
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceSynthetic biology strategies for cyanobacterial systems to heterologously produce cyanobacterial natural products
Manyun
Chen†
 a,
Dipesh
Dhakal†
a,
Campbell W.
Eckhardt
a,
Hendrik
Luesch
*ab and
Yousong
Ding
*a
a,
Dipesh
Dhakal†
a,
Campbell W.
Eckhardt
a,
Hendrik
Luesch
*ab and
Yousong
Ding
*a
aDepartment of Medicinal Chemistry, Center for Natural Products, Drug Discovery and Development, University of Florida, Gainesville, FL 32610, USA. E-mail: luesch@cop.ufl.edu; yding@cop.ufl.edu
bProgram in Cancer and Stem Cell Biology, Duke-NUS Medical School, Singapore, 169857, Singapore
First published on 16th April 2025
Abstract
Covering: 2014 to 2024
Cyanobacteria are prolific producers of bioactive natural products, including promising drug leads for FDA-approved cancer therapeutics. Advances in genome sequencing and computational tools have revealed a wealth of cyanobacterial biosynthetic gene clusters (BGCs). However, progress in genome-driven discovery has been hindered by challenges in manipulating native hosts and the limited availability of efficient heterologous expression platforms. This highlight focuses on recent synthetic biology innovations on cyanobacterial systems that address these obstacles, facilitating the production of diverse cyanobacterial natural product families. We discuss key features of widely used cyanobacterial chassis, such as Synechocystis sp. PCC 6803, Synechococcus elongatus UTEX 2973, Anabaena sp. PCC 7120, and emerging hosts. Advances in BGC cloning, combinatorial biosynthesis, transcriptional and translational regulation, and host engineering are also highlighted. Together, these synthetic biology developments provide a powerful framework for expanding cyanobacterial natural product discovery and production.
 Manyun Chen | Manyun Chen, PhD, earned her B.S. from Tsinghua University and M.S. in Pharmaceutical Sciences from the University of Southern California. She completed her PhD in Medicinal Chemistry at the University of Florida in 2022, studying genomics-guided discovery and biosynthetic pathways of natural products under Prof. Yousong Ding. She then served as a Postdoctoral Associate, focusing on enzyme mechanisms and biocatalyst development. In 2023, she was appointed Assistant Scientist at UF's Center for Natural Products, Drug Discovery and Development (CNPD3), where her research centers on genome mining and biosynthetic production of marine-derived natural products. |
 Dipesh Dhakal | Dipesh Dhakal, PhD, is a Postdoctoral Associate in the Department of Medicinal Chemistry and the Center for Natural Products, Drug Discovery and Development at the University of Florida. He earned his B.Tech in Biotechnology from Kathmandu University (Nepal) in 2007 and M.S. from Guru Nanak Dev University (India) in 2010. He completed his PhD at Sun Moon University (Korea) in 2016, focusing on the biosynthetic and regulatory pathways of natural products in actinobacteria. Since 2020, Dr Dhakal has been investigating the biosynthesis of cyanobacterial natural products, including lyngbyastatins, apratoxins, and dolastatin-10, with an emphasis on their enzymatic mechanisms. |
 Campbell W. Eckhardt | Campbell Eckhardt earned his B.S. in Biochemistry from the University of Tulsa in 2024, where he studied metabolite-driven modulation of salamander morphology under Prof. Ron Bonett. He is currently pursuing a PhD in Pharmaceutical Sciences at the University of Florida in Prof. Yousong Ding's lab. His research focuses on uncovering the molecular mechanisms by which proteins mediate adaptive responses to environmental stressors, ultimately shaping host survival and evolutionary trajectories across diverse biological systems. |
 Hendrik Luesch | Hendrik Luesch, PhD, is a Professor of Medicinal Chemistry at the University of Florida. He earned his Diplom in Chemistry from the University of Siegen (Germany) in 1997 and a PhD in Marine Natural Products Chemistry from the University of Hawaii in 2002 under Prof. Richard E. Moore. He completed postdoctoral training as an Irving S. Sigal Fellow at The Scripps Research Institute with Prof. Peter G. Schultz. Since joining UF in 2005, Dr Luesch has led a multidisciplinary program integrating chemistry, genomics, synthetic biology, and early drug development, with a focus on cyanobacterial natural products for over 25 years. |
 Yousong Ding | Yousong Ding, PhD, is an Associate Professor of Medicinal Chemistry at the University of Florida. He received his B.S. from Peking University, M.S. from the University of Nebraska–Lincoln (with Prof. Liangcheng Du), and PhD from the University of Michigan (with Prof. David H. Sherman). His research centered on the biosynthesis, enzymology, and chemistry of natural products from cyanobacteria, Streptomyces, and fungi. After postdoctoral work with Prof. Frances Arnold at Caltech and a stint at Pfizer, he joined UF in 2013. His lab integrates multidisciplinary approaches to discover and develop nature-inspired bioactive molecules addressing pressing societal needs. |
1. Introduction
Cyanobacteria are a diverse group of Gram-negative, photoautotrophic prokaryotes that have thrived in almost all environments, particularly illuminated niches, for over 3.5 billion years, from aquatic ecosystems to extreme terrestrial habitats such as deserts. Their remarkable ecological adaptability has driven the evolution of diverse survival strategies, including intricate symbiotic relationships and metabolic specialization.1 A key outcome of this evolutionary history is the biosynthesis of an extensive repertoire of structurally and functionally diverse natural products (NPs),2 which play crucial roles in interspecies competition and environmental resilience. These bioactive compounds, ranging from anti-UV agents such as mycosporine-like amino acids to antimicrobials like hapalindoles and antivirals such as cyanovirin-N, provide cyanobacteria with a competitive advantage in their natural habitats.3 Notably, certain cyanobacterial NPs, including dolastatin 10, gatorbulin-1, and cryptophycins, exhibit potent cytotoxic activity by modulation of tubulin dynamics, a validated anticancer strategy, making them valuable drug leads.4 Among these leads, six synthetic analogs of dolastatin 10 have received FDA approval, with five currently on the market, including brentuximab vedotin, polatuzumab vedotin, enfortumab vedotin, disitamab vedotin, and tisotumab vedotin. This remarkable chemical diversity allows cyanobacterial NPs to interact with a wide range of biological targets, contributing to their broad pharmacological potential.3–5Over the past decades, the discovery of bioactive cyanobacterial NPs has extended beyond traditional isolation-based approaches to include in silico strategies, unveiling a vast, untapped chemical diversity encoded within biosynthetic gene clusters (BGCs).6 BGCs are co-localized genes responsible for synthesizing diverse NPs, such as nonribosomal peptides (NRPs), polyketides (PKs), ribosomally synthesized and post-translationally modified peptides (RiPPs), terpenes, glycosides, phosphonates, and biosynthetic hybrids. However, despite notable progress in associating BGCs with known NP families, more than 80% of cyanobacterial BGCs remain uncharacterized or orphaned.5 A major challenge in assigning NPs to their corresponding BGCs stems from the widespread presence of silent or cryptic clusters, which are not expressed under standard laboratory conditions or when native producers are harvested from the field. On the other hand, recent advances in genome sequencing and bioinformatics have greatly accelerated efforts to decipher cyanobacterial biosynthetic potential. The National Center for Biotechnology Information (NCBI) GenBank serves as the most comprehensive repository of cyanobacterial genomes, classifying assemblies into four levels: complete genome, chromosome, scaffold, and contig. As of December 2024, GenBank contained 360 complete genomes, 83 chromosomes, 1672 scaffolds, and 4958 contigs, totalling 6516 cyanobacterial sequences, an approximate 40-fold increase from 178 sequences available between 1980 and 2013.7 Additionally, at the 14th Workshop on Cyanobacteria held in June 2022 at Michigan State University, the newly curated cyanobacterial genomic database CyanoCyc was introduced and later integrated into the BioCyc collection. CyanoCyc encompasses 277 cyanobacterial genomes across 56 genera, with five genomes manually annotated using information from over 1765 publications, providing detailed insights into gene functions and metabolic pathways.8 Other notable genome repositories facilitating cyanobacterial NP discovery include the Joint Genome Institute (JGI) Genome Portal and UniProt Knowledgebase.
Another major advance in in silico cyanobacterial NPs discovery is the increasing availability of bioinformatics tools for mining BGCs and predicting their associated NPs. Antibiotics & Secondary Metabolite Analysis Shell (antiSMASH) 7.0 enables rapid BGC detection, annotation, and comparative analysis using profile hidden Markov models.9 Since its initial release in 2011, antiSMASH has integrated many other tools to enhance BGC mining capabilities. Another key resource, Minimum Information about a Biosynthetic Gene Cluster (MIBiG), serves as a reference database, linking BGCs to known NPs and providing insights into sequence–structure–function relationships.10 These datasets have also been incorporated into more advanced computational frameworks, such as the Biosynthetic Gene Similarity Clustering and Prospecting Engine (BiG-SCAPE) and CORe Analysis of Syntenic Orthologues (CORASON) to prioritize NP BGCs.11 BiG-SCAPE classifies BGCs through sequence similarity networks, while CORASON maps their evolutionary relationships across species. Furthermore, the recently released secondary metabolism collaboratory holds 13.1 million BGC regions from bacteria and archaea, including over 56![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 cyanobacterial BGCs.12 Despite these advancements, bottlenecks remain functionally characterizing BGCs and activating silent clusters to produce encoded NPs. In this regard, heterologous expression has emerged as a powerful strategy to overcome these limitations, unlocking the full biosynthetic potential of cyanobacteria.
000 cyanobacterial BGCs.12 Despite these advancements, bottlenecks remain functionally characterizing BGCs and activating silent clusters to produce encoded NPs. In this regard, heterologous expression has emerged as a powerful strategy to overcome these limitations, unlocking the full biosynthetic potential of cyanobacteria.
Early efforts in the heterologous expression of cyanobacterial NPs have primarily focused on RiPPs in Escherichia coli, largely due to their small BGC sizes. As one of the most extensively developed microbial chassis, E. coli offers several advantages, including rapid growth, a wealth of genetic tools, and well-established protocols for cultivation and genetic manipulation. Recent advancements in large BGC assembly and engineered E. coli strains have enabled the production of more complex cyanobacterial NPs. However, many attempts to express cyanobacterial BGCs in E. coli or other conventional heterologous hosts have been unsuccessful, underscoring the need for alternative expression systems, particularly cyanobacteria.13 Transcriptional regulation in cyanobacteria differs significantly from that in E. coli, including distinct promoter architectures, RNA polymerase compositions, and global regulatory networks, as reviewed previously.14 Moreover, cyanobacteria's photoautotrophic nature makes them promising biofactories for sustainable, high-value NP production in an environmentally friendly manner. However, despite several successful examples (Table 1 and Fig. 1), their potential as heterologous hosts remain largely untapped. In this research highlight, we focus on recent advances in chassis development and synthetic biology tools for heterologous expression in cyanobacteria. Successfully expressed cyanobacterial NPs are discussed in the following two sections to illustrate key strategies employed. These developments could bridge the gap between the numerous BGCs identified through genome mining and the limited number of NPs discovered.
| NP name | NP class | BGC size (kb) | BGC origin | Heterologous host | Assembly method | Major modifications | Maximum yield |
|---|---|---|---|---|---|---|---|
| a (1) NRP: non-ribosomal peptide; (2) TAR: transformation-associated recombination; (3) The use of codon optimized genes; (4) PK: polyketide; (5) DCW: dry biomass weight; (6) ND: not determined. | |||||||
| Lyngbyatoxin A (1) | NRP (1) | 11.3 | Moorena producens | Anabaena sp. PCC 7120 | TAR (2) | Native | 2307 ng mg−1 DCW (5) |
| Pendolmycin (2) | NRP | 12 | M. producens (3) | Anabaena sp. PCC 7120 | TAR | Combinatorial biosynthesis, promoter change | 180 ng mg−1 DCW |
| Teleocidin B-4 (3) | NRP | 12 | M. producens (3) | Anabaena sp. PCC 7120 | TAR | Combinatorial biosynthesis, promoter change | 1035 ng mg−1 DCW |
| APK (PK fragment of apratoxin A, 4) | PK (4) | 25 | M. bouillonii (3) | Anabaena sp. PCC 7120 | Gibson assembly | Promoter change | 9.7 mg L−1 |
| Tolypodiol (5) | Mero-terpenoid | 55 | Brasilonema sp. HT-58-2 | Anabaena sp. PCC 7120 | TAR | Codon optimization, promoter change | ND (6) |
| Cryptomaldamide (6) | PK-NRP | 28.7 | M. producens JHB | Anabaena sp. PCC 7120 | TAR | Native | 15.3 mg g−1 DCW |
| Columbamide (7) | PK-NRP | 28.5 | M. bouillonii PNG5-198 | Anabaena sp. PCC 7120 | TAR | Native | ND |
| Shinorine (8) | NRP | 6.4 | Fischerella sp. PCC 9339 | Synechocystis sp. PCC 6803 | Restriction-ligation | Native and refactored | 2.4 mg g−1 DCW |
| Hapalindoles (9) | Alkaloid | 42 | F. ambigua UTEX 1903 | Synechococcus elongatus UTEX 2973 | Gibson assembly | Refactored | 2.0 mg g−1 DCW |
 | ||
| Fig. 1 Structures of cyanobacterial natural products successfully produced in heterologous cyanobacterial hosts. | ||
2. Cyanobacterial chassis for heterologous production
An ideal heterologous host for NP expression should possess key attributes such as a fast growth rate, biosynthetic compatibility with specific NP classes, and genetic tractability for efficient manipulation. Model bacteria with well-established protocols for conjugation and gene editing are often prioritized. Additionally, hosts with minimal secondary metabolism are advantageous for maximizing the biosynthetic efficiency of exogenous BGCs, whereas strains harboring specific BGC types may enhance expression, facilitate proper protein folding, and provide essential metabolic precursors. Here, we provide a brief overview of the genomic features of selected cyanobacterial strains that have been utilized, or hold potential, as heterologous hosts to produce cyanobacterial NPs (Table 2) and then highlight their applications.| Strain | Genome size (Mb) | GC content (%) | The number of predicted BGCs | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Terpene | RiPP | NRPS | PKS | NRPS/PKS hybrid | Others | Total | |||
| Anabaena 7120 | 7.2 | 41.5 | 4 | 3 | 1 | 1 | 3 | 1 | 13 |
| Synechocystis 6803 | 3.9 | 47.5 | 3 | 3 | |||||
| Synechococcus 7942 | 2.8 | 55.5 | 2 | 2 | |||||
| Synechococcus 2973 | 2.7 | 55.5 | 2 | 2 | |||||
| Synechococcus 7002 | 3.4 | 49.0 | 1 | 1 | 1 | 3 | |||
| Synechococcus 11901 | 3.5 | 49.0 | 1 | 1 | 1 | 3 | |||
2.1 Anabaena sp. PCC 7120
The filamentous freshwater cyanobacterium Anabaena sp. PCC 7120 (hereafter Anabaena 7120) has emerged as the most effective cyanobacterial chassis for heterologous production of cyanobacterial NPs (Table 1). As a well-established model organism with a doubling time of approximately 14–15 hours, Anabaena 7120 has been extensively utilized to elucidate key cyanobacterial biological processes, such as nitrogen fixation and cell differentiation. Although Anabaena 7120 cannot grow under strictly heterotrophic conditions in darkness, it has demonstrated the ability to support mixotrophic growth when supplemented with fructose,15 offering a potential strategy for high cell-density cultivation. This approach could improve NP yields by optimizing carbon flux and enhancing cellular metabolism.16The genome of Anabaena 7120 encodes 13 NP BGCs including five NRPS and PKS BGCs (Table 2), underscoring its inherent biosynthetic capacity. Notably, it harbors a promiscuous Sfp-type phosphopantetheinyl transferase (PPTase), capable of activating carrier proteins of fatty acid synthases (FASs), PKSs, and NRPSs.17 The Philmus group first demonstrated the utility of Anabaena 7120 as a heterologous host for NP production by expressing lyngbyatoxin A (1) from a three-gene BGC derived from the marine cyanobacterium Moorena producens (Fig. 1 and 2A).18 This pathway includes the dimodular NRPS LtxA, which synthesizes the N-methyl-valyl-tryptophan thioester, the cytochrome P450 LtxB, which catalyzes indolactam-forming cyclization, and the prenyltransferase LtxC, which installs the geranyl moiety. Introduction of the ltxA-C genes enabled Anabaena 7120 to produce lyngbyatoxin A (1) in both solid and liquid cultures. Optimization studies revealed that nitrogen source selection and promoter engineering significantly improved titers (Fig. 2A), reaching up to 2307 ng mg−1 dry cell weight (DCW). Furthermore, this chassis has facilitated the heterologous production of NRP/PK hybrid compounds such as cryptomaldamide (6) and columbamide (7) (Fig. 1),19,20 and currently remains the only cyanobacterial host capable of heterologous expression of complex NRP/PK-type cyanobacterial NPs (Table 1). More recently, Anabaena 7120 has been used to produce the cyanobacterial meroterpenoid tolypodiol (5) and its analogs by expressing a 21-kb BGC,21 further demonstrating its versatility in supporting the heterologous production of diverse NP classes. Beyond native pathway expression, it has been successfully applied in combinatorial biosynthesis approaches. For example, swapping the prenyltransferase gene with mpnD from the pendolmycin BGC in marine Actinobacteria or fusing the methyltransferase gene tleD from the teleocidin BGC in Streptomyces yielded diverse indolelactam derivatives (Table 1 and Fig. 2A),16 highlighting cross-phylum BGC engineering. Similarly, we recently modified the PKS subcluster of apratoxin A by incorporating the type-II thioesterase from the erythromycin BGC to facilitate the release of the final product APK (4) in Anabaena 7120 (Fig. 2B).22
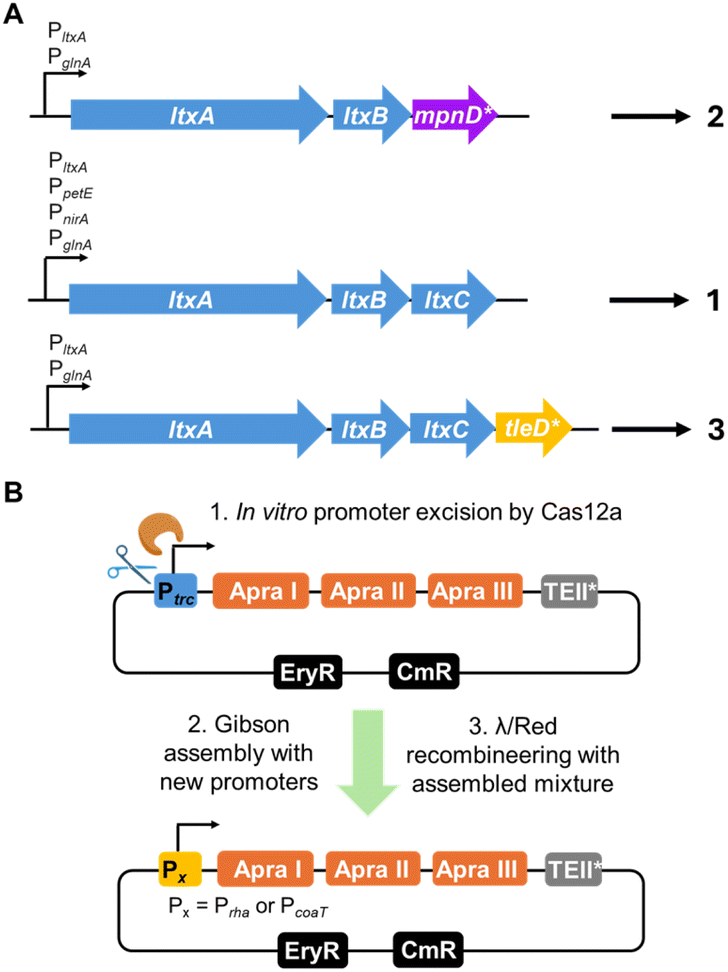 | ||
| Fig. 2 Representative recent advances in engineering cyanobacterial BGCs for heterologous expression in Anabaena 7120. LtxA: NRPS; LtxB: P450; LtxC: prenyltransferase; MpnD*: prenyltransferase; TleD*: methyltransferase-cyclase; *: codon optimized gene. (A) Combinatorial biosynthesis and promoter optimization for the heterologous production of indolelactams (1–3). (B) Promoter engineering for the heterologous expression of APK (4). The APK BGC was divided into three fragments (Apra I–III) and the codon optimized type-II thioesterase (TEII*) domain of the erythromycin cluster was employed to release APK (4). The strong constitutive promoter Ptrc in the cloned BGC was replaced with inducible promoters Prha or PcoaT through a combination of in vitro Cas12a-directed cleavage followed by Gibson assembly and in vivo λRed/ET recombineering. EryR and CmR: erythromycin and chloramphenicol resistance markers, respectively. | ||
2.2 Unicellular cyanobacteria
Compared to Anabaena 7120, the unicellular freshwater model cyanobacteria Synechocystis sp. PCC 6803 (hereafter Synechocystis 6803) and Synechococcus elongatus PCC 7942 (hereafter Synechococcus 7942) exhibit significantly faster growth rates, with doubling times of approximately 7–12 hours and 6–7 hours, respectively, which can be further optimized under controlled conditions.23 Notably, S. elongatus UTEX 2973 (hereafter Synechococcus 2973), a close relative of Synechococcus 7942, demonstrates exceptionally rapid autotrophic growth with a doubling time of less than 2 hours, comparable to many heterotrophic industrial hosts.24 These cyanobacterial strains have been extensively engineered as biofactories for economically valuable compounds, including biofuels, sugars, hydrocarbons, and alcohols. Due to their relatively small genome sizes and lack of native PKS and NRPS pathways (Table 2), heterologous expression of NPs in these chassis has primarily focused on terpenoids.25 One notable exception is the heterologous expression of hapalindole alkaloids (9) in Synechococcus 2973 (Table 1 and Fig. 1), achieved by introducing a 42-kb hapalindole BGC on a replicative plasmid.26 Additionally, despite the absence of a native NRPS BGC, Synechocystis 6803 successfully expressed the shinorine (8) gene cluster from Fischerella sp. PCC 9339 (Table 1 and Fig. 1),27 which includes a single-modular NRPS-like gene, through co-expression with the PPTase of Anabaena 7120. These studies highlight the potential of synthetic biology approaches to enhance the suitability of fast-growing cyanobacterial chassis for the sustainable biosynthesis of diverse cyanobacterial NPs (Table 1 and Fig. 1).Since many bioactive cyanobacterial NPs originate from marine environments, marine cyanobacteria could serve as promising alternative hosts for heterologous expression of these compounds. These organisms often possess specialized metabolic pathways and unique transcriptional regulatory systems that may facilitate the biosynthesis of complex NP precursors and tailoring enzymes not readily available in traditional hosts.28 Moreover, marine cyanobacteria are naturally adapted to various environmental stressors, including high light intensity, elevated temperatures, and fluctuating salinity, enabling them to thrive under dynamic conditions. The model marine cyanobacterium Synechococcus sp. PCC 7002 (hereafter Synechococcus 7002) has been successfully engineered for high-yield sucrose production under high salt concentration.29 Furthermore, Synechococcus sp. PCC 11901 (hereafter Synechococcus 11901), a fast-growing marine cyanobacterium, has been equipped with comprehensive genetic toolkits, including genomic integration systems, constitutive and inducible promoters, and CRISPR-based genome editing.30 Notably, both Synechococcus 7002 and 11901 harbor native NRPS/PKS-type BGCs (Table 2), further supporting their potential as viable hosts for the heterologous biosynthesis of complex NPs.
2.3 New opportunities
One major challenge in developing cyanobacteria as heterologous hosts is polyploidy, a common trait among cyanobacteria, with most strains carrying between 3 to 200 genome copies per cell.31 While polyploidy has been linked to beneficial traits such as enhanced stress tolerance and increased metabolic capacity, it complicates genetic engineering by requiring multiple selection steps to achieve a homozygous strain and a genetically homogenous population. However, certain cyanobacterial species, including Synechocystis 6803, Synechococcus 7942 and 2973, are naturally competent, allowing them to uptake exogenous DNA and integrate it into their genome via homologous recombination.32 Although neutral chromosomal sites for heterologous gene integration have been identified to minimize pleiotropic effects, polyploidy remains a key limitation of this approach. To overcome these challenges, advances in alternative genetic editing methods have been developed to facilitate rational strain engineering in polyploid cyanobacteria. For industrial-scale applications, continued optimization of cyanobacterial chassis is essential. Strategies such as engineering precursor and cofactor pathways, modulating regulatory networks, and fine-tuning biosynthetic pathways can significantly enhance metabolite production efficiency.33 Continued research in this area, particularly the development of new chassis, will be critical to fully harnessing cyanobacteria as sustainable platforms for NP discovery and biomanufacturing.3. Synthetic biology strategies for heterologous production of cyanobacterial NPs in cyanobacterial hosts
Heterologous expression can potentially circumvent the context dependency of BGC activation, which is often influenced by native regulatory networks and signaling molecules. Additionally, it enables the discovery of valuable new NPs and the generation of novel derivatives through pathway and strain engineering. The heterologous expression process involves several key steps, including the identification of target BGC, assembly of the BGC, and delivery of the BGC into a suitable host.13 After selecting a chassis with a well-characterized genome and favorable metabolic traits, the “design-build-test-learn” cycle of synthetic biology can serve as an engineering framework to facilitate successful heterologous expression. This iterative process integrates (1) standardized synthetic biology parts tailored for host compatibility, (2) efficient cloning and delivery of genetic materials, and (3) metabolic engineering and genome editing strategies to enhance target production. While cyanobacterial synthetic biology toolkits are less developed than common chassis such as E. coli and yeast, significant progress has been made. The following section highlights the recent advances of synthetic biology strategies used to enhance heterologous expression in cyanobacteria (Fig. 3). | ||
| Fig. 3 Schematic illustration of key synthetic biology parts for heterologous production of cyanobacterial NPs. (A) An expression cassette consists of a promoter, ribosome-binding site (RBS), gene of interest (GOI), and a terminator, essential for controlled gene expression. (B) Two types of plasmids utilized for BGC cloning. A replicative plasmid includes an origin of replication (ori) for independent maintenance in the host. An integrative plasmid features homology arms (depicted in blue and red) to facilitate targeted genomic integration via homologous recombination. (C) CRISPR-Cas9/Cpf1 and CRISPRi-dCas9 tools for host engineering. | ||
3.1 Synthetic biology toolkits for cyanobacterial BGC engineering
Standardized genetic elements, including promoters, RBSs, and terminators, are essential for fine-tuning heterologous BGC expression by regulating transcription and translation (Fig. 3A). However, these components must be optimized for organism-specific requirements such as GC content, regulatory signals, and codon usage. While broad-host-range genetic parts remain limited, strain-specific genetic tools are becoming increasingly accessible.34Among all synthetic biology parts, promoter refactoring has been a primary focus for optimizing cyanobacterial BGC expression. Promoters, either constitutive or inducible, regulate downstream gene expression in response to environmental factors such as light, intracellular CO2 concentration, metal ions (Ni2+, Co2+, Cu2+), and metabolites like nitrate. Many cyanobacterial promoters have been adapted from those used for E. coli, including tetracycline, rhamnose, and IPTG-inducible systems.35 Promoter engineering has significantly enhanced NP production. For instance, refactoring the shinorine (8). BGC from Fischerella sp. PCC 9339 with the strong constitutive Pcpc560 promoter in Synechocystis 6803 increased the yield 10-fold to 2.37 ± 0.21 mg g−1 DCW (Table 1).27 Similarly, heterologous expression of hapalindoles (9) in Synechococcus 2973 was achieved by reconstructing the native BGC from Fischerella ambigua UTEX 1903 into synthetic operons with Ptrc, PcpcB, and PJ23119 promoters (Table 1), reaching titers of 0.75–3 mg L−1.26 A similar increase in yield was observed when the native promoter in the lyngbyatoxin A (1) BGC was replaced with the constitutive promoter PglnA (Table 1, Fig. 2A).16,18 Moreover, recent studies highlight the importance of promoter selection for both expression and toxicity mitigation. For instance, the tolypodiol (5) BGC was only successfully expressed under the PpetE promoter (Table 1), not its native counterpart.21 Similarly, heterologous expression of APK (4) in Anabaena 7120 was initially unsuccessful with the constitutive PTrc promoter likely due to the product toxicity. This challenge was overcome by switching to rhamnose- and cobalt-inducible promoters (Table 1 and Fig. 2B), achieving a yield of 10 mg L−1 of APK (4).22 These findings underscore the role of promoter tuning in both yield improvement and toxicity management, a crucial factor given that the resistance mechanisms of many cyanobacterial NPs have been poorly characterized. However, promoter function can vary across species, necessitating the development of well-characterized promoter libraries tested in multiple cyanobacterial strains. The CyanoGate MoClo system assembled a suite of constitutive and inducible promoters for diverse cyanobacteria,30,34 though its application in optimizing BGC expression remains limited. Expanding such engineering strategies could unlock new opportunities in cyanobacterial NP discovery and drug development.
In addition to promoters, terminators play a crucial role in transcriptional control during heterologous expression. These sequences, located downstream of a gene, signal the end of transcription by facilitating the release of the transcript from the transcription complex. Terminators are classified into two main types, Rho-dependent and Rho-independent (intrinsic) terminators. However, no homologs of Rho have been identified in cyanobacterial genomes to date,36 and Rho-independent terminators are the primary mechanism of transcription termination in cyanobacteria. A recent work screened 19 intrinsic terminators in Synechocystis 6803 and observed varying efficiencies,37 underscoring the importance of testing terminator strength in different cyanobacterial hosts. On the other hand, a study evaluating 34 terminators in E. coli, Synechococcus 2973, and Synechocystis 6803 identified five terminators with >96% termination efficiency across all three species,38 suggesting that some terminators function consistently in both heterotrophic and cyanobacterial hosts. Despite their importance, terminator optimization for improving transcription efficiency in heterologous expression of cyanobacterial NPs remains underexplored. However, widely used E. coli terminators, such as rrnB T1 and T7 terminator, have been successfully applied for both protein and BGC expression in cyanobacteria.34 Expanding studies on terminator engineering could further enhance gene expression control in cyanobacterial synthetic biology applications, particularly for the controlled expression of multiple genes in the BGCs.
Beyond promoters and terminators, RBSs play a crucial role by influencing translation efficiency and, consequently, product formation. RBS sequences are transcribed and recognized by ribosomes to initiate translation, and their modification allows for fine-tuned control of protein expression. The strength of both native Synechocystis 6803 RBS sequences and synthetic variants has been characterized.39 In addition to direct RBS engineering, various regulatory strategies have been employed to modulate expression levels in cyanobacteria, including riboregulators,40 NOT gates,41 and riboswitches.42 Several computational tools exist for predicting optimized RBS sequences.43 However, these were primarily developed for E. coli and have not been optimized for cyanobacteria. Furthermore, to our knowledge, optimized RBSs have yet to be systematically evaluated for enhancing heterologous production of cyanobacterial NPs in cyanobacterial hosts, presenting a promising yet unexplored opportunity.
3.2 Expression construct generation and delivery
In metabolic engineering and synthetic biology, assembling genetic parts into replicable and expressible DNA molecules is essential for constructing functional pathways. A replicative or integrative plasmid is critical for the rapid prototyping of metabolic pathways or regulatory elements (Fig. 3B). Replicative plasmids are extrachromosomal, circular DNA elements that are autonomously maintained. The origin of replication (ori) determines the plasmid's replication initiation site (Fig. 3B) and influences its copy number, which in turn affects gene dosage and expression levels. While replicative plasmids facilitate rapid functional testing, their variable copy number makes precise control of gene expression challenging. Additionally, they require selectable markers to ensure plasmid maintenance through successive cell divisions.Several origins of replication have been successfully used in cyanobacteria. The broad-host-range ori RSF1010 functions across diverse species,44 while pCA and pCB support replication in Synechocystis 6803 and pANS (also called pUH24) is functional in Synechococcus 7942. Another broad-host-range replicon, RK2, has also been shown to support replication in various cyanobacteria.34 Recently, the Watanabe group identified and characterized a Rep protein from Synechocystis 6803, enabling the development of broad-host-range vectors applicable to multiple cyanobacterial species.45 Chimeric shuttle vectors incorporating distinct origins of replication (ori) have been engineered to support dual-host maintenance, enabling replication in both E. coli (for cloning and assembly) and specific cyanobacterial species. Cyanobacterial plasmid replicons tested so far include pDC1 from Nostoc sp. MAC PCC 8009, pDU1 from Nostoc sp. PCC 7524, pAQ1 from Synechococcus 7002, pBA1 from Anacystis nidulans 6301, pFDA from Fremyella diplosiphon, and pANS.46
In integrative plasmids, a homology arm complementary to a target locus in the host genome enables site-specific integration of the plasmid, carrying the expression cassette (gene of interest, GOI, or BGC), via homologous recombination (Fig. 3B). This approach allows for the stable maintenance of heterologous functions in cyanobacterial hosts and is also commonly used for gene knockouts in loss-of-function studies. In cyanobacteria, “neutral sites” are chromosomal loci where foreign DNA can be integrated without disrupting metabolism or cellular physiology. For example, pSyn6 is a widely used integrative vector used for protein expression in Synechococcus strains, which includes built-in homology arms for targeted genome integration, while 51 putative neutral sites were identified in the Synechococcus 7002 genome.47 Similarly, three major chromosomal neutral sites (NS1, NS2, NS3) have been established for the engineering of Synechococcus 7942.32 For Anabaena 7120, several neutral sites have been used for integrative vectors, while it was further engineered to carry the NS2 site of Synechococcus 7942 in its genome, allowing for heterologous expression of NPs such as cryptomaldamide (6) and columbamide (7).19,20 Further exploration and characterization of neutral sites in cyanobacterial chassis could enable multiplexed genetic engineering, paving the way for more sophisticated synthetic biology applications in cyanobacteria, particularly for the heterologous production of cyanobacterial NPs.
Whether replicative or integrative expression, the next fundamental step after selecting a plasmid is assembling genetic parts within it for functional characterization. While chemical DNA synthesis is economically feasible for relatively short sequences (e.g., <10 kb), constructing larger fragments still depends on enzymatic assembly methods.13 Several DNA assembly techniques have been developed, building on traditional restriction digestion and ligation methods. Golden Gate assembly uses type IIS restriction enzymes for seamless cloning, while alternative strategies such as Gibson assembly and DNA Assembler leverage different enzymatic mechanisms. For manipulating larger BGCs, recombination-based methods like lambda Red (λ/Red) and Rac phage protein (RecET) in E. coli have been widely utilized.48 Our recent work demonstrated the application of Gibson assembly for cloning the APK (4) BGC, while λ/Red recombination was employed to enhance the efficiency of promoter exchange (Fig. 2B).22 Additionally, transformation-associated recombination (TAR) has been utilized for assembling large constructs containing cyanobacterial BGCs in Saccharomyces cerevisiae, followed by heterologous expression in cyanobacteria (Table 1).
Efficient heterologous expression can be hindered by the presence of rare codons or limited tRNA availability, particularly when expressing genes from distantly related species. Codon optimization, which adjusts the nucleotide sequence to match the codon usage preferences of the target host, is an effective strategy for improving BGC expression. Indeed, codon optimization has been employed to produce indolactams (2, 3),16 APK (4, Fig. 2)22 and tolypodiol (5)21 in Anabaena 7120. Additionally, the cloning of rare tRNA genes has been used to enhance NP production, as demonstrated in the heterologous expression of epothilone in Burkholderiales DSM 7029, where this strategy significantly improved titers.49 Though not yet explored in cyanobacteria, incorporating rare tRNAs could support the heterologous production of a broader range of non-native compounds.
Regardless of whether a BGC is cloned through DNA synthesis or enzymatic assembly, its efficient delivery into a cyanobacterial host is essential for successful heterologous production. Many species require facilitated transformation methods to introduce foreign DNA. Electroporation, commonly used for Anabaena 7120, employs an electric field to temporarily increase membrane permeability, allowing plasmid entry. Alternatively, conjugation is an effective method for transformation-resistant hosts like Synechococcus 2973 and Anabaena 7120, relying on direct cell-to-cell contact for DNA transfer. Most transformations require antibiotic resistance markers for selection, raising concerns about the potential release of resistance genes into the environment. To mitigate this issue, markerless genetic engineering strategies are being explored as an alternative. Recently, a two-plasmid CRE recombinase system was developed for Synechocystis 6803, enabling the generation of markerless mutants and providing a more sustainable approach to genetic modification.50
3.3 Host engineering to support heterologous production
Cyanobacteria offer a promising platform for sustainable bioproduction of bioactive cyanobacterial NPs. Metabolic engineering of the cyanobacterial hosts can play a crucial role in optimizing secondary metabolite production by enhancing precursor availability, optimizing cofactor supply, and regulating metabolic flux. These strategies focus on enriching target pathways while minimizing competition from unwanted metabolic routes, ensuring efficient biosynthesis. In cyanobacteria, these strategies have been successfully applied to increase the production of fatty acids, carbohydrates, and terpenoids.51 More recently, systems biology-driven metabolic engineering has been explored to enhance secondary metabolite production for industrial applications.52 However, the application of host engineering and systems biology approaches to heterologous expression of cyanobacterial NPs remains largely unexplored, awaiting further development.Genome-editing technologies such as CRISPR-Cas9 hold immense potential for advancing cyanobacterial strain development (Fig. 3C), enabling precise metabolic rewiring, multi-gene editing, and faster genetic segregation. However, high Cas9 expression is toxic to cyanobacterial cells, posing a challenge for widespread adoption. To address this, a transient Cas9 expression system using the pCRISPomyces-2 vector was employed to successfully knock out endogenous genes like nblA in Synechococcus 2973.53 Compared to Cas9, Cpf1 (now known as Cas12a) exhibits lower toxicity and has been adapted for genome editing in fast-growing cyanobacteria such as Synechococcus 2973 and S. elongatus PCC 11801.54 Notably, a single-plasmid system (pSL2680) was designed, which carried Cpf1 under the lac promoter and crRNA under the PJ23119 promoter, along with homology arms for targeted gene editing. Similarly, the CRISPR-Cpf1 system was utilized to integrate the Synechococcus 7942, NS2 neutral site into the Anabaena PCC 7120 chromosome.19 These advancements demonstrate the growing potential of CRISPR-based genome editing in cyanobacteria chassis, paving the way for more efficient heterologous production of cyanobacterial NPs.
4. Conclusions and perspectives
Despite the vast diversity of BGCs revealed through metagenomic sequencing of cyanobacterial samples, many strains remain unculturable due to unknown or highly specific growth requirements.5 Additionally, challenges associated with axenic culture isolation and the lack of robust genetic tools hinder both BGC characterization and the development of optimized production strains. As a result, the discovery of cyanobacterial NPs has largely relied on direct extraction from environmental samples, which has inherent limitations. Heterologous expression provides a promising alternative, enabling the production of cyanobacterial NPs in genetically tractable model hosts.This highlight has outlined recent advances in synthetic biology strategies for expressing cyanobacterial NPs in cyanobacterial hosts. While Anabaena PCC 7120 has demonstrated success in heterologous NP production, high-yield biosynthesis remains a challenge. As metagenomic sequencing efforts rapidly expand, a wider range of BGCs should be explored across different cyanobacterial chassis to enhance NP discovery. The continued development of genetic and metabolic engineering tools will be critical for optimizing cyanobacteria as sustainable production platforms for synthesizing structurally diverse cyanobacterial NPs.
Genome engineering tools, particularly CRISPR-Cas systems, are expected to revolutionize cyanobacterial strain engineering, enhancing the synthetic capability of chassis. Beyond conventional gene knockout and integration, CRISPRi-dCas9 (a catalytically inactive version of Cas9) has been explored for transcriptional regulation in diverse cyanobacteria (Fig. 3C).55 However, a major bottleneck in CRISPR-based genome editing is the dependence on host recombination machinery for DNA insertion, leading to low efficiency and accuracy. A groundbreaking approach to overcome this limitation is RNA-guided transposition with CRISPR-associated transposases (CAST), recently demonstrated in Anabaena PCC 7120.56 This system precisely inserts genetic payloads 63 bases downstream of the protospacer adjacent motif (PAM) without activating endogenous transposons, making it a powerful tool for genome editing in both cultured and even complex cyanobacterial communities.
Beyond genetic engineering, machine learning and deep learning are emerging as transformative tools in synthetic biology. These computational approaches have been successfully applied to gene expression optimization and metabolic engineering, though their potential for cyanobacterial NP heterologous production remains largely untapped. Additionally, systems biology approaches are being increasingly utilized to refine computational simulations, reducing the number of wet-lab experiments required for robust genetic design. These methods focus on two key aspects: (1) predicting global metabolic flux distributions using genome-scale metabolic (GSM) models, and (2) calculating mass balance of metabolites through 13C-metabolic flux analysis (MFA). While GSM models have been developed for a few cyanobacterial species and successfully applied to chemical biosynthesis, recent advances in isotopically nonstationary 13C MFA have enabled real-time tracking of carbon flux in Synechococcus 7942 and 2973,57 providing deeper insights into carbon assimilation and biochemical pathway regulation. Moreover, recent breakthroughs in massively parallel assays, biophysical modeling, and AI-driven synthetic biology have significantly improved our understanding of gene regulation and transcriptional control in E. coli.58 Integrating these techniques with cyanobacterial synthetic biology is expected to accelerate the rational design of genetic parts, enabling more efficient metabolic engineering strategies.
Overall, significant progress has been made in developing synthetic biology approaches for cyanobacterial heterologous expression, leading to the successful production of several cyanobacterial NPs in engineered cyanobacterial hosts (Table 1 and Fig. 1). The application of emerging engineering strategies, computational modeling, and machine learning will likely enhance cyanobacterial chassis development, unlocking the full potential of cyanobacteria for discovering new cyanobacterial NPs directly from genomes and producing complex and industrially relevant NPs.
5. Conflicts of interest
H. L. is co-founder of Oceanyx Pharmaceuticals, Inc. that has licensed patents and patent applications related to cyanobacterial natural products. The other authors do not have conflicts to declare.6. Acknowledgements
This work is supported by NIH grant R35GM128742 (Y. D.) and NIH grant RM1GM145426 (H. L., Y. D.). H. L. is also supported by the Debbie and Sylvia DeSantis Chair professorship (H. L.). M. C. and D. D. are partially supported by the Division of Sponsored Research and College of Pharmacy at the University of Florida. Fig. 3 was created using BioRender.7. References
- N. Wan, D. M. DeLorenzo, L. He, L. You, C. M. Immethun, G. Wang, E. E. K. Baidoo, W. Hollinshead, J. D. Keasling, T. S. Moon and Y. J. Tang, Biotechnol. Bioeng., 2017, 114, 1593–1602 CrossRef CAS PubMed.
- M. B. Weiss, R. M. Borges, P. Sullivan, J. P. B. Domingues, F. H. S. da Silva, V. G. S. Trindade, S. Luo, J. Orjala and C. M. Crnkovic, Nat. Prod. Rep., 2025, 42, 6–49 RSC.
- J. Demay, C. Bernard, A. Reinhardt and B. Marie, Mar. Drugs, 2019, 17, 320 CrossRef CAS PubMed.
- H. Luesch, E. K. Ellis, Q. Y. Chen and R. Ratnayake, Nat. Prod. Rep., 2025, 42, 208–256 RSC.
- M. Baunach, A. Guljamow, M. Miguel-Gordo and E. Dittmann, Nat. Prod. Rep., 2024, 41, 347–369 RSC.
- F. I. Saldivar-Gonzalez, V. D. Aldas-Bulos, J. L. Medina-Franco and F. Plisson, Chem. Sci., 2022, 13, 1526–1546 RSC.
- E. W. Sayers, J. Beck, E. E. Bolton, J. R. Brister, J. Chan, D. C. Comeau, R. Connor, M. DiCuccio, C. M. Farrell, M. Feldgarden, A. M. Fine, K. Funk, E. Hatcher, M. Hoeppner, M. Kane, S. Kannan, K. S. Katz, C. Kelly, W. Klimke, S. Kim, A. Kimchi, M. Landrum, S. Lathrop, Z. Lu, A. Malheiro, A. Marchler-Bauer, T. D. Murphy, L. Phan, A. B. Prasad, S. Pujar, A. Sawyer, E. Schmieder, V. A. Schneider, C. L. Schoch, S. Sharma, F. Thibaud-Nissen, B. W. Trawick, T. Venkatapathi, J. Wang, K. D. Pruitt and S. T. Sherry, Nucleic Acids Res., 2024, 52, D33–D43 CrossRef CAS PubMed.
- L. R. Moore, R. Caspi, D. A. Campbell, J. R. Casey, S. Crevecoeur, D. J. Lea-Smith, B. Long, N. M. Omar, S. M. Paley, N. M. Schmelling, A. Torrado, J. P. Zehr and P. D. Karp, Front. Microbiol., 2024, 15, 1340413 CrossRef PubMed.
- K. Blin, S. Shaw, H. E. Augustijn, Z. L. Reitz, F. Biermann, M. Alanjary, A. Fetter, B. R. Terlouw, W. W. Metcalf, E. J. N. Helfrich, G. P. van Wezel, M. H. Medema and T. Weber, Nucleic Acids Res., 2023, 51, W46–W50 CrossRef CAS PubMed.
- M. M. Zdouc, K. Blin, N. L. L. Louwen, J. Navarro, C. Loureiro, C. D. Bader, C. B. Bailey, L. Barra, T. J. Booth, K. A. J. Bozhuyuk, J. D. D. Cediel-Becerra, Z. Charlop-Powers, M. G. Chevrette, Y. H. Chooi, P. M. D'Agostino, T. de Rond, E. Del Pup, K. R. Duncan, W. Gu, N. Hanif, E. J. N. Helfrich, M. Jenner, Y. Katsuyama, A. Korenskaia, D. Krug, V. Libis, G. A. Lund, S. Mantri, K. D. Morgan, C. Owen, C. S. Phan, B. Philmus, Z. L. Reitz, S. L. Robinson, K. S. Singh, R. Teufel, Y. Tong, F. Tugizimana, D. Ulanova, J. M. Winter, C. Aguilar, D. Y. Akiyama, S. A. A. Al-Salihi, M. Alanjary, F. Alberti, G. Aleti, S. A. Alharthi, M. Y. A. Rojo, A. A. Arishi, H. E. Augustijn, N. E. Avalon, J. A. Avelar-Rivas, K. K. Axt, H. B. Barbieri, J. C. J. Barbosa, L. G. Barboza Segato, S. E. Barrett, M. Baunach, C. Beemelmanns, D. Beqaj, T. Berger, J. Bernaldo-Aguero, S. M. Bettenbuhl, V. A. Bielinski, F. Biermann, R. M. Borges, R. Borriss, M. Breitenbach, K. M. Bretscher, M. W. Brigham, L. Buedenbender, B. W. Bulcock, C. Cano-Prieto, J. Capela, V. J. Carrion, R. S. Carter, R. Castelo-Branco, G. Castro-Falcon, F. O. Chagas, E. Charria-Giron, A. A. Chaudhri, V. Chaudhry, H. Choi, Y. Choi, R. Choupannejad, J. Chromy, M. S. C. Donahey, J. Collemare, J. A. Connolly, K. E. Creamer, M. Crusemann, A. A. Cruz, A. Cumsille, J. F. Dallery, L. C. Damas-Ramos, T. Damiani, M. de Kruijff, B. D. Martin, G. D. Sala, J. Dillen, D. T. Doering, S. R. Dommaraju, S. Durusu, S. Egbert, M. Ellerhorst, B. Faussurier, A. Fetter, M. Feuermann, D. P. Fewer, J. Foldi, A. Frediansyah, E. A. Garza, A. Gavriilidou, A. Gentile, J. Gerke, H. Gerstmans, J. P. Gomez-Escribano, L. A. Gonzalez-Salazar, N. E. Grayson, C. Greco, J. E. G. Gomez, S. Guerra, S. G. Flores, A. Gurevich, K. Gutierrez-Garcia, L. Hart, K. Haslinger, B. He, T. Hebra, J. L. Hemmann, H. Hindra, L. Hoing, D. C. Holland, J. E. Holme, T. Horch, P. Hrab, J. Hu, T. H. Huynh, J. Y. Hwang, R. Iacovelli, D. Iftime, M. Iorio, S. Jayachandran, E. Jeong, J. Jing, J. J. Jung, Y. Kakumu, E. Kalkreuter, K. B. Kang, S. Kang, W. Kim, G. J. Kim, H. Kim, H. U. Kim, M. Klapper, R. A. Koetsier, C. Kollten, A. T. Kovacs, Y. Kriukova, N. Kubach, A. M. Kunjapur, A. K. Kushnareva, A. Kust, J. Lamber, M. Larralde, N. J. Larsen, A. P. Launay, N. T. Le, S. Lebeer, B. T. Lee, K. Lee, K. L. Lev, S. M. Li, Y. X. Li, C. Licona-Cassani, A. Lien, J. Liu, J. A. V. Lopez, N. V. Machushynets, M. I. Macias, T. Mahmud, M. Maleckis, A. M. Martinez-Martinez, Y. Mast, M. F. Maximo, C. M. McBride, R. M. McLellan, K. M. Bhatt, C. Melkonian, A. Merrild, M. Metsa-Ketela, D. A. Mitchell, A. V. Muller, G. S. Nguyen, H. T. Nguyen, T. H. J. Niedermeyer, J. H. O'Hare, A. Ossowicki, B. O. Ostash, H. Otani, L. Padva, S. Paliyal, X. Pan, M. Panghal, D. S. Parade, J. Park, J. Parra, M. P. Rubio, H. T. Pham, S. J. Pidot, J. Piel, B. Pourmohsenin, M. Rakhmanov, S. Ramesh, M. H. Rasmussen, A. Rego, R. Reher, A. J. Rice, A. Rigolet, A. Romero-Otero, L. R. Rosas-Becerra, P. Y. Rosiles, A. Rutz, B. Ryu, L. A. Sahadeo, M. Saldanha, L. Salvi, E. Sanchez-Carvajal, C. Santos-Medellin, N. Sbaraini, S. M. Schoellhorn, C. Schumm, L. Sehnal, N. Selem, A. D. Shah, T. K. Shishido, S. Sieber, V. Silviani, G. Singh, H. Singh, N. Sokolova, E. C. Sonnenschein, M. Sosio, S. T. Sowa, K. Steffen, E. Stegmann, A. B. Streiff, A. Struder, F. Surup, T. Svenningsen, D. Sweeney, J. Szenei, A. Tagirdzhanov, B. Tan, M. J. Tarnowski, B. R. Terlouw, T. Rey, N. U. Thome, L. R. Torres Ortega, T. Torring, M. Trindade, A. W. Truman, M. Tvilum, D. W. Udwary, C. Ulbricht, L. Vader, G. P. van Wezel, M. Walmsley, R. Warnasinghe, H. G. Weddeling, A. N. M. Weir, K. Williams, S. E. Williams, T. E. Witte, S. M. W. Rocca, K. Yamada, D. Yang, D. Yang, J. Yu, Z. Zhou, N. Ziemert, L. Zimmer, A. Zimmermann, C. Zimmermann, J. J. J. van der Hooft, R. G. Linington, T. Weber and M. H. Medema, Nucleic Acids Res., 2025, 53, D678–D690 CrossRef PubMed.
- J. C. Navarro-Munoz, N. Selem-Mojica, M. W. Mullowney, S. A. Kautsar, J. H. Tryon, E. I. Parkinson, E. L. C. De Los Santos, M. Yeong, P. Cruz-Morales, S. Abubucker, A. Roeters, W. Lokhorst, A. Fernandez-Guerra, L. T. D. Cappelini, A. W. Goering, R. J. Thomson, W. W. Metcalf, N. L. Kelleher, F. Barona-Gomez and M. H. Medema, Nat. Chem. Biol., 2020, 16, 60–68 CrossRef CAS PubMed.
- D. W. Udwary, D. T. Doering, B. Foster, T. Smirnova, S. A. Kautsar and N. J. Mouncey, Nucleic Acids Res., 2025, 53, D717–D723 CrossRef PubMed.
- D. Dhakal, M. Chen, H. Luesch and Y. Ding, J. Ind. Microbiol. Biotechnol., 2021, 48, kuab003 CrossRef CAS PubMed.
- G. C. Gordon and B. F. Pfleger, Adv. Exp. Med. Biol., 2018, 1080, 281–315 CrossRef CAS PubMed.
- R. Stebegg, B. Wurzinger, M. Mikulic and G. Schmetterer, J. Bacteriol., 2012, 194, 4601–4607 CrossRef CAS PubMed.
- P. Videau, K. N. Wells, A. J. Singh, J. Eiting, P. J. Proteau and B. Philmus, ACS Synth. Biol., 2020, 9, 63–75 CrossRef CAS PubMed.
- G. Yang, Y. Zhang, N. K. Lee, M. A. Cozad, S. E. Kearney, H. Luesch and Y. Ding, Sci. Rep., 2017, 7, 11888 CrossRef PubMed.
- P. Videau, K. N. Wells, A. J. Singh, W. H. Gerwick and B. Philmus, ACS Synth. Biol., 2016, 5, 978–988 CrossRef CAS PubMed.
- A. Taton, A. Ecker, B. Diaz, N. A. Moss, B. Anderson, R. Reher, T. F. Leao, R. Simkovsky, P. C. Dorrestein, L. Gerwick, W. H. Gerwick and J. W. Golden, ACS Synth. Biol., 2020, 9, 3364–3376 CrossRef CAS PubMed.
- A. Taton, S. Rohrer, B. Diaz, R. Reher, A. M. Caraballo Rodriguez, M. L. Pierce, P. C. Dorrestein, L. Gerwick, W. H. Gerwick and J. W. Golden, ACS Chem. Biol., 2022, 17, 1910–1923 CrossRef CAS PubMed.
- D. Back, T. J. O'Donnell, K. K. Axt, J. R. Gurr, J. M. Vanegas, P. G. Williams and B. Philmus, ACS Chem. Biol., 2023, 18, 1797–1807 CrossRef CAS PubMed.
- D. Dhakal, D. Kallifidas, M. Chen, S. Kokkaliari, Q. Y. Chen, V. J. Paul, Y. Ding and H. Luesch, Org. Lett., 2023, 25, 2238–2242 CrossRef CAS PubMed.
- P. van Alphen, H. Abedini Najafabadi, F. Branco Dos Santos and K. J. Hellingwerf, Biotechnol. J., 2018, 13, e1700764 CrossRef PubMed.
- J. Yu, M. Liberton, P. F. Cliften, R. D. Head, J. M. Jacobs, R. D. Smith, D. W. Koppenaal, J. J. Brand and H. B. Pakrasi, Sci. Rep., 2015, 5, 8132 CrossRef CAS PubMed.
- C. J. Knoot, J. Ungerer, P. P. Wangikar and H. B. Pakrasi, J. Biol. Chem., 2018, 293, 5044–5052 CrossRef CAS PubMed.
- C. J. Knoot, Y. Khatri, R. M. Hohlman, D. H. Sherman and H. B. Pakrasi, ACS Synth. Biol., 2019, 8, 1941–1951 CrossRef CAS PubMed.
- G. Yang, M. A. Cozad, D. A. Holland, Y. Zhang, H. Luesch and Y. Ding, ACS Synth. Biol., 2018, 7, 664–671 CrossRef CAS PubMed.
- B. Bourgade and K. Stensjo, Front. Microbiol., 2022, 13, 994365 CrossRef PubMed.
- X. Han, W. Wang and X. Lu, Algal Res., 2023, 74, 103212 CrossRef.
- A. J. Victoria, T. T. Selao, J. A. Moreno-Cabezuelo, L. A. Mills, G. A. R. Gale, D. J. Lea-Smith and A. J. McCormick, Plant Physiol., 2024, 196, 1674–1690 CrossRef CAS PubMed.
- M. Griese, C. Lange and J. Soppa, FEMS Microbiol. Lett., 2011, 323, 124–131 CrossRef CAS PubMed.
- A. Taton, C. Erikson, Y. Yang, B. E. Rubin, S. A. Rifkin, J. W. Golden and S. S. Golden, Nat. Commun., 2020, 11, 1688 CrossRef CAS PubMed.
- L. F. Brey, A. J. Wlodarczyk, J. F. Bang Thofner, M. Burow, C. Crocoll, I. Nielsen, A. J. Zygadlo Nielsen and P. E. Jensen, Metab. Eng., 2020, 57, 129–139 CrossRef CAS PubMed.
- R. Vasudevan, G. A. R. Gale, A. A. Schiavon, A. Puzorjov, J. Malin, M. D. Gillespie, K. Vavitsas, V. Zulkower, B. Wang, C. J. Howe, D. J. Lea-Smith and A. J. McCormick, Plant Physiol., 2019, 180, 39–55 CrossRef CAS PubMed.
- T. Sun, S. Li, X. Song, J. Diao, L. Chen and W. Zhang, Biotechnol. Adv., 2018, 36, 1293–1307 CrossRef CAS PubMed.
- C. J. Ramey, A. Baron-Sola, H. R. Aucoin and N. R. Boyle, ACS Synth. Biol., 2015, 4, 1186–1196 CrossRef CAS PubMed.
- C. L. Kelly, G. M. Taylor, A. Satkute, L. Dekker and J. T. Heap, Microorganisms, 2019, 7, 263 CrossRef CAS PubMed.
- G. A. R. Gale, B. Wang and A. J. McCormick, Front. Microbiol., 2020, 11, 624011 CrossRef PubMed.
- E. Englund, F. Liang and P. Lindberg, Sci. Rep., 2016, 6, 36640 CrossRef CAS PubMed.
- K. Abe, Y. Sakai, S. Nakashima, M. Araki, W. Yoshida, K. Sode and K. Ikebukuro, Biotechnol. Lett., 2014, 36, 287–294 CrossRef CAS PubMed.
- A. Taton, A. T. Ma, M. Ota, S. S. Golden and J. W. Golden, ACS Synth. Biol., 2017, 6, 2175–2182 CrossRef CAS PubMed.
- Y. Nakahira, A. Ogawa, H. Asano, T. Oyama and Y. Tozawa, Plant Cell Physiol., 2013, 54, 1724–1735 CrossRef CAS PubMed.
- H. M. Salis, E. A. Mirsky and C. A. Voigt, Nat. Biotechnol., 2009, 27, 946–950 CrossRef CAS PubMed.
- B. Bishe, A. Taton and J. W. Golden, iScience, 2019, 20, 216–228 CrossRef CAS PubMed.
- K. Ohdate, M. Sakata, K. Maeda, Y. Sakamaki, K. Nimura-Matsune, R. Ohbayashi, W. R. Hess and S. Watanabe, Front. Microbiol., 2024, 15, 1311290 CrossRef PubMed.
- A. Taton, F. Unglaub, N. E. Wright, W. Y. Zeng, J. Paz-Yepes, B. Brahamsha, B. Palenik, T. C. Peterson, F. Haerizadeh, S. S. Golden and J. W. Golden, Nucleic Acids Res., 2014, 42, e136 CrossRef PubMed.
- M. Wang, G. Luan and X. Lu, J. Biotechnol., 2019, 295, 37–40 CrossRef CAS PubMed.
- J. Fu, X. Bian, S. Hu, H. Wang, F. Huang, P. M. Seibert, A. Plaza, L. Xia, R. Muller, A. F. Stewart and Y. Zhang, Nat. Biotechnol., 2012, 30, 440–446 CrossRef CAS PubMed.
- X. Bian, B. Tang, Y. Yu, Q. Tu, F. Gross, H. Wang, A. Li, J. Fu, Y. Shen, Y. Z. Li, A. F. Stewart, G. Zhao, X. Ding, R. Muller and Y. Zhang, ACS Chem. Biol., 2017, 12, 1805–1812 CrossRef CAS PubMed.
- C. M. Jones, S. Parrish and D. R. Nielsen, ACS Synth. Biol., 2021, 10, 2371–2382 CrossRef CAS PubMed.
- A. Satta, L. Esquirol and B. E. Ebert, Microorganisms, 2023, 11 CrossRef CAS PubMed.
- H. U. Kim, P. Charusanti, S. Y. Lee and T. Weber, Nat. Prod. Rep., 2016, 33, 933–941 RSC.
- K. E. Wendt, J. Ungerer, R. E. Cobb, H. Zhao and H. B. Pakrasi, Microb. Cell Fact., 2016, 15, 115 CrossRef PubMed.
- J. Ungerer and H. B. Pakrasi, Sci. Rep., 2016, 6, 39681 CrossRef CAS PubMed.
- X. D. Li, L. M. Liu, Y. C. Xi, Q. W. Sun, Z. Luo, H. L. Huang, X. W. Wang, H. B. Jiang and W. Chen, Commun. Biol., 2024, 7, 994 CrossRef CAS PubMed.
- S. Arevalo, D. Perez Rico, D. Abarca, L. W. Dijkhuizen, C. Sarasa-Buisan, P. Lindblad, E. Flores, S. Nierzwicki-Bauer and H. Schluepmann, ACS Synth. Biol., 2024, 13, 901–912 CrossRef CAS PubMed.
- A. Srivastava and P. Shukla, Trends Biotechnol., 2022, 40, 4–7 CrossRef CAS PubMed.
- T. L. LaFleur, A. Hossain and H. M. Salis, Nat. Commun., 2022, 13, 5159 CrossRef CAS PubMed.
Footnote |
| † Equal contributions. |
| This journal is © The Royal Society of Chemistry 2025 |