
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicencePrivileged natural product compound classes for anti-inflammatory drug development
Malcolm Z. Y.
Choo
a,
Julian A. T.
Chua
a,
Sean X. Y.
Lee
 a,
Yuet
Ang
b,
W. S. Fred
Wong
bc and
Christina L. L.
Chai
*ac
a,
Yuet
Ang
b,
W. S. Fred
Wong
bc and
Christina L. L.
Chai
*ac
aDepartment of Pharmacy and Pharmaceutical Sciences, National University of Singapore, 18 Science Drive 4, 117543, Singapore. E-mail: phacllc@nus.edu.sg
bDepartment of Pharmacology, Yong Loo Lin School of Medicine, National University Health System, 16 Medical Drive, 117600, Singapore. E-mail: phcwongf@nus.edu.sg
cDrug Discovery and Optimization Platform, Yong Loo Lin School of Medicine, National University Health System, 117600, Singapore
First published on 11th March 2025
Abstract
Covering: up to early 2025
Privileged compound classes of anti-inflammatory natural products are those where there are many reported members that possess anti-inflammatory properties. The identification of these classes is of particular relevance to drug discovery, as they could serve as valuable starting points in developing effective and safe anti-inflammatory agents. The privileged compound classes of natural products include the polyphenols, coumarins, labdane diterpenoids, sesquiterpene lactones, isoquinoline and indole alkaloids, each offering a variety of molecular scaffolds and functional groups that enable diverse interactions with biological targets. From a medicinal chemistry point of view, natural products are both a boon and a bane. The multi-targeting nature of natural products is a boon in the treatment of multi-factorial diseases such as inflammation, but promiscuity, poor potency and pharmacokinetic properties are significant hurdles that must be addressed to ensure these compounds can be effectively used as therapeutics. In addition, there are continued controversies regarding the efficacies of some of these natural products that will continue to polarise their use. In this review, examples of natural products of six privileged compound classes will be discussed for their potential use and possible further development as anti-inflammatory drugs.
 Malcolm Z. Y. Choo | Dr Malcolm Choo received his BSc (Hons) in Chemistry and PhD in Medicinal Chemistry from the National University of Singapore (NUS). His PhD research focuses on nature-inspired development of small molecule anti-inflammatory drugs under the supervision of Prof. Christina Chai, and co-supervision by Prof. W. S. Fred Wong. His research interests lie in the syntheses of natural products and their use as therapeutics against various diseases. |
 Julian A. T. Chua | Mr Julian Chua received his BSc (Hons) in Pharmaceutical Science from the National University of Singapore (NUS). He is currently pursuing a PhD in the Department of Pharmacy and Pharmaceutical Sciences of NUS, under the supervision of Prof Christina Chai. His research interests lie in the use of natural products in drug discovery and development, and his current research is on the nature-inspired design of small molecule drugs against infectious diseases. |
 Sean X. Y. Lee | Mr Sean Lee received his BSc (Hons) in Pharmaceutical Science from the National University of Singapore (NUS). He is currently pursuing his PhD in the Department of Pharmacy and Pharmaceutical Sciences, NUS, under the supervision of Prof. Christina Chai. His current research is on the polypharmacology of natural products for drug discovery and development. |
 Yuet Ang | Ms Ang Yuet is a Research Assistant working under the supervision of Prof. W. S. Fred Wong in the Department of Pharmacology at the National University of Singapore (NUS). Her research primarily focuses on respiratory pharmacology, with an emphasis on exploring the therapeutic effects of naturally derived compounds in models of respiratory diseases, including idiopathic pulmonary fibrosis and pulmonary arterial hypertension. Yuet holds a BSc in Biomedical Science from Griffith University in Queensland, Australia. Her research interests are in disease pathology, cell biology, and molecular biology, with the goal of developing therapeutic strategies in the field of respiratory health. |
 W. S. Fred Wong | Professor W. S. Fred Wong received his B. Pharm (cum laude) from the St. Johns' University, New York and his PhD from Ohio State University. He was a postdoctoral scientist at Lilly Research Laboratories, focusing on respiratory drug development, and a Research Fellow in Medicine at Brigham and Women's Hospital. He joined the Department of Pharmacology at NUS in 1995 and established the Respiratory Pharmacology Laboratory. His research focuses on studying the pathogenesis of airway inflammation, oxidative damage, DNA damage in asthma, COPD, pulmonary fibrosis, and developing mechanism-based pharmacological agents using small molecules, herbal bioactive molecules, natural/nutritional products and antisense oligonucleotides/siRNA. |
 Christina L. L. Chai | Professor Christina Chai obtained her BSc (Hons) from the University of Canterbury, Christchurch, New Zealand and her PhD in synthetic organic chemistry from the Australian National University. She has held academic positions in Victoria University of Wellington NZ; Australian National University; Institute of Chemical and Engineering Sciences, Singapore and is now at the Department of Pharmacy and Pharmaceutical Sciences, National University of Singapore. Professor Chai has received numerous awards, including the Alexander von Humboldt Fellowship and the Violette and Samuel Glasstone Research Fellowship. Her research is on the development of small-molecule therapeutics for diseases such as inflammation and infectious diseases. |
Inflammation is an important defence mechanism of the innate immunity that is responsible for protecting the host against damaging environmental factors and to maintain/restore tissue homeostasis.1 Prolonged and unresolved inflammation, however, can lead to chronic inflammation, which is the underlying cause of a number of debilitating conditions, such as asthma, arthritis, and atherosclerosis, among others.2 The process of inflammation is complex and involves various signalling pathways that can be triggered by pathogens which are recognised by pattern-recognition receptors (PRRs) such as toll-like receptors (TLRs). This then recruits leukocytes to the affected area(s) and triggers downstream signalling pathways, including nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB),3 mitogen-activated protein kinase (MAPK),4 and cyclooxygenase (COX) pathways.5 In general, activation of inflammatory pathways can result in the upregulation of genes that are directly related to inflammation, and/or lead to the production of pro-inflammatory mediators like as tumour necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), nitric oxide (NO), and prostaglandin E2 (PGE2). For example, activation of the NF-κB and MAPK pathways enhances transcriptional activity in the nucleus, resulting in elevated gene expression levels of inflammatory mediators (e.g., TNF-α, NO) and other molecules involved in immune responses.3,6–8 Similarly, activation of the COX pathway results in the production of PGE2.5,9 As elevated or dysregulated levels of these mediators can be harmful to the host, there is much interest in the development of drugs that can inhibit the production of these inflammatory molecules. It is important to note that many inflammatory pathways crosstalk with each other, and thus the activation of one pathway often triggers others.3,10 Consequently, inflammation is recognized as a complex, multi-faceted problem, likely necessitating the modulation of multiple pathways to achieve effective control or cure.
Nature has proven to be an invaluable source of anti-inflammatory compounds, with aspirin serving as a prime example. As one of the most widely used nonsteroidal anti-inflammatory drugs (NSAIDs), aspirin originates from the natural product salicin, the active compound in willow bark.11 The medicinal use of willow bark dates back to ancient times, where it was employed to treat various ailments like fevers and pains.12 Today, aspirin's applications include the treatment and relief of cardiovascular disease symptoms, general pain, and inflammation due to its anti-inflammatory, analgesic, and antiplatelet effects.12 There are numerous other examples of natural products that display anti-inflammatory properties, including curcumin,13 berberine,14 and andrographolide,15–17 and some of these are active components of traditional herbs and medicines. Natural products with anti-inflammatory properties are more prevalent in certain classes of compounds, suggesting that these compound classes may be privileged. The concept of privileged structures was first introduced by Evans to describe sub-structures that are “capable of providing useful ligands for more than one receptor”.18 Over time, this definition has evolved, with contemporary interpretations considering the favourable pharmacokinetic and pharmacodynamic properties associated with privileged structures.19 Others utilise this term to describe compounds sharing the same core scaffold exhibiting a wide range of bioactivities.20–22 In this perspective review, we define privileged compound classes of anti-inflammatory natural products as those where there are many reported members that possess anti-inflammatory properties. The identification of such scaffolds is clearly a boon for drug discovery as they can serve as starting points in the search for hit compounds for attenuating inflammation. This knowledge may accelerate the discovery of small molecule drugs for the treatment of inflammatory disorders, for example by enabling the curation of compound libraries for high throughput screening.
In this perspective, we will highlight six privileged compound classes of natural products for anti-inflammatory drug development. These include the coumarins, polyphenols, labdane diterpenoids, sesquiterpene lactones, isoquinolines and indole alkaloids. Selected examples from the literature will be highlighted to illustrate the potential for applications in anti-inflammatory drug development.
1. Coumarins as a privileged compound class with anti-inflammatory properties
Coumarins, also known as 2H-chromen-2-one, belong to the phenylpropanoid class of natural products that are biosynthesized via the shikimic acid pathway using the amino acids phenylalanine and tyrosine.23 The term “phenylpropanoid” is derived from the structure of the natural products, which feature an aromatic phenyl group and the 3-carbon propene tail of coumaric acid. A key intermediate in the biosynthesis of coumarins is 4-coumaroyl-CoA, a compound that also serves as a critical precursor for the formation of various other natural products including, flavonoids, isoflavonoids, aurones, stilbenes, and many more.24Structurally, coumarins consist of a benzene ring that is fused with a pyrone moiety (Fig. 1A). These heterocyclic compounds are typically found in plants that belong to the natural orders of Orchidaceae, Leguminocaeae, Rutacaeae, Umbelliferae and Labiatae.29 Although coumarins are distributed throughout all parts of the plant, the highest concentrations are typically found in the fruits, followed by the roots, stems and leaves.29 Naturally occurring coumarins can be classified into 6 types: simple coumarins, furanocoumarins, pyranocoumarins (linear), pyranocoumarins (angular), phenyl coumarins and biscoumarins (Fig. 1B).30
 | ||
| Fig. 1 (A) Structure and numbering of coumarin. (B) Different classes of coumarins. Coumarins and their analogues/derivatives represent an important class of anti-inflammatory agents. Numerous naturally occurring and synthetic coumarins have demonstrated potent anti-inflammatory effects, acting through various mechanisms, such as the inhibition of inducible nitric oxide synthase (iNOS), cyclooxygenase (COX), NF-kB, MAPK, JAK-STAT pathways, as well as the suppression of pro-inflammatory cytokines like TNF-α, NO, PGE2 and IL-6.25–28 | ||
Interestingly, many natural and synthetic coumarins known for their anti-inflammatory activities often possess relatively simple structures, characterized by few functional groups and substituents, and low fraction of sp3 carbons (Fsp3). An example is umbelliferone, i.e. 7-hydroxycoumarin, a simple, naturally occurring coumarin primarily recognized for its antioxidant and anti-inflammatory properties (Fig. 2A).
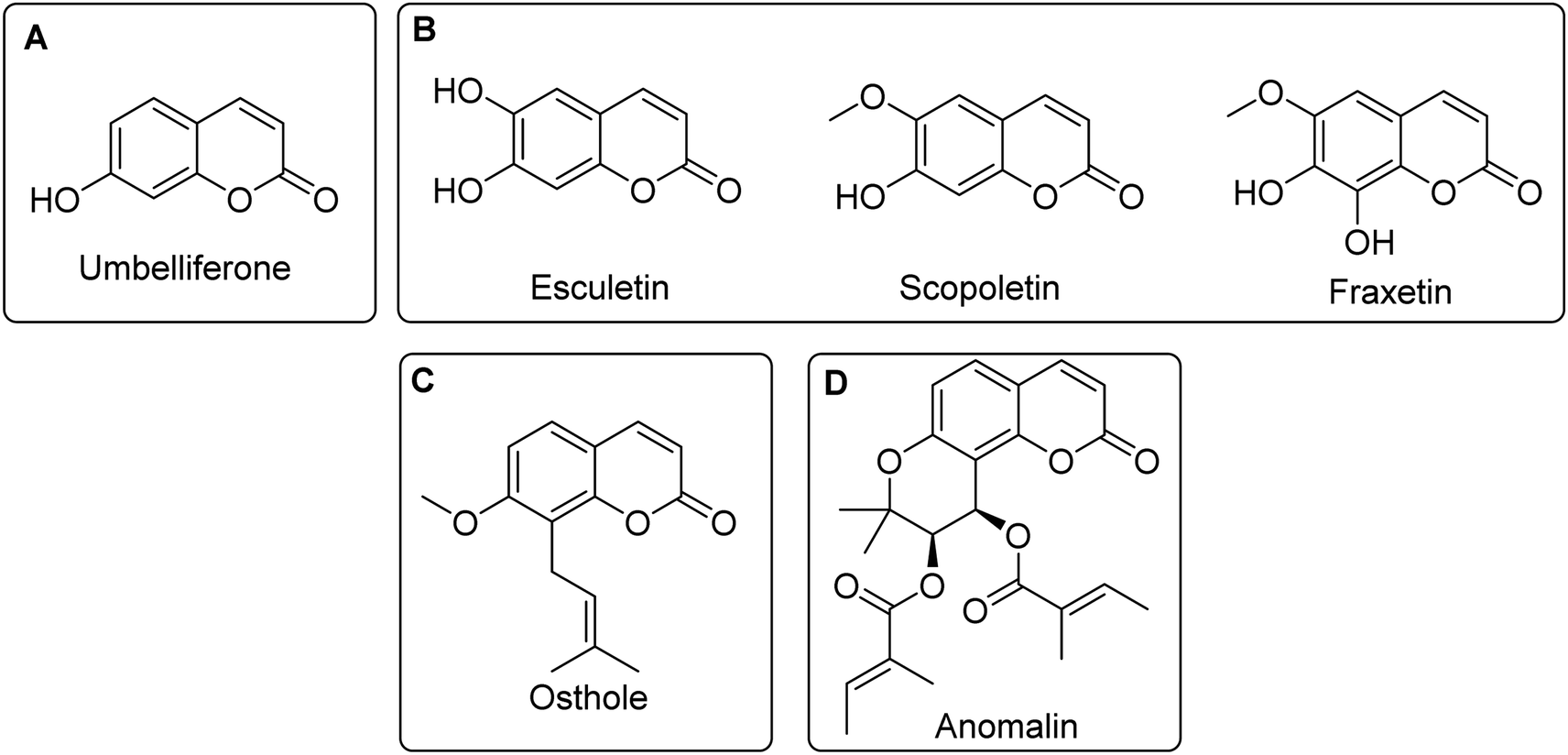 | ||
| Fig. 2 Structures of (A) umbelliferone and (B) the naturally occurring analogues/derivatives of umbelliferone that have been reported to possess similar anti-inflammatory effects, (C) osthole and (D) anomalin. | ||
Umbelliferone has been reported to suppress the production of pro-inflammatory mediators and pathways in several in vivo models. Notably, in a lipopolysaccharide (LPS)-induced acute lung injury (ALI) model, umbelliferone demonstrated significant inhibitory effects on the production of TNF-α, IL-1β, CCL2/monocyte chemoattractant protein-1 (MCP-1) and IL-6 by targeting the TLR4 and NF-κB pathways at doses of 10 to 40 mg kg−1. Umbelliferone also displayed anti-inflammatory effects on Freund's complete adjuvant (FCA)-induced arthritis mouse models via inhibition of the NF-κB and MAPK signalling pathways.31 In another study, oral administration of umbelliferone was found to reduce atopic dermatitis symptoms induced by 2,4-dinitrochlorobenzene (DNCB) and dermatophagoides farinae extract (DFE) in mice, through the suppression of pro-inflammatory cytokines and chemokines. Additionally, in the same study, umbelliferone treatment inhibited the production of pro-inflammatory cytokines in TNF-α/IFN-γ-treated HaCAT cells.32 The mode of anti-inflammatory action was found to be via inhibition of IkBα degradation and NF-kB nuclear translocation, inhibition of signal transducer and activator of transcription 1 (STAT1), p38, extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK) phosphorylation.32 Other anti-inflammatory mechanisms of action for umbelliferone include the inhibition of NLR family pyrin domain containing 3 (NLRP3), nuclear factor erythroid 2-related factor 2 (Nrf2), and Janus kinase-STAT (JAK-STAT) pathways, among others.26 Importantly, structurally simple naturally occurring analogues and derivatives of umbelliferone, such as esculetin, scopoletin, and fraxetin (Fig. 2B), have also demonstrated similar mechanisms for their anti-inflammatory activities.26
Indeed, the umbelliferone core has been used for the further development of many other synthetic coumarin compounds with anti-inflammatory activities. One example is a study by Timonen et al. who synthesized a library of twenty umbelliferone analogues/derivatives using a Pechmann condensation reaction between various substituted resorcinols and the appropriate β-keto esters.33 This led to the identification of a number of compounds with good anti-inflammatory activities in J774 macrophages. The most active compound was able to inhibit iNOS activity by 95% and NO and IL6 production by 92% at 100 µM, compared to umbelliferone (47% iNOS, 65% NO and 34% IL-6 inhibition).33
Aside from umbelliferone, osthole (Fig. 2C) is perhaps the most extensively studied coumarin natural product for its anti-inflammatory properties. Although the precise molecular targets of osthole have not been elucidated, it has been shown to affect a number of inflammatory signalling pathways in several in vitro and in vivo studies. For instance, osthole inhibits the production of key pro-inflammatory mediators, including NO, PGE2, TNF-α, and IL-6 in LPS-stimulated RAW 264.7 macrophages. Furthermore, osthole treatment led to a marked reduction in the protein expression of iNOS, COX-2, p38, and IκBα.26,34–36 In another study, the effects of osthole on inflammatory skin conditions were explored. In histamine/LPS-induced inflammation models using normal human epithelial keratinocytes (NHEK) and normal human dermal fibroblasts (NHDF), osthole significantly attenuated the levels of IL-1β, TNF-α, MCP-1, and CCL5/RANTES.37 Additionally, osthole reduced the protein expression levels of NF-κB, COX-2, and toll-like receptor 2 (TLR2).37 The anti-inflammatory effects of osthole in skin keratinocytes was subsequently found to be due to the direct inhibition of transient receptor potential vanilloid-3 (TRPV3), a member of a non-selective cation channel family.38 Recently, Neuberger et al. solved the cryo-EM structure of osthole complexed with TRPV3, revealing that the oxygen atom of the lactone moiety of osthole forms hydrogen bonds with His 426 (Fig. 3), thus highlighting the critical role of the coumarin scaffold in binding.39 Additionally, the methoxy oxygen of osthole is involved in hydrogen bonding interactions with His 430 (Fig. 3).
 | ||
| Fig. 3 Cryo-EM structure of osthole bound to TRPV3 (PDB: 7RAU). Osthole forms hydrogen bond interactions with His 426 and 430. Crystal structure was visualized using PyMOL 2.5.5. Osthole is represented in yellow whereas interacting amino acid residues are represented in red. Hydrogen bond interactions are represented by yellow dashed lines. | ||
Another relatively more complex coumarin natural product with anti-inflammatory action is anomalin (Fig. 2D). Belonging to the angular pyranocoumarin subclass, anomalin has been evaluated in a number of in vitro and in vivo studies. For instance, anomalin inhibited the transcript and protein expression levels of iNOS and COX-2 in LPS-stimulated RAW 264.7 macrophages.40 qRT-PCR further showed that anomalin significantly reduced the levels of several pro-inflammatory cytokines, including TNF-α and IL-6. In addition, anomalin inhibited the phosphorylation and degradation of IκBα in RAW 264.7 macrophages.40 In another report, Khan et al. studied the effects of anomalin in a mouse model of LPS-induced acute lung injury.41 It was observed that mice that were administered with LPS experienced a significant mortality rate that was markedly improved upon anomalin treatment. Moreover, anomalin decreased the production of NO, IL-1β, IL-6 and TNF-α in plasma and lung tissue.41 To elucidate the mechanisms underlying the anti-inflammatory effects of anomalin, Khan et al. revealed through a western blot analysis that anomalin treatment markedly inhibited the phosphorylation of Akt, p38, JNK and ERK1/2 in LPS-stimulated RAW 264.7 cells.41 Taken together, the anti-inflammatory effects of anomalin could be ascribed to its ability to target multiple signalling pathways like NF-κB, Akt and MAPK.
As illustrated by the examples above, it is clear that coumarins possess significant anti-inflammatory properties. Although only three examples are highlighted here, numerous other anti-inflammatory coumarins have been reported in the literature.26,28,29 Moreover, coumarins are able to inhibit a number of signalling pathways like NF-κB, MAPK, JAK-STAT, iNOS, COX and TLR to exert their anti-inflammatory effects, which contribute to their multi-targeting properties.
2. Polyphenols as a privileged compound class with anti-inflammatory properties
Polyphenols are a structurally diverse group of plant-derived natural products that also belong to the phenylpropanoid family, and are characterized by the presence of one or more aromatic rings bearing multiple hydroxyl groups.42 These compounds can exist in oligomeric or polymeric forms and are often conjugated with sugars, organic acids, plant amines, or other molecules.42 Polyphenols can be divided into two major categories: flavonoids and non-flavonoids. Flavonoids, the largest subclass, are distinguished by their characteristic C6-C3-C6 skeleton (Fig. 4A) and are further categorized into subgroups such as chalcones, flavones, isoflavones, flavonols, flavanones, aurones, and anthocyanins, each defined by specific structural features (Fig. 4B). In contrast, the non-flavonoids comprise of phenolic acids, lignans, stilbenes, xanthones and tannins. | ||
| Fig. 4 Structures of (A) the core flavonoids backbone, (B) different classes of flavonoids and (C) quercetin, a well characterized flavonol known for its anti-inflammatory activities. | ||
Polyphenols have been reported to exhibit a wide range of biological activities such as anti-viral, anti-bacterial, anti-cancer, anti-oxidant and anti-inflammatory etc.43 Amongst their reported biological activities, their anti-inflammatory properties are perhaps one of the most well characterized. A key aspect of their anti-inflammatory effects is their ability to modulate the anti-oxidant pathway (e.g., Kelch-like ECH-associated protein 1 (Keap1)/Nrf2), which plays a pivotal role in inflammation.44 Triggers for oxidative stress include the overproduction of reactive oxygen species (ROS) and other highly reactive molecules that can interact with proteins, lipids, and nucleic acids (DNA/RNA), leading to tissue damage and inflammation.44 Indeed, the crosstalk between the Nrf2 and NF-κB pathways has been well documented.45 Specifically, the activation of Nrf2 can lead to a decrease in NF-κB transcriptional activity, and hence anti-inflammatory effects. Thus, it is evident that by acting as antioxidants and activating the Nrf2 pathway, polyphenols can neutralize ROS and reduce oxidative damage, thereby contributing to their overall anti-inflammatory properties.
Amongst the flavonoids reported with anti-inflammatory activities, quercetin is the most well-studied (Fig. 4C). The anti-inflammatory activity of quercetin can be ascribed to its ability to alleviate oxidative stress and regulate inflammatory modulators.
Quercetin has been reported to reduce oxidative stress in vivo through the activation of Nrf2. In a study by Zardak et al., Wistar rats were subjected to ischemia and reperfusion, leading to oxidative stress, inflammation, and immune system activation.46 Treatment with quercetin resulted in a significant reduction of inflammatory markers such as IL-1β and TNF-α, along with oxidative stress markers including malondialdehyde, superoxide dismutase, and glutathione peroxidase. Further mechanistic studies revealed that administration with quercetin led to an increase in Nrf2 activity, which resulted in repressed NF-kB activities and hence anti-inflammatory effects.46 In another study, the mycotoxin ochratoxin A (OTA) was used to induce oxidative stress in HepG2 cells, leading to an increased production of ROS, activation of NF-κB nuclear translocation and expression, and decreased Nrf2 activity.47 COX-2 protein expression levels were also elevated following OTA stimulation. However, pre-treatment of HepG2 cells with quercetin resulted in increased Nrf2 activity and nuclear translocation, leading to significantly reduced ROS production. Moreover, quercetin prevented NF-κB nuclear translocation and activation leading to a decrease in COX-2 protein expression and OTA-induced NO production.47 Taken together, the anti-inflammatory properties of quercetin can, in part, be ascribed to its anti-oxidant activities.
Quercetin has also been reported to reduce inflammation in a number of in vitro and in vivo models through various mechanisms. These include the suppression of NF-κB and MAPK pathways, as well as inhibition of pro-inflammatory molecules. For example, quercetin has been reported to inhibit NO, TNF-α and IL-6 production in LPS-induced RAW264.7 macrophages through the inhibition of NF-κB and MAPK pathways.48–51In vivo studies using female Lewis rat models found that adjuvant-induced arthritis severity scores were reduced following oral and intracutaneous administration of quercetin.52 This observation was further corroborated by ex vivo data, which showed reduced levels of NO and TNF-α in freshly isolated peritoneal macrophages from the rat model. Quercetin has also been evaluated in randomized clinical trials as an adjuvant therapy for COVID-19.53 These studies demonstrated that patients reported improved clinical symptoms following treatment compared to the control group, with no significant side effects. The proposed mechanism for its efficacy is linked to quercetin's ability to modulate the host inflammatory response following SARS-CoV-2 infection. It was reported that patients in the quercetin-treated group exhibited a significant reduction in lactate dehydrogenase (LDH) in their serum levels.53 As elevated LDH is often associated with tissue damage and inflammation, the reduction in LDH levels suggests that quercetin may help mitigate inflammation and tissue injury that is associated with severe COVID-19 infections.
In addition to quercetin, a recent systematic review analysed 184 naturally occurring, structurally diverse flavonoids reported since 2000 for their anti-inflammatory properties.54 This growing body of evidence supports the notion that the flavonoid scaffold is indeed privileged for anti-inflammatory drug development.
Apart from the flavonoids, other non-flavonoid polyphenols such as resveratrol has also been reported to display anti-inflammatory effects (Fig. 5A).
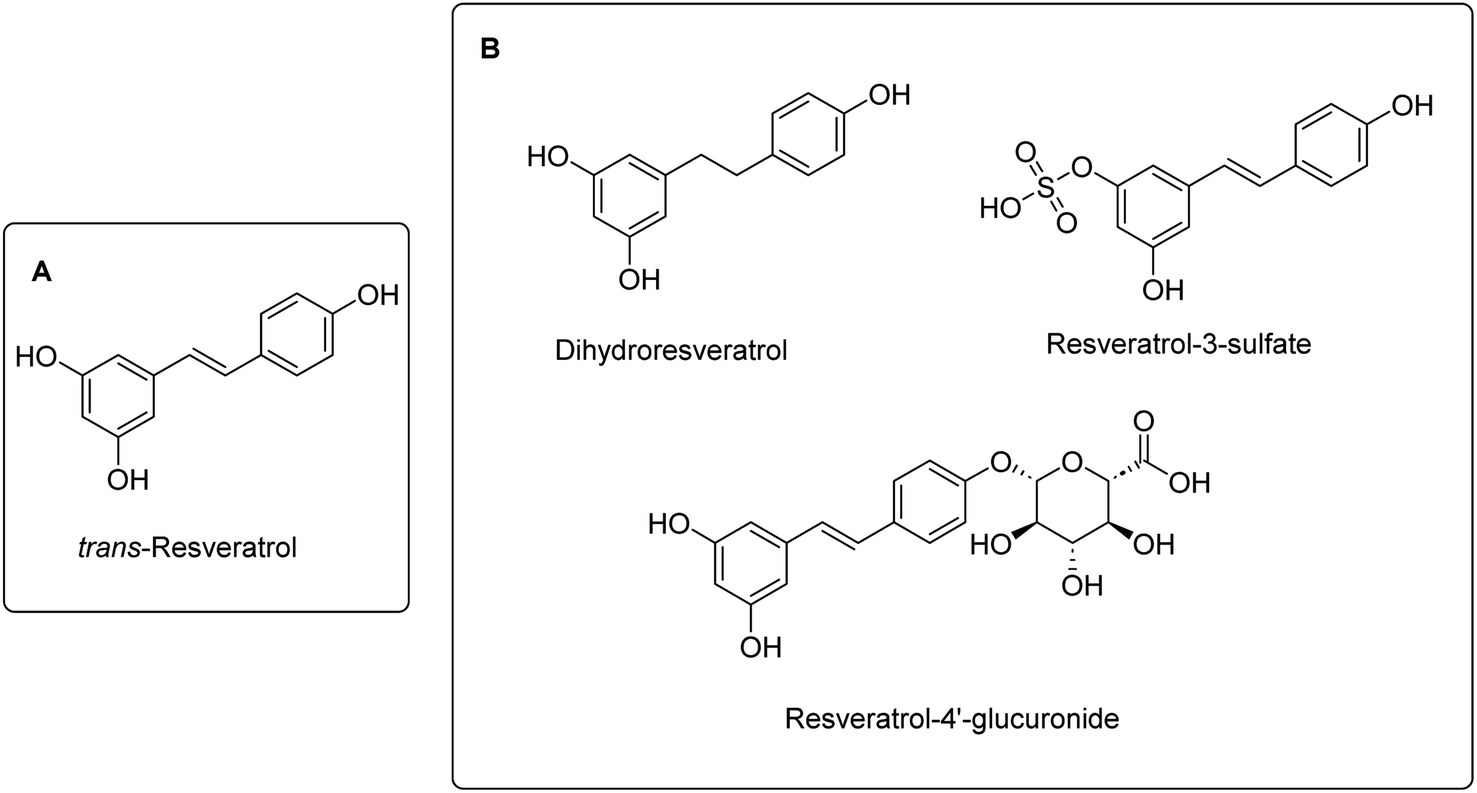 | ||
| Fig. 5 Structures of (A) resveratrol and (B) key metabolites of resveratrol with reported anti-inflammatory activities. | ||
Similar to quercetin, the anti-inflammatory activities of resveratrol can be attributed to its ability to modulate multiple signalling pathways. Ma et al. showed that resveratrol suppressed the release of NO and IL-6 in LPS-stimulated RAW264.7 cells in a concentration-dependent manner.55 In another study, the effects of resveratrol on palmitate-induced inflammation was investigated. It was observed that pretreatment of C2C12 cells with resveratrol significantly alleviated palmitate-induced TNF-α and IL-6 production, as well as their mRNA expression.56 In terms of inflammatory pathways, resveratrol was found to modulate MAPK and NF-κB signalling via the complete inhibition of the phosphorylation of ERK1/2, JNK, and IκB kinases (IKKα/IKKβ) in palmitate-treated cells.56 Additionally, resveratrol reduced the production of pro-inflammatory mediators TNF-α, IL-8, and MCP-1 in LPS-stimulated U937 cells.57 In the same study, RNA sequencing was used to investigate the transcriptional response in U937 cells following resveratrol treatment. Of the 2098 downregulated genes, the top 30 were involved in the positive regulation of inflammatory responses, cell cycle arrest, apoptosis, and transcription.57 Undoubtedly, the literature has established resveratrol as a promising anti-inflammatory agent, and its activities can be ascribed to its ability to modulate key signalling pathways such as NF-κB and MAPK, as well as the suppression of pro-inflammatory cytokines.
Despite the promising anti-inflammatory activities of resveratrol, it suffers from low oral bioavailability (<1%) resulting from extensive metabolism in the liver and intestine.58 However, several studies indicate that the metabolites of resveratrol may also contribute to its anti-inflammatory effects (Fig. 5B). Epidemiological data from the French population indicated a low incidence of coronary heart disease (CHD) – a form of chronic inflammatory disease – despite a diet high in saturated fats.59 This phenomenon is often referred to as the “French Paradox”.60 Subsequent research suggested that this observation could be attributed to the benefits of regular and moderate wine consumption, particularly due to its polyphenol content, including compounds like resveratrol.55 Indeed, the anti-inflammatory effects of the metabolites of resveratrol have been well documented. For example, the metabolite resveratrol-3-sulfate was shown to inhibit the activities of NF-κB, COX-1, and COX-2.61 Lu et al. reported that resveratrol-4′-glucuronide (IC50 = 40.6 µM) and dihydroresveratrol (IC50 = 57.1 µM) exhibited similar COX-2 inhibitory activity as resveratrol (IC50 = 37.5 µM).62 Moreover, dihydroresveratrol was also able to inhibit NO production (IC50 = 64.4 µM) with similar activity to resveratrol (IC50 = 46.1 µM).62 Thus, it is evident that the anti-inflammatory effects of resveratrol can largely be ascribed to its bioactive metabolites, which effectively target multiple inflammatory pathways. Moreover, its demonstrated in vivo efficacy suggests that this natural compound is promising for the further development as an anti-inflammatory agent.
Importantly, novel polyphenols with anti-inflammatory properties continue to emerge as promising leads for anti-inflammatory drug development. For example, Wang et al. isolated two novel polyphenolic compounds, Chinesins B and C from the methanolic extract of Castanopsis chinensis leaves (Fig. 6).63 Both compounds displayed anti-inflammatory activity, inhibiting LPS induced NO production in RAW264.7 cells by approximately 50% at concentrations of 10 µM without significant cytotoxicities.
 | ||
| Fig. 6 Recently isolated natural polyphenols Chinesin B and C with anti-inflammatory activities. | ||
In conclusion, numerous structurally diverse polyphenols have demonstrated anti-inflammatory activities, with some acting through multiple mechanisms. Similar to the coumarins, the ability of polyphenols to modulate various targets and pathways makes them particularly effective for treating complex, multifactorial conditions like inflammation. As highlighted in the examples above, both in vitro and in vivo studies have yielded promising results regarding the use of polyphenols for inflammation. In the case of resveratrol, its anti-inflammatory effects may arise from a combination of the parent compound and its active metabolites.
3. Labdane diterpenoids as a privileged compound class with anti-inflammatory properties
Terpenoids are a large and diverse class of natural products, derived from (C5)n isoprene units, where n is an integer. They are biosynthesized through the mevalonate and methylerythritol phosphate pathways, and can be further classified as hemiterpenes (n = 1), monoterpenes (n = 2), sesquiterpenes (n = 3), diterpenes (n = 4) and so on depending on the number of C5 carbon units that are present. It should however be noted that a number of terpenoids have undergone extensive chemical modifications such as cyclisation, rearrangement and functionalization as well as combination with carbon skeletons derived from other sources (e.g., acetate pathway). There are numerous examples of natural occurring terpenoids with anti-inflammatory properties, although only a few have progressed to clinical trials and/or are marketed drugs. In a recent review of 281 representative natural terpenoids with anti-inflammatory activities, the most common mechanism for anti-inflammatory action was reported to be via attenuation of the expression of iNOS, leading to reduced production of the pro-inflammatory mediator NO.64 Other targets of natural terpenoids reduce the production of other pro-inflammatory mediators such as cytokines (e.g., IL-6), TNF-α, prostaglandins. As outlined in the introduction, the NF-κB signaling pathway ultimately leads to the transcription of a number of genes, including that related to inflammation. As such, any disruptions of the NF-κB pathway can potentially lead to anti-inflammatory effects, for example by reducing the production of cytokines like TNF-α. Artemisinin, an anti-malarial drug is reported to inhibit the DNA binding of the NF-κB complex,65 as is ginkgolides, diterpenes isolated from Gingko biloba leaves.66 In this perspective, two privileged subclasses of terpenoids – the labdane diterpenoids, and the sesquiterpene lactones – where many members have been reported to have anti-inflammatory activities, will be highlighted.Andrographis paniculata, or green chiretta is a well-known herb that is used as traditional medicine in many parts of the world. The major active component of Andrographis paniculata is a labdane diterpenoid known as Andrographolide (Fig. 7). Not surprisingly this natural product has been reported to display polypharmacology and is purported to have anti-cancer, anti-viral, anti-inflammatory properties to name a few.16,67 Of these, the anti-inflammatory properties of andrographolide is the most well-studied and validated, at least in in vitro studies as well as in selected animal models.68,69 Specifically, in vitro studies have shown that the production of TNF-α, IL-6 and NO are inhibited in RAW264.7 cells that has been stimulated with LPS.70 There is also evidence that treatment with andrographolide is also able to reduce inflammation in a rat paw edema assay.71 A number of biological targets identified are purported to be responsible for its anti-inflammatory activities. Of this, through pull-down studies, cysteine-62 of the p50 subunit of NF-κB has been shown to interact covalently with the exocyclic α, β-unsaturated γ-butyrolactone ring of andrographolide.72 This inhibition of NF-κB will then lead to decreased transcription of genes that are responsible for the production of a number of pro-inflammatory cytokines as noted in the earlier sections. Saturating the C12–C13 double bond of andrographolide led to loss of activity. This is consistent with the notion that inhibition of NF-κB occurs via an irreversible covalent reaction with andrographolide. Surprisingly, the analogue of andrographolide that does not possess a C-14 hydroxy group but retain the exocyclic ΔC12–13 bond which can still undergo a Michael reaction with the nucleophilic cysteines (albeit in a reversible fashion), does not possess anti-inflammatory activities as is measured by quantifying the production of pro-inflammatory cytokines.73 It is interesting to note that the concentrations of andrographolide that is needed for 50% inhibition of NF-κB are high and cannot adequately account for the observed anti-inflammatory effects (IC50 (TNF-α) ca. 4 µM), suggesting that there are other major pathways that may be responsible for the anti-inflammatory properties of andrographolide. In a recent paper by Tran et al., a novel mechanism that may contribute to the anti-inflammatory properties of andrographolide was identified.15 In this mechanism, andrographolide is proposed to disrupt the formation of a p38 MAPK-MK2 complex through the binding of andrographolide at an activation loop of MK2, which is located at the interface of p38α and MK2. This disruption causes the p38 MAPK-MK2 complex to disassociate and the MK2 that is released is degraded naturally. As MK2 is responsible for the phosphorylation of RNA binding proteins, e.g. tristetraprolein (TTP), which is in turn responsible for the transcription of genes that are responsible for pro-inflammatory cytokines production, a reduction of MK2 will lead to an overall anti-inflammatory effect.74 Another noteworthy target of andrographolide is the cysteine residue/s of the Keap1 protein. Covalent modification of Keap1 by andrographolide will affect the formation of the Nrf2-Keap1 protein complex, causing the release and translocation of Nrf2, an antioxidant transcription factor, and antioxidant enzymes are produced.75 A related labdane diterpenoid, calcaratarin D has an identical warhead to andrographolide but has a different labdane skeleton (normal-labdane) (Fig. 7). Calcaratarin D displays similar anti-inflammatory effects as andrographolide but intriguingly, and in contrast to andrographolide, inhibits NF-κB strongly.76 Studies suggest that calcaratarin D is likely to exert its anti-inflammatory properties through a selective Akt- NF-κB mediated mechanism.
 | ||
| Fig. 7 Structures of labdane diterpenoids with reported anti-inflammatory activities. Michael-acceptor warheads are highlighted in red. | ||
Apart from andrographolide and calcaratarin D, numerous natural labdane diterpenoids with ent- and normal-labdane stereochemistry have been reported to possess anti-inflammatory properties.77 Although these are not as well-studied as andrographolide, some potential biological targets have been identified. In these labdane diterpenoids, there is variation in the substituents on the labdane ring but more intriguingly is the variety of warheads present (Fig. 7). For example, 14-deoxy-11,12-didehydroandrographolide (DDA), which is also found in Andrographis paniculata has a diene warhead with an endocyclic Michael acceptor, while andalusol, isolated from Sideritis foetens Clemen, has a diene moiety that is not capable of undergoing a Michael addition.78 For DDA, the anti-inflammatory activity is reported to be much weaker than andrographolide and it is believed that the anti-inflammatory properties arise from the inhibition of NF-κB, through modification of the p50 subunit of NF-κB.79 The reported anti-inflammatory activities of andalusol was reported to be ca. 10.5 µM for the inhibition of NO production.80 The mechanism of anti-inflammatory action of andalusol is thought to be due to the suppression of IκBα phosphorylation and degradation, which then leads to the inhibition of the NF-κB pathway. In another example, forsypensins, isolated from the fruits of Forsythia suspensa have also been reported to possess anti-inflammatory properties, comparable to that of ginkgolide.81 Forsypensins are examples of normal-labdanes with ene carboxylic acids as warheads (Fig. 7).
The number of labdane diterpenoids with anti-inflammatory properties is astounding and detailed investigation of possible targets are warranted for the future development of new anti-inflammatory drugs. Issues of selectivity, efficacy, pharmacokinetics and solubilities can potentially be resolved with the identification and understanding of the pharmacophore that is needed for activities as well as through the application of medicinal chemistry in the development of drug-like compounds.
4. Sesquiterpene lactones as a privileged compound class with anti-inflammatory properties
Sesquiterpene lactones (STLs) are a diverse group of naturally occurring compounds predominantly found in plants of the Asteraceae family. These secondary metabolites belong to the family of sesquiterpenoids (C15) and are distinguished by the presence of a lactone ring, which is central to their biological activities. Many STLs bear an electrophilic α-methylene-γ-lactone, which can act as a Michael acceptor to covalently bind to target proteins.82 STLs are widely recognised for their diverse bioactivities, most notably their anti-inflammatory properties.83One of the most well studied STL is parthenolide (Fig. 8A), isolated from the feverfew plant (Tanacetum parthenium).84,85 The plant's name reflects its historical use in traditional medicine for alleviating ailments such as fever, migraines, rheumatoid arthritis, and general pain, amongst others.86–88 The identification of parthenolide as the major bioactive component of feverfew has driven extensive research into its pharmacological properties, with a particular focus on its anti-inflammatory effects.86–90
 | ||
| Fig. 8 Structures of (A) parthenolide and (B) recently reported STLs siegesbeckialide I and glabrescone A. | ||
The first identified and most well-characterized target of parthenolide is IκB kinase β (IKKβ), a subunit of the IκB kinase (IKK) complex that regulates the NF-κB signalling pathway.91 Using a biotinylated parthenolide analogue, followed by several rounds of affinity purification and anti-IKKβ immunoblot analysis, Kwok et al. demonstrated that parthenolide forms a covalent bond with IKKβ in a dose-dependent manner. Site-directed mutagenesis combined with tandem mass spectrometry identified Cys179 as the key residue forming a covalent adduct with parthenolide.91 This covalent inhibition of IKKβ underlies parthenolide's anti-inflammatory effects by suppressing the NF-κB pathway. Notably, parthenolide inhibited LPS-induced IκBα degradation and p65 nuclear translocation in murine vascular smooth muscle cells (VSMCs).92 Interestingly, parthenolide did not affect other common inflammatory pathways like ERK, p38, and JNK in the same cell line. Further in vivo studies demonstrated that parthenolide was able to reduce inflammation in atherosclerotic mice. These findings are aligned with previous reports demonstrating its protective effects against myocardial ischemia and renal diseases like glomerulonephritis via NF-κB inhibition.93,94 In a separate study, parthenolide was shown to abrogate colon inflammation via regulation of gut microbiota.95 Treatment with parthenolide led to a reduction in proinflammatory cytokines such as IL-1β, TNF-α, IL-6, and IL-17A, while simultaneously enhancing the expression of the immunosuppressive cytokine IL-10 in colon tissue. Furthermore, parthenolide treated mice displayed significantly improved gut microbial profile compared to the control group, highlighting its potential as a therapeutic agent for the treatment and/or prevention of inflammatory bowel disease (IBD).95
More recently, the STLs siegesbeckialide I and glabrescone A (Fig. 8B) isolated from Sigesbeckia glabrescens were reported to exhibit anti-inflammatory activity by inhibiting NO release in LPS-stimulated RAW264.7 cells with IC50 values of 1.4 ± 0.2 µM and 15.4 ± 1.3 µM respectively.96 Siegesbeckialide I was also able to inhibit the production of other pro-inflammatory mediators like PGE2, IL-1β, IL-6, and TNF-α. Using a Cellular Thermal Shift Assay (CETSA), the authors further showed that Siegesbeckialide I binds directly to IKKα/β, in a manner that is similar to parthenolide.96 Computational docking studies revealed that Cys46 of IKKβ can form a covalent bond with the α, β-unsaturated lactone moiety of siegesbeckialide I. This then leads to the inactivation of IKKα/β, and consequently abrogation of LPS-stimulated NF-κB activation and thus anti-inflammatory effects.
5. Isoquinoline alkaloids as a privileged compound class with anti-inflammatory properties
Alkaloids are defined as natural products that contain one or more nitrogen atoms in their structure, and can be classified as true alkaloids, protoalkaloids, and pseudoalkaloids. These classes differ based on their biosynthetic origins. Both true alkaloids and protoalkaloids are derived from amino acids, with the key distinction being that the nitrogen atom in true alkaloids is part of a heterocycle, while in protoalkaloids, the nitrogen atom is not. Conversely, pseudoalkaloids are synthesized from the amination or transamination of other biological precursors rather than directly from amino acids.78 Interestingly, alkaloids not only contain carbon, hydrogen, and nitrogen atoms but may also include oxygen or sulfur atoms in their structures. Even rarer, some alkaloids can incorporate elements such as phosphorus, chlorine, and bromine, adding to their structural diversity and potential biological activities.97 Today, alkaloids represent an important class of natural products used to treat a variety of diseases, including cancer, bacterial infections, viruses, inflammation and so on. Notably, two subclasses of alkaloids, isoquinoline alkaloids and indole alkaloids, have been particularly effective in mitigating inflammation.Berberine (Fig. 9A), is a naturally occurring isoquinoline alkaloid found in a family of herbs including Rhizoma coptidis, Berberis vulgaris, Berberis aristata etc.98 Berberine has been demonstrated to exhibit anti-inflammatory activity in both in vitro and in vivo models.
 | ||
| Fig. 9 Structures of (A) berberine and its metabolites of berberine with anti-inflammatory activities. Highlighted portions in red indicate structural differences of the compounds from the parent compound berberine. (B) Recently isolated isoquinoline alkaloid natural products with anti-inflammatory activities. | ||
In an LPS-induced endometritis mouse model, berberine significantly reduced the concentration of NO in uterine tissue and inhibited the expression of pro-inflammatory cytokines TNF-α and IL-1β.99 Western blot analysis of NF-κB and IκBα levels revealed that berberine treatment inhibited NF-κB activation and IκBα degradation, suggesting that the anti-inflammatory effects of berberine may stem from its ability to inhibit the NF-κB signalling pathway.99 Similar results were observed in Aβ-stimulated murine microglial BV2 cells, where berberine demonstrated significant inhibitory effects on the production of Aβ-stimulated inflammatory mediators and cytokines, including IL-6, MCP-1, iNOS, and COX-2. Additionally, western blot analysis showed that berberine inhibited the phosphorylation of IκBα at Ser32/36, preventing its degradation and subsequent activation of NF-κB.100
The anti-inflammatory effects of berberine can also be attributed to its inhibition of phospholipase A2 (PLA2), the enzyme responsible for converting phospholipids into arachidonic acid.101 In a study conducted by Chandra et al., a crystal structure of berberine complexed with PLA2 was solved, revealing that berberine binds to the hydrophobic channel within the active site of PLA2. As illustrated in Fig. 10, one of the methylenedioxy oxygens of berberine forms a hydrogen bond with the amide proton of Gly30, while the same oxygen atom also engages in a water-mediated hydrogen bond with the imidazole ring of His48. Additionally, hydrophobic interactions with residues such as Leu2, Phe5, Ile9, Ala18, Ile19, Tyr22, Ser23, Cys45, His48, Lys69, and Phe106 further stabilize the binding of berberine to PLA2. The inhibitory effects of berberine against PLA2 were corroborated by enzyme kinetics assays, reporting an IC50 value of 87 µM.101
 | ||
| Fig. 10 Crystal structure of berberine-PLA2 complex (PDB: 2QVD). Methylenedioxy oxygen of berberine forms hydrogen bonds with Gly30 and His 48 residues in the hydrophobic channel of PLA2. Crystal structure was visualized using PyMOL 2.5.5. Berberine is represented in yellow whereas interacting amino acid residues are represented in red. Hydrogen bond interactions are represented by yellow dashed lines. Water molecules are represented as red crosses. | ||
Similar to resveratrol, the utility of berberine as a lead compound is limited by poor bioavailability, largely due to extensive phase I and II metabolism. For instance, in rats, a single oral dose of berberine results in a bioavailability of only 0.37%.102 The main metabolites of berberine include berberrubine, demethyleneberberine, jatrorrhizine, palmatine and columbamine (Fig. 9A).103
Considering the low bioavailability of berberine, its in vivo anti-inflammatory activities are rather astonishing. Thus, it is not unreasonable to surmise that its effectiveness in vivo largely stems from its bioactive metabolites. Indeed, the anti-inflammatory activities of the metabolites of berberine (Fig. 9A) have been reported. For example, metabolites like berberrubine and demethyleneberberine have demonstrated anti-inflammatory effects across various cell and animal models, including a human retinal pigment epithelial cell line,104 dextran sulfate sodium (DSS)-induced colitis,105,106 and a P. aeruginosa-induced pneumonia mouse model.107 Palmatine and columbamine were shown to inhibit NF-κB activation and nitric oxide production in LPS-stimulated RAW264.7 macrophages.108 Additionally, jatrorrhizine exhibited NF-κB inhibitory activity in a DSS-induced colitis mouse model.109 Unsurprisingly, the bioactive metabolites of berberine (Fig. 9A) bear a high degree of structural similarity to berberine and thus underscores the privileged nature of the berberine scaffold for anti-inflammatory activity and further development.
Aside from berberine, other isoquinoline alkaloids have been demonstrated to possess good anti-inflammatory activities. For example, the simple isoquinoline alkaloid litcubanine A was recently isolated from L. cubeba in 2021 (Fig. 9B).110In vitro studies showed that litcubanine A significantly inhibited LPS-induced iNOS expression in both RAW 264.7 cells and primary mouse peritoneal macrophages by approximately 85% at concentrations as low as 100 nM. Mechanistic studies further revealed that litcubanine A exerts its anti-inflammatory effects through modulation of the NF-κB signaling pathway. Specifically, treatment with litcubanine A suppressed the phosphorylation of IκK, IκB, and p65 NF-κB in LPS-stimulated RAW 264.7 cells. In a separate study, the novel isoquinoline alkaloid hypectumine B (Fig. 9B), isolated from the herb Hypecoum erectum L. in 2024 was shown to inhibit LPS induced NO production in RAW 264.7 cells with an IC50 value of 24.4 µM. Furthermore, hypectumine B attenuated the mRNA expression levels of TNF-α, IL-6, thus abrogating the TNF-α and IL-6 cytokine release in LPS-induced RAW264.7 cells.
6. Indoles as a privileged compound class with anti-inflammatory properties
Indoles are a class of naturally occurring aromatic heterocyclic compounds belonging to the alkaloid family and are characterized by a six-membered benzene ring fused to a pyrrole. Notably, the indole moiety constitutes the side chain of the canonical amino acid tryptophan, thus underscoring its significance in biological systems.Rutaecarpine is a polycyclic indole alkaloid isolated from the fruits of Evodia rutaecarpa. and is also well known for its anti-inflammatory properties (Fig. 11A). Rutaecarpine exhibits its anti-inflammatory activity via inhibition of the NF-κB signaling pathway.111 When lipoteichoic acid (LTA)-induced RAW267.4 cells were treated with rutaecarpine, a concentration-dependent inhibition of inflammatory mediators and cytokines such as NO, TNF-α, and IL-1β was observed. This anti-inflammatory effect was attributed to inhibition of the NF-κB signaling pathway as shown by western blot studies and confocal microscopy, where rutaecarpine treatment led to significantly lowered nuclear translocation of fluorescently labeled NF-κB.111 Another target of rutaecarpine is COX-2, as shown in studies conducted by Moon et al.112 Rutaecarpine selectively inhibited COX-2 over COX-1 in both bone marrow-derived mast cells (BMMC) and HEK293 cells. In BMMC, it exhibits a 30-fold selectivity for COX-2, inhibiting the enzyme with an IC50 value of 0.28 µM. In HEK293 cells, rutaecarpine has an IC50 value of 2.8 µM, showcasing an impressive selectivity of over 140-fold for COX-2 compared to COX-1.112
 | ||
| Fig. 11 Structures of (A) rutaecarpine, (B) known selective COX-2 inhibitors that were withdrawn from the market by FDA due to adverse cardiovascular effect and (C) newly isolated novel indole natural products with anti-inflammatory activities. Indole moiety is denoted in red. | ||
Interestingly, despite its selective COX-2 inhibition, rutaecarpine does not suffer from the adverse cardiovascular effects associated with NSAIDs such as rofecoxib and valdecoxib (Fig. 11B).113–119 This can be ascribed to the anti-platelet activity of rutaecarpine via the inhibition of phospholipase C. This reduces phosphoinositide breakdown, subsequently decreasing thromboxane A2 formation and intracellular Ca2+ mobilization, ultimately preventing platelet aggregation induced by various agonists.120,121 Notably, this anti-platelet mechanism is distinct from that of aspirin, which operates through the irreversible covalent inhibition of platelet COX-1.118,122 As such, rutaecarpine serves as an excellent starting point for the further development of anti-inflammatory drugs as it does not result in increased cardiovascular complications. Its non-covalent mechanism of action is also advantageous as it reduces the likelihood of off-target binding that can lead to toxicities.
Numerous other indole-based natural products have demonstrated anti-inflammatory activities. For example, two newly identified indole alkaloids, fissindoalkas B and C, were isolated in 2024 from the roots of Fissistigma oldhamii (Hemsl.) Merr.123 These compounds were shown to effectively suppress LPS-induced NO production in RAW 264.7 cells, with IC50 values of 2.52 ± 0.18 µM and 2.33 ± 0.16 µM, respectively (Fig. 11C). In another study, the indole natural product axilline B (Fig. 11C), isolated from the ethanol extract of M. axillaris, exhibited significant anti-inflammatory effects in vitro. Specifically, axilline B abrogated LPS-induced NO release in RAW 264.7 cells, with an IC50 value of 3.7 ± 0.9 µM. Mechanistic studies further demonstrated that axilline B effectively reduced the protein expression levels of iNOS and COX-2 in inflamed cells. Axilline B was also found to modulate NF-κB signalling by inhibiting the phosphorylation of IκBα in LPS-stimulated RAW 264.7 cells, highlighting its potential as an anti-inflammatory agent.
7. Concluding remarks
Natural products have traditionally provided humankind with life-saving therapeutics and are poised to continue to play a critical role in drug discovery and development. This is evidenced from the fact that ca. 40% of all small molecule FDA approved drugs have origins from natural products – a testament to their enduring value and potential.124 Natural products are good sources of lead compounds due to the presence of privileged structures that increase their propensity to bind to one or more biological receptors (i.e., multi-targeting/polypharmacology).90,125 Indeed, the average number of known target proteins for FDA-approved and clinically investigated NPs in the Drug Bank is 13.89.19 In the context of inflammation, the polypharmacology of natural products is advantageous as treating inflammation is a multi-faceted problem, and is likely to require multi-targeting drugs for effective control or cure.126–128 This is illustrated by examples from the privileged compound classes discussed above, which have been widely reported to modulate multiple inflammatory pathways simultaneously.It is well known that natural products exhibit higher success rates in clinical trials during the drug development process.129 For example, the proportion of natural products and natural product-derived compounds in Phase 1 clinical trials are ca. 35% while in Phase III clinical trials, this percentage is ca. 45%. In contrast, for synthetic compounds, the percentages for Phase I and Phase III clinical trials are ca. 65% and ca. 55% respectively, showing a greater amount of attrition as compared to natural products and natural product derived compounds.129 Thus, prioritizing natural products and/or natural product derived compounds can improve the chances of clinical trials success in the drug development pipeline.129 In another study, Heinzke et al. evaluated the natural product likeness of marketed drugs and compounds in Phase I to III clinical trials.130 Their findings revealed an increasing trend in natural product-like properties among a set of over 1000 clinical compounds published since 2008. This can be explained by the rise of clinical pseudo-natural products (PNPs)—compounds created by combining natural product fragments in ways that would not naturally occur through biosynthesis. PNPs have been rapidly increasing over time and now make up 67% of clinical compounds developed after 2010. A possible advantage for PNPs is that compounds with increased natural product-likeness may facilitate drug distribution through membrane transporters.131 Additionally, incorporating natural product fragments into drug candidates may reduce attrition rates that result from toxicities, which is a leading cause of preclinical failure.132 The overall findings support the idea that a form of “natural selection” is at play, with successful drug discovery campaigns increasingly favouring the use of natural product-like structures.130
Despite the advantages of natural products, there are still limitations that need to be addressed to effectively utilise these remarkable small molecules in drug discovery and development. For example, the biological activities of some of the natural products outlined here are not without controversy and have been the subject of much discussion. First and foremost is the knowledge that low to medium molecular weight (less than 500) natural products such as that belonging to the coumarins and the polyphenols family typically have poor bioavailability due to low solubility and rapid metabolism, thus raising concerns about their efficacies and true usefulness as drugs. This was highlighted using resveratrol and berberine as examples. In another example, andrographolide is also rapidly metabolised and has an elimination half-life of 2.39 hours in humans.133 Nonetheless, all three natural products display remarkable in vivo efficacies. As mentioned above, this could be rationalised by the formation of bioactive metabolites. Furthermore, these metabolites can also bind to different biological targets to synergistically/additively elicit their anti-inflammatory effects, a phenomenon known as metabolism-activated multi-targeting (MAMUT).134 That said, not all natural products generate active metabolites, and their therapeutic potential remains limited by low bioavailability and rapid metabolism. Fortunately, this can be addressed by designing pro-drugs and/or with formulation technology.135–137 Taken together, the metabolic liabilities of natural products suggest that demonstrating anti-inflammatory activity is only the first step in the development of effective anti-inflammatory agents. To fully harness their potential, a comprehensive understanding of their pharmacokinetic and pharmacodynamic (PK/PD) properties, identification of bioactive metabolites, and rigorous in vivo analyses are essential.16,134 While this is a challenging and laborious endeavour, it is a pursuit well worth undertaking to exploit the full therapeutic potential of natural products and MAMUT for the treatment of inflammation.
The second issue is that pertaining to privileged compound classes like the polyphenols, which has been labelled as pan-assay interference compounds (PAINS) or invalid metabolic panaceas (IMPS).138–143 This means that the assay conditions should be scrutinised to ensure that the results are real. In our view, a PAINS label does not inherently preclude these compounds from drug development but rather emphasizes that assay conditions should account for potential interferences.144
From the examples in the compound classes above, it is evident that there is no single distinct structural feature in each of the privileged compound class that can be solely ascribed to be responsible for its privileged anti-inflammatory activities. The precise biological targets are dependent on the actual structure of the compounds such that even slightly structurally different natural products may have very different targets. The anti-inflammatory potencies also vary and by and large, small to medium sized natural products are not highly potent. However, it is clear from the many examples in the literature that potencies can be improved through understanding of the mechanism/s of anti-inflammatory action, and this enables rational structural optimisation of the natural products. Additionally, as demonstrated by the aforementioned examples, newly reported active natural products are still being discovered, and this underscores their continuing value as lead compounds for anti-inflammatory drugs. Hence, from a drug discovery and development viewpoint, nature-inspired drug design offers tremendous opportunities for anti-inflammatory drug research.
8. Data availability
No primary research results, software or code have been included and no new data were generated or analysed as part of this review.9. Conflicts of interest
There are no conflicts to declare.10. Acknowledgements
This work was supported by the NUHS Seed Fund (NUHSRO/2024/023/T1/Seed-Sep 23/01) to W. S. F. W., and an MOE Tier 1 Grant A-8001493-00-00 from the Ministry of Education Singapore to C. L. L. C.11. References
- L. Chen, H. Deng, H. Cui, J. Fang, Z. Zuo, J. Deng, Y. Li, X. Wang and L. Zhao, Inflammatory responses and inflammation-associated diseases in organs, Oncotarget, 2017, 9(6), 7204–7218 CrossRef.
- D. Furman, J. Campisi, E. Verdin, P. Carrera-Bastos, S. Targ, C. Franceschi, L. Ferrucci, D. W. Gilroy, A. Fasano, G. W. Miller, A. H. Miller, A. Mantovani, C. M. Weyand, N. Barzilai, J. J. Goronzy, T. A. Rando, R. B. Effros, A. Lucia, N. Kleinstreuer and G. M. Slavich, Chronic inflammation in the etiology of disease across the life span, Nat. Med., 2019, 25(12), 1822–1832 CrossRef.
- M. G. Dorrington and I. D. C. Fraser, NF-κB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration, Front. Immunol., 2019, 10, 705 CrossRef.
- J. Wang, Y. Liu, Y. Guo, C. Liu, Y. Yang, X. Fan, H. Yang, Y. Liu and T. Ma, Function and inhibition of P38 MAP kinase signaling: Targeting multiple inflammation diseases, Biochem. Pharmacol., 2024, 220, 115973 CrossRef.
- K. Jin, C. Qian, J. Lin and B. Liu, Cyclooxygenase-2-Prostaglandin E2 pathway: A key player in tumor-associated immune cells, Front. Oncol., 2023, 13, 1099811 CrossRef.
- A. R. Liu and P. Ramakrishnan, Regulation of Nuclear Factor-kappaB Function by O-GlcNAcylation in Inflammation and Cancer, Front. Cell Dev. Biol., 2021, 9, 751761 CrossRef.
- B. M. Facchin, G. O. Dos Reis, G. N. Vieira, E. T. B. Mohr, J. S. da Rosa, I. F. Kretzer, I. G. Demarchi and E. M. Dalmarco, Inflammatory biomarkers on an LPS-induced RAW 264.7 cell model: a systematic review and meta-analysis, Inflamm. Res., 2022, 71(7–8), 741–758 Search PubMed.
- G. van Loo and M. J. M. Bertrand, Death by TNF: a road to inflammation, Nat. Rev. Immunol., 2023, 23(5), 289–303 Search PubMed.
- M. Nakanishi and D. W. Rosenberg, Multifaceted roles of PGE2 in inflammation and cancer, Semin. Immunopathol., 2013, 35(2), 123–137 CrossRef.
- A. Oeckinghaus, M. S. Hayden and S. Ghosh, Crosstalk in NF-κB signaling pathways, Nat. Immunol., 2011, 12(8), 695–708 CrossRef PubMed.
- J. Lei, Y. Zhou, D. Xie and Y. Zhang, Mechanistic insights into a classic wonder drug--aspirin, J. Am. Chem. Soc., 2015, 137(1), 70–73 CrossRef.
- M. J. R. Desborough and D. M. Keeling, The aspirin story – from willow to wonder drug, Br. J. Haematol., 2017, 177(5), 674–683 CrossRef PubMed.
- Y. Peng, M. Ao, B. Dong, Y. Jiang, L. Yu, Z. Chen, C. Hu and R. Xu, Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures, Drug Des., Dev. Ther., 2021, 15, 4503–4525 CrossRef.
- X. Ai, P. Yu, L. Peng, L. Luo, J. Liu, S. Li, X. Lai, F. Luan and X. Meng, Berberine: A Review of its Pharmacokinetics Properties and Therapeutic Potentials in Diverse Vascular Diseases, Front. Pharmacol, 2021, 12, 762654 CrossRef CAS PubMed.
- Q. T. N. Tran, P. X. L. Gan, W. Liao, Y. K. Mok, C. L. L. Chai and W. S. F. Wong, Degradation of MK2 with natural compound andrographolide: A new modality for anti-inflammatory therapy, Pharmacol. Res., 2023, 194, 106861 CrossRef CAS PubMed.
- Q. T. N. Tran, W. S. D. Tan, W. S. F. Wong and C. L. L. Chai, Polypharmacology of andrographolide: beyond one molecule one target, Nat. Prod. Rep., 2021, 38(4), 682–692 RSC.
- R. A. Burgos, P. Alarcon, J. Quiroga, C. Manosalva and J. Hancke, Andrographolide, an anti-inflammatory multitarget drug: all roads lead to cellular metabolism, Molecules, 2021, 26(1), 5 Search PubMed.
- B. E. Evans, K. E. Rittle, M. G. Bock, R. M. DiPardo, R. M. Freidinger, W. L. Whitter, G. F. Lundell, D. F. Veber, P. S. Anderson and R. S. Chang, et al., Methods for drug discovery: development of potent, selective, orally effective cholecystokinin antagonists, J. Med. Chem., 1988, 31(12), 2235–2246 CrossRef CAS.
- J. Fang, C. Cai, Q. Wang, P. Lin, Z. Zhao and F. Cheng, Systems Pharmacology-Based Discovery of Natural Products for Precision Oncology Through Targeting Cancer Mutated Genes, CPT Pharmacometrics Syst. Pharmacol., 2017, 6(3), 177–187 CrossRef CAS.
- P. Schneider and G. Schneider, Privileged Structures Revisited, Angew Chem. Int. Ed. Engl., 2017, 56(27), 7971–7974 CrossRef CAS PubMed.
- M. E. Welsch, S. A. Snyder and B. R. Stockwell, Privileged scaffolds for library design and drug discovery, Curr. Opin. Chem. Biol., 2010, 14(3), 347–361 CrossRef CAS PubMed.
- E. K. Davison and M. A. Brimble, Natural product derived privileged scaffolds in drug discovery, Curr. Opin. Chem. Biol., 2019, 52, 1–8 CrossRef CAS PubMed.
- J. Barros, J. C. Serrani-Yarce, F. Chen, D. Baxter, B. J. Venables and R. A. Dixon, Role of bifunctional ammonia-lyase in grass cell wall biosynthesis, Nat. Plants, 2016, 2(6), 16050 CrossRef CAS.
- T. Vogt, Phenylpropanoid biosynthesis, Mol. Plant, 2010, 3(1), 2–20 CrossRef CAS PubMed.
- M. Z. Y. Choo, L. W. T. Khaw and C. L. L. Chai, Syntheses of Minutuminolate and Related Coumarin Natural Products and Evaluation of Their TNF-α Inhibitory Activities, ACS Omega, 2023, 8, 41785–41791 CrossRef CAS.
- B. Rostom, R. Karaky, I. Kassab and M. Sylla-Iyarreta Veitía, Coumarins derivatives and inflammation: Review of their effects on the inflammatory signaling pathways, Eur. J. Pharmacol., 2022, 922, 174867 Search PubMed.
- A. Stefanachi, F. Leonetti, L. Pisani, M. Catto and A. Carotti, Coumarin: A Natural, Privileged and Versatile Scaffold for Bioactive Compounds, Molecules, 2018, 23(2), 250 Search PubMed.
- J. Grover and S. M. Jachak, Coumarins as privileged scaffold for anti-inflammatory drug development, RSC Adv., 2015, 5(49), 38892–38905 Search PubMed.
- M. K. Hussain, S. Khatoon, M. F. Khan, M. S. Akhtar, S. Ahamad and M. Saquib, Coumarins as versatile therapeutic phytomolecules: A systematic review, Phytomedicine, 2024, 134, 155972 CrossRef CAS PubMed.
- J. Sharifi-Rad, N. Cruz-Martins, P. López-Jornet, E. P. Lopez, N. Harun, B. Yeskaliyeva, A. Beyatli, O. Sytar, S. Shaheen, F. Sharopov, Y. Taheri, A. O. Docea, D. Calina and W. C. Cho, Natural Coumarins: Exploring the Pharmacological Complexity and Underlying Molecular Mechanisms, Oxid. Med. Cell. Longev., 2021, 2021, 6492346 CrossRef.
- L. Ouyang, Y. Dan, Z. Shao, S. Yang, C. Yang, G. Liu, W. Zhou and D. Duan, Effect of umbelliferone on adjuvant-induced arthritis in rats by MAPK/NF-κB pathway, Drug Des., Dev. Ther., 2019, 13, 1163–1170 CrossRef CAS.
- J. Y. Lim, J. H. Lee, D. H. Lee, J. H. Lee and D. K. Kim, Umbelliferone reduces the expression of inflammatory chemokines in HaCaT cells and DNCB/DFE-induced atopic dermatitis symptoms in mice, Int. Immunopharmacol., 2019, 75, 105830 CrossRef CAS PubMed.
- J. M. Timonen, R. M. Nieminen, O. Sareila, A. Goulas, L. J. Moilanen, M. Haukka, P. Vainiotalo, E. Moilanen and P. H. Aulaskari, Synthesis and anti-inflammatory effects of a series of novel 7-hydroxycoumarin derivatives, Eur. J. Med. Chem., 2011, 46(9), 3845–3850 CrossRef CAS PubMed.
- M. Sun, M. Sun and J. Zhang, Osthole: an overview of its sources, biological activities, and modification development, Med. Chem. Res., 2021, 30(10), 1767–1794 CrossRef CAS.
- H. Fan, Z. Gao, K. Ji, X. Li, J. Wu, Y. Liu, X. Wang, H. Liang, Y. Liu, X. Li, P. Liu, D. Chen and F. Zhao, The in vitro and in vivo anti-inflammatory effect of osthole, the major natural coumarin from Cnidium monnieri (L.) Cuss, via the blocking of the activation of the NF-κB and MAPK/p38 pathways, Phytomedicine, 2019, 58, 152864 CrossRef CAS.
- Y. Zhou, Z. Du, Q. Wu, M. Guo, Z. Chen, C. Sun, X. Li, Y. Zou, Z. Zheng, P. Chen, W. J. Cho, Y. C. Cho, N. Chattipakorn, Y. Wang, G. Liang and Q. Tang, Discovery of novel osthole derivatives exerting anti-inflammatory effect on DSS-induced ulcerative colitis and LPS-induced acute lung injury in mice, Eur. J. Med. Chem., 2024, 268, 116252 CrossRef CAS PubMed.
- N. Kordulewska, J. Topa, A. Cieślińska and B. Jarmołowska, Osthole Regulates Secretion of Pro-Inflammatory Cytokines and Expression of TLR2 and NF-κB in Normal Human Keratinocytes and Fibroblasts, J. Inflamm. Res., 2022, 15, 1501–1519 Search PubMed.
- X. Y. Sun, L. L. Sun, H. Qi, Q. Gao, G. X. Wang, N. N. Wei and K. Wang, Antipruritic Effect of Natural Coumarin Osthole through Selective Inhibition of Thermosensitive TRPV3 Channel in the Skin, Mol. Pharmacol., 2018, 94(4), 1164–1173 CrossRef CAS PubMed.
- A. Neuberger, K. D. Nadezhdin, E. Zakharian and A. I. Sobolevsky, Structural mechanism of TRPV3 channel inhibition by the plant-derived coumarin osthole, EMBO Rep., 2021, 22(11), e53233 CrossRef CAS.
- S. Khan, E. M. Shin, R. J. Choi, Y. H. Jung, J. Kim, A. Tosun and Y. S. Kim, Suppression of LPS-induced inflammatory and NF-κB responses by anomalin in RAW 264.7 macrophages, J. Cell. Biochem., 2011, 112(8), 2179–2188 CrossRef CAS PubMed.
- A. Khan, S. Khan, H. Ali, K. U. Shah, H. Ali, O. Shehzad, A. Onder and Y. S. Kim, Anomalin attenuates LPS-induced acute lungs injury through inhibition of AP-1 signaling, Int. Immunopharmacol., 2019, 73, 451–460 CrossRef CAS PubMed.
- N. V. Zagoskina, M. Y. Zubova, T. L. Nechaeva, V. V. Kazantseva, E. A. Goncharuk, V. M. Katanskaya, E. N. Baranova and M. A. Aksenova, Polyphenols in Plants: Structure, Biosynthesis, Abiotic Stress Regulation, and Practical Applications (Review), Int. J. Mol. Sci., 2023, 24(18), 13874 CrossRef CAS PubMed.
- K. Stromsnes, R. Lagzdina, G. Olaso-Gonzalez, L. Gimeno-Mallench and J. Gambini, Pharmacological Properties of Polyphenols: Bioavailability, Mechanisms of Action, and Biological Effects in In Vitro Studies, Animal Models, and Humans, Biomedicines, 2021, 9(8), 1074 CrossRef PubMed.
- Y. Zhou, Z. Jiang, H. Lu, Z. Xu, R. Tong, J. Shi and G. Jia, Recent Advances of Natural Polyphenols Activators for Keap1-Nrf2 Signaling Pathway, Chem. Biodivers., 2019, 16(11), e1900400 CrossRef PubMed.
- S. Saha, B. Buttari, E. Panieri, E. Profumo and L. Saso, An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation, Molecules, 2020, 25(22), 5474 CrossRef PubMed.
- M. Yousefi Zardak, F. Keshavarz, A. Mahyaei, M. Gholami, F. S. Moosavi, E. Abbasloo, F. Abdollahi, M. Hossein Rezaei, E. Madadizadeh, N. Soltani, F. Bejeshk, N. Salehi, F. Rostamabadi, F. Bagheri, M. Jafaraghae, M. Ranjbar Zeydabadi, M. Baghgoli, G. Sepehri and M. A. Bejeshk, Quercetin as a therapeutic agent activate the Nrf2/Keap1 pathway to alleviate lung ischemia-reperfusion injury, Sci. Rep., 2024, 14(1), 23074 CrossRef PubMed.
- P. Ramyaa, R. Krishnaswamy and V. V. Padma, Quercetin modulates OTA-induced oxidative stress and redox signalling in HepG2 cells - up regulation of Nrf2 expression and down regulation of NF-κB and COX-2, Biochim. Biophys. Acta, 2014, 1840(1), 681–692 CrossRef PubMed.
- G. B. Lee, Y. Kim, K. E. Lee, R. Vinayagam, M. Singh and S. G. Kang, Anti-Inflammatory Effects of Quercetin, Rutin, and Troxerutin Result From the Inhibition of NO Production and the Reduction of COX-2 Levels in RAW 264.7 Cells Treated with LPS, Appl. Biochem. Biotechnol., 2024, 196, 8431–8452 CrossRef PubMed.
- R. Zhong, L. Miao, H. Zhang, L. Tan, Y. Zhao, Y. Tu, M. Angel Prieto, J. Simal-Gandara, L. Chen, C. He and H. Cao, Anti-inflammatory activity of flavonols via inhibiting MAPK and NF-κB signaling pathways in RAW264.7 macrophages, Curr. Res. Food Sci., 2022, 5, 1176–1184 CrossRef PubMed.
- J. Tang, P. Diao, X. Shu, L. Li and L. Xiong, Quercetin and Quercitrin Attenuates the Inflammatory Response and Oxidative Stress in LPS-Induced RAW264.7 Cells: In Vitro Assessment and a Theoretical Model, BioMed Res. Int., 2019, 2019, 7039802 Search PubMed.
- H. N. Lee, S. A. Shin, G. S. Choo, H. J. Kim, Y. S. Park, B. S. Kim, S. K. Kim, S. D. Cho, J. S. Nam, C. S. Choi, J. H. Che, B. K. Park and J. Y. Jung, Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264.7 macrophages and DNCB-induced atopic dermatitis animal models, Int. J. Mol. Med., 2018, 41(2), 888–898 Search PubMed.
- M. Mamani-Matsuda, T. Kauss, A. Al-Kharrat, J. Rambert, F. Fawaz, D. Thiolat, D. Moynet, S. Coves, D. Malvy and M. D. Mossalayi, Therapeutic and preventive properties of quercetin in experimental arthritis correlate with decreased macrophage inflammatory mediators, Biochem. Pharmacol., 2006, 72(10), 1304–1310 CrossRef PubMed.
- F. Di Pierro, A. Khan, S. Iqtadar, S. U. Mumtaz, M. N. A. Chaudhry, A. Bertuccioli, G. Derosa, P. Maffioli, S. Togni, A. Riva, P. Allegrini, M. Recchia and N. Zerbinati, Quercetin as a possible complementary agent for early-stage COVID-19: Concluding results of a randomized clinical trial, Front. Pharmacol, 2022, 13, 1096853 CrossRef PubMed.
- J. Li, R. Zhao, P. Miao, F. Xu, J. Chen, X. Jiang, Z. Hui, L. Wang and R. Bai, Discovery of anti-inflammatory natural flavonoids: Diverse scaffolds and promising leads for drug discovery, Eur. J. Med. Chem., 2023, 260, 115791 CrossRef PubMed.
- C. Ma, Y. Wang, L. Dong, M. Li and W. Cai, Anti-inflammatory effect of resveratrol through the suppression of NF-κB and JAK/STAT signaling pathways, Acta Biochim. Biophys. Sin., 2015, 47(3), 207–213 CrossRef PubMed.
- A. Sadeghi, S. S. Seyyed Ebrahimi, A. Golestani and R. Meshkani, Resveratrol Ameliorates Palmitate-Induced Inflammation in Skeletal Muscle Cells by Attenuating Oxidative Stress and JNK/NF-κB Pathway in a SIRT1-Independent Mechanism, J. Cell. Biochem., 2017, 118(9), 2654–2663 CrossRef PubMed.
- D. M. L. Pinheiro, A. H. S. de Oliveira, L. G. Coutinho, F. L. Fontes, R. K. de Medeiros Oliveira, T. T. Oliveira, A. L. F. Faustino, V. Lira da Silva, J. T. A. de Melo Campos, T. B. P. Lajus, S. J. de Souza and L. F. Agnez-Lima, Resveratrol decreases the expression of genes involved in inflammation through transcriptional regulation, Free Radic. Biol. Med., 2019, 130, 8–22 CrossRef PubMed.
- C. Sergides, M. Chirilă, L. Silvestro, D. Pitta and A. Pittas, Bioavailability and safety study of resveratrol 500 mg tablets in healthy male and female volunteers, Exp. Ther. Med., 2016, 11(1), 164–170 CrossRef PubMed.
- B. Catalgol, S. Batirel, Y. Taga and N. K. Ozer, Resveratrol: French paradox revisited, Front. Pharmacol, 2012, 3, 141 Search PubMed.
- S. Renaud and M. de Lorgeril, Wine, alcohol, platelets, and the French paradox for coronary heart disease, Lancet, 1992, 339(8808), 1523–1526 CrossRef PubMed.
- J. Hoshino, E. J. Park, T. P. Kondratyuk, L. Marler, J. M. Pezzuto, R. B. van Breemen, S. Mo, Y. Li and M. Cushman, Selective synthesis and biological evaluation of sulfate-conjugated resveratrol metabolites, J. Med. Chem., 2010, 53(13), 5033–5043 CrossRef PubMed.
- D. L. Lu, D. J. Ding, W. J. Yan, R. R. Li, F. Dai, Q. Wang, S. S. Yu, Y. Li, X. L. Jin and B. Zhou, Influence of glucuronidation and reduction modifications of resveratrol on its biological activities, Chembiochem, 2013, 14(9), 1094–1104 CrossRef PubMed.
- Y. F. Wang, P. Lin, N. Pang, Z. B. Liu, B. Y. Yang, R. J. He and Y. L. Huang, Two novel butyrolactone lignan dimers from leaves of Castanopsis chinensis and their anti-inflammatory activities, J. Nat. Med., 2025, 79(1), 258–267 CrossRef PubMed.
- J. Ge, Z. Liu, Z. Zhong, L. Wang, X. Zhuo, J. Li, X. Jiang, X. Y. Ye, T. Xie and R. Bai, Natural terpenoids with anti-inflammatory activities: Potential leads for anti-inflammatory drug discovery, Bioorg. Chem., 2022, 124, 105817 Search PubMed.
- K. S. Wang, J. Li, Z. Wang, C. Mi, J. Ma, L. X. Piao, G. H. Xu, X. Li and X. Jin, Artemisinin inhibits inflammatory response via regulating NF-κB and MAPK signaling pathways, Immunopharmacol. Immunotoxicol., 2017, 39(1), 28–36 Search PubMed.
- C. W. Woo, F. Cheung, V. W. Chan, Y. L. Siow and K. O, Homocysteine stimulates inducible nitric oxide synthase expression in macrophages: antagonizing effect of ginkgolides and bilobalide, Mol. Cell. Biochem., 2003, 243(1–2), 37–47 Search PubMed.
- H. Zhang, S. Li, Y. Si and H. Xu, Andrographolide and its derivatives: Current achievements and future perspectives, Eur. J. Med. Chem., 2021, 224, 113710 Search PubMed.
- X. Li, W. Yuan, J. Wu, J. Zhen, Q. Sun and M. Yu, Andrographolide, a natural anti-inflammatory agent: An Update, Front. Pharmacol, 2022, 13, 920435 CrossRef PubMed.
- M. Low, H. Suresh, X. Zhou, D. J. Bhuyan, M. A. Alsherbiny, C. Khoo, G. Münch and C. G. Li, The wide spectrum anti-inflammatory activity of andrographolide in comparison to NSAIDs: A promising therapeutic compound against the cytokine storm, PLoS One, 2024, 19(7), e0299965 CrossRef PubMed.
- A. A. Abu-Ghefreh, H. Canatan and C. I. Ezeamuzie, In vitro and in vivo anti-inflammatory effects of andrographolide, Int. Immunopharmacol., 2009, 9(3), 313–318 CrossRef PubMed.
- L. Wen, N. Xia, X. Chen, Y. Li, Y. Hong, Y. Liu, Z. Wang and Y. Liu, Activity of antibacterial, antiviral, anti-inflammatory in compounds andrographolide salt, Eur. J. Pharmacol., 2014, 740, 421–427 CrossRef CAS PubMed.
- Y. F. Xia, B. Q. Ye, Y. D. Li, J. G. Wang, X. J. He, X. Lin, X. Yao, D. Ma, A. Slungaard, R. P. Hebbel, N. S. Key and J. G. Geng, Andrographolide attenuates inflammation by inhibition of NF-kappa B activation through covalent modification of reduced cysteine 62 of p50, J. Immunol., 2004, 173(6), 4207–4217 CrossRef CAS.
- Q. T. N. Tran, D. W. S. Tan, W. S. F. Wong and C. L. L. Chai, From irreversible to reversible covalent inhibitors: Harnessing the andrographolide scaffold for anti-inflammatory action, Eur. J. Med. Chem., 2020, 204, 112481 CrossRef CAS.
- S. Duraisamy, M. Bajpai, U. Bughani, S. G. Dastidar, A. Ray and P. Chopra, MK2: a novel molecular target for anti-inflammatory therapy, Expert Opin. Ther. Targets, 2008, 12(8), 921–936 CrossRef CAS.
- J. Y. Seo, E. Pyo, J. P. An, J. Kim, S. H. Sung and W. K. Oh, Andrographolide Activates Keap1/Nrf2/ARE/HO-1 Pathway in HT22 Cells and Suppresses Microglial Activation by Aβ(42) through Nrf2-Related Inflammatory Response, Mediators Inflammation, 2017, 2017, 5906189 Search PubMed.
- Q. T. N. Tran, W. S. F. Wong and C. L. L. Chai, The identification of naturally occurring labdane diterpenoid calcaratarin D as a potential anti-inflammatory agent, Eur. J. Med. Chem., 2019, 174, 33–44 CrossRef CAS PubMed.
- Q. T. N. Tran, W. S. F. Wong and C. L. L. Chai, Labdane diterpenoids as potential anti-inflammatory agents, Pharmacol. Res., 2017, 124, 43–63 CrossRef CAS.
- A. Navarro, B. de las Heras and A. M. Villar, Andalusol, a diterpenoid with anti-inflammatory activity from Siderits foetens Clemen, Z. Naturforsch., C: J. Biosci., 1997, 52(11–12), 844–849 CrossRef CAS PubMed.
- V. S. Nguyen, X. Y. Loh, H. Wijaya, J. Wang, Q. Lin, Y. Lam, W. S. Wong and Y. K. Mok, Specificity and inhibitory mechanism of andrographolide and its analogues as antiasthma agents on NF-κB p50, J. Nat. Prod., 2015, 78(2), 208–217 CrossRef CAS.
- B. de las Heras, A. Navarro, M. J. Díaz-Guerra, P. Bermejo, A. Castrillo, L. Boscá and A. Villar, Inhibition of NOS-2 expression in macrophages through the inactivation of NF-kappaB by andalusol, Br. J. Pharmacol., 1999, 128(3), 605–612 CrossRef CAS.
- K. L. Xiang, R. X. Liu, L. Zhao, Z. P. Xie, S. M. Zhang and S. J. Dai, Labdane diterpenoids from Forsythia suspensa with anti-inflammatory and anti-viral activities, Phytochemistry, 2020, 173, 112298 CrossRef CAS PubMed.
- R. Lagoutte and N. Winssinger, Following the Lead from Nature with Covalent Inhibitors, Chimia, 2017, 71(10), 703–711 CrossRef CAS.
- M. S. Matos, J. D. Anastácio and C. Nunes Dos Santos, Sesquiterpene Lactones: Promising Natural Compounds to Fight Inflammation, Pharmaceutics, 2021, 13(7), 991 CrossRef CAS.
- A. Ghantous, A. Sinjab, Z. Herceg and N. Darwiche, Parthenolide: from plant shoots to cancer roots, Drug Discov. Today, 2013, 18(17–18), 894–905 CrossRef CAS PubMed.
- A. Pareek, M. Suthar, G. S. Rathore and V. Bansal, Feverfew (Tanacetum parthenium L.): A systematic review, Pharmacogn. Rev., 2011, 5(9), 103–110 CrossRef PubMed.
- J. Liu, M. Cui, Y. Wang and J. Wang, Trends in parthenolide research over the past two decades: A bibliometric analysis, Heliyon, 2023, 9(7), e17843 CrossRef.
- R. R. A. Freund, P. Gobrecht, D. Fischer and H.-D. Arndt, Advances in chemistry and bioactivity of parthenolide, Nat. Prod. Rep., 2020, 37(4), 541–565 RSC.
- S. Dey, M. Sarkar and B. Giri, Anti-inflammatory and Anti-tumor Activities of Parthenolide: An Update, J. Chem. Biol. Ther., 2016, 01, 107 Search PubMed.
- V. B. Mathema, Y. S. Koh, B. C. Thakuri and M. Sillanpää, Parthenolide, a sesquiterpene lactone, expresses multiple anti-cancer and anti-inflammatory activities, Inflammation, 2012, 35(2), 560–565 CrossRef.
- M. Z. Y. Choo and C. L. L. Chai, Chapter Two - The polypharmacology of natural products in drug discovery and development, in Annu. Rep. Med. Chem., ed. Altmann, K.-H., Academic Press, 2023, vol. 61, pp. 55–100 Search PubMed.
- B. H. Kwok, B. Koh, M. I. Ndubuisi, M. Elofsson and C. M. Crews, The anti-inflammatory natural product parthenolide from the medicinal herb Feverfew directly binds to and inhibits IkappaB kinase, Chem. Biol., 2001, 8(8), 759–766 Search PubMed.
- O. López-Franco, P. Hernández-Vargas, G. Ortiz-Muñoz, G. Sanjuán, Y. Suzuki, L. Ortega, J. Blanco, J. Egido and C. Gómez-Guerrero, Parthenolide modulates the NF-kappaB-mediated inflammatory responses in experimental atherosclerosis, Arterioscler. Thromb. Vasc. Biol., 2006, 26(8), 1864–1870 CrossRef.
- B. Zingarelli, P. W. Hake, A. Denenberg and H. R. Wong, Sesquiterpene lactone parthenolide, an inhibitor of IkappaB kinase complex and nuclear factor-kappaB, exerts beneficial effects in myocardial reperfusion injury, Shock, 2002, 17(2), 127–134 CrossRef PubMed.
- O. López-Franco, Y. Suzuki, G. Sanjuán, J. Blanco, P. Hernández-Vargas, Y. Yo, J. Kopp, J. Egido and C. Gómez-Guerrero, Nuclear factor-kappa B inhibitors as potential novel anti-inflammatory agents for the treatment of immune glomerulonephritis, Am. J. Pathol., 2002, 161(4), 1497–1505 Search PubMed.
- Y. J. Liu, B. Tang, F. C. Wang, L. Tang, Y. Y. Lei, Y. Luo, S. J. Huang, M. Yang, L. Y. Wu, W. Wang, S. Liu, S. M. Yang and X. Y. Zhao, Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner, Theranostics, 2020, 10(12), 5225–5241 Search PubMed.
- X. Gao, X. Shen, Y. Zheng, L. Yang, X. Zhang, G. Hu, J. Jia and A. Wang, Sesquiterpene Lactones from Sigesbeckia glabrescens Possessing Potent Anti-inflammatory Activity by Directly Binding to IKKα/β, J. Nat. Prod., 2021, 84(11), 2808–2821 CrossRef.
- P. Dey, A. Kundu, A. Kumar, M. Gupta, B. M. Lee, T. Bhakta, S. Dash and H. S. Kim, Chapter 15 – Analysis of alkaloids (indole alkaloids, isoquinoline alkaloids, tropane alkaloids), in Recent Adv. Nat. Prod. Anal., 2020, pp 505–567 Search PubMed.
- M. A. Neag, A. Mocan, J. Echeverría, R. M. Pop, C. I. Bocsan, G. Crişan and A. D. Buzoianu, Berberine: Botanical Occurrence, Traditional Uses, Extraction Methods, and Relevance in Cardiovascular, Metabolic, Hepatic, and Renal Disorders, Front. Pharmacol, 2018, 9, 557 CrossRef PubMed.
- K. Fu, X. Lv, W. Li, Y. Wang, H. Li, W. Tian and R. Cao, Berberine hydrochloride attenuates lipopolysaccharide-induced endometritis in mice by suppressing activation of NF-κB signal pathway, Int. Immunopharmacol., 2015, 24(1), 128–132 CrossRef.
- L. Jia, J. Liu, Z. Song, X. Pan, L. Chen, X. Cui and M. Wang, Berberine suppresses amyloid-beta-induced inflammatory response in microglia by inhibiting nuclear factor-kappaB and mitogen-activated protein kinase signalling pathways, J. Pharm. Pharmacol., 2012, 64(10), 1510–1521 CrossRef.
- D. N. Chandra, G. K. Prasanth, N. Singh, S. Kumar, O. Jithesh, C. Sadasivan, S. Sharma, T. P. Singh and M. Haridas, Identification of a novel and potent inhibitor of phospholipase A(2) in a medicinal plant: crystal structure at 1.93Å and Surface Plasmon Resonance analysis of phospholipase A(2) complexed with berberine, Biochim. Biophys. Acta, 2011, 1814(5), 657–663 CrossRef.
- X. Feng, K. Wang, S. Cao, L. Ding and F. Qiu, Pharmacokinetics and Excretion of Berberine and Its Nine Metabolites in Rats, Front. Pharmacol, 2020, 11, 594852 CrossRef.
- K. Wang, X. Feng, L. Chai, S. Cao and F. Qiu, The metabolism of berberine and its contribution to the pharmacological effects, Drug Metab. Rev., 2017, 49(2), 139–157 CrossRef CAS PubMed.
- H. S. Cui, S. Hayasaka, X. Y. Zhang, Y. Hayasaka, Z. L. Chi and L. S. Zheng, Effect of berberrubine on interleukin-8 and monocyte chemotactic protein-1 expression in human retinal pigment epithelial cell line, Life Sci., 2006, 79(10), 949–956 Search PubMed.
- Y. Y. Chen, R. Y. Li, M. J. Shi, Y. X. Zhao, Y. Yan, X. X. Xu, M. Zhang, X. T. Zhao and Y. B. Zhang, Demethyleneberberine alleviates inflammatory bowel disease in mice through regulating NF-κB signaling and T-helper cell homeostasis, Inflamm. Res., 2017, 66(2), 187–196 CrossRef CAS.
- X. T. Yu, Y. F. Xu, Y. F. Huang, C. Qu, L. Q. Xu, Z. R. Su, H. F. Zeng, L. Zheng, T. G. Yi, H. L. Li, J. P. Chen and X. J. Zhang, Berberrubine attenuates mucosal lesions and inflammation in dextran sodium sulfate-induced colitis in mice, PLoS One, 2018, 13(3), e0194069 CrossRef.
- Y. Han, C. Ge, J. Ye, R. Li and Y. Zhang, Demethyleneberberine alleviates Pseudomonas aeruginosa-induced acute pneumonia by inhibiting the AIM2 inflammasome and oxidative stress, Pulm. Pharmacol. Ther., 2023, 83, 102259 CrossRef CAS.
- X. Liu, Z. Hu, Q. Shi, H. Zeng, Y. Shen, H. Jin and W. Zhang, Anti-inflammatory and anti-nociceptive activities of compounds from Tinospora sagittata (Oliv.) Gagnep, Arch Pharm. Res., 2010, 33(7), 981–987 Search PubMed.
- S. Niu, M. Jing, J. Wen, S. Wei, H. Li, X. Li, X. Ma and Y. Zhao, Jatrorrhizine Alleviates DSS-Induced Ulcerative Colitis by Regulating the Intestinal Barrier Function and Inhibiting TLR4/MyD88/NF-κB Signaling Pathway, J. Evidence-Based Complementary Altern. Med., 2022, 2022, 3498310 Search PubMed.
- H. Xia, Y. Liu, G. Xia, Y. Liu, S. Lin and L. Guo, Novel Isoquinoline Alkaloid Litcubanine A - A Potential Anti-Inflammatory Candidate, Front. Immunol., 2021, 12, 685556 Search PubMed.
- T. Jayakumar, C. M. Yang, T. L. Yen, C. Y. Hsu, J. R. Sheu, C. W. Hsia, M. Manubolu, W. C. Huang, C. Y. Hsieh and C. H. Hsia, Anti-Inflammatory Mechanism of An Alkaloid Rutaecarpine in LTA-Stimulated RAW 264.7 Cells: Pivotal Role on NF-κB and ERK/p38 Signaling Molecules, Int. J. Mol. Sci., 2022, 23(11), 5889 CrossRef CAS.
- T. C. Moon, M. Murakami, I. Kudo, K. H. Son, H. P. Kim, S. S. Kang and H. W. Chang, A new class of COX-2 inhibitor, rutaecarpine from Evodia rutaecarpa, Inflamm. Res., 1999, 48(12), 621–625 CrossRef CAS.
- K. M. Tian, J. J. Li and S. W. Xu, Rutaecarpine: A promising cardiovascular protective alkaloid from Evodia rutaecarpa (Wu Zhu Yu), Pharmacol. Res., 2019, 141, 541–550 CrossRef CAS PubMed.
- S. Harirforoosh, W. Asghar and F. Jamali, Adverse effects of nonsteroidal antiinflammatory drugs: an update of gastrointestinal, cardiovascular and renal complications, J. Pharm. Nutr. Sci., 2013, 16(5), 821–847 Search PubMed.
- T. Grosser, S. Fries and G. A. FitzGerald, Biological basis for the cardiovascular consequences of COX-2 inhibition: therapeutic challenges and opportunities, J. Clin. Invest., 2006, 116(1), 4–15 CrossRef CAS PubMed.
- C. Patrono, Cardiovascular effects of cyclooxygenase-2 inhibitors: a mechanistic and clinical perspective, Br. J. Clin. Pharmacol., 2016, 82(4), 957–964 CrossRef CAS PubMed.
- A. A. El-Malah, M. M. Gineinah, P. K. Deb, A. N. Khayyat, M. Bansal, K. N. Venugopala and A. S. Aljahdali, Selective COX-2 Inhibitors: Road from Success to Controversy and the Quest for Repurposing, Pharmaceuticals, 2022, 15(7), 827 Search PubMed.
- C. P. Cannon and P. J. Cannon, COX-2 Inhibitors and Cardiovascular Risk, Science, 2012, 336(6087), 1386–1387 CrossRef CAS.
- Y. Yu, E. Ricciotti, R. Scalia, S. Y. Tang, G. Grant, Z. Yu, G. Landesberg, I. Crichton, W. Wu, E. Puré, C. D. Funk and G. A. FitzGerald, Vascular COX-2 modulates blood pressure and thrombosis in mice, Sci. Transl. Med., 2012, 4(132), 132ra54 Search PubMed.
- J. R. Sheu, Y. C. Kan, W. C. Hung, C. H. Su, C. H. Lin, Y. M. Lee and M. H. Yen, The antiplatelet activity of rutaecarpine, an alkaloid isolated from Evodia rutaecarpa, is mediated through inhibition of phospholipase C, Thromb. Res., 1998, 92(2), 53–64 CrossRef.
- S. Jia and C. Hu, Pharmacological effects of rutaecarpine as a cardiovascular protective agent, Molecules, 2010, 15(3), 1873–1881 CrossRef PubMed.
- G. J. Roth, N. Stanford and P. W. Majerus, Acetylation of prostaglandin synthase by aspirin, Proc. Natl. Acad. Sci. U. S. A., 1975, 72(8), 3073–3076 CrossRef PubMed.
- D. C. Dai, X. F. Xu, H. Yan and Y. Zhang, Three new indole alkaloid derivatives from Fissistigma oldhamii Levl. and their anti-inflammatory effects, Fitoterapia, 2024, 175, 105910 CrossRef PubMed.
- D. J. Newman and G. M. Cragg, Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019, J. Nat. Prod., 2020, 83(3), 770–803 CrossRef.
- T. T. Ho, Q. T. N. Tran and C. L. L. Chai, The polypharmacology of natural products, Future Med. Chem., 2018, 10(11), 1361–1368 CrossRef PubMed.
- M. L. Bolognesi, Harnessing Polypharmacology with Medicinal Chemistry, ACS Med. Chem. Lett., 2019, 10(3), 273–275 CrossRef.
- A. Kabir and A. Muth, Polypharmacology: The science of multi-targeting molecules, Pharmacol. Res., 2022, 176, 106055 CrossRef.
- E. Proschak, H. Stark and D. Merk, Polypharmacology by Design: A Medicinal Chemist's Perspective on Multitargeting Compounds, J. Med. Chem., 2019, 62(2), 420–444 CrossRef.
- D. Domingo-Fernández, Y. Gadiya, A. J. Preto, C. A. Krettler, S. Mubeen, A. Allen, D. Healey and V. Colluru, Natural Products Have Increased Rates of Clinical Trial Success throughout the Drug Development Process, J. Nat. Prod., 2024, 87(7), 1844–1851 CrossRef.
- A. L. Heinzke, A. Pahl, B. Zdrazil, A. R. Leach, H. Waldmann, R. J. Young and P. D. Leeson, Occurrence of "Natural Selection" in Successful Small Molecule Drug Discovery, J. Med. Chem., 2024, 67(13), 11226–11241 CrossRef.
- S. O'Hagan and D. B. Kell, Consensus Rank Orderings of Molecular Fingerprints Illustrate The Most Genuine Similarities Between Marketed Drugs and Small Endogenous Human Metabolites, but Highlight Exogenous Natural Products as The Most Important ‘Natural’ Drug Transporter Substrates, ADMET and DMPK, 2017, 5, 2, pp. 85–125 Search PubMed.
- M. J. Waring, J. Arrowsmith, A. R. Leach, P. D. Leeson, S. Mandrell, R. M. Owen, G. Pairaudeau, W. D. Pennie, S. D. Pickett, J. Wang, O. Wallace and A. Weir, An analysis of the attrition of drug candidates from four major pharmaceutical companies, Nat. Rev. Drug Discovery, 2015, 14(7), 475–486 CrossRef PubMed.
- A. Panossian, A. Hovhannisyan, G. Mamikonyan, H. Abrahamian, E. Hambardzumyan, E. Gabrielian, G. Goukasova, G. Wikman and H. Wagner, Pharmacokinetic and oral bioavailability of andrographolide from Andrographis paniculata fixed combination Kan Jang in rats and human, Phytomedicine, 2000, 7(5), 351–364 CrossRef.
- P. Mátyus and C. L. Chai, Metabolism-Activated Multitargeting (MAMUT): An Innovative Multitargeting Approach to Drug Design and Development, ChemMedChem, 2016, 11(12), 1197–1198 CrossRef.
- P. Ma and R. J. Mumper, Paclitaxel Nano-Delivery Systems: A Comprehensive Review, J. Nanomed. Nanotechnol., 2013, 4(2), 1000164 Search PubMed.
- M. Markovic, S. Ben-Shabat and A. Dahan, Prodrugs for Improved Drug Delivery: Lessons Learned from Recently Developed and Marketed Products, Pharmaceutics, 2020, 12(11), 1031 CrossRef PubMed.
- A. B. Karbasi, J. D. Barfuss, T. C. Morgan, D. Collins, D. A. Costenbader, D. G. Dennis, A. Hinman, K. Ko, C. Messina, K. C. Nguyen, R. C. Schugar, K. A. Stein, B. B. Williams, H. Xu, J. P. Annes and M. Smith, Sol-moiety: Discovery of a water-soluble prodrug technology for enhanced oral bioavailability of insoluble therapeutics, Nat. Commun., 2024, 15(1), 8487 CrossRef PubMed.
- I. Kaya and G. Colmenarejo, Analysis of nuisance substructures and aggregators in a comprehensive database of food chemical compounds, J. Agric. Food Chem., 2020, 68(33), 8812–8824 CrossRef.
- J. Bisson, J. B. McAlpine, J. B. Friesen, S. N. Chen, J. Graham and G. F. Pauli, Can Invalid Bioactives Undermine Natural Product-Based Drug Discovery?, J. Med. Chem., 2016, 59(5), 1671–1690 CrossRef.
- R. F. Dantas, T. C. S. Evangelista, B. Neves Jr, M. R. Senger, C. H. Andrade, S. B. Ferreira and F. P. Silva-Junior, Dealing with frequent hitters in drug discovery: a multidisciplinary view on the issue of filtering compounds on biological screenings, Expet Opin. Drug Discov., 2019, 14(12), 1269–1282 CrossRef.
- R. Sheridan and K. Spelman, Polyphenolic promiscuity, inflammation-coupled selectivity: whether PAINs filters mask an antiviral asset, Front. Pharmacol, 2022, 13, 909945 CrossRef.
- J. B. Baell, Screening-based translation of public research encounters painful problems, ACS Med. Chem. Lett., 2015, 6(3), 229–234 CrossRef PubMed.
- J. Baell and M. A. Walters, Chemistry: Chemical con artists foil drug discovery, Nature, 2014, 513(7519), 481–483 CrossRef PubMed.
- M. Z. Y. Choo and C. L. L. Chai, Promoting GAINs (Give Attention to Limitations in Assays) over PAINs Alerts: no PAINS, more GAINs, ChemMedChem, 2022, 17(7), e202100710 CrossRef PubMed.
| This journal is © The Royal Society of Chemistry 2025 |