DOI:
10.1039/D6CB00048G
(Review Article)
RSC Chem. Biol., 2026, Advance Article
Let the peptides shine: SOX (Sulfonamido-OXine)-labelled peptides for direct kinase and phosphatase monitoring
Received
4th February 2026
, Accepted 16th May 2026
First published on 19th May 2026
Abstract
Protein kinases and their cognate phosphatases are central regulators of cellular signalling, acting through the opposing processes of phosphorylation and dephosphorylation. Defects in kinase/phosphatase signalling are implicated in many diseases, including cancer, neurodegenerative conditions and metabolic disorders. Rapid and economical methods for measuring kinase activities are critically needed as diagnostic tools and for the evaluation of kinase inhibitors, including approaches that allow direct and quantitative measurement of kinase activities in complex media like cell lysates. SOX (sulfonamido-oxine)-labelled peptides, that upon phosphorylation chelate Mg2+ and undergo Chelation-Enhanced Fluorescence (CHEF), have emerged as a convenient class of modular kinase sensors. The development of an array of SOX-peptides has allowed researchers to gain insights into biological processes including binding events, conformational constraints, enzyme activities, protein trafficking and localization. This focused review provides an overview of the use of SOX-labelled peptides for probing kinase and phosphatase activity in real-time assays, including their development, synthesis, applications and strengths and weaknesses compared to other kinase assays.
Introduction
Common kinase assays and limitations
Protein phosphorylation on Ser, Thr and Tyr is a common post-translational modification (PTM).1,2 This PTM is controlled by the opposing actions of kinases and phosphatases, ubiquitous and highly conserved elements of signalling pathways.2–6 Although different protein kinases process different substrates, they share broadly conserved structural motifs, including ATP-binding domains and catalytic site residues.7–9 Aberrant levels of kinase activity are associated with cancer, infectious diseases, neurodegenerative conditions and metabolic disorders (Fig. 1).2,8,10,11 Therefore, detection of kinase activity is crucial for screening novel kinase inhibitors and identification of kinase substrates.10 Sensitive, selective and high-throughput chemosensors are invaluable in academic and pharmaceutical settings for unravelling these diverse signalling cascades.12,13
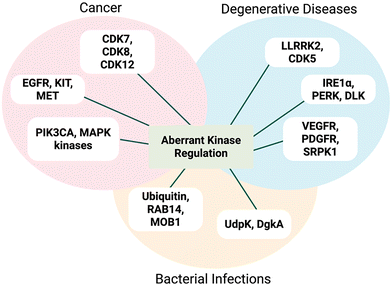 |
| | Fig. 1 Representative examples of different classes of kinase aberrant activity in many diseases, including cancer, neurodegenerative disorders and bacterial infections. | |
The crucial roles of kinases and phosphatases in normal cellular communication and physiology underscore the unmet need for robust analytical tools.2,14,15 Functional in vitro assays facilitate kinase profiling, evaluating drug efficacy prior to in vivo analyses. Typically, phosphorylated substrates can be quantified indirectly or directly using endpoint (discontinuous) or continuous (real-time) assays, respectively.16,17 In endpoint assays, an aliquot of the reaction mixture is removed prior to analysis of the reaction product (ADP or phosphorylated substrate). For these assays to be valid, the chosen timepoint must fall in the linear portion of the progress curve so that the endpoint signal reflects the initial reaction rate.18,19 While endpoint assays are valuable for early profiling and selectivity screening, they provide limited information on enzyme kinetics and inhibitor mechanism.12 Thus, time-dependent inhibition (TDI) can be missed or mischaracterised.20 Unlike endpoint methods, real-time assays track reactions over the entire course of an experiment. By capturing full progress curves, continuous assays detect TDI accurately, facilitating the screening of potent inhibitors.16,17,21 Moreover, they differentiate between reversible and irreversible inhibition mechanisms in complex biological samples.22,23 This is particularly effective for quantifying kinetic parameters and providing PK/PD strategies. Commonly employed approaches for monitoring kinase activity include mass spectrometry-based, immunoassays, and radioactivity-based approaches.2,24 While these techniques have proven effective, they also have several limitations.2,22,25,26 Quantitative mass spectrometry platforms require sample enrichment to detect low-abundance analytes.27 Immunoassays rely on validation of the antibodies from batch to batch to ensure their specificity.28 32P-based assays are inherently discontinuous and hence not practical for high-throughput screening applications.2,29,30 Fluorescence sensing is a compelling alternative in biomedical research and clinical applications, offering operational simplicity, rapid response kinetics and suitability for bioimaging with spatiotemporal resolution.24,31,32 Various kinds of kinase sensing systems including fluorophore dyes, nanomaterials and lanthanides are used to interrogate signalling processes at the molecular level. Despite these contributions, several technical, biological and material-related concerns remain. Fluorophore dyes (for example Alexa Fluor 647, cyanine dyes and rhodamine derivatives) yield signals that are strongly influenced by photostability, duty cycle, sensitivity to buffer conditions and activation rates by light sources.33–35 Additionally, they can introduce cellular perturbations which complicate their application in kinase assays.36–38 Fluorescent nanomaterials (FNMs) include gold nanoclusters (AuNCs), carbon nanodots (CDs) and quantum dots (QDs). However, most nanomaterial-based kinase assays are endpoint.39–42 In the synthesis process, the surface of FNMs can be hard to distribute uniformly due to agglomeration of small particles. FNMs also suffer from variable quantum yields that compromise quantitative accuracy of assays.43–47 Lanthanide-based compounds are being actively developed in biomedical imaging, yet limited progress in kinase sensing has been achieved so far. Surface quenching owing to surface defects, adsorbed water, or hydroxyl groups encourages non-radiative relaxation.48,49 Rare-earth elements are associated with high extraction costs and environmentally damaging mining practices. Moreover, most of the advanced synthesis methods are not easily translatable to industrial-scale production.50,51 Until now, only a few simple and rapid single step continuous assays have been developed. Förster Resonance Energy Transfer (FRET) assays offer a strong distance-dependent mechanism for the quantification of biomolecular distances and concentrations via fluorescent biosensing. Genetically encoded FRET partners undergo conformational changes upon phosphorylation.26,27 The distance of the donor and the acceptor is related to the efficiency of FRET. While FRET assays are convenient tools for continuous monitoring of peptidases, their application to kinase assays typically relies on indirect readouts.4,52–56 Several fluorescent analogues for FRET have been developed and in theory can be applied for kinase sensing. However, most FRET changes are less than 50% which greatly reduces the sensitivity of these probes and kinase activity has to be high to be detected.14,57,58
Early chelation-enhanced fluorescence (CHEF) probes for kinase sensing
In kinase-sensor design, CHEF probes have emerged as a class of modular scaffolds. These probes have no fluorescence (or very weak fluorescence) themselves but upon chelation with a metal ion, a significant fluorescence enhancement is observed.24,59 CHEF probes are distinguished by their heteroatoms (for example amines) containing a suitably positioned lone pair that quenches fluorescence through photoinduced electron transfer (PET) and thereby increases fluorescence.10,60–63 The coordination effectively stabilizes the energy of the lone-pair-derived orbital relative to the fluorophore, restoring emission.64 Early CHEF probes for Zn2+ sensing and in turn inorganic phosphate detection were developed by Huston and co-workers. They synthesized a structurally simple anthracene fluorophore (1) that reported dramatic fluorescence increase (over 1000-fold) upon Zn2+chelation in acetonitrile.65 Over the next several years the same group developed additional anthrylazamacrocycle conjugate probes to explore their CHEF properties.66 Probes 2 and 3 (Fig. 2) showed remarkable fluorescence enhancements (from 108- up to 140-fold) by addition of Zn2+ and Cd2+ in aqueous media.67 In 1994, Vance and Czarnik established the first non-destructive, real-time assay using a high affinity anthrylpolyamine CHEF chemosensor (3). This probe showed micromolar affinity for pyrophosphate, over 2000-fold higher than for phosphates, allowing direct monitoring of inorganic phosphatase.68 Although potent, probes 1–3 lacked specificity and selectivity. These sensors could not be translated to kinase monitoring, leaving an innovation gap until the late 1990s, when the Imperiali group pioneered the use of SOX-labelled peptides for monitoring kinase activity.12,14,69,70 This offered considerable scope for the development of self-sensing sensors for assaying kinase activity in real-time.2,14,71,72
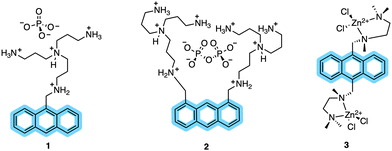 |
| | Fig. 2 Early CHEF probes (1–3) developed by Czarnik and co-workers. Anthracene moieties conjugated with polyamine binding sites yielded strong CHEF “ON” responses in organic and aqueous media. | |
SOX-labelled peptides for continuous kinase assays
Given the ability of anthracene probes to chelate phosphate anions and Zn2+, chelation-sensitive fluorophores were soon incorporated into peptides to create self-sensing kinase sensors. The modular design of these peptides relies on a kinase recognition motif, tagged with the phosphorylation-sensitive SOX fluorophore (Scheme 1).14,15,73 Unlike conventional fluorophores, SOX (λex =365 nm and λem =485 nm) reports direct readouts of kinase activity in their native chemical environment.14 SOX is covalently attached to a peptide substrate and utilized to report tyrosine or serine/threonine phosphorylation via the CHEF effect.70,74 SOX-peptides have low fluorescence but upon phosphorylation and chelation to Mg2+, a large increase in fluorescence is observed due to the CHEF effect.73 The SOXylating agent required for the introduction of the SOX fluorophore into peptides, SOX-Br, is synthesized from commercially available 8-hydroxy-2-methylquinoline (Scheme 1). Chlorosulfonylation introduces a sulfonyl chloride group, followed by displacement of chloride with dimethylamine. This intermediate is then protected as a tert-butyldiphenylsilyl ether before the benzylic position is brominated via a Riley oxidation, reduction and bromination to yield the quinoline-based fluorophore SOX-Br.
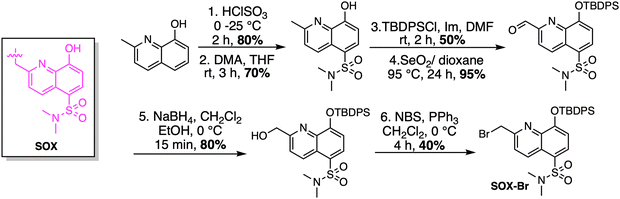 |
| | Scheme 1 Synthesis of SOX-Br. DMA: dimethylamine, TBDPSCl: tert-butyldiphenylsilyl ether, Im: imidazole, NBS: N-bromosuccinimide. | |
The Imperiali lab established two types of modular SOX sensors: (i) tunable Zn2+ sensors, which are beyond the scope of this review; and (ii) versatile probes of kinase activity. Three distinct classes of SOX-peptides have been profiled: (1) β-turn focused (BTF); (2) recognition-domain focused (RDF); and (3) chimeric.2,29,69,73 These peptide sensors have the SOX group incorporated within the nonnatural amino acids Ala(SOX) or Cys(SOX). The BTF-peptides have a β-turn motif containing Ala(SOX), and either an N- or C-terminal kinase recognition motif (Fig. 3).14,15,75 Although active under a variety of conditions, the BTF-peptides are limited by this conformationally constrained β-turn moiety.12,69 This separates the fluorophore from the phosphorylation site, rendering some sensors less selective and specific (i.e., more promiscuous) due to loss of sequence recognition determinants.15,76,77 To address this challenge, a more powerful class of SOX-peptides was developed, the recognition-domain focused (RDF) peptides. In the RDF design, S-alkylation of a cysteine residue with SOX-Br affords Cys(SOX). This approach circumvents the constrained nature of β-turn motifs.76 With the aim of developing optimal sensors, the RDF strategy has been extended to protein-based probes, termed chimeric reporters. In chimeric reporters, the Cys(SOX) can be placed anywhere in the protein to discriminate challenging targets with short or ubiquitous consensus sequences.78,79
 |
| | Fig. 3 Distinct classes of SOX-peptide probes. Recognition elements are fully conserved in the RDF design and only partially so in the BTF design due to the required β-turn. The SOX chromophore can be positioned N- or C-terminal to the Ser/Thr/Tyr residue in both designs. | |
SOX-peptides and -proteins can be prepared using published methods.80 For the synthesis of the BTF class of SOX-peptides, the unnatural amino acid Fmoc-Ala(SOX)-OH was synthesized and used (Scheme 2).14,70,74 SOX-Br is used to alkylate a glycine derivative in the presence of a phase-transfer catalyst, which upon acid hydrolysis yields Ala(SOX). Fmoc protection is then carried out, yielding Fmoc-Ala(SOX)-OH for peptide synthesis.69,70,74
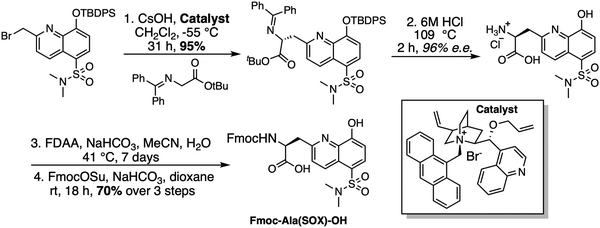 |
| | Scheme 2 Synthesis of Fmoc-Ala(SOX)-OH for use in Fmoc SPPS. FDAA = 1-Fluoro-2-4-dinitrophenyl-5-L-alanine amide; Fmoc = 9-fluorenylmethyloxycarbonyl. | |
The RDF class of SOX-peptides are also constructed via Fmoc-based SPPS but rather than utilizing Fmoc-Ala(SOX)-OH, they were prepared using Fmoc-Cys(SOX(TBDPS))-OH (Scheme 3).69 This includes an appropriately placed cysteine protected with a trityl group. After selective on-resin sulfhydryl group deprotection, the free thiol is alkylated with SOX-Br. Standard TFA cleavage and concomitant removal of all the side chain protecting groups afford the non-natural Cys(SOX) amino acid with excellent conversion (>95%). Cys(SOX) is then utilized in automated SPPS or SPOT synthesis to generate RDF peptide libraries.12,69,80
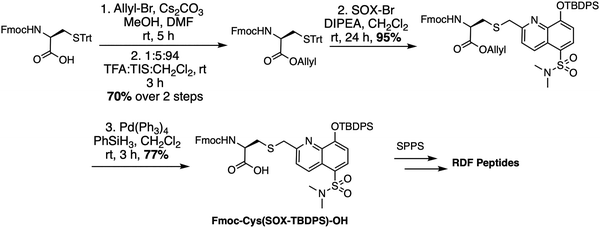 |
| | Scheme 3 Synthesis of Fmoc-Cys(SOX(TBDPS))-OH for use in Fmoc SPPS. Trt: trityl, DIPEA: N,N-diisopropylethylamine, TFA: trifluoroacetic acid. | |
β-Turn focused (BTF) sensors
BTF sensors have been successfully applied as chemosensors (with 4–12-fold fluorescence increases upon phosphorylation) to monitor kinase activity with pure enzyme and in unfractionated cell extracts.76 The BTF design contains Ala(SOX) (−)2 or (+)2 amino acids away from the phosphorylation site, a kinase recognition motif that includes the amino acid being phosphorylated, and a β-turn that preorganizes a binding site for interaction with Mg2+.14,69,75,77 The β-turn also prevents interference of the SOX moiety with the enzyme by placing it further away from the phosphorylation site (Fig. 4). The kinase recognition motif is varied to target the desired kinase. BTF sensors successfully targeted kinases including PKC, PKA, Akt and MK2 (Table 1).14 Notably, these kinase assays required less than 0.1 mg total cell protein for measurements compared to the ∼0.5 mg for complex kinase assays.75 Across the range of probes, serine yields larger increases in fluorescence than threonine (cf. PKC-S1 with PKC-S2 and Akt-S1 with Akt-S4). Another meaningful trend is that Mg2+ coordination is enhanced with rigidity (cf. Cdk2-S1 with PKC-S2 and PKA-S3 with Pim2-S2). Incorporation of an additional aspartate residue (cf. PKC-S5 and PKC-S6) didn’t significantly change the fluorescence compared to the closely related peptide PKC-S4.69,76
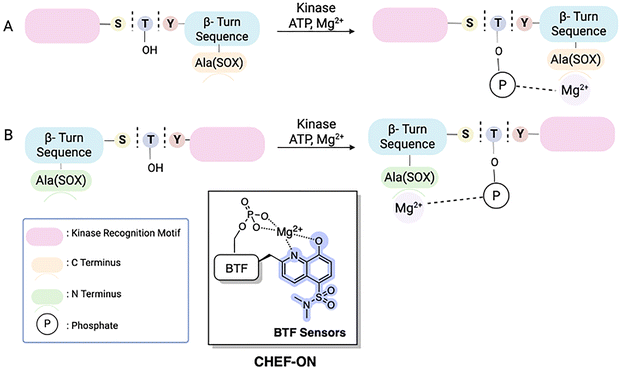 |
| | Fig. 4 Schematic representation of 1st generation SOX-peptides, termed BTF sensors. The key modules include the SOX fluorophore, separated from the phosphorylation site by a β-turn. The binding site for Mg2+ is preorganized by the β-turn sequence. Design of kinase recognition motif for C-terminus (A) and N-terminus (B).75 | |
Table 1 Comparison of BTF and RDF SOX-peptides as kinase sensors
| Target kinase |
Design |
Substrate sequence (all C-terminal amides) |
KM (µM) |
Fold fluorescence increase |
| PKCα |
BTF |
Ac-Ala(SOX)-P-G-![[S with combining low line]](https://www.rsc.org/images/entities/char_0053_0332.gif) -F-R-R-R -F-R-R-R |
8.6 ± 2.9 |
5.7 |
| PKA |
Ac-L-R-R-A--L-P-Ala(SOX) |
1.8 ± 0.5 |
1.3 |
| Akt |
Ac-A-R-K-R-E-R-A-Y--F-DP-Ala(SOX)-G |
3.8 ± 0.1 |
9.3 |
| MK2 |
Ac-A-H-L-Q-R-Q-L--I-DP-Ala(SOX)-G |
21 ± 1 |
8.5 |
| PKA |
Ac-L-R-R-A--L-DP-Ala(SOX)-G |
2.9 ± 0.3 |
5.6 |
| PKC-S1 |
Ac-Ala(SOX)-P-G--F-R-R-R |
8.6 ± 2.9 |
6.7 ± 0.6 |
| PKC-S2 |
Ac-Ala(SOX)-P-G-![[T with combining low line]](https://www.rsc.org/images/entities/char_0054_0332.gif) -F-R-R-R -F-R-R-R |
23 ± 2.3 |
3.8 |
| Cdk2-S1 |
Ac-Ala(SOX)-P-K--P-K-K-A-K-K-L |
0.6 ± 0.1 |
4.7 ± 0.5 |
| PKA-S2 |
Ac-L-R-R-A--L-P- Ala(SOX) |
1.8 ± 0.5 |
4.0 |
| PKA-S3 |
Ac-L-R-R-A--L-DP- Ala(SOX)-G |
2.9 ± 0.6 |
6.5 ± 0.7 |
| Akt-S1 |
Ac-A-R-K-R-E-R-A-Y--F-DP-Ala(SOX)-G |
3.8 ± 0.2 |
7.6 ± 0.6 |
| Akt-S4 |
Ac-A-R-K-R-E-R-T-Y--F-DP-Ala(SOX)-G |
3.8 ± 0.7 |
4.5± 0.3 |
| MK2-S1 |
Ac-A-H-L-Q-R-Q-L--I- DP-Ala(SOX)-G |
21 ± 2 |
7.7 ± 0.9 |
| Pim2-S2 |
Ac-A-R-K-R-R-R-H-DP--G-P-Ala(SOX)-G |
8.6 ± 2.9 |
3.0 ± 0.4 |
| Src |
RDF |
Ac-A-E-E-Cys(SOX)-I-![[Y with combining low line]](https://www.rsc.org/images/entities/char_0059_0332.gif) -G-E-F-E-A-K-K-K-K -G-E-F-E-A-K-K-K-K |
7.0 ± 1.0 |
2.0 ± 0.1 |
| Abl |
Ac-E-Cys(SOX)-I--A-A-P-F-A-K-K-K |
10.5 ± 1.5 |
4.6 ± 0.5 |
| IRK |
Ac-R-Cys(SOX)-D--Nle-T-M-Q-I-G-K-K |
27.1 ± 3.9 |
4.2 ± 0.1 |
| PKCα |
Ac-R-R-R-Cys(SOX)-A--F-R-R-R |
0.13 ± 0.02 |
3.7 ± 0.2 |
| PKCβI |
Ac-L-K-R-Cys(SOX)-A--F-K-K-F-A |
0.81 ± 0.18 |
9.7 ± 0.5 |
| PKCδ |
Ac-R-K-R-K-G--F-Cys(SOX)-Y-G-G |
0.48 ± 0.07 |
7.3 ± 0.1 |
| Pim2 |
Ac-A-R-K-R-R-R-H-P--G-Cys(SOX)-P-T-A |
1.4 ± 0.1 |
3.2 ± 0.1 |
| Akt1 |
Ac-A-R-K-R-E-R-A-Y--F-Cys(SOX)-H-H-A |
0.69 ± 0.11 |
3.9 ± 0.3 |
| MK2 |
Ac-A-H-L-Q-R-Q-L--I-Cys(SOX)-H-H |
1.2 ± 0.2 |
4.4 ± 0.2 |
| PKA |
Ac-A-L-R-R-A--L-Cys(SOX)-A-A |
2.6 ± 0.3 |
5.0 ± 0.2 |
Recognition-domain focused (RDF) sensors
RDF-sensors enabled the inclusion of extended binding sequences, which maximized recognition by the cognate kinase (Fig. 5). Cys(SOX) is placed either (−)2 or (+)2 amino acids from the phosphorylation site, similarly to BTF-peptides. RDF-peptides offer several advantages compared to BTF peptides: (i) kinase recognition sequences can be included on both sides of the phosphorylation site; (ii) SOX addition to cysteine can be performed either during SPPS or on unprotected peptides via cysteine SOXylation; (iii) up to 28-fold improved catalytic activity; and (iv) improved kinetic parameters (up to 66-fold lower KM compared to their BTF counterparts).69,77 A variety of different RDF-peptides have been constructed that target representative Ser/Thr (PKCα, PKCβI, PKCδ, Pim2, Akt1, MK2 and PKA), receptor (IRK) and nonreceptor Tyr (Src, Abl) kinases.77 Based on the RDF design, new SOX-peptides have been developed using high-throughput mass-spectrometry.12 Furthermore, these RDF-peptides were used in a 384 well plate format to monitor the activity of multiple kinases at once (multiplexing). This shows the amenability of SOX probes for the screening of kinase inhibitors, with the potential for such inhibitors to be taken forward as therapeutic leads. Indeed, the Dalby laboratory detailed the use of SOX-based sensors to identify eEF-2 inhibitor scaffolds from a 32![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 960-member inhibitor library.81,82 The “SOXtide” substrate (Ac-RKK
960-member inhibitor library.81,82 The “SOXtide” substrate (Ac-RKK![[Y with combining low line]](https://www.rsc.org/images/entities/b_char_0059_0332.gif) KFNC-Cys(SOX)-DRRRFL-NH2) resulted in a five-fold increase in fluorescence emission. This allowed for the continuous monitoring of eEF-2 activity in vitro, including samples of tissue homogenates and lysates.81 Stains and colleagues described three optimal sensor constructs (FAKtide-S2, Nictide-S2 and ROCK-S1) for focal adhesion kinase (FAK), leucine-rich repeat kinase 2 (LRRK2) and Rho-associated protein kinase (ROCK) respectively.83–86 FAKtide-S2 (Ac-VSETDDA-Cys(SOX)-IIDEEDT-NH2) and Nictide-S2 (Ac-RLGWWR-Cys(SOX)-YLRRARQGNTKQR-NH2) detected concentrations of their targets as low as 1.0 and 2.5 nM and are highly reproducible (Z’ = 0.91 and 0.7 respectively). Similarly, ROCK-S1 (KPARKKRYV-Cys(SOX)-GNPYWM) sensitively reported ROCK activity with a limit of detection of 10 pM. As a proof-of-principle ROCK-S1 also rapidly identified inhibitor scaffolds (PHA665752 and IKK16) against ROCK2 from a 78 small molecule library. FAK, and ROCK are actively associated with hepatocellular carcinoma tumors (HCC), while LRRK2 with forms of Parkinson's disease.87–89
KFNC-Cys(SOX)-DRRRFL-NH2) resulted in a five-fold increase in fluorescence emission. This allowed for the continuous monitoring of eEF-2 activity in vitro, including samples of tissue homogenates and lysates.81 Stains and colleagues described three optimal sensor constructs (FAKtide-S2, Nictide-S2 and ROCK-S1) for focal adhesion kinase (FAK), leucine-rich repeat kinase 2 (LRRK2) and Rho-associated protein kinase (ROCK) respectively.83–86 FAKtide-S2 (Ac-VSETDDA-Cys(SOX)-IIDEEDT-NH2) and Nictide-S2 (Ac-RLGWWR-Cys(SOX)-YLRRARQGNTKQR-NH2) detected concentrations of their targets as low as 1.0 and 2.5 nM and are highly reproducible (Z’ = 0.91 and 0.7 respectively). Similarly, ROCK-S1 (KPARKKRYV-Cys(SOX)-GNPYWM) sensitively reported ROCK activity with a limit of detection of 10 pM. As a proof-of-principle ROCK-S1 also rapidly identified inhibitor scaffolds (PHA665752 and IKK16) against ROCK2 from a 78 small molecule library. FAK, and ROCK are actively associated with hepatocellular carcinoma tumors (HCC), while LRRK2 with forms of Parkinson's disease.87–89
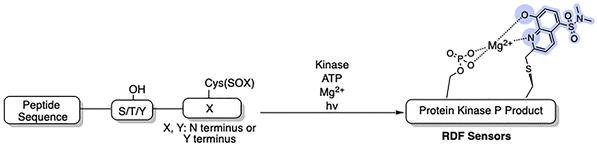 |
| | Fig. 5 RDF designs from optimized nonfluorescent substrates. Circumventing the β-turn requirement allows the recognition elements to be fully conserved in the RDF design. The SOX chromophore can be positioned N- or C- terminus to the Ser/Thr/Tyr residue.77 | |
Chimeric sensors
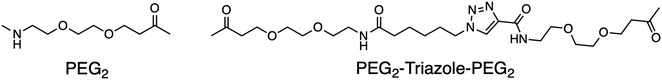
BTF- and RDF-sensors gave sensitive and robust signals with direct readouts under physiological conditions when the SOX group is placed within (+2) or (−2) amino acids away from the phosphorylation site.76,77 However, several physiologically important kinases including mitogen-activated protein kinases (MAPKs) are acutely reliant on secondary interactions distal to the phosphorylation site that impart substrate specificity.2,15,76,78,79 MAPKs are extremely challenging in terms of sensor design due to the minimal consensus sequences required for recognition, such as a Ser-Pro or a Thr-Pro dipeptide motif.2,78 To achieve higher selectivity for MAPKs, the third generation of SOX biosensors was developed, known as chimeric sensors. These are protein-based reporters, where the Cys(SOX) is placed anywhere in the protein, enabling the detection of challenging targets with short or ubiquitous consensus sequences.90 Two chimeric sensors were reported for ERK1/2 and p38-α, named SOX-PNT and MEF2A-CSOX respectively (Fig. 6). SOX-PNT consists of the recombinant PNT domain from the Ets-1 transcription factor and the synthetic ERK1/2 consensus motif.79,91–93 In MEF2A-CSOX the phosphorylation site is linked to the docking peptide sequence via a flexible linker AOO (8-amino-3,6-dioxaoctanoic acid). Both probes showed good affinities (KMSOX-PNT = 14.9 µM and KMMEF2A-CSOX = 1.3 µM) alongside excellent fluorescence responses (4–17-fold) which would be impossible to achieve with simple peptide probes.78,79 Background affinity is minimal in the presence of closely related isoforms.78,79 Using a docking domain-based strategy, three activity sensors were developed for the kinase subfamilies, ERK1/2, p38α/β and JNK1/2/3 (Table 2). Each manifested a 70-fold enhanced affinity. Triazole and polyethylene glycol were used as linkers to join the two sensor modules. The three chimeric sensors showed improved binding affinities (KM (ca 0.14–3.3 µM)) compared to SOX-PNT and MEF2A-CSOX. These MAP kinase sensors quantified kinase activities in cell lysates over an extended time course. Kinase inhibitor activities were also monitored in either recombinant protein or cell-lysate based assays.2 The MAP sensors were also applied in vitro in a breast cancer progression model based on the MCF10A series, where p38α/β activity correlated with increased tumorigenicity.2,13,94–96 This further underscored the practical utility of SOX-chimeric sensors in profiling kinase activity and quantifying disease progression in cell culture models.
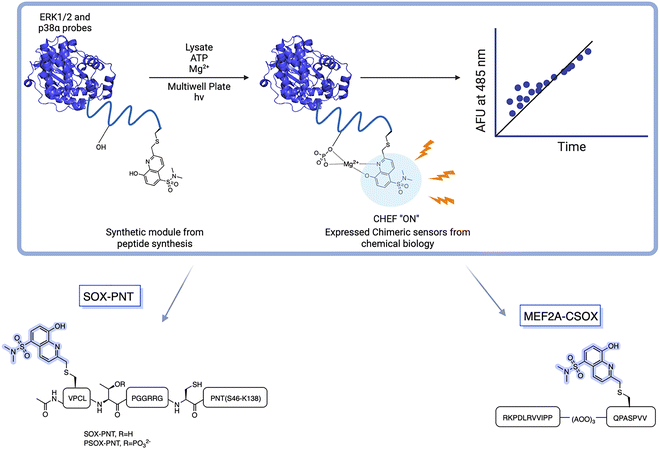 |
| | Fig. 6 Rational design of Cys(SOX)-based chimeric sensors SOX-PNT and MEF2A-CSOX in which a known peptide sequence is linked to CSOX via a flexible linker. SOX-PNT probes the ERK1/2, utilizing the PNT domain from Ets-1 for specific binding to the enzyme. The docking domain motif of MEF2A-CSOX is linked to the phosphorylation site with AOO linker. | |
Table 2 Sequences of Modular MAPK Activity Sensors
| Kinase |
Docking motif |
Linker |
Phosphorylation site |
| JNK1/2/3 |
ERPSRDHLYLPLEP |
PEG2 |
SANLL![[P with combining low line]](https://www.rsc.org/images/entities/char_0050_0332.gif) -Cys(SOX)-PA -Cys(SOX)-PA |
| p38α/β |
IKIKKIEDASNPLLLKRRKKG |
PEG2-Triazole- PEG2 |
QP-Cys(SOX)-AVV |
| ERK1/2 |
PQLKPIESSILAQRRVRKLG |
PEG2-Triazole- PEG2 |
VP-Cys(SOX)-LGGRR |
 |
SOX peptides for differential sensing of MAPKs
While RDF and chimeric sensors reported accurate kinase readouts via pre-organized Mg2+ chelation, discrimination among closely related kinases, including MAPKs, remains challenging. Conversely, an array of SOX peptides can fingerprint kinase isoforms via a single pattern recognition approach, known as differential sensing (DS).27,97,98 Anslyn et al. reported five SOX peptides with distinctive docking sites and selectivities for MAPK families that contain several closely related isoforms. These display ca 80% identity over the catalytic domains. Their array differentiated each of the following families at nanomolar levels (0.5–5 nM) from a single pattern response: ERK1/2, JNK1/2/3 and p38α/β/δ. The discriminatory power of differential sensing was further validated by qualitatively detecting specific inhibitor concentration in one in vitro assay. In 2022, the same team followed the activity of numerous kinases at once (multiplexing) using DS. Retaining the use of four SOX-peptides, they expanded the original array to a much broader one of 14 SOX-peptides in total (Fig. 7). Using machine learning (e.g. principal component analysis (PCA)) they achieved excellent discrimination of nine isoforms of JNK and p38 families. In the DS array, ERK isoform displayed the poorest discrimination, consistent with their 87–88% similar identity. Notably, this viable quantitation method successfully multiplexed the three kinase activities in A549 cell lysates, albeit with increased errors.27,97
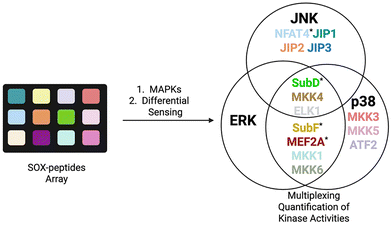 |
| | Fig. 7 Multiplexing of MAP kinase activity with 14 SOX-peptides, using differential sensing. The peptides of the original array are denoted with an asterisk. | |
Rationally designed SOX-peptides for interrogating Arg kinase and phosphatase
While most studies have been performed on protein alcohol (Ser/Thr/Tyr) kinases, protein guanidine (e.g., Arg) kinases also exist.99–102 In Gram-positive bacteria arginine phosphorylation is regulated by Arg kinase McsB and the cognate phosphatase YwlE, which are crucial regulators of stress responses and pathogenesis.103,104 In response to heat stress, McsB phosphorylates arginine to produce phosphoarginine (pArg).105,106 To probe both McsB and YwlE, Kee and coworkers reported SOX-CtsR(-2), a reversible SOX-peptide that reports a five-fold fluorescence increase upon phosphorylation.107 Notably, in the presence of YwlE both the Arg phosphorylation level and fluorescence intensity decreased, validating the dual function of SOX-CtsR(-2) (Ac-KVIQ-Cys(SOX)-K![[R with combining low line]](https://www.rsc.org/images/entities/b_char_0052_0332.gif) GGGGYIK-NH2) (Fig. 8). Arg phosphorylation is dynamically controlled by the opposing actions of McsB and YwlE. Yet, SOX-CtsR(-2) suggests that McsB mediates the dephosphorylation of the pArg residue to generate ATP.107,108 SOX-CtsR(-2) is the first example of a SOX-peptide used to study the interplay between McsB and YwlE. Using this sensor, it was demonstrated that McsB kinetic activity is modulated by factors including autophosphorylation state and nucleotide levels. While counterintuitive, this study revealed that McsB can dephosphorylate pArg even without the presence of YwlE. In addition to these applications, SOX-peptides can also provide insights into such complex mechanisms that until now remained elusive.
GGGGYIK-NH2) (Fig. 8). Arg phosphorylation is dynamically controlled by the opposing actions of McsB and YwlE. Yet, SOX-CtsR(-2) suggests that McsB mediates the dephosphorylation of the pArg residue to generate ATP.107,108 SOX-CtsR(-2) is the first example of a SOX-peptide used to study the interplay between McsB and YwlE. Using this sensor, it was demonstrated that McsB kinetic activity is modulated by factors including autophosphorylation state and nucleotide levels. While counterintuitive, this study revealed that McsB can dephosphorylate pArg even without the presence of YwlE. In addition to these applications, SOX-peptides can also provide insights into such complex mechanisms that until now remained elusive.
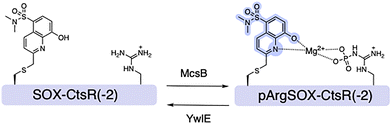 |
| | Fig. 8 Design of SOX-CtsR(−2) sensor for bacterial Arg Kinase McsB and the cognate phosphatase YwlE, and representative spectra of SOX-CtsR(−2) and the phosphorylated form pArgSOX-CtsR(−2) in the presence of Mg2+. | |
Compared to the >500 known kinases, the human genome encodes only ∼30 protein serine/threonine phosphatases (PPs), which have been referred to as the “ugly ducklings” of cellular signalling.6,106,109 However, increasing amounts of evidence suggests that dysregulation of phosphatases can play a critical role in human disease states, including hepatocellular carcinoma (HCC) and the metabolic disorder known as NAFLD.110–113 Being able to continuously monitor their enzymatic activity via fluorescent readouts is desirable for mechanistic investigations. For this purpose, the Stains lab demonstrated that SOX-peptides can be repurposed to afford direct activity assays for protein phosphatases.114,115 Dephosphorylation of the CHEF sensor by a phosphatase results in a decrease in the fluorescence over time, which is directly proportional to the activity of the target enzyme (Fig. 9). The first proof-of principle probe termed PT1Btide-pS3 (ARDI-![[p with combining low line]](https://www.rsc.org/images/entities/b_char_0070_0332.gif) -R-Cys(SOX)-FFRKG), was based on the autophosphorylation sequence of ALK, a known substrate for protein tyrosine phosphatase-1B (PTP1B).114,116,117 PT1Btide-pS3 can detect as little as 12 pg full-length recombinant PTP1B in a remarkably selective trend.114 This optimal chemosensor also provided clear evidence for elevated PTP1B catalytic activity in a rat model of NAFLD disease that afflicts 20–30% of the US population.115 The generality of this approach was further investigated with the PSPtide sensor (Ac-DRRV-pS-V-Cys(SOX)-NH2).118,119 Utilizing this sensor the temporal dynamics of protein phosphatase-2A (PP2A) activity during insulin stimulation of liver hepatocytes were profiled.120–122 Background subtraction of any off-target serine/threonine phosphatase activity using calyculin A allowed isolation of inhibitor-sensitive phosphatase activity.114,116,123,124 Assignment to PP2A was further supported by borthogonal validation through siRNA knockdown and immunodepletion, which reliably quantified PP2A activity in unfractionated cell lysates.116 This validated assay demonstrated a significant modulation of PP2A activity across a panel of human carcinoma cell lines (HCT116, HeLa and HepG2).114 Another Cys(SOX)-based phosphatase CHEF probe is the pPEST1tide (Ac-YDEDFpYD-Cys(SOX)-EF-NH2) for monitoring the phosphatase non-receptor type 12 (PTPN12 or PTP-PEST) activity.125 Gratifyingly, decreasing concentrations of PTP-PEST yielded a limit of detection of 0.2 nM with the pPEST1tide. This level of sensitivity is comparable with PT1Btide-pS3 and PSPtide. However, the applicability of pPEST1tide in interrogating the role of PTP-PEST in HCC and other aggressive tumors remains underexplored.111,125,126
-R-Cys(SOX)-FFRKG), was based on the autophosphorylation sequence of ALK, a known substrate for protein tyrosine phosphatase-1B (PTP1B).114,116,117 PT1Btide-pS3 can detect as little as 12 pg full-length recombinant PTP1B in a remarkably selective trend.114 This optimal chemosensor also provided clear evidence for elevated PTP1B catalytic activity in a rat model of NAFLD disease that afflicts 20–30% of the US population.115 The generality of this approach was further investigated with the PSPtide sensor (Ac-DRRV-pS-V-Cys(SOX)-NH2).118,119 Utilizing this sensor the temporal dynamics of protein phosphatase-2A (PP2A) activity during insulin stimulation of liver hepatocytes were profiled.120–122 Background subtraction of any off-target serine/threonine phosphatase activity using calyculin A allowed isolation of inhibitor-sensitive phosphatase activity.114,116,123,124 Assignment to PP2A was further supported by borthogonal validation through siRNA knockdown and immunodepletion, which reliably quantified PP2A activity in unfractionated cell lysates.116 This validated assay demonstrated a significant modulation of PP2A activity across a panel of human carcinoma cell lines (HCT116, HeLa and HepG2).114 Another Cys(SOX)-based phosphatase CHEF probe is the pPEST1tide (Ac-YDEDFpYD-Cys(SOX)-EF-NH2) for monitoring the phosphatase non-receptor type 12 (PTPN12 or PTP-PEST) activity.125 Gratifyingly, decreasing concentrations of PTP-PEST yielded a limit of detection of 0.2 nM with the pPEST1tide. This level of sensitivity is comparable with PT1Btide-pS3 and PSPtide. However, the applicability of pPEST1tide in interrogating the role of PTP-PEST in HCC and other aggressive tumors remains underexplored.111,125,126
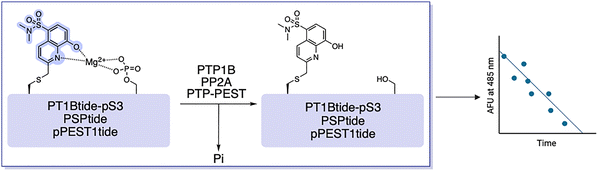 |
| | Fig. 9 Reported SOX-based protein phosphatase sensors. Phosphatase activity removes the phosphate group from the peptide probe and greatly reduces the affinity for Mg2+ leading to a decrease in fluorescence. | |
SOX-peptide technologies now commercially available
SOX-peptides are now commercially available through the Phosphosens® platform from AssayQuant Technologies Inc. (https://www.assayquant.com), who have developed a set of validated assay formats for a broad range of kinases and phosphatases. Their standardized continuous kinetic assays allow direct monitoring of enzyme activity in real time, without the need for coupling enzymes, antibodies, or wash steps, and can be performed on purified recombinant enzymes and cell and tissue lysates. In addition, they have developed a red-shifted “PhosphoSens-Red” format that uses Eu3+ salts to generate a red-shifted time-resolved fluorescence, which helps reduce background interference and compound autofluorescence. However, because Eu3+ is introduced after the kinase reaction, this format converts the assay to an endpoint readout rather than a continuous one, improving practicality for screening while sacrificing the real-time kinetic information available from the original Mg2+-dependent CHEF format.
Conclusions and future outlooks
This review has described how SOX-peptides allow direct, continuous measurement of kinase and phosphatase activity, providing access to kinetic information that is difficult to obtain using conventional endpoint assays. In appropriate systems, these tools offer a practical alternative to antibody- and radioactivity-based methods, particularly for studies in unfractionated lysates and inhibitor screening. Despite these advantages, CHEF probes have several important limitations. The synthesis of SOX-labelled substrates and related fluorophores is not trivial as it involves multiple steps, fragile phenol protecting groups, limited overall yields and often challenging workup and purification steps due to product solubility and sensitivity. Thus, custom SOX probes are not easily accessible and large-scale preparation remains demanding for laboratories without expertise in chemical synthesis. In addition, current CHEF sensors require chemical synthesis by SPPS, which is resource-intensive and generates substantial solvent and reagent waste. Unlike genetically encoded FRET reporters, SOX-peptides have yet to be generated inside cells, limiting their use in long-term or spatially resolved live-cell studies. From an optical perspective, SOX-based probes are also constrained by their short excitation wavelengths (typically 360–365 nm), which increases background interference and phototoxicity in biological samples. For phosphatase assays, additional complications can arise from enzyme sensitivity to oxidation, which can introduce artefacts unless strictly controlled. Several development directions address these issues. One approach is the incorporation of red-shifted fluorophores or lanthanide complexes to enable excitation in the visible range and improve signal-to-noise ratios. Time-resolved fluorescence (TRF) formats using lanthanides (e.g. Tb3+, Eu3+) further reduce background interference by red-shifting the detection wavelength. Another avenue is the design of alternative Mg2+-responsive chromophores that retain the CHEF mechanism but exhibit more favorable photophysical properties. Progress in this area has been limited over the past decade, with relatively few new scaffolds beyond SOX and closely related analogues. Overall, CHEF probes represent a robust platform for real-time enzymatic assays, particularly in biochemical and screening contexts. Broader adoption of lysate-compatible assays can accelerate discovery, profiling and optimization.
Conflicts of interest
There are no conflicts to declare.
Abbreviations
| Abl | Abelson kinase |
| ADP | Adenosine diphosphate |
| Akt | Protein kinase B |
| ALK | Anaplastic lymphoma kinase |
| AOO | 8-amino-3,6-dioxaoctanoic acid |
| ATP | Adenosine triphosphate |
| BTF | β-turn focused |
| Cdk | Cyclin-dependent kinase |
| CHEF | Chelation-enhanced fluorescence |
| DS | Differential sensing |
| eEF-2 | Eukaryotic elongation factor 2 |
| EGFR | Epidermal growth factor receptor |
| ERK | Extracellular signal-regulated kinase |
| FAK | Focal adhesion kinase |
| FDAA | 1-fluoro-2,4-dinitrophenyl-5-L-alanine amide |
| Fmoc | 9-fluorenylmethoxycarbonyl |
| FRET | Förster resonance energy transfer |
| HCC | Hepatocellular carcinoma |
| HTS | High-throughput screening |
| IRK | Insulin receptor kinase |
| JNK | c-Jun N-terminal kinase |
| KM | Michaelis constant |
| LOD | Limit of detection |
| LRRK2 | Leucine-rich repeat kinase 2 |
| MAPK | Mitogen-activated protein kinase |
| McsB | Protein arginine kinase McsB |
| MEF2A | Myocyte enhancer factor 2A |
| MK2 | MAPK-activated protein kinase 2 |
| NAFLD | Non-alcoholic fatty liver disease |
| NBS | N-bromosuccinimide |
| PCA | Principal component analysis |
| PEG | Polyethylene glycol |
| PET | Photoinduced electron transfer |
| pArg | Phosphoarginine |
| PHA665752 | A small-molecule kinase inhibitor |
| Pim2 | Proviral integration site for Moloney murine leukemia virus 2 kinase |
| PK/PD | Pharmacokinetic/pharmacodynamic |
| PKA | Protein kinase A |
| PKC | Protein kinase C |
| PP | Protein phosphatase |
| PP2A | Protein phosphatase 2A |
| PTM | Post-translational modification |
| PTP1B | Protein tyrosine phosphatase 1B |
| PTPN12 | Protein tyrosine phosphatase non-receptor type 12 |
| RDF | Recognition-domain focused |
| ROCK | Rho-associated protein kinase |
| SH2 | Src homology 2 |
| SOX | Sulfonamido-oxine |
| SPPS | Solid-phase peptide synthesis |
| Src | Proto-oncogene tyrosine-protein kinase Src |
| TBDPS | Tert-butyldiphenylsilyl |
| TDI | Time-dependent inhibition |
| TFA | Trifluoroacetic acid |
| Trt | Trityl |
| YwlE | Arginine phosphatase YwlE |
Data availability
No new data were generated or analyzed in this study.
References
- D. Ochoa, A. F. Jarnuczak, C. Viéitez, M. Gehre, M. Soucheray, A. Mateus, A. A. Kleefeldt, A. Hill, L. Garcia-Alonso, F. Stein, N. J. Krogan, M. M. Savitski, D. L. Swaney, J. A. Vizcaíno, K.-M. Noh and P. Beltrao, Nat. Biotechnol., 2020, 38, 365–373 CrossRef CAS PubMed.
- L. B. Peterson, M. B. Yaffe and B. Imperiali, Biochemistry, 2014, 53, 5771–5778 CrossRef CAS PubMed.
- S. Müller-Dott, E. J. Jaehnig, K. P. Munchic, W. Jiang, T. M. Yaron-Barir, S. R. Savage, M. Garrido-Rodriguez, J. L. Johnson, A. Lussana, E. Petsalaki, J. T. Lei, A. Dugourd, K. Krug, L. C. Cantley, D. R. Mani, B. Zhang and J. Saez-Rodriguez, Nat. Commun., 2025, 16, 4771 CrossRef PubMed.
- S. M. Gomez, A. D. Axtman, T. M. Willson, M. B. Major, R. R. Townsend, P. K. Sorger and G. L. Johnson, Drug Discovery Today, 2024, 29, 103881 CrossRef CAS PubMed.
- H. Zhang, X. Cao, M. Tang, G. Zhong, Y. Si, H. Li, F. Zhu, Q. Liao, L. Li, J. Zhao, J. Feng, S. Li, C. Wang, M. Kaulich, F. Wang, L. Chen, L. Li, Z. Xia, T. Liang, H. Lu, X.-H. Feng and B. Zhao, eLife, 2021, 10, e64943 CrossRef CAS PubMed.
- G. Manning, D. B. Whyte, R. Martinez, T. Hunter and S. Sudarsanam, Science, 2002, 298, 1912–1934 CrossRef CAS PubMed.
- S. K. Hanks and T. Hunter, FASEB J., 1995, 9, 576–596 CrossRef CAS.
- M. M. Attwood, D. Fabbro, A. V. Sokolov, S. Knapp and H. B. Schiöth, Nat. Rev. Drug Discovery, 2021, 20, 839–861 CrossRef CAS PubMed.
- R. Roskoski, Pharmacol. Res., 2025, 216, 107723 CrossRef CAS PubMed.
- J. Hu and G. Li, Chem. – Asian J., 2022, 17, e202200182 CrossRef CAS PubMed.
- A. P. Valaka, H. Nyström, L. Håversen, C. Benitez-Martin, C. Schäfer, W. S. Jang, A. Camponeschi, J. Andréasson, J. Borén and M. Grøtli, RSC Chem. Biol., 2025, 6, 618–629 RSC.
- J. A. González-Vera, E. Luković and B. Imperiali, Bioorg. Med. Chem. Lett., 2009, 19, 1258–1260 CrossRef PubMed.
- J. González-Vera and M. Morris, Proteomes, 2015, 3, 369–410 CrossRef PubMed.
- M. D. Shults and B. Imperiali, J. Am. Chem. Soc., 2003, 125, 14248–14249 CrossRef CAS PubMed.
- H. Waldmann and P. Janning, Concepts and case studies in chemical biology, Wiley, Hoboken, 2014 Search PubMed.
- A. Single, H. Beetham, B. J. Telford, P. Guilford and A. Chen, SLAS Discovery, 2015, 20, 1286–1293 CrossRef CAS PubMed.
- H. Ma, S. Deacon and K. Horiuchi, Expert Opin. Drug Discovery, 2008, 3, 607–621 CrossRef CAS PubMed.
- Y. Wang and H. Ma, Drug Discovery Today Technol., 2015, 18, 1–8 CrossRef CAS PubMed.
- O. Kepp, L. Galluzzi, M. Lipinski, J. Yuan and G. Kroemer, Nat. Rev. Drug Discovery, 2011, 10, 221–237 CrossRef CAS PubMed.
- S. Fowler and H. Zhang, AAPS J., 2008, 10, 410–424 CrossRef CAS PubMed.
- H. Bisswanger, Perspect. Sci., 2014, 1, 41–55 CrossRef.
- B. Zhang, H. Chen, L. Shi, R. Guo, Y. Wang, Y. Zheng, R. Bai, Y. Gao, B. Liu and X. Zhang, ACS Sens., 2024, 9, 4560–4567 CrossRef CAS PubMed.
- J. Han, Y. Wang, J. Yu, X. Zhang, Q. Duan, R. Zhang, J. Jing and X. Zhang, Anal. Chem., 2025, 97, 4915–4922 CrossRef CAS PubMed.
- F. Yang, Y. Ma, H. Zou, Y. Liu, X. Li and J. Huang, Sci. Rep., 2025, 15, 42630 CrossRef CAS PubMed.
- M. Khatun, P. Ghorai, J. Mandal, S. Ghosh Chowdhury, P. Karmakar, S. Blasco, E. García-España and A. Saha, ACS Omega, 2023, 8, 7479–7491 CrossRef CAS PubMed.
- Z. D. Nagel, A. A. Beharry, P. Mazzucato, G. J. Kitange, J. N. Sarkaria, E. T. Kool and L. D. Samson, PLoS One, 2019, 14, e0208341 CrossRef CAS PubMed.
- L. Zeng, T. S. Kaoud, D. Zamora-Olivares, A. L. Bohanon, Y. Li, J. R. Pridgen, Y. E. Ekpo, D. L. Zhuang, J. R. Nye, M. Telles, M. Winkler, S. Rivera, F. Marini, K. N. Dalby and E. V. Anslyn, J. Am. Chem. Soc., 2022, 144, 4017–4025 CrossRef CAS PubMed.
- J.-Q. Chen, L. M. Wakefield and D. J. Goldstein, J. Transl. Med., 2015, 13, 182 CrossRef PubMed.
- B. Alberts, R. Heald, K. Hopkin, A. Johnson, D. Morgan, K. Roberts and P. Walter, Essential cell biology, W.W. Norton & Company, New York, 6th edn, 2023 Search PubMed.
- E. Blondeel, S. Ernst, F. De Vuyst, Á. Diósdi, C. Pinheiro, D. Estêvão, P. Rappu, R. Boiy, S. Dedeyne, L. Craciun, V. Goossens, J. Dehairs, T. Cruz, D. Audenaert, W. Ceelen, M. Linnebacher, T. Boterberg, J. Vandesompele, P. Mestdagh, J. Swinnen, J. Heino, P. Horvath, M. J. Oliveira, A. Hendrix, P. Demetter and O. De Wever, Nat. Protoc., 2025, 20, 2899–2941 CrossRef CAS PubMed.
- D. S. Lawrence and Q. Wang, ChemBioChem, 2007, 8, 373–378 CrossRef CAS PubMed.
- Q. Wang and D. S. Lawrence, J. Am. Chem. Soc., 2005, 127, 7684–7685 CrossRef CAS PubMed.
- M. S. Frei, S. Mehta and J. Zhang, Annu. Rev. Biophys., 2024, 53, 275–297 CrossRef CAS PubMed.
- N. W. Nkune, K. Moloudi, B. P. George and H. Abrahamse, RSC Adv., 2025, 15, 22267–22284 RSC.
- G. T. Dempsey, J. C. Vaughan, K. H. Chen, M. Bates and X. Zhuang, Nat. Methods, 2011, 8, 1027–1036 CrossRef CAS PubMed.
- Y. Nasu, Y. Shen, L. Kramer and R. E. Campbell, Nat. Chem. Biol., 2021, 17, 509–518 CrossRef CAS PubMed.
- T. Kagan, M. Gabay, A. Meenakshisundaram, Y. Levi, S. Eid, N. Malchenko, M. Maman, A. Nitzan, L. Ravotto, R. Zaidel-Bar, B. J. Eickholt, M. Gal and T. Laviv, Nat. Methods, 2025, 22, 764–777 CrossRef CAS PubMed.
- F. Brøndsted and L. G. Wang, Nat. Methods, 2025, 22, 1137–1139 CrossRef PubMed.
- C. Ferrag and K. Kerman, Front. Sens., 2020, 1, 583822 CrossRef.
- G. Jamalipour Soufi and S. Iravani, Green Chem., 2020, 22, 2662–2687 RSC.
- P. Koutsogiannis, E. Thomou, H. Stamatis, D. Gournis and P. Rudolf, Adv. Phys.: X, 2020, 5, 1758592 CAS.
- X. Liu, Y. Li, X. Xu, P. Li, Z. Nie, Y. Huang and S. Yao, TrAC, Trends Anal. Chem., 2014, 58, 40–53 CrossRef CAS.
- A. T. Krasley, E. Li, J. M. Galeana, C. Bulumulla, A. G. Beyene and G. S. Demirer, Chem. Rev., 2024, 124, 3085–3185 CrossRef CAS PubMed.
- M. Minoshima, S. I. Reja, R. Hashimoto, K. Iijima and K. Kikuchi, Chem. Rev., 2024, 124, 6198–6270 CrossRef CAS PubMed.
- A. Roy, M. Singha and S. Singha, ChemBioChem, 2025, 26, e202500175 CrossRef CAS PubMed.
- D. Xiao, H. Qi, Y. Teng, D. Pierre, P. T. Kutoka and D. Liu, Nanoscale Res. Lett., 2021, 16, 167 CrossRef CAS PubMed.
- X. Yan, H. Li, W. Zheng and X. Su, Anal. Chem., 2015, 87, 8904–8909 CrossRef CAS PubMed.
- W. Cui and L. L. Parker, Chem. Commun., 2015, 51, 362–365 RSC.
- S. Fernández-Fariña, O. Kotova, S. R. Donohoe and T. Gunnlaugsson, Chem. Soc. Rev., 2025, 54, 11226–11265 RSC.
- J. Li, X. Tian, W. Bao, J. Zhao and Z. Jiang, Discovery Appl. Sci., 2025, 7, 1310 CrossRef CAS.
- A. Ramström, D. R. S. Pooler, H. Abasov, M. Tomar, S. Crespi and F. Schaufelberger, Angew. Chem., Int. Ed., 2025, 64, e202505666 CrossRef PubMed.
- M. Ahmadi, Z. Zhao and I. J. Dmochowski, Chem. Sci., 2025, 16, 8125–8135 RSC.
- J. Liao, Y. Song and Y. Liu, Acta Pharmacol. Sin., 2015, 36, 1408–1415 Search PubMed.
- M. D. Allen, L. M. DiPilato, B. Ananthanarayanan, R. H. Newman, Q. Ni and J. Zhang, Sci. Signaling, 2008, 1, pt6 CrossRef PubMed.
- T. Mizutani, T. Kondo, S. Darmanin, M. Tsuda, S. Tanaka, M. Tobiume, M. Asaka and Y. Ohba, Clin. Cancer Res., 2010, 16, 3964–3975 CrossRef CAS PubMed.
- A. Y. Ting, K. H. Kain, R. L. Klemke and R. Y. Tsien, Proc. Natl. Acad. Sci. U. S. A., 2001, 98, 15003–15008 CrossRef CAS PubMed.
- N. D. Rawlings and A. J. Barrett, Encyclopedia of Life Sciences, Wiley, 2014 Search PubMed.
- A. K. Carmona, M. A. Juliano and L. Juliano, An. Acad. Bras. Ciênc., 2009, 81, 381–392 Search PubMed.
- J. Wang, Q. Meng, Y. Yang, S. Zhong, R. Zhang, Y. Fang, Y. Gao and X. Cui, ACS Sens., 2022, 7, 2521–2536 CrossRef CAS PubMed.
- K. P. Carter, A. M. Young and A. E. Palmer, Chem. Rev., 2014, 114, 4564–4601 CrossRef CAS PubMed.
- Principles of Fluorescence Spectroscopy, ed J. R. Lakowicz, Springer, Boston, 3rd edn, 2006 Search PubMed.
- B. Valeur and M. N. Berberan-Santos, Molecular Fluorescence: Principles and Applications, Wiley, 1st edn, 2012 Search PubMed.
- H. Lee, H.-S. Lee, J. H. Reibenspies and R. D. Hancock, Inorg. Chem., 2012, 51, 10904–10915 CrossRef CAS PubMed.
- C. E. Housecroft and A. G. Sharpe, Inorganic chemistry, Pearson, Harlow, 5th edn, 2018 Search PubMed.
- M. E. Huston, K. W. Haider and A. W. Czarnik, J. Am. Chem. Soc., 1988, 110, 4460–4462 CrossRef CAS.
- M. E. Huston, E. U. Akkaya and A. W. Czarnik, J. Am. Chem. Soc., 1989, 111, 8735–8737 CrossRef CAS.
- E. U. Akkaya, M. E. Huston and A. W. Czarnik, J. Am. Chem. Soc., 1990, 112, 3590–3593 CrossRef CAS.
- D. H. Vance and A. W. Czarnik, J. Am. Chem. Soc., 1994, 116, 9397–9398 CrossRef CAS.
- M. D. Shults, D. Carrico-Moniz and B. Imperiali, Anal. Biochem., 2006, 352, 198–207 CrossRef CAS PubMed.
- M. D. Shults, D. A. Pearce and B. Imperiali, J. Am. Chem. Soc., 2003, 125, 10591–10597 CrossRef CAS PubMed.
- R. Rahier, A. Noiriel and A. Abousalham, Anal. Chem., 2016, 88, 666–674 Search PubMed.
- Y.-X. Sun, X.-X. Li, L.-L. Gan, M.-X. Du, B. Zhao, W.-D. Li, J.-G. Li and W.-K. Dong, Chem. Pap., 2024, 78, 7001–7013 CrossRef CAS.
- A. T. Krueger and B. Imperiali, ChemBioChem, 2013, 14, 788–799 CrossRef CAS PubMed.
- D. A. Pearce, N. Jotterand, I. S. Carrico and B. Imperiali, J. Am. Chem. Soc., 2001, 123, 5160–5161 CrossRef CAS PubMed.
- M. D. Shults, K. A. Janes, D. A. Lauffenburger and B. Imperiali, Nat. Methods, 2005, 2, 277–284 CrossRef CAS PubMed.
- J. A. González-Vera, Chem. Soc. Rev., 2012, 41, 1652–1664 RSC.
- E. Luković, J. A. González-Vera and B. Imperiali, J. Am. Chem. Soc., 2008, 130, 12821–12827 Search PubMed.
- C. I. Stains, E. Luković and B. Imperiali, ACS Chem. Biol., 2011, 6, 101–105 CrossRef CAS PubMed.
- E. Luković, E. Vogel Taylor and B. Imperiali, Angew. Chem., Int. Ed., 2009, 48, 6828–6831 CrossRef PubMed.
- J. R. Beck, L. B. Peterson, B. Imperiali and C. I. Stains, Curr. Protoc. Chem. Biol., 2014, 6, 10–18 Search PubMed.
- A. K. Devkota, M. Warthaka, R. Edupuganti, C. D. J. Tavares, W. H. Johnson, B. Ozpolat, E. J. Cho and K. N. Dalby, SLAS Discovery, 2014, 19, 445–452 CrossRef CAS PubMed.
- C. C. Ayala-Aguilera, T. Valero, Á. Lorente-Macías, D. J. Baillache, S. Croke and A. Unciti-Broceta, J. Med. Chem., 2022, 65, 1047–1131 CrossRef CAS PubMed.
- D. A. Szalewski, J. R. Beck and C. I. Stains, Bioorg. Med. Chem. Lett., 2014, 24, 5648–5651 CrossRef CAS PubMed.
- J. R. Beck, X. Zhou, G. R. Casey and C. I. Stains, Anal. Chim. Acta, 2015, 897, 62–68 CrossRef CAS PubMed.
- R. G. Silva, K. F. Geoghegan, X. Qiu and A. Aulabaugh, Anal. Biochem., 2014, 450, 63–69 CrossRef CAS PubMed.
- M. I. Kelly, T. J. Bechtel, D. R. Reddy, E. D. Hankore, J. R. Beck and C. I. Stains, Anal. Chim. Acta, 2015, 891, 284–290 CrossRef CAS PubMed.
- G.-H. Nam, E. J. Lee, Y. K. Kim, Y. Hong, Y. Choi, M.-J. Ryu, J. Woo, Y. Cho, D. J. Ahn, Y. Yang, I.-C. Kwon, S.-Y. Park and I.-S. Kim, Nat. Commun., 2018, 9, 2165 CrossRef PubMed.
- Q. Rui, H. Ni, D. Li, R. Gao and G. Chen, Curr. Neuropharmacol., 2018, 16, 1348–1357 CrossRef CAS PubMed.
- D. Gnani, I. Romito, S. Artuso, M. Chierici, C. De Stefanis, N. Panera, A. Crudele, S. Ceccarelli, E. Carcarino, V. D’Oria, M. Porru, E. Giorda, K. Ferrari, L. Miele, E. Villa, C. Balsano, D. Pasini, C. Furlanello, F. Locatelli, V. Nobili, R. Rota, C. Leonetti and A. Alisi, Cell Death Differ., 2017, 24, 889–902 CrossRef CAS PubMed.
- E. J. Goldsmith, R. Akella, X. Min, T. Zhou and J. M. Humphreys, Chem. Rev., 2007, 107, 5065–5081 CrossRef CAS PubMed.
- H.-M. Lee, D. R. Larson and D. S. Lawrence, ACS Chem. Biol., 2009, 4, 409–427 CrossRef CAS PubMed.
- D. M. Rothman, M. D. Shults and B. Imperiali, Trends Cell Biol., 2005, 15, 502–510 CrossRef CAS PubMed.
- E. F. Wagner and Á. R. Nebreda, Nat. Rev. Cancer, 2009, 9, 537–549 CrossRef CAS PubMed.
- C. Prével, M. Pellerano, T. N. N. Van and M. C. Morris, Biotechnol. J., 2014, 9, 253–265 CrossRef PubMed.
- M. C. Lawrence, A. Jivan, C. Shao, L. Duan, D. Goad, E. Zaganjor, J. Osborne, K. McGlynn, S. Stippec, S. Earnest, W. Chen and M. H. Cobb, Cell Res., 2008, 18, 436–442 CrossRef CAS PubMed.
- Y. J. Feng and Y. Y. Li, J. Dig. Dis., 2011, 12, 327–332 CrossRef CAS PubMed.
- D. Zamora-Olivares, T. S. Kaoud, J. Jose, A. Ellington, K. N. Dalby and E. V. Anslyn, Angew. Chem., Int. Ed., 2014, 53, 14064–14068 CrossRef CAS PubMed.
- B. A. Rose, T. Force and Y. Wang, Physiol. Rev., 2010, 90, 1507–1546 Search PubMed.
- S. Fu, C. Fu, Q. Zhou, R. Lin, H. Ouyang, M. Wang, Y. Sun, Y. Liu and Y. Zhao, Sci. China: Chem., 2020, 63, 341–346 CrossRef CAS.
- A. K. W. Elsholz, K. Turgay, S. Michalik, B. Hessling, K. Gronau, D. Oertel, U. Mäder, J. Bernhardt, D. Becher, M. Hecker and U. Gerth, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, 7451–7456 CrossRef CAS PubMed.
- F. Falcioni, R. W. Molt, Y. Jin, J. P. Waltho, S. Hay, N. G. J. Richards and G. M. Blackburn, ACS Catal., 2024, 14, 6650–6658 CrossRef CAS PubMed.
- D. B. Trentini, M. J. Suskiewicz, A. Heuck, R. Kurzbauer, L. Deszcz, K. Mechtler and T. Clausen, Nature, 2016, 539, 48–53 CrossRef CAS PubMed.
- J. Fuhrmann, B. Mierzwa, D. B. Trentini, S. Spiess, A. Lehner, E. Charpentier and T. Clausen, Cell Rep., 2013, 3, 1832–1839 CrossRef CAS PubMed.
- M. J. Suskiewicz, B. Hajdusits, R. Beveridge, A. Heuck, L. D. Vu, R. Kurzbauer, K. Hauer, V. Thoeny, K. Rumpel, K. Mechtler, A. Meinhart and T. Clausen, Nat. Chem. Biol., 2019, 15, 510–518 CrossRef CAS PubMed.
- H. Ouyang, C. Fu, S. Fu, Z. Ji, Y. Sun, P. Deng and Y. Zhao, Org. Biomol. Chem., 2016, 14, 1925–1929 RSC.
- D. B. Trentini, J. Fuhrmann, K. Mechtler and T. Clausen, Mol. Cell. Proteomics, 2014, 13, 1953–1964 CrossRef CAS PubMed.
- H. Jung, Y. Choi, D. Lee, J. K. Seo and J.-M. Kee, Chem. Commun., 2019, 55, 7482–7485 RSC.
- B. Hajdusits, M. J. Suskiewicz, N. Hundt, A. Meinhart, R. Kurzbauer, J. Leodolter, P. Kukura and T. Clausen, eLife, 2021, 10, e63505 CrossRef CAS PubMed.
- D. L. Brautigan, FEBS J., 2013, 280, 324–325 CrossRef CAS PubMed.
- S. Pouwels, N. Sakran, Y. Graham, A. Leal, T. Pintar, W. Yang, R. Kassir, R. Singhal, K. Mahawar and D. Ramnarain, BMC Endocr. Disord., 2022, 22, 63 CrossRef CAS PubMed.
- Z. M. Younossi, P. Golabi, J. M. Paik, A. Henry, C. Van Dongen and L. Henry, Hepatology, 2023, 77, 1335–1347 CrossRef PubMed.
- P. Ye, J. Liu, W. Xu, D. Liu, X. Ding, S. Le, H. Zhang, S. Chen, M. Chen and J. Xia, Hepatology, 2019, 69, 1946–1964 CrossRef CAS PubMed.
- J.-S. Yoon and C.-W. Lee, Exp. Mol. Med., 2022, 54, 1799–1813 CrossRef CAS PubMed.
- J. R. Beck, T. Truong and C. I. Stains, ACS Chem. Biol., 2016, 11, 3284–3288 CrossRef CAS PubMed.
- G. R. Casey, J. R. Beck and C. I. Stains, Methods Enzymology, Elsevier, 2019, vol. 622, pp. 29–53 Search PubMed.
- G. R. Casey and C. I. Stains, Chem. – Eur. J., 2018, 24, 7810–7824 CrossRef CAS PubMed.
- M. C. Boutterin, P. Mazot, C. Faure, S. Doly, N. Gervasi, M. L. Tremblay and M. Vigny, Cell Signal., 2013, 25, 1505–1513 CrossRef CAS PubMed.
- J. R. Beck, A. Lawrence, A. S. Tung, E. N. Harris and C. I. Stains, ACS Chem. Biol., 2016, 11, 284–290 CrossRef CAS PubMed.
- K. A. Kalesh, L. P. Tan, K. Lu, L. Gao, J. Wang and S. Q. Yao, Chem. Commun., 2010, 46, 589–591 RSC.
- A. Brewer, G. Sathe, B. E. Pflug, R. G. Clarke, T. J. Macartney and G. P. Sapkota, iScience, 2024, 27, 109302 CrossRef CAS PubMed.
- J. Oh, Y. Park, S. Park, O.-G. Woo, J.-H. Lee, J.-S. Lee and T. Kim, Mol. Cells, 2025, 48, 100255 CrossRef CAS PubMed.
- M. Elchebly, P. Payette, E. Michaliszyn, W. Cromlish, S. Collins, A. L. Loy, D. Normandin, A. Cheng, J. Himms-Hagen, C.-C. Chan, C. Ramachandran, M. J. Gresser, M. L. Tremblay and B. P. Kennedy, Science, 1999, 283, 1544–1548 CrossRef CAS PubMed.
- M. J. Peake, M. Pejakovic and G. H. White, J. Clin. Pathol., 1988, 41, 202–206 CrossRef CAS PubMed.
- A. Donella-Deana, M. H. Krinks, M. Ruzzene, C. Klee and L. A. Pinna, Eur. J. Biochem., 1994, 219, 109–117 CrossRef CAS PubMed.
- G. R. Casey and C. I. Stains, Analyst, 2020, 145, 6713–6718 RSC.
- Z. Chen, J. E. Morales, P. A. Guerrero, H. Sun and J. H. McCarty, Cancer Res., 2018, 78, 3809–3822 CrossRef CAS PubMed.
|
| This journal is © The Royal Society of Chemistry 2026 |
Click here to see how this site uses Cookies. View our privacy policy here. 
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a  and
Stephen A. Cochrane
and
Stephen A. Cochrane











