
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Itaconate and its derivatives as anti-pathogenic agents
Rizkallah Al Akiki Dit Al Mazraania,
Naglis Malys *ab and
Vida Malienec
*ab and
Vida Malienec
aBioprocess Research Centre, Faculty of Chemical Technology, Kaunas University of Technology, Radvilėnų st. 19, Kaunas, LT-50254, Lithuania
bDepartment of Organic Chemistry, Faculty of Chemical Technology, Kaunas University of Technology, Radvilėnų st. 19, Kaunas, LT-50254, Lithuania. E-mail: n.malys@gmail.com
cBuilt Environment and Sustainable Technologies Research Institute, Faculty of Health, Innovation, Technology and Science, Liverpool John Moores University, Byrom Street, Liverpool L3 3AF, UK
First published on 10th February 2025
Abstract
Pathogenic microorganisms and viruses cause outbreaks and pandemics that affect millions of people worldwide. Despite recent advances in pharmacology and medicine, the ability of infectious diseases to spread in the modern era is accelerating due to various factors contributing to increased human-to-human and human–animal contacts. With the global rise of drug resistance among pathogens and frequently occurring viral outbreaks, alternative drugs and therapies that specifically inhibit microbial virulence or regulate immune responses are attracting growing interest. The present review focuses on itaconate and its derivatives as potential anti-pathogenic agents. It summarizes the current state of research on itaconate metabolism in bacteria, fungi and mammals. This is followed by a comprehensive review of recent advances studying itaconate and its derivatives as anti-inflammatory, immunoregulatory, antimicrobial and antiviral compounds, along with their mechanisms of action. Finally, the review emphasises the existing challenges and future research directions for the application of itaconate and its derivatives as anti-pathogenic agents.
Introduction
Itaconate is a versatile compound with a wide range of applications, including its use as a monomer in the synthesis of plastics and food packaging.1–3 Importantly, it exhibits anti-pathogenic4,5 and immunoregulatory properties.6 Itaconate was originally discovered by Gustav Crasso and Jean Louis Laissaigne around 1840,7 who revealed that itaconate is formed from aconitate while studying the thermal decomposition of citric acid. The itaconate biosynthesis by the fungus Aspergillus was first reported nearly a hundred years later in 1931.8–10 Furthermore, the biosynthesis of itaconate at an industrial scale has been demonstrated through the whole-cell bioconversion of citrate to itaconate in engineered Escherichia coli.11,12 The chemical synthesis of itaconate by the carbonization of citric acid, followed by hydrolyzation of the anhydride, has been attempted to increase the yield for industrial needs.13 However, no chemical method can rival fungal production of itaconate.14Over a decade ago, mammalian immune cells, specifically macrophages, were shown to produce itaconate in mitochondria in large quantities in response to activation by the inflammatory stimulus lipopolysaccharide (LPS).15–17 The cis-aconitate decarboxylase (ACOD1), encoded by the immunoresponsive gene 1 (Irg1), was identified as the enzyme responsible for catalysing the decarboxylation of cis-aconitate, a tricarboxylic acid (TCA) cycle intermediate, to itaconate.18 Since its emergence as a mitochondrial metabolite with inflammation-regulatory properties, interest in itaconate has grown significantly.19–23
With the increase in pathogen resistance to conventional medical treatments and the recent severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) pandemic,24–28 the immunomodulatory, antimicrobial and anti-pathogenic properties of itaconate have attracted substantial research.29–33 Itaconate and its derivatives have also been demonstrated to possess antiviral properties.22
This review will delve into the anti-pathogenic properties of itaconate and its derivatives. Their inhibitory effects will be discussed in detail, although not all mechanisms of action are fully understood. The medicinal potential of itaconate and its derivatives as anti-inflammatory, antimicrobial and antiviral agents will be explored. Additionally, we will examine recent discoveries that may shed light on the mode of action of itaconate derivatives as antiviral agents, potentially explaining previously observed effects where the mechanisms were unknown.
Itaconate's chemical properties in the biological context
Itaconate (1) is an α,β-unsaturated dicarboxylic acid (C5H6O4) with a characteristic double bond and two carboxyl groups. The unsaturated double bond can accept an electron pair, enabling it to act as an acceptor in the Michael reaction.34 Due to this chemical property, itaconate can alkylate the cysteine residue (Fig. 1) of the peptide.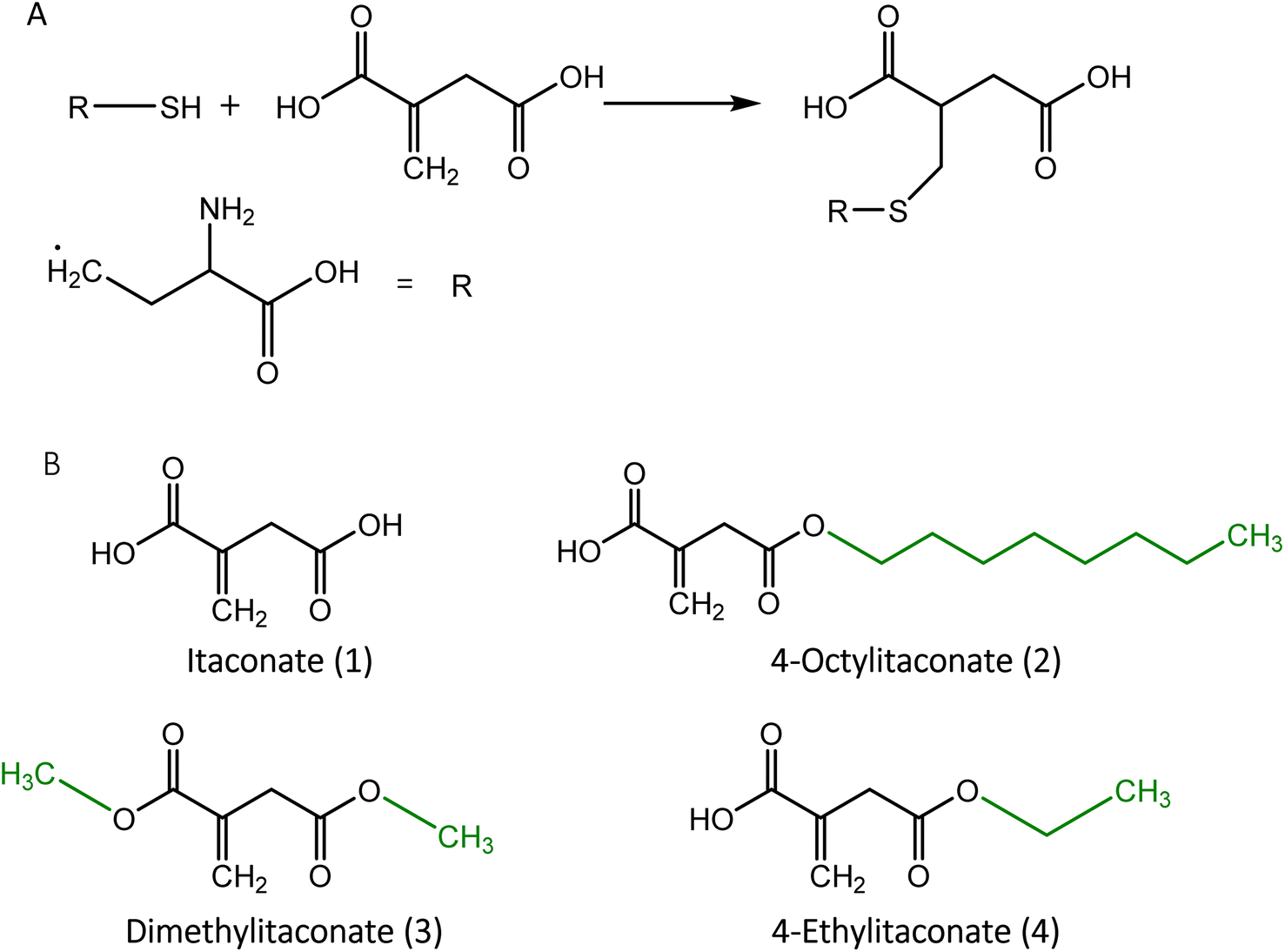 | ||
| Fig. 1 (A) Alkylation of cysteine by itaconate. (B) Structural representation of itaconate and its derivatives. Residues used to generate itaconate derivatives are shown in green. | ||
Similar to other dicarboxylic acids, itaconate exhibits strong polarity and hydrophilicity, causing weak permeability across the membrane,35 which hinders the investigation of its effects in the cell. To improve itaconate's ability to enter the cell, esterification was applied to derive substitute compounds such as 4-octylitaconate (2) (4-OI),35 dimethylitaconate (3) (DI)19 and 4-ethylitaconate (4) (4-EI).36,37 The most studied derivative, 4-OI, was found to be converted to itaconate upon entry into the cell by the esterase,36 whereas DI exhibited a short-term effect without the formation of itaconate and was metabolized rapidly in the cell.38 DI also showed a wide spectrum of effects on metabolism caused by the covalent modification of metabolic enzymes.37 Although 4-EI has a similar structure and polarity to DI, its effect was less pronounced in the cell.39
Chemical synthesis of itaconate and its derivatives
Various chemical methods were developed for the synthesis of itaconate. The pyrolysis of citric acid followed by hydrolysis of itaconic anhydride (Fig. 2) was one of the early methods developed by Baup.40 Later, the carboxylation of aconitic acid was introduced by Crasso.7 Other variations of chemical synthesis of itaconate were developed. However, neither of these chemical synthesis methods cannot rival the microbial biosynthesis using fungi, which is currently preferred for the commercial production of itaconate.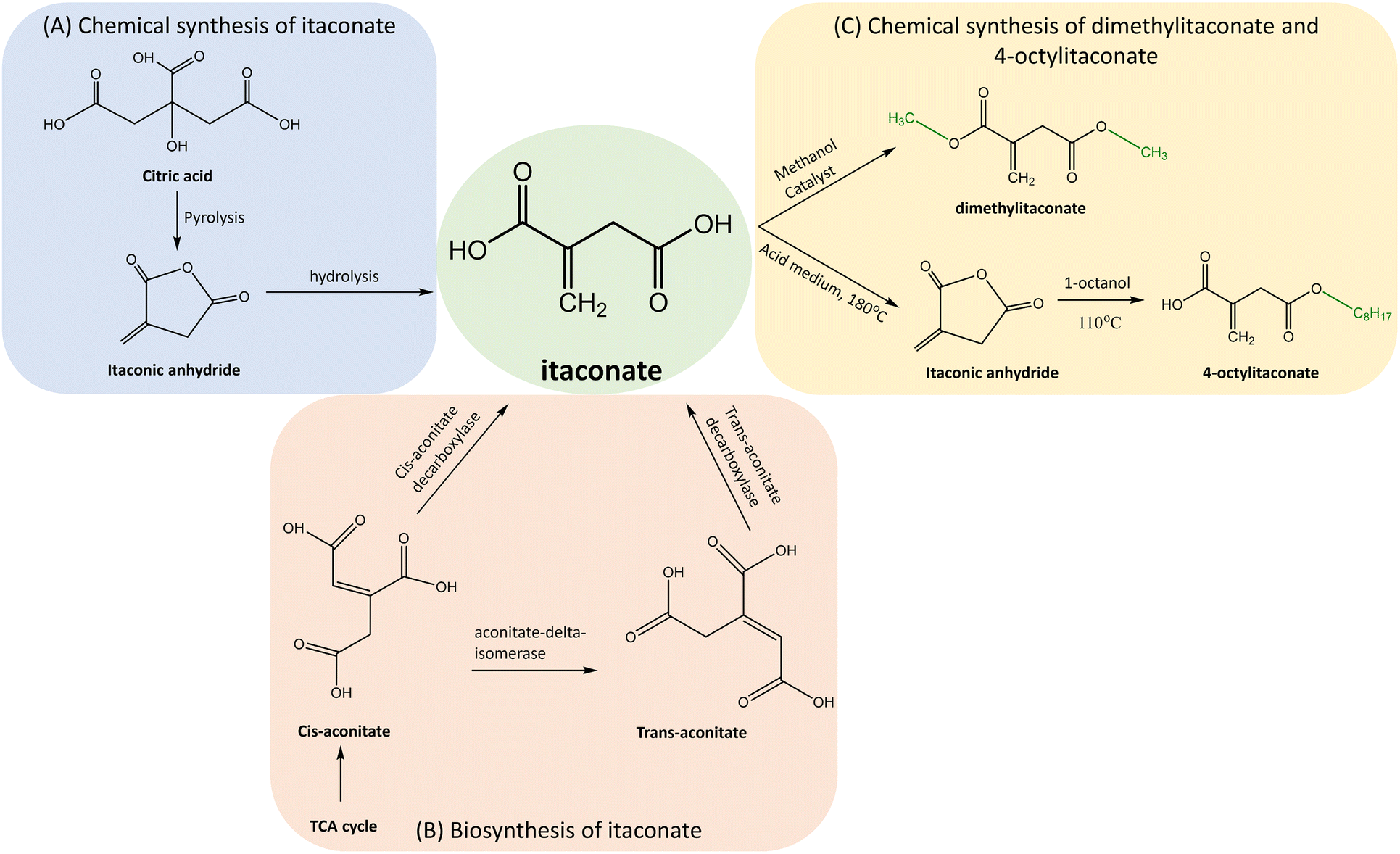 | ||
| Fig. 2 Synthesis of itaconate and its derivatives. (A) Chemical synthesis of itaconate by pyrolysis of citric acid followed by hydrolysis of itaconic anhydride. (B) Biosynthesis of itaconate in fungi from cis-aconitate by cis-aconitate decarboxylase, either directly or by passing through trans-aconitate. (C) Chemical synthesis of itaconate derivatives DI and 4-OI by esterification of itaconate. | ||
The itaconate derivatives such as 4OI and DI can be chemically synthesised by esterification of itaconate. The early synthesis of 4-OI described by Gargallo et al.41 was based on one-step esterification by 1-octanol and using an acid as a catalyst. This method was not environment-friendly and presented a low yield of 35%. Therefore, a two-step method was developed.42 It involved convertion of itaconate to itaconic anhydride using sulphuric acid (H2SO4) as a catalyst at a temperature of 180 °C and pressure of 10 mm Hg followed by anhydride's esterification to 4-OI by 1-octanol at a temperature of 110 °C. Although the yield increased to 95% and selectivity for the derivative was 94%, the itaconic anhydride was susceptible to the hydrolysis and the production cost was high. In order to reduce the cost and increase the selectivity, a novel method was developed.43 The esterification by 1-octanol was applied in the presence of toluene and immobilized lipase (Novozyme 435) at a temperature of 50 °C achieving the 4-OI's yield of 99% with a 99% selectivity.
DI can also be synthesized by the esterification of itaconate with methanol and using H2SO4 as the catalyst.44 Other catalysts such as La3+ ∼ SO42−/TiO2–SiO2 and Ce4+ ∼ SO42−/TiO2–SiO2 were applied to achieve the yield of 94.31% and 93.43%, respectively.
Metabolism of itaconate
 | ||
| Fig. 3 Biosynthesis of itaconate in macrophages. The TLR4 receptor, activated by LPS from Gram-negative bacteria, triggers the upregulation of Irg1 expression, encoding ACOD1, which leads to itaconate biosynthesis in macrophages. | ||
Despite shared transport mechanisms for itaconate production in both fungi, their metabolic pathways diverge. A. terreus features cis-aconitate decarboxylase,52 whereas U. maydis expresses aconitate-delta-isomerase in conjunction with trans-aconitate decarboxylase for the conversion of cis-aconitate to itaconate51 (Fig. 2).
Metabolic engineering for the biosynthesis of itaconate
Itaconate has been commercially produced using the filamentous fungus A. terreus.56 Despite achieving a 60% yield, the interest in improving the productivity of this process for industrial use remains high. Several microorganisms, including A. terreus, Ustilago sp. and Candida sp., have the capability to produce itaconic acid, but their production levels are low using glucose as a carbon source.55,57–60 The yield of itaconate varies depending on several factors, mainly the carbon source. When mannose was used with A. terreus, the yield of itaconate was 0.46 g g−1;59 however, when glucose was used as the carbon source with A. terreus, the yield increased to 0.62 g g−1.61 Therefore, in this section, we will discuss the metabolic engineering methods used to synthesize itaconate.To address industrial needs, recombinant production strategies were applied for itaconate production. This involved genetic engineering and overexpression of A. terreus genes in E. coli62 and Corynebacterium glutamicum.63 Though the native E. coli lacks the cis-aconitate decarboxylase gene and ability to synthesize itaconate,12 it was engineered for this dicarboxylic acid production. In one of the studies, itaconate synthesis was achieved using glucose as a substrate and integrating the cis-aconitate decarboxylase gene cad1 from A. terreus.62 In addition, genes of citrate synthase and aconitase from C. glutamicum were heterologously expressed and the gene encoding lactate dehydrogenase was inactivated, resulting in an itaconate yield of 0.09 mol per mol glucose.62 In another study, a modified strategy was explored by integrating aconitase gene from C. glutamicum and cad1 from A. terreus and utilizing citric acid as a source.
C. glutamicum, which naturally tolerates itaconate well, was also engineered for the production of this compound.63,64 By reducing isocitrate dehydrogenase activity and integrating cad1 gene from A. terreus, the yields of 0.29 and 0.02 g g−1 were achieved using glucose and acetate as carbon sources, respectively.
Anti-pathogenic activities of itaconate and its derivatives
 | ||
| Fig. 4 Effects of itaconate on anti-inflammatory signalling pathways. Itaconate (1) alkylates Cys151 of KEAP1, leading to the dissociation of the KEAP1–Nrf2 complex and subsequent translocation of Nrf2 to the cell nucleus, (2) increases ATF3 protein levels, facilitating its translocation to the cell nucleus, where it inhibits IκBζ at the translational level, (3) disrupts the NLRP3–NEK7 interaction through a modification known as decarboxypropylation on C548 of NLRP3, thereby preventing NLRP3-dependent IL-1β release, (4) inhibits glycolysis by alkylating Cys22 of GAPDH, (5) inhibits succinate dehydrogenase due to its structural resemblance to succinate, and (6) alkylates STING, causing a decrease in its phosphorylation and exerting an anti-inflammatory effect. | ||
Itaconate was also shown to induce the electrophilic stress response. However, this compound showed a less immunosuppressive phenotype compared to DI and 4-OI.37 The comparative analysis of itaconate and its derivatives revealed divergent inflammasome and immune response in macrophages. By identifying itaconate as an immunoregulatory metabolite, this study highlights the importance of using the unmodified form of this compound in future studies.37
In conclusion, based on what was discussed and presented in Fig. 3, itaconate and its derivatives were able to inhibit the inflammation by binding covalently to the cysteine of proteins KEAP1, NLRP3 and GAPDH or by binding to AFT3, SDH and STING, leading to the decrease of the synthesis of interleukin IL-1β and IL-6. Interestingly, in a recent study, DI showed enhanced inflammation.79 This pro- and anti-inflammatory dichotomy of DI highlights the complexity of immune responses when considering derivatives of itaconate for therapeutic application.
Antimicrobial properties of itaconate
Itaconate was proven to possess antimicrobial properties by inhibiting the bacterial growth.80 Itaconate inhibits key enzymes from the glyoxylate cycle such as isocitrate lyase and propionyl CoA-carboxylase, which leads to the inhibition of bacterial growth.81,82 In addition, itaconate was shown to activate the biosynthesis of lysosome in macrophages for the phagocytosis of pathogen.83 The mechanism of inhibition of enzymes as well as the activation of the biosynthesis of lysosome is explained below. | ||
| Fig. 5 Antimicrobial effect of itaconate is shown in (A) activating macrophages and inducing the synthesis of lysosomal biogenesis by activating TFEB, and (B) inhibiting key enzymes in the glyoxylate shunt, (1) isocitrate lyase and (2) propionyl-CoA carboxylase. | ||
In addition to inhibiting isocitrate lyase, itaconate was reported to decrease the assimilation of acetate and propionate by inhibiting the activity of the propionyl-CoA carboxylase (PCAC)82 (Fig. 4B) in Rhodospirillum rubrum. Itaconate selectively inhibits the oxidation of propionate by reducing oxygen intact and competitively inhibiting PCAC since the precursor of propionate, succinate, was not inhibited. Based on the obtained data, itaconate can target other enzymes involved in the conversion of propionyl-CoA to succinate, including methylmalonyl-CoA epimerase, methylmalonyl-CoA mutase and succinate thiokinase. However, the exact mechanism of action was not fully characterized in this study.82
Other itaconate-related mechanisms contributing to the inhibition of pathogen growth, including those restricting the replication, have been recently studied.32,54,85 Studies on Salmonella typhimurium, Mycobacterium avium and other pathogenic bacteria provide links between itaconate produced in the mitochondria after stimulation of innate immune receptors and cell defence mechanisms restricting the propagation of intracellular pathogens.
In summary, the antimicrobial properties of itaconate extends from the direct inhibition of key bacterial metabolic pathways, targeting enzymes such as isocitrate lyase and propionyl CoA-carboxylase, to an indirect effect by triggering the synthesis of lysosomes in macrophages to activate the phagocytosis of bacteria.
Itaconate and its derivatives as immunoregulators
The immunoregulatory role of itaconate and its derivatives has come to the attention recently with relevant research still in its pioneering phase.37,39,87–89 They have been recognized to suppress the inflammatory response in pro-inflammatory M1 macrophages. Furthermore, Runtsch et al. demonstrated that itaconate and 4-OI impede M2 macrophage polarization and metabolic remodelling by inhibiting JAK1 (Janus kinase 1) and STAT6 (signal transducer and activator of transcription 6) phosphorylation.90 4-OI was shown to modify JAK1 at multiple residues including cysteines 715, 816, 943, and 1130, leading to this kinase inhibition. Using another derivative of itaconate, the DI, it was shown that a long-term trained immunity can be induced.79 DI was able to alter the central metabolism and mitochondrial energetics at the transcriptional, epigenomic, and metabolic levels, leading to increased responsiveness and survival to infection of pathogens such as Staphylococcus aureus. In addition, naturally occurring isomers mesaconate and citraconate, which have similar strong electrophile properties to itaconate, were revealed as immunoregulatory compounds.48Altogether, recent pioneering studies on itaconate and its derivatives have shown their immunoregulatory potential in the treatment of immune-mediated diseases including allergy, asthma and fibrosis by targeting macrophages.
Anti-viral properties of itaconate and its derivatives
Itaconate and its derivatives have been reported to inhibit the replication of multiple viruses including influenza A, SARS-CoV-2, PRRSV (porcine reproductive and respiratory syndrome virus) and Zika.20 The antiviral effects of these compounds studied so far are presented in Fig. 6. Itaconate derivatives 4-OI and DI have gained the most attention as potential antiviral agents due to their cell-permeable properties. Moreover, itaconate and its naturally occurring isomers citramalate and mesaconate were shown to modulate immune response and amino acid metabolism, leading to the reduced release of viral particles of influenza A.48 | ||
| Fig. 6 Antiviral effect of itaconate and its derivatives. (A) Neuron cells by inhibiting SDH. (B) Epithelial, kidney, and lung cells by inhibiting viral replication, which can be explained by the inhibition of CRM1 in influenza A, and may involve a similar mechanism for SARS-CoV-2, PRRSV and other viruses. | ||
Considering the importance of CRM1 function to various pathogenic viruses, itaconate and its derivatives can be applied to the formulation of anti-inflammatory and antiviral therapies.
Itaconate has been shown to possess antiviral activity against Zika virus in neurons47 by inhibiting SDH (Fig. 6A). Since the neural cells have low capacity for recovery and adaptive immune response is highly regulated in the central nervous system, the effect of itaconate against Zika virus was found to be of high importance.95 In this study, it was observed that viral response involved the sensing of viral RNA by ZBP1 (Z-DNA-binding protein 1), which led to the activation of receptor-interacting serine/threonine kinase RIPK1 and RIPK3 regulating key inflammatory and cell death receptors.47,96 These activities induced Irg1 expression, ultimately resulting in itaconate production. Knocking out genes Zbp1 and Ripk3 demonstrated that these genes play essential roles in the expression of the virus and inhibiting them led to virus suppression. This effect was not mediated by Nrf2, as demonstrated by qPCR analysis, which revealed no significant changes in Nrf2 expression or its canonical targets, such as Hmox1, Nqo1 and Gclm in Ripk3−/− or Irg1−/− neurons. Comparative metabolomics analysis revealed that 40 distinct metabolites exhibited significant alterations in concentration, most prominently an increase in succinate and a decrease in fumarate and malate in infected cells, which are immediate downstream products of succinate oxidation. No significant changes were observed in Ripk3−/− or Irg1−/− neurons. This study showed that ZBP1 and RIPK3 induced Irg1 expression and subsequent itaconate production and TCA cycle inhibition, thus promoting antiviral metabolic state in neurons.47
To summarise the efficacy of itaconate and its derivatives against bacteria and viruses in a dose-dependent manner, data are presented in Table 1, providing information of the minimal inhibitory concentration (MIC) of itaconate, DI and 4-OI, as well as the proposed mode of action. Both, itaconate and DI were tested against enterohemorrhagic E. coli and Salmonella typhimurium,103 revealing no significance in MIC (24 and 39.52 mM, respectively). Similarly, a subtle difference in MIC was observed when itaconate and DI were tested with M. tuberculosis (1 and 0.866 mM, respectively).101,104 These data suggest that there is no significant difference between the efficacies of itaconate and its derivative DI. Notably, the antimicrobial effect of DI against M. tuberculosis, Bacillus Calmette Guérin and multidrug-resistant M. tuberculosis104 was lower when the derivative was used directly on the bacteria than that with the macrophage.
| Molecule | Bacteria | MIC (mM) | Mode of action | Reference |
|---|---|---|---|---|
| a Hypothetical mode of action.b In the article, the concentration is referred to IC50: half-maximal inhibitory concentration.c The concentration when antiviral activity was observed. | ||||
| Itaconate | Escherichia coli | 5 | Inhibition of isocitrate lyase or proton-shuttle effecta | 101 |
| Salmonella enterica ser. Typhimurium | 20 | Inhibition of isocitrate lyase or proton-shuttle effecta | ||
| Pseudomonas aeruginosa | 20 | ND | ||
| Klebsiella pneumoniae | 10 | ND | ||
| Acinetobacter baumannii | 20 | ND | ||
| Enterobacter faecium | 20 | ND | ||
| Mycobacterium tuberculosis | 1 | ND | ||
| Salmonella typhimurium | ND | ROS production | 102 | |
| Enterohemorrhagic E. coli | 24 | Inhibition of SDH | 103 | |
| Salmonella typhimurium | 24 | |||
| DI | Mycobacterium tuberculosisb | 0.866b | ND | 104 |
| Bacillus Calmette Guérinb | 1.2b | ND | ||
| Multidrug-resistant Mycobacterium tuberculosisb | 1.2b | ND | ||
| Enterohemorrhagic E. coli | 39.52 | Inhibition of SDH | 103 | |
| Salmonella typhimurium | 39.52 | |||
| 4-OI | Escherichia coli | ND | ND | 105 |
| Salmonella typhimurium | ||||
Concluding remarks and prospects
Itaconate is an important metabolite and industrially useful chemical compound. However, the interest in its anti-pathogenic and immunoregulatory properties has come to focus recently. The comparative analysis of itaconate and its derivatives shows divergent inflammasome regulation in macrophages, revealing the immunoregulatory role of itaconate and highlighting the importance of using the unmodified version of this compound in future studies on the inflammation and immune response.37 On the other hand, itaconate derivatives can also find applications in therapeutic treatments. As recently shown, 4-OI can enhance oncolytic virotherapy by suppressing antiviral immunity in cancer cells through the modification of cysteine residues in the mitochondrial antiviral signalling (MAVS)-IKKβ pathway proteins.99 The viral replication of influenza A, a positive-strand RNA (−ssRNA) virus, occurs within the nucleus of the host. Itaconate and its derivative modify the protein CRM1 in Cys528, protein nuclear export in eukaryotes, and hindering the nuclear transportation of viral ribonucleoprotein inhibiting the replication of influenza A.92,93 However, SARS-CoV-2, PRRSV and Zika are positive-strand RNA (+ssRNA) viruses that replicate in the cytoplasm. Itaconate obstruct the metabolism of infected neuron cell by inhibiting SDH, demonstrating the antiviral property against ZIKA.47 Alternatively, NF-κB plays a crucial role in the replication of SARS-CoV-2 and PRRSV.106–109 NF-κB is attenuated by itaconate and its derivative either in the anti-inflammatory pathway through inhibiting the IκBζ protein68 or by inhibiting the inflammation caused in osteoarthrosis through the STAT-dependent NF-κB pathway.110 Thus, it can be hypothesized that itaconate manifests its antiviral properties against SARS-CoV-2 and PRRSV either through inhibition of NF-κB or by obstruction of cell metabolism. Future research is needed to clarify the exact mechanism. The mechanism of inhibition of viral replication by itaconate and its derivative can differ and result in contrasting antiviral and immune responses. As itaconate impacts the immune response and, particularly, type I interferons, targeting the metabolism of this compound presents new therapeutic possibilities to improve the host defense or to tackle autoimmune disorders.88Data availability
No primary research results, software or code have been included and no new data were generated or analysed as part of this review.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work has received funding from the Research Council of Lithuania (LMTLT), agreement no. S-MIP-24-48 (to NM).References
- K. L. Wasewar, D. Shende and A. Keshav, Ind. Eng. Chem. Res., 2011, 50, 1003–1011 CrossRef CAS.
- O. B. Moore, P. A. Hanson, J. W. Comerford, A. Pellis and T. J. Farmer, Front. Chem., 2019, 7, 501 CrossRef CAS PubMed.
- S. Brännström, M. Finnveden, M. Johansson, M. Martinelle and E. Malmström, Eur. Polym. J., 2018, 103, 370–377 CrossRef.
- J. Sasikaran, M. Ziemski, P. K. Zadora, A. Fleig and I. A. Berg, Nat. Chem. Biol., 2014, 10, 371–377 CrossRef CAS PubMed.
- K. L. Tomlinson, T. W. F. Lung, F. Dach, M. K. Annavajhala, S. J. Gabryszewski, R. A. Groves, M. Drikic, N. J. Francoeur, S. H. Sridhar, M. L. Smith, S. Khanal, C. J. Britto, R. Sebra, I. Lewis, A.-C. Uhlemann, B. C. Kahl, A. S. Prince and S. A. Riquelme, Nat. Commun., 2021, 12, 1399 CrossRef CAS PubMed.
- J. Domínguez-Andrés, B. Novakovic, Y. Li, B. P. Scicluna, M. S. Gresnigt, R. J. W. Arts, M. Oosting, S. J. C. F. M. Moorlag, L. A. Groh, J. Zwaag, R. M. Koch, R. ter Horst, L. A. B. Joosten, C. Wijmenga, A. Michelucci, T. van der Poll, M. Kox, P. Pickkers, V. Kumar, H. Stunnenberg and M. G. Netea, Cell Metab., 2019, 29, 211–220 CrossRef PubMed.
- G. L. Crasso, Justus Liebigs Ann. Chem., 1840, 34, 53–84 CrossRef.
- C. T. Calam, A. E. Oxford and H. Raistrick, Biochem. J., 1939, 33, 1488–1495 CrossRef CAS PubMed.
- P. Bonnarme, B. Gillet, A. M. Sepulchre, C. Role, J. C. Beloeil and C. Ducrocq, J. Bacteriol., 1995, 177, 3573–3578 CrossRef CAS PubMed.
- K. Kinoshita, Shokubutsugaku Zasshi, 1931, 45, 45–61 CrossRef.
- T. Willke and K. D. Vorlop, Appl. Microbiol. Biotechnol., 2001, 56, 289–295 CrossRef CAS PubMed.
- J. Kim, H. M. Seo, S. K. Bhatia, H. S. Song, J. H. Kim, J. M. Jeon, K. Y. Choi, W. Kim, J. J. Yoon, Y. G. Kim and Y. H. Yang, Sci. Rep., 2017, 7, 39768 CrossRef CAS PubMed.
- R. E. Dunbar, J. Chem. Educ., 1942, 19, 398 CrossRef.
- B. E. Tate, in Kirk-Othmer Encyclopedia of Chemical Technology, 1981, DOI:10.1002/0471238961.09200103.a01.
- J.-H. Shin, J.-Y. Yang, B.-Y. Jeon, Y. Yoon, S.-H. Oh, Y.-H. Kang, D. Ryu and G.-S. Hwang, J. Proteome Res., 2011, 10, 2238–2247 CrossRef CAS PubMed.
- C. L. Strelko, W. Lu, F. J. Dufort, T. N. Seyfried, T. C. Chiles, J. D. Rabinowitz and M. F. Roberts, J. Am. Chem. Soc., 2011, 133, 16386–16389 CrossRef CAS PubMed.
- M. Sugimoto, H. Sakagami, Y. Yokote, H. Onuma, M. Kaneko, M. Mori, Y. Sakaguchi, T. Soga and M. Tomita, Metabolomics, 2012, 8, 624–633 CrossRef CAS.
- A. Michelucci, T. Cordes, J. Ghelfi, A. Pailot, N. Reiling, O. Goldmann, T. Binz, A. Wegner, A. Tallam, A. Rausell, M. Buttini, C. L. Linster, E. Medina, R. Balling and K. Hiller, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 7820–7825 CrossRef CAS PubMed.
- V. Lampropoulou, A. Sergushichev, M. Bambouskova, S. Nair, E. E. Vincent, E. Loginicheva, L. Cervantes-Barragan, X. Ma, S. C. Huang, T. Griss, C. J. Weinheimer, S. Khader, G. J. Randolph, E. J. Pearce, R. G. Jones, A. Diwan, M. S. Diamond and M. N. Artyomov, Cell Metab., 2016, 24, 158–166 CrossRef CAS PubMed.
- A. F. McGettrick and L. A. O'Neill, Curr. Opin. Immunol., 2023, 80, 102268 CrossRef CAS PubMed.
- C. G. Peace and L. A. J. O’Neill, J. Clin. Invest., 2022, 132, e148548 CrossRef CAS PubMed.
- F. J. Sánchez-García, C. A. Pérez-Hernández, M. Rodríguez-Murillo and M. M. B. Moreno-Altamirano, Front. Cell. Infect. Microbiol., 2021, 11, 725043 CrossRef PubMed.
- N. C. Williams and L. A. J. O’Neill, Front. Immunol., 2018, 9, 141 CrossRef PubMed.
- R. Singh, M. S. Smitha and S. P. Singh, J. Nanosci. Nanotechnol., 2014, 14, 4745–4756 CrossRef CAS PubMed.
- K. Shatzkes, N. D. Connell and D. E. Kadouri, Future Microbiol., 2017, 12, 469–472 CrossRef CAS PubMed.
- E. Garcia-Gutierrez, M. J. Mayer, P. D. Cotter and A. Narbad, Gut Microbes, 2019, 10, 1–21 CrossRef CAS PubMed.
- P. V. Markov, M. Ghafari, M. Beer, K. Lythgoe, P. Simmonds, N. I. Stilianakis and A. Katzourakis, Nat. Rev. Microbiol., 2023, 21, 361–379 CrossRef CAS PubMed.
- B. Hu, H. Guo, P. Zhou and Z.-L. Shi, Nat. Rev. Microbiol., 2021, 19, 141–154 CrossRef CAS PubMed.
- D. Olagnier, E. Farahani, J. Thyrsted, J. Blay-Cadanet, A. Herengt, M. Idorn, A. Hait, B. Hernaez, A. Knudsen, M. B. Iversen, M. Schilling, S. E. Jørgensen, M. Thomsen, L. S. Reinert, M. Lappe, H.-D. Hoang, V. H. Gilchrist, A. L. Hansen, R. Ottosen, C. G. Nielsen, C. Møller, D. van der Horst, S. Peri, S. Balachandran, J. Huang, M. Jakobsen, E. B. Svenningsen, T. B. Poulsen, L. Bartsch, A. L. Thielke, Y. Luo, T. Alain, J. Rehwinkel, A. Alcamí, J. Hiscott, T. H. Mogensen, S. R. Paludan and C. K. Holm, Nat. Commun., 2020, 11, 4938 CrossRef CAS PubMed.
- S. M. O'Carroll and L. A. J. O'Neill, Immunother. Adv., 2021, 1, ltab013 CrossRef PubMed.
- C. Michalaki, G. J. Albers and A. J. Byrne, Clin. Exp. Immunol., 2023, 215, 120–125 CrossRef PubMed.
- M. Chen, H. Sun, M. Boot, L. Shao, S. J. Chang, W. Wang, T. T. Lam, M. Lara-Tejero, E. H. Rego and J. E. Galán, Science, 2020, 369, 450–455 CrossRef CAS PubMed.
- M. Ruetz, G. C. Campanello, M. Purchal, H. Shen, L. McDevitt, H. Gouda, S. Wakabayashi, J. Zhu, E. J. Rubin, K. Warncke, V. K. Mootha, M. Koutmos and R. Banerjee, Science, 2019, 366, 589–593 CrossRef CAS PubMed.
- T. Robert and S. Friebel, Green Chem., 2016, 18, 2922–2934 RSC.
- J. Henderson, S. Dayalan Naidu, A. T. Dinkova-Kostova, S. Przyborski, R. Stratton and S. O′Reilly, Cells, 2021, 10, 2053 CrossRef CAS PubMed.
- E. L. Mills, D. G. Ryan, H. A. Prag, D. Dikovskaya, D. Menon, Z. Zaslona, M. P. Jedrychowski, A. S. H. Costa, M. Higgins, E. Hams, J. Szpyt, M. C. Runtsch, M. S. King, J. F. McGouran, R. Fischer, B. M. Kessler, A. F. McGettrick, M. M. Hughes, R. G. Carroll, L. M. Booty, E. V. Knatko, P. J. Meakin, M. L. J. Ashford, L. K. Modis, G. Brunori, D. C. Sévin, P. G. Fallon, S. T. Caldwell, E. R. S. Kunji, E. T. Chouchani, C. Frezza, A. T. Dinkova-Kostova, R. C. Hartley, M. P. Murphy and L. A. O'Neill, Nature, 2018, 556, 113–117 CrossRef CAS PubMed.
- A. Swain, M. Bambouskova, H. Kim, P. S. Andhey, D. Duncan, K. Auclair, V. Chubukov, D. M. Simons, T. P. Roddy, K. M. Stewart and M. N. Artyomov, Nat. Metab., 2020, 2, 594–602 CrossRef CAS PubMed.
- M. ElAzzouny, C. T. Tom, C. R. Evans, L. L. Olson, M. J. Tanga, K. A. Gallagher, B. R. Martin and C. F. Burant, J. Biol. Chem., 2017, 292, 4766–4769 CrossRef CAS PubMed.
- M. Bambouskova, L. Gorvel, V. Lampropoulou, A. Sergushichev, E. Loginicheva, K. Johnson, D. Korenfeld, M. E. Mathyer, H. Kim, L.-H. Huang, D. Duncan, H. Bregman, A. Keskin, A. Santeford, R. S. Apte, R. Sehgal, B. Johnson, G. K. Amarasinghe, M. P. Soares, T. Satoh, S. Akira, T. Hai, C. de Guzman Strong, K. Auclair, T. P. Roddy, S. A. Biller, M. Jovanovic, E. Klechevsky, K. M. Stewart, G. J. Randolph and M. N. Artyomov, Nature, 2018, 556, 501–504 CrossRef CAS PubMed.
- S. Baup, Ann. Pharm., 1836, 19, 29–38 CrossRef.
- L. Gargallo, D. Radić and A. León, Die Makromolekulare Chem., 1985, 186, 1289–1296 CrossRef CAS.
- J.-V. Richard, C. Delaite, G. Riess and A.-S. Schuller, Thermochim. Acta, 2016, 623, 136–143 CrossRef CAS.
- C. Liu, Y. Wang, J. Liu, A. N. Chen, J. Xu, R. Zhang, F. Wang, K. Nie and L. Deng, J. Org. Chem., 2021, 86, 7895–7903 CrossRef CAS PubMed.
- L. Li, S. Liu, J. Xu, S. Yu, F. Liu, C. Xie, X. Ge and J. Ren, J. Mol. Catal. A: Chem., 2013, 368–369, 24–30 CAS.
- A. K. Jha, S. C.-C. Huang, A. Sergushichev, V. Lampropoulou, Y. Ivanova, E. Loginicheva, K. Chmielewski, K. M. Stewart, J. Ashall, B. Everts, E. J. Pearce, E. M. Driggers and M. N. Artyomov, Immunity, 2015, 42, 419–430 CrossRef CAS PubMed.
- M. Bambouskova, L. Potuckova, T. Paulenda, M. Kerndl, D. A. Mogilenko, K. Lizotte, A. Swain, S. Hayes, R. D. Sheldon, H. Kim, U. Kapadnis, A. E. Ellis, C. Isaguirre, S. Burdess, A. Laha, G. K. Amarasinghe, V. Chubukov, T. P. Roddy, M. S. Diamond, R. G. Jones, D. M. Simons and M. N. Artyomov, Cell Rep., 2021, 34, 108756 CrossRef CAS PubMed.
- B. P. Daniels, S. B. Kofman, J. R. Smith, G. T. Norris, A. G. Snyder, J. P. Kolb, X. Gao, J. W. Locasale, J. Martinez, M. Gale Jr, Y. M. Loo and A. Oberst, Immunity, 2019, 50, 64–76 CrossRef CAS PubMed.
- F. Chen, W. A. M. Elgaher, M. Winterhoff, K. Büssow, F. H. Waqas, E. Graner, Y. Pires-Afonso, L. Casares Perez, L. de la Vega, N. Sahini, L. Czichon, W. Zobl, T. Zillinger, M. Shehata, S. Pleschka, H. Bähre, C. Falk, A. Michelucci, S. Schuchardt, W. Blankenfeldt, A. K. H. Hirsch and F. Pessler, Nat. Metab., 2022, 4, 534–546 CrossRef CAS PubMed.
- L. van der Straat, M. Vernooij, M. Lammers, W. van den Berg, T. Schonewille, J. Cordewener, I. van der Meer, A. Koops and L. H. de Graaff, Microb. Cell Factories, 2014, 13, 11 CrossRef PubMed.
- N. Wierckx, G. Agrimi, P. S. Lübeck, M. G. Steiger, N. P. Mira and P. J. Punt, Curr. Opin. Biotechnol., 2020, 62, 153–159 CrossRef CAS PubMed.
- E. Geiser, S. K. Przybilla, A. Friedrich, W. Buckel, N. Wierckx, L. M. Blank and M. Bölker, Microb. Biotechnol., 2016, 9, 116–126 CrossRef CAS PubMed.
- W. M. Jaklitsch, C. P. Kubicek and M. C. Scrutton, Microbiology, 1991, 137, 533–539 CrossRef CAS.
- R. D. Joerger, Genes, 2020, 11, 797 CrossRef CAS PubMed.
- H. Wang, A. A. Fedorov, E. V. Fedorov, D. M. Hunt, A. Rodgers, H. L. Douglas, A. Garza-Garcia, J. B. Bonanno, S. C. Almo and L. P. S. de Carvalho, Proc. Natl. Acad. Sci. U. S. A., 2019, 116, 15907–15913 CrossRef CAS PubMed.
- T. Zambanini, H. Hosseinpour Tehrani, E. Geiser, D. Merker, S. Schleese, J. Krabbe, J. M. Buescher, G. Meurer, N. Wierckx and L. M. Blank, Biotechnol. Biofuels, 2017, 10, 131 CrossRef PubMed.
- M. Okabe, D. Lies, S. Kanamasa and E. Y. Park, Appl. Microbiol. Biotechnol., 2009, 84, 597–606 CrossRef CAS PubMed.
- S. Krull, A. Hevekerl, A. Kuenz and U. Prüße, Appl. Microbiol. Biotechnol., 2017, 101, 4063–4072 CrossRef CAS PubMed.
- G. E. N. Nelson, D. H. Traufler, S. E. Kelley and L. B. Lockwood, Ind. Eng. Chem., 1952, 44, 1166–1168 CrossRef CAS.
- B. C. Saha and G. J. Kennedy, Lett. Appl. Microbiol., 2017, 65, 527–533 CrossRef CAS PubMed.
- I. S. Kolláth, Á. P. Molnár, Á. Soós, E. Fekete, E. Sándor, B. Kovács, C. P. Kubicek and L. Karaffa, Front. Microbiol., 2019, 10, 1589 CrossRef PubMed.
- B. C. Saha, J. Ind. Microbiol. Biotechnol., 2017, 44, 303–315 CrossRef CAS PubMed.
- K. S. Vuoristo, A. E. Mars, J. V. Sangra, J. Springer, G. Eggink, J. P. Sanders and R. A. Weusthuis, Appl. Microbiol. Biotechnol., 2015, 99, 221–228 CrossRef CAS PubMed.
- M. Merkel, D. Kiefer, M. Schmollack, B. Blombach, L. Lilge, M. Henkel and R. Hausmann, Bioresour. Technol., 2022, 351, 126994 CrossRef CAS PubMed.
- A. Otten, M. Brocker and M. Bott, Metab. Eng., 2015, 30, 156–165 CrossRef CAS PubMed.
- Y. Zhang, Y.-J. Zhou, J.-S. Tang, J.-Q. Lan, Y.-Y. Kang, L. Wu and Y. Peng, J. Asian Nat. Prod. Res., 2022, 24, 577–588 CrossRef CAS PubMed.
- S. T. Liao, C. Han, D. Q. Xu, X. W. Fu, J. S. Wang and L. Y. Kong, Nat. Commun., 2019, 10, 5091 CrossRef PubMed.
- T. Cordes, M. Wallace, A. Michelucci, A. S. Divakaruni, S. C. Sapcariu, C. Sousa, H. Koseki, P. Cabrales, A. N. Murphy, K. Hiller and C. M. Metallo, J. Biol. Chem., 2016, 291, 14274–14284 CrossRef CAS PubMed.
- Y. Feng, Z. Chen, Y. Xu, Y. Han, X. Jia, Z. Wang, N. Zhang and W. Lv, Front. Immunol., 2023, 14, 1188253 CrossRef CAS PubMed.
- N. Donnelly, A. M. Gorman, S. Gupta and A. Samali, Cell. Mol. Life Sci., 2013, 70, 3493–3511 CrossRef CAS PubMed.
- A. Hooftman, S. Angiari, S. Hester, S. E. Corcoran, M. C. Runtsch, C. Ling, M. C. Ruzek, P. F. Slivka, A. F. McGettrick, K. Banahan, M. M. Hughes, A. D. Irvine, R. Fischer and L. A. J. O'Neill, Cell Metab., 2020, 32, 468–478 CrossRef CAS PubMed.
- K. V. Swanson, M. Deng and J. P. Ting, Nat. Rev. Immunol., 2019, 19, 477–489 CrossRef CAS PubMed.
- W. Qin, K. Qin, Y. Zhang, W. Jia, Y. Chen, B. Cheng, L. Peng, N. Chen, Y. Liu, W. Zhou, Y.-L. Wang, X. Chen and C. Wang, Nat. Chem. Biol., 2019, 15, 983–991 CrossRef CAS PubMed.
- W. W. Ackermann and V. R. Potter, Proc. Soc. Exp. Biol. Med., 1949, 72, 1–9 CrossRef CAS PubMed.
- C. Yang, T. Liu and G. P. Shi, EBioMedicine, 2020, 59, 102938 CrossRef CAS PubMed.
- C. Zhang, G. Shang, X. Gui, X. Zhang, X. C. Bai and Z. J. Chen, Nature, 2019, 567, 394–398 CrossRef CAS PubMed.
- X. Hu, H. Zhang, Q. Zhang, X. Yao, W. Ni and K. Zhou, J. Neuroinflammation, 2022, 19, 242 CrossRef CAS PubMed.
- W. Yang, Y. Wang, K. Tao and R. Li, Cell. Mol. Biol. Lett., 2023, 28, 100 CrossRef CAS PubMed.
- W. Li, Y. Li, J. Kang, H. Jiang, W. Gong, L. Chen, C. Wu, M. Liu, X. Wu, Y. Zhao and J. Ren, Cell Rep., 2023, 42, 112145 CrossRef CAS PubMed.
- A. V. Ferreira, S. Kostidis, L. A. Groh, V. A. C. M. Koeken, M. Bruno, I. Baydemir, G. Kilic, Ö. Bulut, T. Andriopoulou, V. Spanou, K. D. Synodinou, T. Gkavogianni, S. J. C. F. M. Moorlag, L. Charlotte de Bree, V. P. Mourits, V. Matzaraki, W. J. H. Koopman, F. L. van de Veerdonk, G. Renieris, M. Giera, E. J. Giamarellos-Bourboulis, B. Novakovic and J. Domínguez-Andrés, Cell Rep., 2023, 42, 112658 CrossRef CAS PubMed.
- H. H. Luan and R. Medzhitov, Cell Metab., 2016, 24, 379–387 CrossRef CAS PubMed.
- B. A. McFadden and S. Purohit, J. Bacteriol., 1977, 131, 136–144 CrossRef CAS PubMed.
- I. A. Berg, L. V. Filatova and R. N. Ivanovsky, FEMS Microbiol. Lett., 2002, 216, 49–54 CrossRef CAS PubMed.
- E.-M. Schuster, M. W. Epple, K. M. Glaser, M. Mihlan, K. Lucht, J. A. Zimmermann, A. Bremser, A. Polyzou, N. Obier, N. Cabezas-Wallscheid, E. Trompouki, A. Ballabio, J. Vogel, J. M. Buescher, A. J. Westermann and A. S. Rambold, Nat. Metab., 2022, 4, 856–866 CrossRef CAS PubMed.
- B. X. C. Kwai, A. J. Collins, M. J. Middleditch, J. Sperry, G. Bashiri and I. K. H. Leung, RSC Med. Chem., 2021, 12, 57–61 RSC.
- A. Gidon, C. Louet, L. M. Røst, P. Bruheim and T. H. Flo, mBio, 2021, 12, e0212121 CrossRef PubMed.
- Z. Zhang, C. Chen, F. Yang, Y. X. Zeng, P. Sun, P. Liu and X. Li, Mol. Cell, 2022, 82, 2844–2857 CrossRef CAS PubMed.
- Z. Zasłona and L. A. J. O'Neill, Mol. Cell, 2020, 78, 814–823 CrossRef PubMed.
- S. M. O'Carroll, F. D. R. Henkel and L. A. J. O'Neill, Immunol. Rev., 2024, 323, 276–287 CrossRef PubMed.
- Z. Li, W. Zheng, W. Kong and T. Zeng, Inflammation, 2023, 46, 1177–1191 CrossRef CAS PubMed.
- M. C. Runtsch, S. Angiari, A. Hooftman, R. Wadhwa, Y. Zhang, Y. Zheng, J. S. Spina, M. C. Ruzek, M. A. Argiriadi, A. F. McGettrick, R. S. Mendez, A. Zotta, C. G. Peace, A. Walsh, R. Chirillo, E. Hams, P. G. Fallon, R. Jayaraman, K. Dua, A. C. Brown, R. Y. Kim, J. C. Horvat, P. M. Hansbro, C. Wang and L. A. J. O'Neill, Cell Metab., 2022, 34, 487–501 CrossRef CAS PubMed.
- K. T. Nguyen, M. P. Holloway and R. A. Altura, Int. J. Biochem. Mol. Biol., 2012, 3, 137–151 CAS.
- P. Ribó-Molina, H. J. Weiss, B. Susma, S. van Nieuwkoop, L. Persoons, Y. Zheng, M. Ruzek, D. Daelemans, R. A. M. Fouchier, L. A. J. O'Neill and B. G. van den Hoogen, J. Virol., 2023, 97, e0132523 CrossRef PubMed.
- F. H. Waqas, M. Shehata, W. A. M. Elgaher, A. Lacour, N. Kurmasheva, F. Begnini, A. E. Kiib, J. Dahlmann, C. Chen, A. Pavlou, T. B. Poulsen, S. Merkert, U. Martin, R. Olmer, D. Olagnier, A. K. H. Hirsch, S. Pleschka and F. Pessler, PLoS Pathog., 2023, 19, e1011506 CrossRef CAS PubMed.
- A. Sohail, A. A. Iqbal, N. Sahini, F. Chen, M. Tantawy, S. F. H. Waqas, M. Winterhoff, T. Ebensen, K. Schultz, R. Geffers, K. Schughart, M. Preusse, M. Shehata, H. Bähre, M. C. Pils, C. A. Guzman, A. Mostafa, S. Pleschka, C. Falk, A. Michelucci and F. Pessler, PLoS Pathog., 2022, 18, e1010219 CrossRef CAS PubMed.
- R. S. Klein and C. A. Hunter, Immunity, 2017, 46, 891–909 CrossRef CAS PubMed.
- L. Mifflin, D. Ofengeim and J. Yuan, Nat. Rev. Drug Discovery, 2020, 19, 553–571 CrossRef CAS PubMed.
- A. Decout, J. D. Katz, S. Venkatraman and A. Ablasser, Nat. Rev. Immunol., 2021, 21, 548–569 CrossRef CAS PubMed.
- C. Su, T. Cheng, J. Huang, T. Zhang and H. Yin, Cell Rep., 2023, 42, 113040 CrossRef CAS PubMed.
- N. Kurmasheva, A. Said, B. Wong, P. Kinderman, X. Han, A. H. F. Rahimic, A. Kress, M. E. Carter-Timofte, E. Holm, D. van der Horst, C. F. Kollmann, Z. Liu, C. Wang, H. D. Hoang, E. Kovalenko, M. Chrysopoulou, K. S. Twayana, R. N. Ottosen, E. B. Svenningsen, F. Begnini, A. E. Kiib, F. E. H. Kromm, H. J. Weiss, D. Di Carlo, M. Muscolini, M. Higgins, M. van der Heijden, A. Bardoul, T. Tong, A. Ozsvar, W. H. Hou, V. R. Schack, C. K. Holm, Y. Zheng, M. Ruzek, J. Kalucka, L. de la Vega, W. A. M. Elgaher, A. R. Korshoej, R. Lin, J. Hiscott, T. B. Poulsen, L. A. O'Neill, D. G. Roy, M. M. Rinschen, N. van Montfoort, J. S. Diallo, H. F. Farin, T. Alain and D. Olagnier, Nat. Commun., 2024, 15, 4096 CrossRef CAS PubMed.
- Y. Pang, Y. Wang, C. Li, J. Liu, C. Duan, Y. Zhou, L. Fang and S. Xiao, Virology, 2022, 577, 84–90 CrossRef CAS PubMed.
- D. Duncan, A. Lupien, M. A. Behr and K. Auclair, Microbiology, 2021, 167, 001050 CrossRef CAS PubMed.
- X. Zhu, Y. Guo, Z. Liu, J. Yang, H. Tang and Y. Wang, Sci. Rep., 2021, 11, 18173 CrossRef CAS PubMed.
- L. Y. Xie, Y. B. Xu, X. Q. Ding, S. Liang, D. L. Li, A. K. Fu and X. A. Zhan, Biomed. Pharmacother., 2023, 167, 115487 CrossRef CAS PubMed.
- Y. J. Kim, E.-J. Park, S.-H. Lee, P. Silwal, J. K. Kim, J. S. Yang, J. Whang, J. Jang, J.-M. Kim and E.-K. Jo, Cell Biosci., 2023, 13, 49 CrossRef CAS PubMed.
- Q. Ding, X. Jing, S. Yao, W. Su, B. Ye, Y. Qu, F. Gao, T. Sun and X. Guo, Biomater. Adv., 2022, 139, 212979 CrossRef CAS PubMed.
- C. R. Simoneau, P.-Y. Chen, G. K. Xing, J. M. Hayashi, I. P. Chen, M. M. Khalid, N. L. Meyers, T. Y. Taha, K. E. Leon, R. K. Suryawanshi, M. McCavitt-Malvido, T. Ashuach, K. A. Fontaine, L. Rodriguez, B. Joehnk, K. Walcott, S. Vasudevan, X. Fang, M. Maishan, S. Schultz, J. P. Roose, M. A. Matthay, A. Sil, M. Arjomandi, N. Yosef and M. Ott, Sci. Rep., 2024, 14, 15351 CrossRef CAS PubMed.
- Q. Zhou, L. Zhang, Y. Dong, Y. Wang, B. Zhang, S. Zhou, Q. Huang, T. Wu and G. Chen, Hypertens. Res., 2024, 47, 375–384 CrossRef PubMed.
- B. E. Nilsson-Payant, S. Uhl, A. Grimont, A. S. Doane, P. Cohen, R. S. Patel, C. A. Higgins, J. A. Acklin, Y. Bram, V. Chandar, D. Blanco-Melo, M. Panis, J. K. Lim, O. Elemento, R. E. Schwartz, B. R. Rosenberg, R. Chandwani and B. R. tenOever, J. Virol., 2021, 91, e01257 Search PubMed.
- Y. Xu, Z. Zhu, M. Zhang, L. Chen, K. Tian and X. Li, Microbiol. Spectrum, 2024, 12, e0347923 CrossRef PubMed.
- L. Ni, Z. Lin, S. Hu, Y. Shi, Z. Jiang, J. Zhao, Y. Zhou, Y. Wu, N. Tian, L. Sun, A. Wu, Z. Pan, X. Zhang and X. Wang, Biochem. Pharmacol., 2022, 198, 114935 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2025 |