
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
POMC-specific modulation of metabolic and immune pathways via melanocortin-3 receptor signaling†
Mariya
Nezhyva‡
a,
Friederike A.
Sandbaumhüter‡
a,
Per E.
Andrén
 ab and
Erik T.
Jansson
*a
ab and
Erik T.
Jansson
*a
aDepartment of Pharmaceutical Biosciences, Uppsala University, 751 24 Uppsala, Sweden. E-mail: erik.jansson@uu.se
bScience for Life Laboratory, Spatial Mass Spectrometry, Uppsala University, 751 24 Uppsala, Sweden
First published on 11th June 2025
Abstract
This proteomic study provides a nuanced mechanistic understanding of the signaling processes upon agonist binding to the melanocortin-3 receptor (MC3R). Utilizing thermal proteome profiling (TPP) combined with LC–MS, we uncovered the distinct influences of the endogenous agonists adrenocorticotropic hormone (ACTH), α-melanocyte-stimulating hormone (α-MSH), and γ-melanocyte-stimulating hormone (γ-MSH) on protein thermal stability and pathway activation. In our 2D-TPP study, transfected HEK293 cells for expression of MC3R were exposed to the three endogenous MC3R-ligands across several concentrations followed by incubation at several temperatures, centrifugation and LC–MS analysis of the resulting supernatants. This enabled us to assess the effects of type of ligand and concentration on the thermal stability of proteins in these cells. We employed a combination of multivariate analysis, differential expression, TPP and pathway analysis to deeply characterize the impact of MC3R activation on molecular mechanisms. All three ligands affected signaling pathways related to the immune system and energy homeostasis. While α-MSH significantly modulated the IL-6 pathway via STAT3, and γ-MSH prominently activated interferon signaling, ACTH uniquely affected NADPH-related proteins. All ligands shared involvement in the cAMP-PKA-CREB and varied impacts on PI3K and ERK pathways, crucial for energy metabolism. All proteomic data are available under DOI: https://doi.org/10.6019/PXD039945.
Introduction
The melanocortin-3 receptor (MC3R), a G-protein coupled receptor prominently expressed in the brain, pancreas, placenta, and immune cells, plays a critical role in various physiological functions, including immune response and energy homeostasis.1–3 The interplay between MC3R and food intake disorders, such as obesity and chronic inflammation, is mediated by hormones and cytokines.4–7 Peptide hormones and neuropeptides, such as cholecystokinin (CCK), leptin, peptide tyrosine–tyrosine (PYY), insulin, and ghrelin, modulate the release of pro-opiomelanocortin (POMC)-derived peptides and Agouti-related protein (AgRP), further regulating MC3R signaling.8,9 The activation of MC3R by POMC-derived peptides like adrenocorticotropic hormone (ACTH), α-melanocyte-stimulating hormone (α-MSH), and γ-melanocyte-stimulating hormone (γ-MSH), has been directly linked to effects such as energy expenditure and reduced food intake.10 Because the structurally closely related neuropeptides ACTH, α-MSH, and γ-MSH share the critical His–Phe–Arg–Trp amino acid sequence crucial for their interaction with MC3R, and have very similar binding affinities (although MC3R shows a slight preference for γ-MSH), these ligands offer the unique opportunity to investigate important differences between the ligands. While the upstream modulators of MC3R and their general physiological roles are relatively well characterized, significant gaps remain in our understanding of the downstream pathways and their specific, ligand-dependent modulation. Previous findings have demonstrated that agonist binding leads to an increase in cAMP levels, impacting inflammatory mediators like cytokines. While these effects are well studied for ACTH and α-MSH, they are less understood for γ-MSH.6,11–13 Understanding the multi-stage signaling cascades that convey chemical messages from initial receptor–ligand contact to biological responses is crucial for developing effective and safe therapies.Advances in data-rich technologies like proteomics lead to large and complex datasets necessitating the development and implementation of novel data analysis strategies to extract meaningful biological insights. Thermal proteome profiling (TPP) has recently emerged as a method that enables the untargeted study of gain or loss of molecular interaction upon stimulation of a cell or tissue with a drug.1,2,14 In contrast to differential expression analysis, the method focuses on finding those proteins that interact within a chain of events due to drug stimulation. In this study, we focus on receptor–ligand interactions that govern physiological and pathophysiological processes, specifically examining the downstream signaling cascades activated by MC3R. We have developed a method combining MS-based TPP with a multidimensional data analysis workflow and applied it to elucidate the effects of endogenous agonist binding to MC3R.3
Here, we provide a detailed analysis of two-dimensional TPP coupled with LC–MS to MC3R-expressing HEK293 cells treated with ACTH, α-MSH, or γ-MSH. Multivariate, differential, and pathway analyses, together with cAMP-readouts allow us to dissect G-protein-mediated activation. We provide information extraction and interpretation, including principal component analysis (PCA) and combine experimental data on thermally affected proteins with differential expression profiles and present the results obtained from a previously acquired dataset3 in their biological context. Further, we provide a molecular context for these findings with ingenuity pathway analysis (IPA).15 By integrating the multiple data analysis methods PCA, differential expression, thermal stability and pathway analysis, we provide deeper insights into protein–ligand interactions.
PCA simplifies our dataset by identifying key clusters that represent distinct biological states, allowing us to visualize patterns and pinpoint proteins significantly affected by ligand binding or receptor activation. These clusters can highlight novel protein interactions and pathways for further investigation. By mapping melting profiles under various conditions, TPP elucidates how ligand binding, and its impact on signaling cascades impact protein stability and function. Integrating data on differential expression, phosphorylation, and transcription factor activity facilitates the identification of proteins relevant to MC3R responses,3 illustrating the complex interplay of biological processes and regulatory mechanisms.
Further, we enhance our analysis by comparing outputs from PCA and TPP with IPA. This side-by-side comparison enriches our understanding by directly highlighting relevant proteins, streamlining the identification of potential biomarkers and therapeutic targets without extensive database searches. We show here how this synergistic approach enables researchers to form a holistic view of the biological processes involved, fostering the generation of robust hypotheses that enable data-driven research. By leveraging the strengths of each analysis method, we examplify here how the complexities of receptor-mediated signaling can be navigated.
Materials and methods
The experimental procedures used to generate the dataset analyzed here have been previously described. For detailed information on the materials and methods used to obtain the dataset, please refer to Sandbaumhüter et al.3 Briefly, human embryonic kidney 293 cells overexpressing human MC3R (HEK-TREx-MC3R) were treated with the endogenous ligands ACTH, α-MSH, and γ-MSH dissolved in DMSO or with DMSO only (control) for 1 h prior to cell harvest. The functionality of the receptor was confirmed with the cAMP-GloTM Assay (Promega Corporation, Madison, WI). During the TPP experiment aliquots were first incubated at various temperatures between 37 and 67 °C for 3 min, followed by snap-freezing in liquid nitrogen. After cell lysis the soluble and aggregated proteins were separated by centrifugation (14![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g at 4 °C for 90 min). The supernatant was purified and present proteins were digested by trypsin using filter-aided sample preparation. The resulting peptides were analyzed with LC–MS (nanoAcquity and Synapt G2-Si, Waters Corporation, Milford, MA, USA).
000g at 4 °C for 90 min). The supernatant was purified and present proteins were digested by trypsin using filter-aided sample preparation. The resulting peptides were analyzed with LC–MS (nanoAcquity and Synapt G2-Si, Waters Corporation, Milford, MA, USA).
Data analysis
Next, we conducted TPP data analysis, described in detail elsewhere.3 Briefly, protein abundance data obtained from ISOQuant underwent log2-transformation and centering. Subsequently, statistical models were employed to scrutinize both temperature and concentration-dependent behavior of the proteins. Two hypotheses were considered: a null hypothesis H0, suggesting no drug–protein interaction, and an H1 alternate hypothesis H1, characterized by a log-logistic function. The F-statistic was used to assess the goodness-of-fit between these models. Additionally, false discovery rates (FDR) were computed using a bootstrap method, applying a cut-off of 0.01 to distinguish proteins with significantly altered thermal stability.3,18,19 Furthermore, we used the protein differential expression results for pathway analysis through the pathway analysis with down-weighting of overlapping genes (PADOG) algorithm in the reactome GSA software.
In addition to what we previously described,3 we here also used PCA on the ISOQuant output after log-transformation and normalized the protein abundance values for 37 °C. The first two principal components were selected for their high explained variance, which collectively accounted for 66.9% of the total variance in the dataset. Scores and loadings plots were generated to visualize the clustering patterns with metascape20 of the samples based on their ligand treatments.
Protein differential expression analysis was conducted on samples treated with ligand at 37 °C with limma package. Pathway analysis was performed using the IPA suite (version 23.0, QIAGEN, DUS, Germany). Core pathway analysis was constructed by uploading a list of differentially expressed proteins and hill values (for TPP data as a measure of stability instead of protein fold change) along with their UniProt identification. Pathway output was generated with a p-value threshold of <0.05.
Results and discussion
Binding of the three structurally related endogenous agonists, ACTH, α-MSH and γ-MSH, to MC3R triggers a signaling cascade resulting in effects on the immune response and energy homeostasis for all three ligands (Fig. 1). Based on differential protein expression and gene set enrichment analysis, we identified biological pathways affected by the binding of the different agonists and placed them into context with proteins that showed a melting temperature shift in the TPP experiments. The melting temperature shift originates from changes in the thermal stability of a protein resulting from direct or indirect ligand effects on its binding status. For the POMC-derived neuropeptides, our results highlight the modulation of key proteins and pathways such as the cAMP-PKA-CREB and RAS-RAF-MAPK pathway21 (Table 1). | ||
| Fig. 1 Ligand-specific and common molecular pathway modulation by ACTH, α-MSH, and γ-MSH. This schematic represents the differential impact of melanocortin ligands on metabolic and immune signaling pathways. Shared effects are depicted centrally (in dark green), illustrating the upregulation of adenylyl cyclase (AC) leading to increased cAMP levels, activation of protein kinase A (PKA), and phosphorylation of cAMP response element-binding protein (CREB) by all ligands. Ligand specific effects are shown in purple for ACTH, light green for α-MSH and yellow for γ-MSH. Proteins are denoted with thermally stabilized (T), phosphorylated (P), upregulated (↑), downregulated (↓) indicating the complex interplay between ligand-specific and universal signaling events mediated by melanocortin interactions. | ||
| Pathway | Protein | Thermal effect | Ligand |
|---|---|---|---|
| a TLR4, TLR5, TLR7/8, TLR9, TLR10, TLR1:TLR2, TLR6:TLR2. | |||
| Activation of the AP-1 family of transcription factors | MK01 | Stabilized | ACTH |
| Costimulation by the CD28 family | 2A5G | Destabilized | γ-MSH |
| ERK/MAPK targets | MK01 | Stabilized | ACTH |
| ERKs are inactivated | MK01 | Stabilized | ACTH |
| FCERI mediated MAPK activation | MK01 | Stabilized | ACTH |
| Growth hormone receptor signaling | MK01 | Stabilized | ACTH |
| Interleukin-17 signaling | MK01; MP2K1 | Stabilized | ACTH |
| Interleukin-3 & Interleukin-5 and GM-CSF signaling | CRK | Stabilized | ACTH |
| Interleukin-4 and Interleukin-13 signaling | VIME | Destabilized | ACTH |
| HSP7C | Stabilized | α-MSH | |
| ISG15 antiviral mechanism | NUP62 | Destabilized | ACTH |
| IF4G1; NUP50 | Stabilized | ACTH | |
| MHC class II antigen presentation | KIF3C; TBA1C | Destabilized | ACTH |
| CAZA2 | Stabilized | ACTH | |
| KLC1 | Stabilized | α-MSH | |
| DCTN1 | Destabilized | γ-MSH | |
| MyD88 cascade initiated on plasma membrane | MK01; MP2K1 | Stabilized | ACTH |
| MyD88 dependent cascade initiated on endosome | MK01; MP2K1 | Stabilized | ACTH |
| MyD88:MAL(TIRAP) cascade initiated on plasma membrane | MK01; MP2K1 | Stabilized | ACTH |
| Regulation of actin dynamics for phagocytic cup formation | CRK; MK01 | Stabilized | ACTH |
| COF1 | Destabilized | α-MSH | |
| Regulation of signaling by CBL | CRK | Stabilized | ACTH |
| Role of phospholipids in phagocytosis | PLD3 | Stabilized | ACTH; γ-MSH |
| Toll like receptor cascadesa | MK01; MP2K1 | Stabilized | ACTH |
| TRAF6 mediated induction of NFκ B and MAP kinases upon TLR7/8 or 9 activation | MK01; MP2K1 | Stabilized | ACTH |
PCA revealed distinct proteome-wide thermal profiles for each POMC peptide, supporting ligand-specific signaling biases. The different agonists cluster in individual areas, while the different concentrations cluster tightly together for the individual agonists (Fig. 2A). The top 50 ranked proteins from the PCA loading matrix elucidate proteins that have the highest impact on discriminating between the ligand-mediated effects, hence, the discrepancy in pathway regulation (Fig. 2B). The list includes regulatory proteins such as transcription factors (transcription factor JunD [JUND], retinoic acid receptor RXR-beta [RXRB], ribonucleoside-diphosphate reductase subunit [RIR2B]) as well as proteins involved in immune response (tubulin beta-4a [TBB4A]). Differences between the POMC peptides are also reflected in affected biological processes such as NFκ B, Rho GTPases signaling-related pathways, and thermally affected proteins. We identified a set of thermally affected proteins linked to diverse pathways involved in signal transduction, immune response, and energy homeostasis that will be discussed in more detail in the further sections (Fig. 2D). These findings align well with our previous report where we identified the transcription factors CCAR2, DDX21, HMGB2, SRSF7, and TET2 that have been thermally affected and phosphorylated besides known immunoregulatory transcription factors (IRF, STAT, NFκB).3 These transcription factors are implicated as downstream targets in the MC3R signaling cascade and are crucial in immune responses.8,9,22,23
 | ||
| Fig. 2 Multivariate and network analysis of POMC-derived peptide effects: (A) scores (purple for ACTH, green for α-MSH, yellow for γ-MSH) and (B) loadings plots obtained from a principal component analysis of the differential expression patterns (relative to vehicle-only negative controls) observed after treatment with the three POMC ligands at three concentrations (20, 100, and 500 nM). The loadings plot shows the proteins with the 50 highest loadings and their positions in the two-dimensional space spanned by principal components 1 and 2. (C) Network of enriched ontology clusters of proteins with Euclidean distance >0.1 based on PCA analysis (metascape,20); (D) Sankey diagram showing connections between the POMC-peptides ACTH, α-, and γ-MSH, the affected biological processes, and the proteins identified with affected thermal stability as a result of treatment. | ||
Signal transduction
Our data corroborates well with known mechanisms of MC3R, where signals are mediated upon activation via the cAMP-PKA-CREB signaling pathway (Fig. 1).24,25 In this pathway, the Gsα subunit plays a key role by activating adenylyl cyclase, leading to elevated cAMP levels. We observed a significant upregulation in Gsα activity, supported by an average log2 fold change (μFC) of 5 (File S1 [XLSX], ESI†). This activation further triggers secondary messenger systems involving protein kinases and Rho GTPases. As downstream consequences, we detected the upregulation of RAS, RAF, and p38 MAPK pathways, along with the stabilization of mitogen-activated protein kinase 1 (MK01) for all tested ligands (File S1 [XLSX], ESI†). These findings are further corroborated by our previous confirmation of an increase in cAMP after target engagement.3Simultaneously, we identified a strong involvement of Wnt signaling, which plays a role in inflammatory and neurovegetative processes. This observation was corroborated by the involvement of ubiquinol biosynthesis in ACTH-treated samples (Fig. 2B and File S1 [XLSX], ESI†). Wnt signaling was previously found to interface with ERK and MAPK pathways through β-catenin phosphorylation by glycogen synthase kinase 3 (GSK3).26 This interaction was particularly evident for γ-MSH and α-MSH, with both canonical FZD proteins and non-canonical ROR receptors involved. Further, our PCA analysis of differential protein expression highlighted the protein RXRB, which interacts with Wnt-canonical pathways and was only observed in samples treated with γ-MSH or ACTH (Fig. 2B and File S1 [XLSX], ESI†).
Regarding ACTH, a cascade of downstream signaling pathways with considerable overlap in their targets was activated and led to significant downstream effects. In detail, FGFR4 pathways were activated, resulting in GSK3 phosphorylation and the subsequent stimulation of β-catenin/TCF signaling.27 Key proteins like MK01, phosphorylated forms of tyrosine–protein phosphatase non-receptor type 11 (PTN11), serine/threonine–protein phosphatase (2AAA), and ubiquitin-60S ribosomal protein (RL40) were implicated (File S1 [XLSX], S2 [CSV], S6 [XLSX], ESI†). Conforming with this, there was a downregulation of STAT1 activity. Fibroblast growth factors (FGFs) exhibited an upregulation with a μFC of 6, which led to further upregulation of MAPK and PI3K signaling. Additionally, PLCG2-mediated IP3 and DAG signaling were activated in response to FGFR4 activity in the ACTH samples (File S1 [XLSX], ESI†).
In contrast to these findings, all three ligands activated ERK/MAPK cascades and SRC family kinases, consistent with known MC4R signaling axes.28 ERK/MAPK signaling can be initiated by both directly through G-protein dependent mechanisms or indirectly by β-arrestin mediated scaffolding. This was supported by increased intracellular Ca2+-mobilization, enhancing tropomyosin alpha chain signaling across all conditions studied. However, here we could identify some ligand-specific differences as well as for the transcription factor JUND. It was among the top 50 PCA loadings, suggesting its central role in ligand differentiation (Fig. 2B). Moreover, nerve growth factor-independent TrkA activation was exclusively upregulated in ACTH samples, while ALK signaling was upregulated in α-MSH and ACTH samples (File S1 [XLSX], ESI†).29 Confirming the previously described signal transduction processes with our experimental results, we could expand the knowledge to ligand-specific modulations and gained confidence for zooming into other modulated pathways such as the immune system.
Immune system
A rapid immune response to environmental, mechanical, chemical, and infectious stressors is based on the inducible regulation of protein expression through transcription factors. Central roles in the coordination of this dynamic response are the interferon regulatory factors (IRFs), signal transducers and activators of transcription (STATs), and the nuclear factor kappa-light-chain-enhancer of activated B-cells (NFκB). Our study reveals that the activation of the MC3R by ACTH, as well as α-MSH and γ-MSH, exerts distinct ligand-specific effects on the signaling pathways both upstream and downstream of these critical mediators (Fig. 3 and Table 1).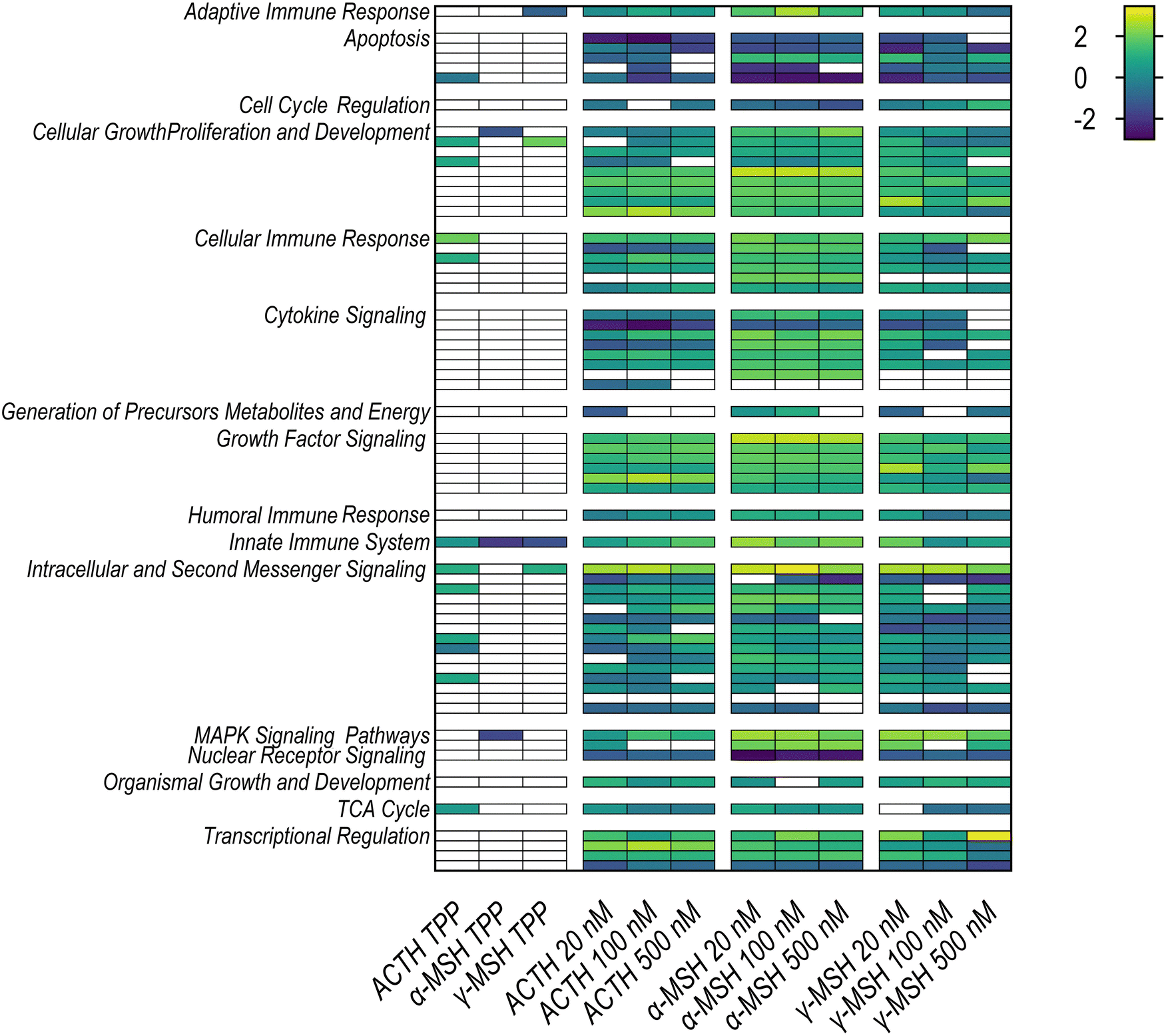 | ||
| Fig. 3 Heatmap showing pathway analysis with IPA as an effect of treatment with ACTH, α-MSH and γ-MSH at various concentrations compared to vehicle-only, and thermally affected proteins. | ||
Of the three studied ligands, ACTH stands out with the highest number of upregulated and thermally stabilized proteins related to the immune system (Table 1). ACTH facilitates an anti-inflammatory response via both a cortisol-dependent and an independent mechanism, with the latter being mediated through MC3R.30 Our study highlights ACTH's prominent role in these MC3R-mediated immunomodulatory processes; a role shared to a lesser extent by α-MSH and γ-MSH. The initial pro-inflammatory phase in immune activation is typically followed by an anti-inflammatory counterbalance.31 Hence, cytokine signaling pathways, prominently those involving interferons (IFNs) and interleukins (ILs), are pivotal in the characterization of the immune modulation by MC3R. Both, IFN and IL signaling were elevated by the treatment of all three POMC-derived ligands (Table 1 and File S1 [XLSX], ESI†).
The influence of ligands on interferon signaling was evaluated by the pathway activity of IRFs and STATs. Remarkably, γ-MSH presented robust activation of IFN signaling, especially for IFNα and IFNβ. ACTH and α-MSH induced these pathways as well, but their effects were more moderate (Fig. 3). When IFNs bind to their receptors, they can initiate the formation of complexes with Janus-associated kinase (JAK) 1 and tyrosine kinase (TYK) 2, leading to phosphorylation of the transcription factor STAT1, known as the JAK-STAT pathway, which has a crucial role in the immune response and its involvement in MC3R signaling.13
In our study, we identified that all ligands (ACTH, α-, and γ-MSH) upregulate transcription factor activity of STAT3 and enhance its phosphorylation through PTK6 signaling, which plays a pivotal role in modulating the IL-6 pathway and is linked to c-SRC kinase activity (File S3-5-5 [XLSX], ESI†). This leads to increased levels of IL-6, its receptor IL6R, and the signal transducer IL6ST, promoting phosphorylation and dimerization of STAT proteins (1, 3, 5A/B) and elevated expression of the cytokines IL6 and IL10. Notably, the degree of IL-6 regulation varies among the ligands, with α-MSH inducing the most significant change, approximately 9.7-fold, as seen in Fig. 3 and File S1 [XLSX] (ESI†). IL-6 acts as a pro-inflammatory cytokine and anti-inflammatory myokine in the initial phase of the immune response. Further, it is highly present in adipose tissue.5
During inflammation, the transcription factor TET-methylcytosine-dioxygenase 2 (TET2) modulates IL-6 activity. Interestingly, we observed a thermal destabilization of TET2 (File S3–S5 [XLSX], ESI†), which generally suggests reduced interaction with other molecules, and an increase in its phosphorylated form indicates a potentially active role in immune response regulation.3 This dualistic nature of TET2's regulation was mirrored in the activities of transcription factors involved in the immune response; a decrease in anti-inflammatory STAT1 activity was coupled with an increase in pro-inflammatory STAT3 activity.3 Further complexities in signaling pathways were highlighted by the thermal destabilization of vimentin (VIME), associated with pro-inflammatory STAT3 signaling and phosphorylation by ACTH (see Table 1 and File S3–S5 [XLSX], ESI†), contrasting with a decrease in activity and phosphorylation of high mobility group box 2 (HMGB2), which is involved in pro-inflammatory processes.3 This activity corresponds with enhanced signaling pathways such as p38, ERK, JNK, and PI3K (File S1 [XLSX], ESI†).
Furthermore, NFκ B, another key player regulated by ILs, interacts with RIR2B, a top 50 PCA loadings protein that was thermally stabilized in the ACTH and γ-MSH treatment groups (Fig. 2 and File S2 [CSV], ESI†).
All three endogenous ligands clearly affected the immune response triggered by MC3R engagement. While ACTH affected the highest number of immune-related proteins, γ-MSH showed the strongest activation of IFN signaling and α-MSH was the major effector of the IL-6 cascade.
Energy homeostasis
MC3R-mediated signaling also extends to energy balance through modulation of metabolic pathways. We identified differential thermal responses among proteins in key metabolic processes following treatment with ACTH, α-MSH, and γ-MSH (File S1 [XLSX], ESI†).All three ligands were found to modulate the critical cAMP-PKA-CREB signaling pathway, which is integral to energy balance and connects to numerous metabolic pathways.21 Interestingly, the treatment with these peptides resulted in varied effects on the PI3K and ERK pathways, crucial to glucose and lipid metabolism.32–34 The observed activation of the ERK pathway is directly connected to AMP-activated protein kinase (AMPK), known as the sensor for energy and regulator of energy homeostasis33–36 (File S1 [XLSX], ESI†).
PCA highlighted mitochondrial proteins, including TIM8A and ATP5J, as discriminators of ligand-specific responses (Fig. 2), suggesting differential regulation of mitochondrial function. Consistent with this, ACTH uniquely modulated NADPH-producing enzymes—such as isocitrate dehydrogenase, succinate dehydrogenase, and aconitase—while α-MSH and γ-MSH preferentially affected proteins involved in pyruvate metabolism and glycolysis, respectively. Beyond canonical metabolic pathways, we observed ligand-dependent changes in histone deacetylases: HDAC1 and HDAC2 were thermally affected by all three peptides, whereas HDAC7 responded exclusively to ACTH (File S1 [XLSX], ESI†). These epigenetic regulators may link MC3R signaling to long-term metabolic homeostasis and require further investigation.
Conclusions
This study offers a comprehensive, high-confidence map of MC3R signaling by comparing the thermal proteome profiles elicited by its three endogenous agonists—ACTH, α-MSH, and γ-MSH—and integrating differential expression, 2D-TPP, and IPA.By aligning TPP-derived stability shifts with pathway enrichment, we confirmed established axes (cAMP-PKA-CREB, JAK-STAT) and uncovered ligand-specific biases: γ-MSH robustly activates interferon cascades, whereas ACTH uniquely reconfigures NADPH-producing enzymes, and α-MSH and γ-MSH preferentially modulate pyruvate and glycolytic proteins. PCA further implicated mitochondrial factors (TIM8A, ATP5J) in ligand discrimination.
Beyond metabolism and immunity, we identified histone deacetylases (HDAC1/2 for all ligands, HDAC7 for ACTH) as potential mediators of MC3R effects, suggesting novel intervention points. Despite the limitation that all findings were obtained within a HEK293-cell line that does not represent a disease model, our study has generated a better understanding of MC3R-signaling and opens avenues for development and further studies. It will be of particular interest to follow up or results presented here with studies of therapeutic interventions targeting metabolic disorders and immune-related conditions in corresponding tissues.
Conflicts of interest
The authors declare no competing financial interest.Data availability
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository and are accessible via the DOI https://doi.org/10.6019/PXD039945.Acknowledgements
The work in this paper was supported by research grants from the Swedish Research Council (Research Environment Grant Interdisciplinary Research, 2021-03293, P. E. A.; Natural and Engineering Science, 2022-04198, P. E. A.; and 2018-03988, E. T. J.); the Science for Life Laboratory (P. E. A.); the Carl Trygger Foundation, the Magnus Bergvall foundation and the Åke Wiberg Foundation (E. T. J.).References
- R. Jafari, H. Almqvist, H. Axelsson, M. Ignatushchenko, T. Lundbäck, P. Nordlund and D. M. Molina, The cellular thermal shift assay for evaluating drug target interactions in cells, Nat. Protoc., 2014, 9, 2100–2122 CrossRef CAS PubMed.
- A. Mateus, T. A. Määttä and M. M. Savitski, Thermal proteome profiling: unbiased assessment of protein state through heat-induced stability changes, Proteome Sci., 2016, 15, 1–7 CrossRef PubMed.
- F. A. Sandbaumhüter, M. Nezhyva, P. E. Andrén and E. T. Jansson, Label-Free Quantitative Thermal Proteome Profiling Reveals Target Transcription Factors with Activities Modulated by MC3R Signaling, Anal. Chem., 2023, 95, 15400–15408 CrossRef PubMed PMID: 37804223.
- S. J. Getting, Targeting melanocortin receptors as potential novel therapeutics, Pharmacol. Ther., 2006, 111, 1–15 CrossRef CAS PubMed.
- B. J. Renquist, R. N. Lippert, J. A. Sebag, K. L. Ellacott and R. D. Cone, Physiological roles of the melanocortin MC3 receptor, Eur. J. Pharmacol., 2011, 660, 13–20 CrossRef CAS PubMed.
- R. P. Lisak and J. A. Benjamins, Melanocortins, melanocortin receptors and multiple sclerosis, Brain Sci., 2017, 7, 104 CrossRef PubMed.
- A. Ferrante Jr, Obesity-induced inflammation: a metabolic dialogue in the language of inflammation, J. Intern. Med., 2007, 262, 408–414 CrossRef CAS PubMed.
- A. S. Garfield, D. D. Lam, O. J. Marston, M. J. Przydzial and L. K. Heisler, Role of central melanocortin pathways in energy homeostasis, Trends Endocrinol. Metab., 2009, 20, 203–215 CrossRef CAS PubMed.
- G. S. Yeo, D. H. M. Chao, A.-M. Siegert, Z. M. Koerperich, M. D. Ericson, S. E. Simonds, C. M. Larson, S. Luquet, I. Clarke and S. Sharma, et al., The melanocortin pathway and energy homeostasis: from discovery to obesity therapy, Mol. Metab., 2021, 48, 101206 CrossRef CAS PubMed.
- Y. Yang and Y. Xu, The central melanocortin system and human obesity, J. Mol. Cell Biol., 2020, 12, 785–797 CrossRef CAS PubMed.
- D. L. Marks, V. Hruby, G. Brookhart and R. D. Cone, The regulation of food intake by selective stimulation of the type 3 melanocortin receptor (MC3R), Peptides, 2006, 27, 259–264 CrossRef CAS PubMed.
- P. Sweeney, M. N. Bedenbaugh, J. Maldonado, P. Pan, K. Fowler, S. Y. Williams, L. E. Gimenez, M. Ghamari-Langroudi, G. Downing and Y. Gui, et al., The melanocortin-3 receptor is a pharmacological target for the regulation of anorexia, Sci. Transl. Med., 2021, 13, eabd6434 CrossRef CAS PubMed.
- A. E. Moscowitz, H. Asif, L. B. Lindenmaier, A. Calzadilla, C. Zhang and M. Mirsaeidi, The importance of melanocortin receptors and their agonists in pulmonary disease, Front. Med., 2019, 6, 145 CrossRef PubMed.
- H. Franken, T. Mathieson, D. Childs, G. Sweetman, T. Werner, I. Tögel, C. Doce, S. Gade, M. Bantscheff, G. Drewes, F. B. M. Reinhard, W. Huber and M. M. Savitski, Thermal proteome profiling for unbiased identification of direct and indirect drug targets using multiplexed quantitative mass spectrometry, Nat. Protoc., 2015, 10, 1567–1593 CrossRef CAS PubMed.
- A. Krämer, J. Green, J. Pollard Jack and S. Tugendreich, Causal analysis approaches in Ingenuity Pathway Analysis, Bioinformatics, 2013, 30, 523–530 CrossRef PubMed.
- U. Distler, J. Kuharev, P. Navarro, Y. Levin, H. Schild and S. Tenzer, Drift time-specific collision energies enable deep-coverage data-independent acquisition proteomics, Nat. Methods, 2014, 11, 167–170 CrossRef CAS PubMed.
- U. Distler, J. Kuharev, P. Navarro and S. Tenzer, Label-free quantification in ion mobility–enhanced data-independent acquisition proteomics, Nat. Protoc., 2016, 11, 795–812 CrossRef CAS PubMed.
- S. Sridharan, N. Kurzawa, T. Werner, I. Günthner, D. Helm, W. Huber, M. Bantscheff and M. M. Savitski, Proteome-wide solubility and thermal stability profiling reveals distinct regulatory roles for ATP, Nat. Commun., 2019, 10, 1–13 CrossRef CAS PubMed.
- B. Zhang, M. C. Chambers and D. L. Tabb, Proteomic parsimony through bipartite graph analysis improves accuracy and transparency, J. Proteome Res., 2007, 6, 3549–3557 CrossRef CAS PubMed.
- Y. Zhou, B. Zhou, L. Pache, M. Chang, A. H. Khodabakhshi, O. Tanaseichuk, C. Benner and S. K. Chanda, Metascape provides a biologist-oriented resource for the analysis of systems-level datasets, Nat. Commun., 2019, 10, 1523 CrossRef PubMed.
- E. London and C. A. Stratakis, The regulation of PKA signaling in obesity and in the maintenance of metabolic health, Pharmacol. Ther., 2022, 108113 CrossRef CAS PubMed.
- E. J. Anderson, I. Çakir, S. J. Carrington, R. D. Cone, M. Ghamari-Langroudi, T. Gillyard, L. E. Gimenez and M. J. Litt, 60 years of POMC: regulation of feeding and energy homeostasis by α-MSH, J. Mol. Endocrinol., 2016, 56, T157–T174 CAS.
- A. A. Butler, The melanocortin system and energy balance, Peptides, 2006, 27, 281–290 CrossRef CAS PubMed.
- E. J. Lee, S.-H. Lee, J.-W. Jung, W. Lee, B. J. Kim, K. W. Park, S.-K. Lim, C.-J. Yoon and J.-H. Baik, Differential regulation of cAMP-mediated gene transcription and ligand selectivity by MC3R and MC4R melanocortin receptors, Eur. J. Biochem., 2001, 268, 582–591 CrossRef CAS PubMed.
- L. Yang, Neuronal cAMP/PKA signaling and energy homeostasis, Neural Reg. Metab., 2018, 31–48 Search PubMed.
- B. A. Ibarra, C. Machen and R. P. Atit, Wnt-dependent activation of ERK mediates repression of chondrocyte fate during calvarial development, J. Dev. Biol., 2021, 9, 23 CrossRef CAS PubMed.
- Y. Zou, S. Zheng, X. Xie, F. Ye, X. Hu, Z. Tian, S.-M. Yan, L. Yang, Y. Kong and Y. Tang, et al., N6-methyladenosine regulated FGFR4 attenuates ferroptotic cell death in recalcitrant HER2-positive breast cancer, Nat. Commun., 2022, 13, 1–18 Search PubMed.
- D. Ramírez, J. Saba, L. Carniglia, D. Durand, M. Lasaga and C. Caruso, Melanocortin 4 receptor activates ERK-cFos pathway to increase brain-derived neurotrophic factor expression in rat astrocytes and hypothalamus, Mol. Cell. Endocrinol., 2015, 411, 28–37 CrossRef PubMed.
- F. Tabbó, A. Barreca, R. Piva, G. Inghirami and E. T.-C. L. S. Group, ALK signaling and target therapy in anaplastic large cell lymphoma, Front. Oncol., 2012, 2, 41 Search PubMed.
- W. Wang, D.-Y. Guo, Y.-J. Lin and Y.-X. Tao, Melanocortin regulation of inflammation, Front. Endocrinol., 2019, 10, 683 CrossRef PubMed.
- S. J. Getting, Y. Riffo-Vasquez, S. Pitchford, M. Kaneva, P. Grieco, C. P. Page, M. Perretti and D. Spina, A role for MC3R in modulating lung inflammation, Pulm. Pharmacol. Ther., 2008, 21, 866–873 CrossRef CAS PubMed.
- M. S. Savova, L. V. Mihaylova, D. Tews, M. Wabitsch and M. I. Georgiev, Targeting PI3K/AKT signaling pathway in obesity, Biomed. Pharmacother., 2023, 159, 114244 CrossRef CAS PubMed.
- Z. Yang and Y.-X. Tao, Biased signaling initiated by agouti-related peptide through human melanocortin-3 and-4 receptors, Biochim. Biophys. Acta, Mol. Basis Dis., 2016, 1862, 1485–1494 CrossRef CAS PubMed.
- X. Huang, G. Liu, J. Guo and Z. Su, The PI3K/AKT pathway in obesity and type 2 diabetes, Int. J. Biol. Sci., 2018, 14, 1483 CrossRef CAS PubMed.
- G. Baldini and K. D. Phelan, The melanocortin pathway and control of appetite-progress and therapeutic implications, J. Endocrinol., 2019, 241, R1–R33 CAS.
- P. Kühnen, H. Krude and H. Biebermann, Melanocortin-4 receptor signalling: importance for weight regulation and obesity treatment, Trends Mol. Med., 2019, 25, 136–148 CrossRef PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available: File 1 (XLSX): full reactome pathway analysis for all drug and concentration conditions at 37 °C. File 2 (XLSX): list of identified proteins exhibiting altered thermal stability. File 3–5, (XLSX) ISOQuant report file for the analysis of relative protein abundance under all drug, temperature and concentration conditions. File 6 (XLSX): ISOQuant report file for the analysis of relative protein abundance under all drug and concentration conditions at 37 °C with phosphorylation used as a variable modification in the preceding PLGS search. See DOI: https://doi.org/10.1039/d4mo00248b |
| ‡ Contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2025 |