K. Liefeith, Biofouling, 2012, 28, 315–327 CrossRef CAS PubMed.
- M. Rohde, Microbiol. Spectrosc., 2019, 7(3), 10 Search PubMed.
- K. Cremin, B. Jones, J. Teahan, G. N. Meloni, D. Perry, C. Zerfass, M. Asally, O. S. Soyer and P. R. Unwin, Anal. Chem., 2020, 92(24), 16024–16032 CrossRef CAS PubMed.
- M. L. F. Murga, G. Font De Valdez and A. E. Disalvo, Cryobiology, 2000, 41, 10–16 CrossRef CAS PubMed.
- J. K. Oh, Y. Yegin, F. Yang, M. Zhang, J. Li, S. Huang, S. V. Verkhoturov, E. A. Schweikert, K. Perez-Lewis, E. A. Scholar, T. M. Taylor, A. Castillo, L. Cisneros-Zevallos, Y. Min and M. Akbulut, Sci. Rep., 2018, 8, 17247 CrossRef PubMed.
- M. C. Van Loosdrecht, J. Lyklema, W. Norde, G. Schraa and A. J. Zehnder, Appl. Environ. Microbiol., 1987, 53, 1898–1901 CrossRef CAS PubMed.
- N. A. R. Gow, J.-P. Latge and C. A. Munro, Microbiol. Spectrosc, 2017, 5(3), 01 Search PubMed.
- A. E. Smith, Z. Zhang, C. R. Thomas, K. E. Moxham and A. P. J. Middelberg, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 9871–9874 CrossRef CAS PubMed.
- D. Kregiel, J. Berlowska and B. Szubzda, J. Ind. Microbiol. Biotechnol., 2012, 39, 1881–1886 CrossRef CAS PubMed.
- L. M. Lavaisse, A. Hollmann, M. A. Nazareno and E. A. Disalvo, Colloids Surf., B, 2019, 174, 63–69 CrossRef CAS PubMed.
- A. Rogowska, P. Pomastowski, M. Złoch, V. Railean-Plugaru, A. Król, K. Rafińska, M. Szultka-Młyńska and B. Buszewski, Sci. Rep., 2018, 8, 7261 CrossRef PubMed.
- in Molecular biology of the cell, ed. B. Alberts, Garland Science, New York, 4th edn, 2002 Search PubMed.
- A. Melelli, O. Arnould, J. Beaugrand and A. Bourmaud, Molecules, 2020, 25, 632 CrossRef CAS PubMed.
- G. Gilliard, E. Huby, S. Cordelier, M. Ongena, S. Dhondt-Cordelier and M. Deleu, Front. Plant Sci., 2021, 12, 749581 CrossRef PubMed.
- T. B. Kinraide, U. Yermiyahu and G. Rytwo, Plant Physiol., 1998, 118, 505–512 CrossRef CAS PubMed.
- P. Durand-Smet, N. Chastrette, A. Guiroy, A. Richert, A. Berne-Dedieu, J. Szecsi, A. Boudaoud, J.-M. Frachisse, M. Bendahmane, O. Hamant and A. Asnacios, Biophys. J., 2014, 107, 2237–2244 CrossRef CAS PubMed.
- O. V. Bondar, D. V. Saifullina, I. I. Shakhmaeva, I. I. Mavlyutova and T. I. Abdullin, Acta Naturae, 2012, 4, 78–81 CrossRef CAS PubMed.
- M. Nishino, I. Matsuzaki, F. Y. Musangile, Y. Takahashi, Y. Iwahashi, K. Warigaya, Y. Kinoshita, F. Kojima and S. Murata, PLoS One, 2020, 15, e0236373 CrossRef CAS PubMed.
- F. Esposito, M. R. Fernandes, R. Lopes, M. Muñoz, C. P. Sabino, M. P. Cunha, K. C. Silva, R. Cayô, W. M. B. S. Martins, A. M. Moreno, T. Knöbl, A. C. Gales and N. Lincopan, J. Clin. Microbiol., 2017, 55, 3454–3465 CrossRef CAS PubMed.
- I. Obi, T. Kakutani, N. Imaizumi, Y. Ichikawa and M. Senda, Plant Cell Physiol., 1990, 31, 1031–1037 Search PubMed.
- M. J. Stoddart, in Mammalian Cell Viability, ed. M. J. Stoddart, Humana Press, Totowa, NJ, 2011, vol. 740, pp. 1–6 Search PubMed.
- M. R. Barer and C. R. Harwood, Advances in Microbial Physiology, Elsevier, 1999, vol. 41, pp. 93–137 Search PubMed.
- H. Wei, X.-Y. Yang, W. Geng, H. C. Van Der Mei and H. J. Busscher, Nanoscale, 2021, 13, 7220–7233 RSC.
- M. A. Luzuriaga, F. C. Herbert, O. R. Brohlin, J. Gadhvi, T. Howlett, A. Shahrivarkevishahi, Y. H. Wijesundara, S. Venkitapathi, K. Veera, R. Ehrman, C. E. Benjamin, S. Popal, M. D. Burton, M. A. Ingersoll, N. J. De Nisco and J. J. Gassensmith, ACS Nano, 2021, 15, 17426–17438 CrossRef CAS PubMed.
- H. Li, A. Kang, B. An, L.-Y. Chou, F.-K. Shieh, C.-K. Tsung and C. Zhong, Mater. Today Bio, 2021, 10, 100097 CrossRef CAS PubMed.
- Z. Ji, H. Zhang, H. Liu, O. M. Yaghi and P. Yang, Proc. Natl. Acad. Sci. U. S. A., 2018, 115, 10582–10587 CrossRef CAS PubMed.
- K. M. McKinnon, CP Immunol., 2018, 120, 5.1.1–5.1.11 Search PubMed.
- L. L.-Y. Chan, K. J. McCulley and S. L. Kessel, in Cell Viability Assays, ed. D. F. Gilbert and O. Friedrich, Springer New York, New York, NY, 2017, vol. 1601, pp. 27–41 Search PubMed.
- in Cell Viability Assays: Methods and Protocols, ed. D. F. Gilbert and O. Friedrich, Springer New York, New York, NY, 2017, vol. 1601 Search PubMed.
- P. Stiefel, S. Schmidt-Emrich, K. Maniura-Weber and Q. Ren, BMC Microbiol., 2015, 15, 36 CrossRef PubMed.
- Lehninger Principles of Biochemistry – ISBN-13 978-1-4292-3414-6, https://www.books-by-isbn.com/1-4292/1429234148-Lehninger-Principles-of-Biochemistry-David-L-Nelson-Michael-M-Cox-1-4292-3414-8.html, (accessed March 3, 2026).
- S. L. Fink and B. T. Cookson, Infect. Immun., 2005, 73, 1907–1916 CrossRef CAS PubMed.
- Z. Darzynkiewicz, S. Bruno, G. Del Bino, W. Gorczyca, M. A. Hotz, P. Lassota and F. Traganos, Cytometry, 1992, 13, 795–808 CrossRef CAS PubMed.
- L. Shi, S. Günther, T. Hübschmann, L. Y. Wick, H. Harms and S. Müller, Cytometry Part A, 2007, 71A, 592–598 CrossRef PubMed.
- M. Rosenberg, N. F. Azevedo and A. Ivask, Sci. Rep., 2019, 9, 6483 CrossRef PubMed.
- A. Permyakova, A. Kakar, J. Bachir, E. Gkaniatsou, B. Haye, N. Menguy, F. Nouar, C. Serre, N. Steunou, T. Coradin, F. M. Fernandes and C. Sicard, ACS Mater. Lett., 2023, 5, 79–84 CrossRef CAS.
- Y. Qin, L. Chen, Y. Cheng, S. Yang, Y. Liu, W. Fan, L. Wang, Q. Wang, L. Zheng and Q. Cao, ACS Appl. Bio Mater., 2020, 3, 3268–3275 CrossRef CAS PubMed.
- T. L. Riss, R. A. Moravec, A. L. Niles, S. Duellman, H. A. Benink, T. J. Worzella and L. Minor, in Assay Guidance Manual, ed. S. Markossian, A. Grossman, M. Arkin, D. Auld, C. Austin, J. Baell, K. Brimacombe, T. D. Y. Chung, N. P. Coussens, J. L. Dahlin, V. Devanarayan, T. L. Foley, M. Glicksman, K. Gorshkov, J. V. Haas, M. D. Hall, S. Hoare, J. Inglese, P. W. Iversen, M. Lal-Nag, Z. Li, J. R. Manro, J. McGee, O. McManus, M. Pearson, T. Riss, P. Saradjian, G. S. Sittampalam, M. Tarselli, O. J. Trask, J. R. Weidner, M. J. Wildey, K. Wilson, M. Xia and X. Xu, Eli Lilly & Company and the National Center for Advancing Translational Sciences, Bethesda (MD), 2004 Search PubMed.
- E. U. Karatop, C. E. Cimenci, A. M. Aksu, E. U. Karatop, C. E. Cimenci and A. M. Aksu, Cytotoxicity – Understanding Cellular Damage and Response, IntechOpen, 2022 Search PubMed.
- K. Präbst, H. Engelhardt, S. Ringgeler and H. Hübner, in Cell Viability Assays, ed. D. F. Gilbert and O. Friedrich, Springer New York, New York, NY, 2017, vol. 1601, pp. 1–17 Search PubMed.
- E. Grela, J. Kozłowska and A. Grabowiecka, Acta Histochem., 2018, 120, 303–311 CrossRef CAS PubMed.
- Q. Chen, S. Tang, Y. Li, Z. Cong, D. Lu, Q. Yang, X. Zhang and S. Wu, ACS Appl. Mater. Interfaces, 2021, 13, 58382–58392 CrossRef CAS PubMed.
- R. Ohtani, K. Kawano, M. Kinoshita, S. Yanaka, H. Watanabe, K. Hirai, S. Futaki, N. Matsumori, H. Uji-i, M. Ohba, K. Kato and S. Hayami, Angew. Chem., Int. Ed., 2020, 59, 17931–17937 CrossRef CAS PubMed.
- L. Ha, K. M. Choi and D.-P. Kim, ACS Appl. Mater. Interfaces, 2021, 13, 18545–18553 CrossRef CAS PubMed.
- S. Neri, E. Mariani, A. Meneghetti, L. Cattini and A. Facchini, Clin. Diagn. Lab. Immunol., 2001, 8, 1131–1135 CrossRef CAS PubMed.
- G. Adam and H. Duncan, Soil Biol. Biochem., 2001, 33, 943–951 CrossRef CAS.
- W. Chen, S. Kong, M. Lu, F. Chen, W. Cai, L. Du, J. Wang and C. Wu, Soft Matter, 2020, 16, 270–275 RSC.
- K. Liang, J. J. Richardson, J. Cui, F. Caruso, C. J. Doonan and P. Falcaro, Adv. Mater., 2016, 28, 7910–7914 CrossRef CAS PubMed.
- H. Arakawa, M. Shiokawa, O. Imamura and M. Maeda, Anal. Biochem., 2003, 314, 206–211 CrossRef CAS PubMed.
- S. Kamiloglu, G. Sari, T. Ozdal and E. Capanoglu, Food Front., 2020, 1, 332–349 CrossRef.
- S. H. Yang, E. H. Ko, Y. H. Jung and I. S. Choi, Angew. Chem., Int. Ed., 2011, 50, 6115–6118 CrossRef CAS PubMed.
- N. Jeon, I. Jeong, E. Cho, I. Choi, J. Lee, E. H. Han, H. J. Lee, P. C. W. Lee and E. Lee, JACS Au, 2023, 3, 154–164 CrossRef CAS PubMed.
- H. Yang, Y. Zhang, L. Zeng, W. Yin, Y. Xu, J. Chen, S. Liu, X. Zou, Z. He and Z. Dai, Small Methods, 2022, 6, 2101391 CrossRef CAS PubMed.
- Y. Jiao, Y. Guo, Y. Fan, R. Wang, X. Li, H. Wu, Z. Meng, X. Yang, Y. Cui, H. Liu, L. Pan, T. Maimaitijuma, J. Zhang, Y. Wang, Y. Cao and T. Zhang, BioMed Res. Int., 2020, 2020, 2846297 CrossRef PubMed.
- Y. Wang, C. Liu, X. Ma, A. Filppula, Y. Cui, J. Ye and H. Zhang, Nanoscale, 2024, 16, 20925–20939 RSC.
- F. Sivandzade, A. Bhalerao and L. Cucullo, Bio-Protocol, 2019, 9, e3128 CrossRef PubMed.
- K. S. Louis and A. C. Siegel, in Mammalian Cell Viability, ed. M. J. Stoddart, Humana Press, Totowa, NJ, 2011, vol. 740, pp. 7–12 Search PubMed.
- A. Kan, D. P. Birnbaum, P. Praveschotinunt and N. S. Joshi, Appl. Environ. Microbiol., 2019, 85, e00434–19 CrossRef CAS PubMed.
- M. Wu, Y. Liao, D. Guo, M. Zhai, D. Xia, Z. Zhang, X. Liu and Y. Huang, RSC Adv., 2024, 14, 14722–14741 RSC.
- H. C. Moon, S. Han, J. Borges, T. Pesqueira, H. Choi, S. Y. Han, H. Cho, J. H. Park, J. F. Mano and I. S. Choi, Soft Matter, 2020, 16, 6063–6071 RSC.
- J. Guo, Y. Ping, H. Ejima, K. Alt, M. Meissner, J. J. Richardson, Y. Yan, K. Peter, D. von Elverfeldt, C. E. Hagemeyer and F. Caruso, Angew. Chem., Int. Ed., 2014, 53, 5546–5551 CrossRef CAS PubMed.
- H. Bunzen, ChemNanoMat, 2021, 7, 998–1007 CrossRef CAS.
- M. R. Hafner, F. Carraro, B. Abbasgholi-NA, S. Dal Zilio, M. Schmallegger, H. Wiltsche, L. Ge, M. R. Reithofer, E. Astria, L. Villanova, H. Amenitsch, C. Doonan and P. Falcaro, Adv. Funct. Mater., 2025, e18940 Search PubMed.
- W. Liang, R. Ricco, N. K. Maddigan, R. P. Dickinson, H. Xu, Q. Li, C. J. Sumby, S. G. Bell, P. Falcaro and C. J. Doonan, Chem. Mater., 2018, 30, 1069–1077 CrossRef CAS.
- M. Ge, Y. Wang, F. Carraro, W. Liang, M. Roostaeinia, S. Siahrostami, D. M. Proserpio, C. Doonan, P. Falcaro, H. Zheng, X. Zou and Z. Huang, Angew. Chem., Int. Ed., 2021, 60, 11391–11397 CrossRef CAS PubMed.
- A. D. Katsenis, A. Puškarić, V. Štrukil, C. Mottillo, P. A. Julien, K. Užarević, M.-H. Pham, T.-O. Do, S. A. J. Kimber, P. Lazić, O. Magdysyuk, R. E. Dinnebier, I. Halasz and T. Friščić, Nat. Commun., 2015, 6, 6662 CrossRef CAS PubMed.
- M. de, J. Velásquez-Hernández, R. Ricco, F. Carraro, F. T. Limpoco, M. Linares-Moreau, E. Leitner, H. Wiltsche, J. Rattenberger, H. Schröttner, P. Frühwirt, E. M. Stadler, G. Gescheidt, H. Amenitsch, C. J. Doonan and P. Falcaro, CrystEngComm, 2019, 21, 4538–4544 RSC.
- G. B. Whitfield, T. D. Brock, A. Ammann, D. Gottlieb and H. E. Carter, J. Am. Chem. Soc., 1955, 77, 4799–4801 CrossRef CAS.
- T. Sirec, A. Strazzulli, R. Isticato, M. De Felice, M. Moracci and E. Ricca, Microb. Cell Fact., 2012, 11, 100 CrossRef CAS PubMed.
- S. A. Polash, A. Poddar, S. Pyreddy, F. Carraro, A. M. D’Angelo, G. Bryant, P. Falcaro and R. Shukla, ACS Appl. Mater. Interfaces, 2025, 17, 3002–3012 CrossRef CAS PubMed.
- Y. Lu, Y. Song, L. Peng, X. Rao, K. B. Tan, S. Zhou and G. Zhan, ACS Appl. Mater. Interfaces, 2023, 15, 35552–35564 CrossRef CAS PubMed.
- L. Zheng, Z. Wang, H. Liu, N. Wang, J. Liu, M. Ma, X. Jia, M. Qian, Y. Liu, M. Li, Z. Wei and Y. Xiang, ACS Nano, 2025, 19, 17900–17916 CrossRef CAS PubMed.
- M. R. Hafner, L. Villanova and F. Carraro, CrystEngComm, 2022, 24, 7266–7271 RSC.
- M. de, J. Velásquez-Hernández, E. Astria, S. Winkler, W. Liang, H. Wiltsche, A. Poddar, R. Shukla, G. Prestwich, J. Paderi, P. Salcedo-Abraira, H. Amenitsch, P. Horcajada, C. J. Doonan and P. Falcaro, Chem. Sci., 2020, 11, 10835–10843 RSC.
- P. K. Lam, T. H. Vo, J.-H. Chen, S.-W. Lin, C.-L. Kuo, J.-J. Liao, K.-Y. Chen, S.-R. Huang, D. Li, Y.-H. Chang, H.-Y. Chen, H.-T. Hsieh, Y.-A. Hsu, H.-K. Tsao, H.-C. Yang and F.-K. Shieh, J. Mater. Chem. A, 2023, 11, 24678–24685 RSC.
- X. Luo, Y. Zhou, Y. Wang, X. Sun, J. Hu, X. Zi and Y. Fu, Mol. Catal., 2024, 560, 114131 CAS.
- P. Horcajada, S. Surblé, C. Serre, D.-Y. Hong, Y.-K. Seo, J.-S. Chang, J.-M. Grenèche, I. Margiolaki and G. Férey, Chem. Commun., 2007, 2820–2822 RSC.
- J. Liang, Q. Chen, J. Yong, H. Suyama, J. Biazik, B. Njegic, A. Rawal and K. Liang, Chem. Sci., 2024, 15, 991–1002 RSC.
- C. Li, M. Feng, B. Li, X. Feng, Y. Zhang and B. Wang, ACS Nano, 2025, 19, 2890–2899 CrossRef CAS PubMed.
- L. Ha, U. Ryu, D.-C. Kang, J.-K. Kim, D. Sun, Y.-E. Kwon, K. M. Choi and D.-P. Kim, ACS Biomater. Sci. Eng., 2021, 7, 3075–3081 CrossRef CAS PubMed.
- L. R. Kummetha, J.-J. Oh, F. H. Van Der Linden and M.-E. Aubin-Tam, Trends Biotechnol., 2025, 43, 812–825 CrossRef CAS PubMed.
- Y. Liu, M. Wu, A. Kang, X. Zhang, R. Xie, Y. Huang, J. Huang, L. Chou and C. Zhong, Adv. Funct. Mater., 2024, 34, 2309288 CrossRef CAS.
- A. Date, P. Pasini and S. Daunert, Anal. Chem., 2007, 79, 9391–9397 CrossRef CAS PubMed.
- L. M. González, N. Mukhitov and C. A. Voigt, Nat. Chem. Biol., 2020, 16, 126–133 CrossRef PubMed.
- J.-J. Oh, F. H. Van Der Linden, K. Malcı, R. A. Van Der Valk, T. Ellis and M.-E. Aubin-Tam, Sci. Adv., 2025, 11, eadw8278 CrossRef CAS PubMed.
- Y. Liu, L. Niu, N. Li, Y. Wang, M. Liu, X. Su, X. Bao, B. Yin and S. Shen, Adv. Sci., 2023, 10, 2205641 CrossRef CAS PubMed.
- X. Wu, J. Zhang, Z. Deng, X. Sun, Y. Zhang, C. Zhang, J. Wang, X. Yu and G. Yang, Biomaterials, 2025, 316, 123000 CrossRef CAS PubMed.
- X. Kuang, Y. Liu, H. Luo, Q. Li, F. Wu, C. Fan and J. Liu, J. Am. Chem. Soc., 2023, 145, 26932–26946 CrossRef CAS PubMed.
- C. Zu and J. Wang, Crit. Rev. Microbiol., 2014, 40, 225–235 CrossRef CAS PubMed.
- N. S. Forbes, Nat. Rev. Cancer, 2010, 10, 785–794 CrossRef CAS PubMed.
- W. Wang, H. Xu, Q. Ye, F. Tao, I. Wheeldon, A. Yuan, Y. Hu and J. Wu, Nat. Biomed. Eng., 2022, 6, 44–53 CrossRef CAS PubMed.
- B. Pang, X. Qiao, L. Janssen, A. Velds, T. Groothuis, R. Kerkhoven, M. Nieuwland, H. Ovaa, S. Rottenberg, O. Van Tellingen, J. Janssen, P. Huijgens, W. Zwart and J. Neefjes, Nat. Commun., 2013, 4, 1908 CrossRef PubMed.
- A. Hak, M. S. Ali, S. A. Sankaranarayanan, V. R. Shinde and A. K. Rengan, ACS Appl. Bio Mater., 2023, 6, 349–364 CrossRef CAS PubMed.
- A. Barbora, O. Bohar, A. A. Sivan, E. Magory, A. Nause and R. Minnes, PLoS One, 2021, 16, e0245350 CrossRef CAS PubMed.
- J. Zhou, C. Liu, Y. Wang, Y. Guo, X. Xu, E. Vuorimaa-Laukkanen, O. Koivisto, A. M. Filppula, J. Ye and H. Zhang, Smart Med., 2025, 4, e134 CrossRef CAS PubMed.
- M. Govender, Y. E. Choonara, P. Kumar, L. C. du Toit, S. van Vuuren and V. Pillay, AAPS PharmSciTech, 2014, 15, 29–43 CrossRef CAS PubMed.
- C. Hill, F. Guarner, G. Reid, G. R. Gibson, D. J. Merenstein, B. Pot, L. Morelli, R. B. Canani, H. J. Flint, S. Salminen, P. C. Calder and M. E. Sanders, Nat. Rev. Gastroenterol. Hepatol., 2014, 11, 506–514 CrossRef PubMed.
- O.-A. Praepanitchai, A. Noomhorm and A. K. Anal, BioMed Res. Int., 2019, 2019, 1–8 CrossRef PubMed.
- M. Saarela, G. Mogensen, R. Fondén, J. Mättö and T. Mattila-Sandholm, J. Biotechnol., 2000, 84, 197–215 CrossRef CAS PubMed.
- C. C. Dodoo, J. Wang, A. W. Basit, P. Stapleton and S. Gaisford, Int. J. of Pharm., 2017, 530, 224–229 CrossRef CAS PubMed.
- J. Levy, Am. J. Gastroenterol. Suppl., 2000, 95, S8–S10 CrossRef CAS PubMed.
- F. J. Rodrigues, M. F. Cedran, J. L. Bicas and H. H. Sato, Food Res. Int., 2020, 137, 109682 CrossRef CAS PubMed.
- K. Vivek, S. Mishra, R. C. Pradhan, M. Nagarajan, P. K. Kumar, S. S. Singh, D. Manvi and N. N. Gowda, Appl. Food Res., 2023, 3, 100248 CrossRef CAS.
- X. Qi, F. Zhu, Z. Chang and Y. Deng, Langmuir, 2024, 40, 16172–16179 CrossRef CAS PubMed.
- S. Ghasemi, J. B. Raoof, M. Ghani and R. Ojani, Talanta, 2024, 269, 125403 CrossRef CAS PubMed.
- M. Liu, L. Zhang, R. Yang, H. Cui, Y. Li, X. Li and H. Huang, J. Hazard. Mater., 2024, 461, 132475 CrossRef CAS PubMed.
- N. Aziminia, M. Hadjipavlou, Y. Philippou, S. S. Pandian, S. Malde and M. Y. Hammadeh, BJU Int., 2019, 123, 753–768 CrossRef PubMed.
- S. Li, X. Zhou, Z. Chen, F. C. Herbert, R. Jayawickramage, S. D. Panangala, M. A. Luzuriaga, S. B. Alahakoon, S. D. Diwakara, X. Meng, L. Fei, J. Ferraris, R. A. Smaldone and J. J. Gassensmith, ACS Appl. Mater. Interfaces, 2020, 12, 11884–11889 CrossRef CAS PubMed.
- L. Ran, Y. Lin, G. Su, Z. Yang and H. Teng, ChemBioChem, 2024, 25, e202400147 CrossRef CAS PubMed.
- Y. Xiang, H. Yan, B. Zheng, A. Faheem and Y. Hu, Anal. Chem., 2020, 92, 12338–12346 CrossRef CAS PubMed.
- A. Karnholz, K. Küsel, A. Gößner, A. Schramm and H. L. Drake, Appl. Environ. Microbiol., 2002, 68, 1005–1009 CrossRef CAS PubMed.
- M. Vázquez-González, C. Wang and I. Willner, Nat Catal, 2020, 3, 256–273 CrossRef.
- X. Liu, W. Qi, Y. Wang, R. Su and Z. He, Nanoscale, 2017, 9, 17561–17570 RSC.
- X. Lian, Y. Huang, Y. Zhu, Y. Fang, R. Zhao, E. Joseph, J. Li, J. Pellois and H. Zhou, Angew. Chem., Int. Ed., 2018, 57, 5725–5730 CrossRef CAS PubMed.
- Z. Xu, G. Xiao, H. Li, Y. Shen, J. Zhang, T. Pan, X. Chen, B. Zheng, J. Wu, S. Li, W. Zhang, W. Huang and F. Huo, Adv. Funct. Mater., 2018, 28, 1802479 CrossRef.
- I. Koltover, T. Salditt, J. O. Rädler and C. R. Safinya, Science, 1998, 281, 78–81 CrossRef CAS PubMed.
- C. LoPresti, H. Lomas, M. Massignani, T. Smart and G. Battaglia, J. Mater. Chem., 2009, 19, 3576 RSC.
- S. Cao, L. C. Da Silva and K. Landfester, Angew. Chem., Int. Ed., 2022, 61, e202205266 CrossRef CAS PubMed.
- N. Martin, J.-P. Douliez, Y. Qiao, R. Booth, M. Li and S. Mann, Nat. Commun., 2018, 9, 3652 Search PubMed.
- H. Cai, Y.-L. Huang and D. Li, Coord. Chem. Rev., 2019, 378, 207–221 CrossRef CAS.
- S. Moon, Y. Liu, J. T. Hupp and O. K. Farha, Angew. Chem., Int. Ed., 2015, 54, 6795–6799 CrossRef CAS PubMed.
- J. Huo, M. Marcello, A. Garai and D. Bradshaw, Adv. Mater., 2013, 25, 2717–2722 Search PubMed.
- J. Zhang, N. Jin, N. Ji, X. Chen, Y. Shen, T. Pan, L. Li, S. Li, W. Zhang and F. Huo, ACS Appl. Mater. Interfaces, 2021, 13, 52215–52233 Search PubMed.
- Z. Xu, J. Zhang, T. Pan, H. Li, F. Huo, B. Zheng and W. Zhang, Chem. Mater., 2020, 32, 3553–3560 CrossRef CAS.
- D. Tian, R. Hao, X. Zhang, H. Shi, Y. Wang, L. Liang, H. Liu and H. Yang, Nat. Commun., 2023, 14, 3226 CrossRef CAS PubMed.
- B. Zhang, J. Zhang, C. Liu, L. Peng, X. Sang, B. Han, X. Ma, T. Luo, X. Tan and G. Yang, Sci. Rep., 2016, 6, 21401 CrossRef CAS PubMed.
- D. Lee and D. A. Weitz, Small, 2009, 5, 1932–1935 Search PubMed.
- K. S. Rabe, J. Müller, M. Skoupi and C. M. Niemeyer, Angew. Chem., Int. Ed., 2017, 56, 13574–13589 Search PubMed.
- A. C. Marr and S. Liu, Trends Biotechnol., 2011, 29, 199–204 Search PubMed.
- J. Zhang, Y. Shen, N. Jin, X. Zhao, H. Li, N. Ji, Y. Li, B. Zha, L. Li, X. Yao, S. Zhang, F. Huo and W. Zhang, Research, 2022, 2022, 9847698 CAS.
- H. Chang, X. Wen, Z. Li, Z. Ling, Y. Zheng and C. Xu, Bioeng. Transla Med., 2023, 8, e10457 CrossRef CAS PubMed.
- G. Kumar, Y. C. Wang, C. Co and C.-C. Ho, Langmuir, 2003, 19, 10550–10556 Search PubMed.
- S. Z. Yow, C. H. Quek, E. K. F. Yim, C. T. Lim and K. W. Leong, Biomaterials, 2009, 30, 1133–1142 CrossRef CAS PubMed.
- D. Dutta, A. Pulsipher, W. Luo and M. N. Yousaf, J. Am. Chem. Soc., 2011, 133, 8704–8713 CrossRef CAS PubMed.
- M. W. Eskew and A. S. Benight, Thermochim. Acta, 2022, 715, 179274 CrossRef CAS.
- M. R. Banki, T. U. Gerngross and D. W. Wood, Protein Sci., 2005, 14, 1387–1395 CrossRef CAS PubMed.
- J. S. Decker, U. Yano, R. M. Melgar and M. D. Lynch, Biotechnol. J., 2024, 19, 2400005 CrossRef CAS PubMed.
- M. Cárdenas-Fernández, W. Neto, C. López, G. Álvaro, P. Tufvesson and J. M. Woodley, Biotechnol. Prog., 2012, 28, 693–698 CrossRef PubMed.
- R. R. Chen, Appl. Microbiol. Biotechnol., 2007, 74, 730–738 Search PubMed.
- Y. Ni and R. R. Chen, Biotech. Bioeng., 2004, 87, 804–811 CrossRef CAS PubMed.
- R. A. Sheldon and J. M. Woodley, Chem. Rev., 2018, 118, 801–838 CrossRef CAS PubMed.
- C. Ning, X. Wang, L. Li, Y. Zhu, M. Li, P. Yu, L. Zhou, Z. Zhou, J. Chen, G. Tan, Y. Zhang, Y. Wang and C. Mao, Chem. Res. Toxicol., 2015, 28, 1815–1822 Search PubMed.
- A. G. Renehan, BMJ, 2001, 322, 1536–1538 CrossRef CAS PubMed.
- S. L. Fink and B. T. Cookson, Infect. Immun., 2005, 73, 1907–1916 CrossRef CAS PubMed.
- K. Simu, K. Holmfeldt, U. L. Zweifel and Å. Hagström, Appl. Environ. Microbiol., 2005, 71, 4793–4800 CrossRef CAS PubMed.
- V. Kilin, O. Glushonkov, L. Herdly, A. Klymchenko, L. Richert and Y. Mely, Biophys. J., 2015, 108, 2521–2531 CrossRef CAS PubMed.
Footnotes |
| † Equal contribution. |
| ‡ Current affiliation: Center for Membrane Separations, Adsorption, Catalysis, and Spectroscopy (cMACS), B-3001 Leuven, Belgium. |
| § Apoptosis describes the orchestrated collapse of a cell characterised by membrane blebbing, cell shrinkage, condensation of chromatin, and fragmentation of DNA followed by rapid engulfment of the corpse by neighbouring cells.215 It is a regulated process essential for maintaining tissue homeostasis, embryonic development, and immune system function. Dysregulation of apoptosis can contribute to various pathological conditions, such as cancer (e.g. cell immortalization) and degenerative diseases. |
| ¶ Necrosis is a passive, accidental cell death triggered by environmental perturbations, leading to the unregulated release of inflammatory cellular components.216 |
| || Culturability is defined as the ability of a single cell to yield a population discernible by the observer, usually a visible colony on the surface of a nutrient agar plate. Such culture-based techniques have been in use for many decades, have generated a coherent body of information, and have a proven track record in protecting public health at relatively low cost.147,217 |
| ** F2N12S: N-[[4′-N,N-diethylamino-3-hydroxy-6-flavonyl]-methyl]-N-methyl-N-(3-sulfopropyl)-1-dodecanaminium is a fluorophore which is highly sensitive to the lipid order of lipid bilayers.218 |
| †† SGF: simulated gastric fluid, a lab-prepared solution mimicking the acidic conditions in the human stomach. |
| This journal is © The Royal Society of Chemistry 2026 |