
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAdvancements in microfluidic technology for rapid bacterial detection and inflammation-driven diseases
Jing
Zhang
 ae,
Yatian
Fu
bc,
Ching Yin
Fong
b,
Haojun
Hua
b,
Wei
Li
bc and
Bee Luan
Khoo
*bcd
ae,
Yatian
Fu
bc,
Ching Yin
Fong
b,
Haojun
Hua
b,
Wei
Li
bc and
Bee Luan
Khoo
*bcd
aCollege of Basic Medicine, Hebei University, Baoding, China
bDepartment of Biomedical Engineering, City University of Hong Kong, 83 Tat Chee Avenue, Kowloon, Hong Kong, China. E-mail: blkhoo@cityu.edu.hk
cHong Kong Centre for Cerebro-Cardiovascular Health Engineering (COCHE), Hong Kong, China
dDepartment of Precision Diagnostic and Therapeutic Technology, City University of Hong Kong Shenzhen-Futian Research Institute, Shenzhen 518057, China
eKey Laboratory of Pathogenesis Mechanism and Control of Inflammatory-autoimmune Diseases in Hebei Province, Hebei University, Baoding 071000, China
First published on 9th April 2025
Abstract
Bacterial detection is pivotal for the timely diagnosis and effective treatment of infectious diseases. Microfluidic platforms offer advantages over traditional methods, including heightened sensitivity, rapid analysis, and minimal sample volume requirements. Traditional clinical methods for bacterial identification often involve extended processing times and necessitate high pathogen concentrations, resulting in delayed diagnoses and missed treatment opportunities. Microfluidic technology overcomes these limitations by facilitating rapid bacterial identification at lower biomass levels, thus ensuring prompt and precise treatment interventions. Additionally, bacteria-driven inflammation has been associated with the development and progression of various diseases, including cancer. Elucidating the complex interplay between bacteria, inflammation, and disease is essential for devising effective disease models and therapeutic strategies. Microfluidic platforms have been used to construct in vitro disease models that accurately replicate the intricate microenvironment that bacteria-driven inflammation affects. These models offer valuable insights into bacteria-driven inflammation and its impact on disease progression, such as cancer metastasis and therapeutic responses. This review examines recent advancements in bacterial detection using microfluidics and assesses the potential of this technology as a robust tool for exploring bacteria-driven inflammation in the context of cancer.
 Jing Zhang | Jing Zhang is currently a Lecturer at Hebei University and received her Ph.D. from the City University of Hong Kong. She has experience in the clinical laboratory, and her research interests include microfluidics-based cancer detection, cancer management, and therapy. |
 Yatian Fu | Dr. Yatian Fu is a Postdoctoral Researcher in the Department of Biomedical Engineering at the City University of Hong Kong. Her research interests are constructing in vitro models to simulate disease and studying disease development and treatment. |
 Ching Yin Fong | Miss Fong Ching Yin is a Ph.D. student in the Department of Biomedical Engineering at the City University of Hong Kong. She focuses on studying the three-dimensional in vitro infectious microfluidic model and invasive cancer metastasis. |
 Haojun Hua | Haojun Hua is pursuing a Ph.D. in Artificial Intelligence at City University of Hong Kong, focusing on deep learning, image recognition, and medical image analysis. He has published several research papers and holds multiple patents in AI and microfluidics. |
 Wei Li | Wei Li is a Ph.D. student in the Department of Biomedical Engineering at the City University of Hong Kong. His research interests include microfluidics-assisted portable disease detection tools, biophotonics, and nanotechnology. |
 Bee Luan Khoo | Dr. Bee Luan Khoo is an Associate Professor of Biomedical Engineering at the City University of Hong Kong (CityU). Dr. Khoo got her Ph.D. from the National University of Singapore, working on tumor models for prognosis evaluation. As a Senior Postdoctoral Associate in the Singapore-MIT Alliance for Research and Technology, she developed microfluidic-based tools for rare cell detection. Dr. Khoo's research group now focuses on detecting, prognosis, and characterising disease heterogeneity using multidisciplinary techniques, including designing and utilising microfluidic devices for personalized disease management and evaluation. |
1. Introduction
Bacterial infections can affect various parts of the human body, causing multiple health issues, from common skin infections and cellulitis to more severe and invasive diseases such as tuberculosis (TB), sepsis, nephritis, and cystitis.1 The timely and accurate detection of bacteria is crucial for effectively treating and managing these conditions.2 It allows healthcare professionals to initiate appropriate treatment promptly, significantly improving patient outcomes. Any delay in detection can lead to severe complications or even life-threatening situations.3 Consequently, developing sensitive and rapid detection methods is essential for identifying bacterial presence and facilitating effective intervention.Bacteria-driven inflammation has been linked to the onset and advancement of diverse diseases. Bacterial infections can manifest as acute, life-threatening conditions or chronic, recurrent issues.4 Microfluidic platforms have been used to construct in vitro disease models that accurately replicate the intricate microenvironment that bacteria-driven inflammation affects. Beyond infectious diseases, bacterial infections are also implicated in the development and progression of specific cancer types.5 Inflammatory responses triggered by bacterial infections can contribute to a tumor-promoting microenvironment, influencing cancer initiation, progression, and metastasis.6 Understanding the complex interactions between bacterial infection and inflammation is critical to developing effective therapeutic strategies and disease models that can accurately simulate the microenvironment affected by bacteria-driven inflammation.
Microfluidic technology has emerged as a powerful tool in detecting and studying bacterial infections, offering several advantages over conventional methods.7 These platforms enable high-sensitivity detection with rapid analysis times and require only small sample volumes, making them particularly useful in clinical settings. Traditional bacterial identification techniques often need extended processing times and high pathogen concentrations, which can result in delayed diagnosis and missed treatment opportunities.8 In contrast, microfluidic platforms have demonstrated the ability to rapidly identify bacteria at lower biomass thresholds, ensuring prompt detection and enabling timely treatment.
This review aims to discuss the recent advancements in bacterial detection using microfluidics and explore the potential of these platforms as a versatile tool for studying bacteria-driven inflammation as an underlying complication for various physiological and pathological conditions, such as cancer. By integrating knowledge from microbiology, oncology, and engineering, this review will highlight how microfluidics can contribute to the early diagnosis of bacterial infections and the development of innovative cancer models, ultimately leading to more personalized and effective therapeutic approaches.
2. Challenges and innovations in bacterial detection
2.1 Conventional techniques
The timely and accurate detection and identification of bacterial strains have become critically important in various fields, including clinical diagnostics, food safety, and environmental monitoring.9,10 In these domains, rapid and precise bacterial detection is essential for preventing the spread of infections, ensuring food quality, and assessing the impact of environmental changes.11 However, traditional bacterial detection methods often cannot meet the demands of rapid and efficient detection processes,12 with some protocols taking up to 7 or 8 days to yield conclusive results.13Conventional pathogen detection techniques typically involve culture and colony counting, polymerase chain reaction (PCR), and immunology-based assays. PCR is widely used for DNA analysis, enabling the detection of specific bacterial genes with high sensitivity.14 Culture and colony-counting methods allow for the enumeration of bacteria by growing them on selective media, providing quantitative insights into bacterial populations.15 Immunology-based techniques, which rely on antigen–antibody interactions, are employed to identify bacterial antigens, offering specificity in pathogen detection.16
While these approaches are well-established and known for their accuracy, they have limitations. The time required for bacterial cultures to grow, the complexity of PCR protocols, and the need for specialized equipment and reagents all contribute to the prolonged analysis times.14 Additionally, implementing these methods often necessitates highly skilled personnel and sophisticated laboratory infrastructure, further complicating their widespread application in urgent or resource-limited settings.
In light of these challenges, there is a growing demand for more efficient bacterial detection methods to deliver rapid, accurate, and accessible results across various applications. This review will explore emerging technologies and methodologies that address these limitations, highlighting their potential to revolutionize bacterial detection and identification practices in critical sectors.
Culture and plating methods have long been the gold standard for bacterial detection. These techniques involve growing bacteria on selective media, allowing for their enumeration and identification. Despite their reliability and widespread use, culture methods are often time-consuming, typically requiring several days to more than a week to produce definitive results.17 This delay can be a significant drawback, especially in clinical settings where timely diagnosis is critical for effective treatment.
Given the limitations of culture-based methods, there is a pressing need to explore and implement alternative bacterial detection techniques to provide quicker and more efficient results.18 PCR has emerged as an alternative, offering the potential for rapid and precise bacterial identification, thereby addressing the urgency of timely diagnostics in various applications. PCR is a widely adopted nucleic acid amplification technology in bacterial detection. It allows for the isolation, amplification, and quantification of specific DNA sequences, including those unique to target bacterial species. PCR offers significant advantages over traditional methods, such as culture and plating, primarily due to its ability to deliver results in a much shorter time frame.19
Immunoassays leverage the highly specific interaction between antibodies and antigens, making them versatile tools for detecting a broad range of targets. Several immunoassay techniques, including enzyme-linked immunoassay (EIA), enzyme-linked fluorescence assay (ELFA), enzyme-linked immunosorbent assay (ELISA), and immunomagnetic beads (IMB) have been employed for pathogen detection.20 Among these, ELISA stands out as the most widely used and well-established technique, serving as a foundational method that has inspired numerous biosensor applications.
ELISA's success lies in combining the specificity of antibody–antigen interactions with the sensitivity of enzymatic assays. This is achieved by using antibodies or antigens conjugated to enzymes that produce easily detectable signals, making ELISA a cornerstone in immunoassay techniques.21
2.2 Emerging microfluidic biosensor technologies for the detection of pathogenic bacteria
Emerging microfluidic-based detection technologies present compelling alternatives to traditional bacterial detection methods. We summarized recent publications, particularly those from this year, focusing on innovative bacterial detection methods that address pressing challenges. The study's selection criteria focused on approaches relevant to rapid, on-site, real-time, and multiplex bacterial analysis using microfluidic technologies. The scope of the review primarily encompasses the detection of common disease-causing bacteria. To ensure a comprehensive overview, we included a diverse range of detection methods based on various principles of microfluidic systems, as well as those capable of detecting multiple bacterial species. Papers that did not focus on microfluidic-based detection methods or were not directly related to bacterial detection were excluded, as they did not align with the scope of this review. We classify the detection methods into four parts based on different bacterial detection principles and introduce them in the following parts, including detection of microfluidic systems for specific identification of gene and protein biomarkers, physical property detection and separation of microfluidic systems, optical detection methods of microfluidic biosensors, and microfluidic electrochemical sensing technology. | ||
| Fig. 1 A) A schematic diagram of the diagnostic process for detecting and identifying periprosthetic joint infection-associated bacteria performed on an integrated microfluidic system. The image is reproduced from a ref. 22 licensed by CC BY 3.0. B) A rapid on-site method for detecting viable E. coli O157:H7, using immunomagnetic separation combined with propidium monoazide and loop-mediated isothermal amplification and nucleic acid lateral flow strip. The image is reproduced from ref. 23 with permission from Elsevier, copyright 2022. C) Valveless microfluidic device for E. coli detection. The image is reproduced from a ref. 24 licensed by CC BY 3.0. D) Representation of the experimental procedure for droplet production and magnetoresistive measurement employed in this work. The image is reproduced from ref. 25 with permission from Elsevier, copyright 2024. | ||
2.2.1.1 Antimicrobial peptides. Antimicrobial peptides (AMPs), short peptide fragments found in a wide range of organisms, play a critical role in the innate immune system, providing essential protection against pathogens.26 These peptides are easy to synthesize and cost-effective and exhibit remarkable stability under extreme conditions, making them versatile tools in pathogen detection. Moreover, AMPs are derived from diverse sources, further enhancing their utility. In a pioneering study, Kim et al. introduced the first portable dual antibiotic-conjugated graphene micropatterned field-effect transistor (ABX-GMFET) integrated with a microfluidic chip. This innovative system facilitates on-site detection of Gram-positive and Gram-negative bacteria in various sample types, with a 10 CFU mL−1 detection limit. The system exploits charge and chemical interactions between the antibiotic probes (vancomycin and magainin I) and the target bacteria.27 A significant challenge in combating antibiotic resistance is the need for timely, on-site clinical data regarding bacterial identification, particularly for Gram-positive and Gram-negative strains. The ABX-GMFET platform addresses this challenge by providing a rapid diagnostic tool to mitigate the risks associated with antibiotic misuse, often leading to untreatable infections. The platform integrates a recognition probe with a transistor within a microfluidic device, minimizing human intervention and enabling streamlined automation, thereby offering a practical solution for improving clinical decision-making in real-time.
2.2.1.2 Antibodies. Antibodies play a vital role in identifying and eradicating pathogens with ease of use and high affinity for target antigens. Antibodies are integrated with microfluidic biosensors to facilitate bacterial detection. Wang et al. developed an integrated microfluidic system for the automatic detection of live Mycobacterium tuberculosis (MTB) and differentiation from dead bacteria. The slow growth rate of MTB sets it apart from other infectious bacteria, requiring at least 6–8 weeks for detection and complicating early diagnosis. This system captures bacteria within 10 minutes using a heparin-conjugated hemagglutinin antibody. To distinguish dead bacteria, a light-reactive dye, propidium monoazide, binds selectively to the double-stranded DNA of deceased bacteria within 20 minutes, preventing gene amplification during subsequent PCR steps. This study represents the first automated bacterial detection process for viable MTB on a single chip, providing results within 90 minutes and achieving a detection limit as low as 100 CFU.28 This advancement holds significant promise for on-site diagnosis and improving the efficiency of TB detection. The chip demonstrates the capability to deliver rapid and precise TB diagnoses, enabling potential adjustments in antibiotic dosages. While this innovative system demonstrates adaptability to various sample conditions and enhances diagnostic specificity, additional research and validation are necessary before clinical translation can be successfully achieved. Additionally, Wen et al. employed immunomagnetic separation technology to enhance the concentration of E. coli O157:H7 through magnetic enrichment (Fig. 1B). This was followed by a modified propidium monoazide method in conjunction with loop-mediated isothermal amplification to amplify the target gene. The presence of viable E. coli O157:H7 was then detected using the nucleic acid lateral flow test strip method.23 This method eliminates pre-culturing and delivers results within a mere 2 hour. Operating without specialized equipment, it relies solely on a standard water bath to maintain consistent temperature. This simplicity renders it highly convenient, particularly in resource-constrained settings. While this method effectively eliminates false positives resulting from dead bacteria, it is important to note that the detection sensitivity is lower than the standard culture method.
2.2.1.3 Phages. Phages can remain active under various extreme conditions (such as high temperatures and organic solvents), and phages can be easily and cheaply mass-produced. Usually, a large number of progeny phages can be obtained by simply infecting a bacterial solution.29 Alonzo et al. developed a microfluidic device and portable instrument prototype to enable rapid and highly sensitive phage-based assays for detecting E. coli cells (Fig. 1C). This advanced microfluidic system integrates membrane filtration and selective enrichment using T7-NanoLuc-CBM, a genetically engineered phage. It successfully detects as few as 4.1 CFU of E. coli in a 100 μL sample within just 5.5 hours.24 This innovation represents a significant step toward creating a faster, portable, semi-automated phage-based microfluidic platform for bacterial detection. In numerous remote low- and middle-income regions, the surveillance of fecal contamination in drinking water is constrained by inadequate facilities, financial constraints, and staff shortages. Introducing on-site, cost-effective, straightforward, precise, and swift assays for detecting E. coli contamination could empower community-driven monitoring efforts, promote behavioral change, and enhance emergency readiness measures. Future optimization endeavors will enhance test throughput and instrument automation, enabling prompt monitoring and user-friendly operation. Bloodstream infections are instigated by pathogens infiltrating the bloodstream, culminating in conditions such as sepsis and septic shock. Costa et al. developed an innovative microfluidic assay that combines a bead-based chip with a phage receptor-binding protein to serve as a highly specific and sensitive recognition molecule. This integrated device enables direct detection of bacteria from whole blood samples. The assay facilitates bacterial enrichment and allows simultaneous multiplex detection of Pseudomonas aeruginosa and E. coli.30 The detection concept and device architecture can be extended to encompass other bacterial species or sample matrices for various biomedical applications and food safety purposes. However, due to the spatial limitations of the microcolumns in the chip and the potential for spectral crosstalk when using multiple fluorescent proteins, whether this method can achieve high-throughput screening needs further experimental confirmation. In another advancement, Nogueira et al. introduced a novel microfluidic system for diagnosing urinary tract infections caused by Klebsiella pneumoniae. Klebsiella pneumoniae has emerged as a prevalent pathogen in catheter-associated urinary tract infections and a form of nosocomial urinary tract infections. Alarmingly, over 50% of Klebsiella pneumoniae strains exhibit resistance to multiple antibiotics, posing a significant challenge in treatment. This system combines magnetoresistive sensors with droplet microfluidics, offering a unique diagnostic approach.25 The system encapsulates Klebsiella pneumoniae suspensions in droplets containing magnetic nanoparticles (MNPs) modified with Klebsiella pneumoniae-specific phage receptor-binding proteins. Magnetically labeled cells within the droplets are detected using an MR readout system (Fig. 1D). MNPs have the advantages of large specific surface area, small detection steric hindrance, and uniform distribution.31 This method allows precise control over the number of MNPs and bacteria in each droplet, facilitating clear differentiation between Klebsiella pneumoniae and non-target E. coli samples. The system demonstrates a dynamic detection range for K. pneumoniae from 104 to 108 CFU mL−1.25 Leveraging droplet microfluidics effectively tackles the issues linked to MNP aggregation encountered during batch sample processing. By ensuring the uniform distribution of MNPs and bacteria in each droplet, a direct and highly reproducible correlation between the target species' concentration and the magnetic peak count was established.
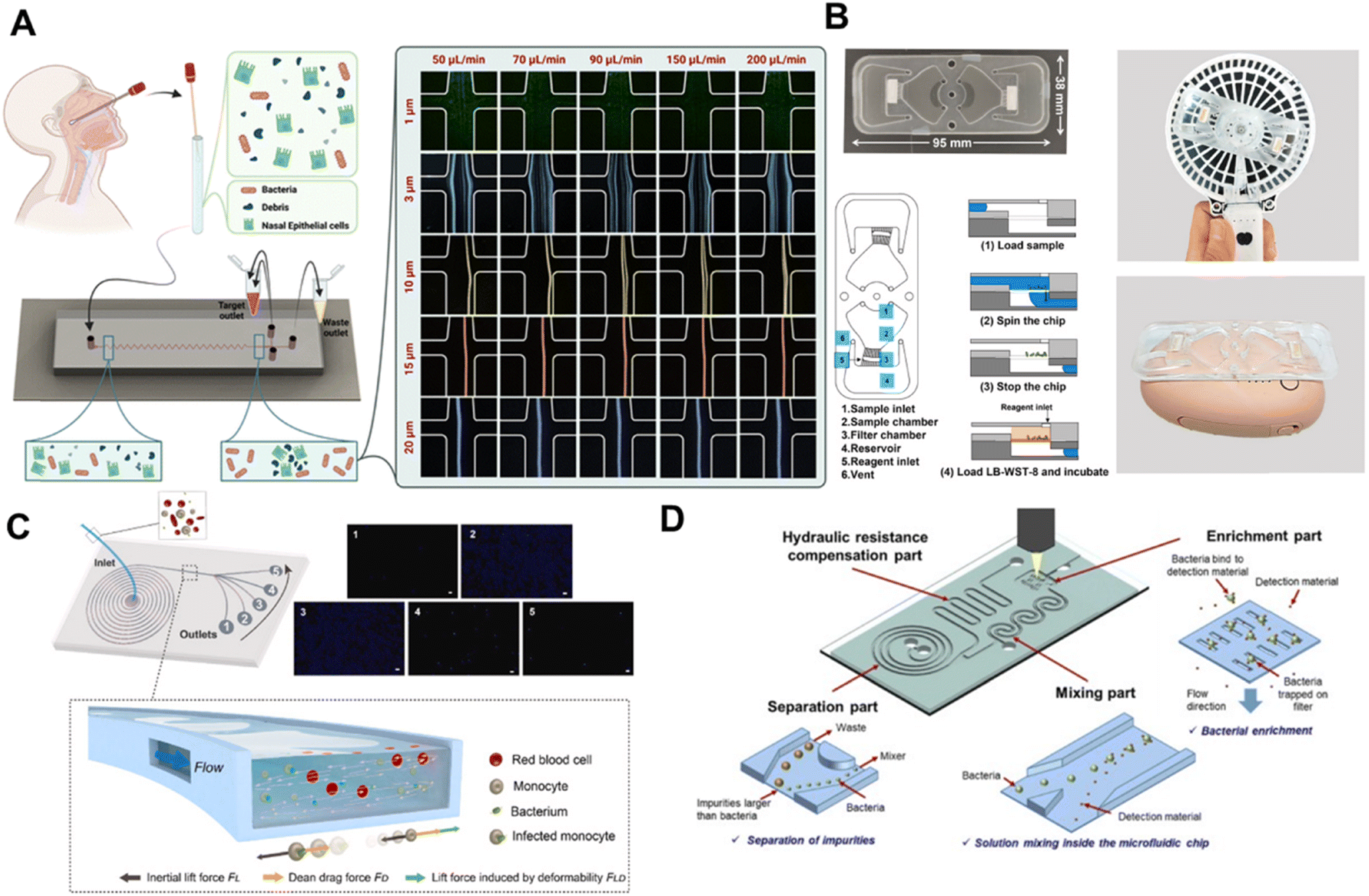 | ||
| Fig. 2 A) Rapid separation of bacteria from primary nasal samples using inertial microfluidics. The image is reproduced from ref. 32 with permission from the Royal Society of Chemistry B) detection of live bacteria using nylon filter membrane-integrated centrifugal microfluidics. The image is reproduced from ref. 33 with permission from Elsevier, copyright 2023. C) Label-free microchip to detect monocytes with internalized pathogens. The image is reproduced from ref. 34 with permission from Elsevier, copyright 2021. D) Microfluidic chip with integrated separation, mixing, and concentration operations for rapid and sensitive bacterial detection utilizing synthetic inorganic antibodies. The image is reproduced from ref. 35 with permission from Elsevier, copyright 2024. | ||
In a recent study, Chang et al. presented a centrifugal microfluidic device integrated with a nylon membrane to efficiently detect viable bacteria in water (Fig. 2B). Utilizing a handheld fan as the rotor and a rechargeable hand warmer as the heat source, this system achieves over 500-fold concentration of bacteria in water. Following incubation with water-soluble tetrazolium-8, color changes on the nylon membrane can be visually assessed or captured with a smartphone camera. The process is completed within 3 hours, with a detection limit of 102 CFU mL−1 and a range extending from 102 to 105 CFU mL−1.33 The platform boasts cost-effectiveness, user-friendliness, high sensitivity, and data reliability. However, the device currently takes 3 hours to complete the test, and future improvements to the culture medium are expected to speed up bacterial growth and reduce the detection time. This platform offers a convenient and sensitive approach for swift water quality monitoring, proving especially invaluable for nations with limited resources.
Liu et al. developed a portable centrifugal microfluidic system that uses motors to rotate magnetic beads for bacterial cell lysis, incorporating loop-mediated isothermal amplification on the same chip. This system allows for the rapid detection of five pathogenic bacteria in water within 70 minutes, achieving a detection limit of approximately 104–105 CFU mL−1.36 The chip is a compact, all-in-one, fully automatic portable analyzer with integrated injection and output functionalities. Michael et al. designed a fingertip spinner with centrifugal microfluidic technology to concentrate and filter urine samples on a nitrocellulose membrane. The sample is then reacted with WST-8 reagent, resulting in color development within 45 minutes. The detection limit for this system is approximately 103 CFU mL−1.37 The issue of slow and inconsistent rotation speeds is mitigated through fluid-assisted separation technology, effectively minimizing overall hydrodynamic drag. The device is designed to be cost-effective, sensitive, specific, rapid, durable, equipment-free, and easily deliverable to those requiring it. Nonetheless, the current proof-of-concept study solely focused on Escherichia coli in urinary tract infections, warranting future testing on additional bacterial species like Klebsiella pneumoniae or Proteus mirabilis for comprehensive validation.
Phagocytes are crucial in identifying and engulfing invading microorganisms to eliminate them. Nevertheless, bacterial pathogens can evade detection by remaining at low levels or replicating inside cells post-engulfment by immune cells, thereby escaping conventional detection methods. Liao et al. demonstrated that phagocytic blood cells undergo physical properties changes when containing intracellular bacteria. Specifically, human monocytes infected with various bacterial species exhibited increased stiffness and size compared to uninfected monocytes. Utilizing these mechanical differences, they developed a microfluidic biosensor based on inertial focusing (Fig. 2C). This biosensor allows for rapid and differential detection of infected monocytes from a large population of uninfected cells.34 The device can deliver a reading within 1.5 hours, requiring only a small blood sample of approximately 1 ml.
Furthermore, Liao et al. have pioneered a microfluidics-based label-free multivariate analysis platform comprising a biofilm enrichment device and a severity detection device. This innovative system enables simultaneous real-time biofilm enrichment and viscosity-based detection of biofilm-associated infection severity.38 Biofilm-associated infections are chronic and challenging to diagnose, often resistant to traditional antibiotic treatments. The biosensor's outcomes remain consistent for biofilms formed by Gram-negative and Gram-positive bacteria, including biofilm-promoting strains, across diverse mucus viscosities. Nam et al. introduced a microfluidic chip that integrates bacterial separation, mixing, and concentration through fluid dynamics alone (Fig. 2D). The chip achieves size-based bacterial separation by leveraging spiral fluidic channels using Dean flow and inertial focusing. Incorporating polydimethylsiloxane (PDMS) membrane filters within the chip enhances bacterial concentration, making the process highly efficient. Bacterial detection was assessed utilizing synthetic inorganic and commercially accessible fluorescent antibodies.35 This microfluidic chip demonstrates rapid and highly sensitive bacteria detection in food, human blood, and urine, offering significant value for effective prevention and treatment strategies. This unified chip architecture integrates multiple functionalities, offering a comprehensive solution for bacterial analysis. The high-throughput characteristics of this method have not been extensively discussed, suggesting that further enhancements may be necessary to optimize its performance in high-throughput screening applications.
 | ||
| Fig. 3 A) Single Escherichia coli bacteria detection using a chemiluminescence digital microwell array chip. The image is reproduced from ref. 40 with permission from Elsevier, copyright 2022. B) Schematic illustration of the multifunctional PEC biosensor based on PDA/MnO2/SA31/GCE for detection and inactivation of bacteria. The image is reproduced from ref. 41 with permission from Elsevier, copyright 2024. C) Raman spectrometer-read CRISPR/Cas biosensor for nucleic acids detection of pathogenic bacteria. The image is reproduced from ref. 42 with permission from Elsevier, copyright 2022. | ||
Additionally, it enables the differentiation between live and dead bacteria by monitoring bacterial proliferation and enzyme expression. This innovative technology promises rapid and reliable detection of E. coli, with potential applications spanning microbiology, environmental monitoring, and clinical diagnostics. The occurrence of hospital-acquired infections involving antibiotic-resistant bacteria has seen a significant surge. These infections lead to prolonged hospital stays and heightened costs and contribute to increased medical complexity. Wang et al. developed a microfluidic sensor capable of detecting three common bacterial species: Acinetobacter baumannii, Escherichia coli, and multidrug-resistant S. aureus. This sensor utilizes a biotin-labeled aptamer immobilized on a nitrocellulose membrane within the chip. Following the binding of the aptamer to the bacteria, a color reaction is induced using tetramethylbenzidine and streptavidin, which produces a blue colour upon binding secondary aptamers to the primary ones. This colorimetric change facilitates the detection of the target bacteria.43 The chip employs highly specific aptamers to detect target bacterial species simultaneously within a mere 35 minute timeframe. Operating without an external power source, it offers the benefits of quicker detection times and multiplexing capabilities compared to antibody-based assays. While this microfluidic system holds promise as a powerful tool for diagnosing various nosocomial bacterial strains in high-risk environments like intensive care units, additional clinical sample testing is necessary to ascertain its suitability for clinical applications. Baltekin et al. investigated the feasibility of developing a rapid susceptibility test for urinary tract infections. Using a specially designed microfluidic chip, this approach directly captures bacteria from samples with low bacterial counts. The growth rates of individual bacterial cells are monitored through microscopy to evaluate their response to antibiotic treatment within a short time frame.44 This method enables the determination of antibiotic susceptibility in less than 30 minutes. The system's rapid turnaround and high sensitivity and specificity make it a crucial tool in combating antimicrobial resistance by facilitating the appropriate use of antimicrobials in primary care settings, where patients typically seek initial treatment.
Developed and commercialized by Sysmex Astrego, the system is user-friendly. Healthcare professionals can place the sample onto the cartridge and insert it into the analyzer to initiate automated measurements, eliminating the need for specialized laboratory expertise. In another study, Cui et al. introduced a multifunctional photoelectrochemical biosensor powered by near-infrared light to detect and inactivate S. aureus (Fig. 3B). A carboxyl-functionalized SA31 aptamer is covalently bonded to a PDA/MnO photoelectrode through an amine-carboxyl linkage. The SA31 aptamer selectively captures S. aureus, which induces steric hindrance and produces a diminished photocurrent signal. This change in photocurrent is linearly related to the logarithm of the bacterial concentration, with a wide detection range from 10 to 107 CFU mL−1 and a low detection limit of 2.0 CFU mL−1. Furthermore, the photocurrent signal is restored upon the desorption of inactivated S. aureus from the photoelectrode.41 The biosensor exhibited high sensitivity and demonstrated excellent antibacterial activity.
Surface-enhanced Raman spectroscopy (SERS) represents a potent analytical technique that relies on the intensified Raman scattering of molecules adsorbed on or near SERS-active surfaces, such as noble metal nanostructures. This method has found extensive application in the sensitive and selective detection of chemical and biological molecules. Zhuang et al. developed a microfluidic paper-based analysis device (μPAD) that combines recombinase polymerase amplification with SERS to achieve highly sensitive bacterial detection (Fig. 3C). In this system, the aggregation level of SERS nanoprobes correlates with the concentration of Salmonella typhimurium (S. typhi). The μPAD captures and quantifies S. typhi, with the results analyzed using a Raman spectrometer. The device demonstrates a detection limit of approximately 3–4 CFU mL−1 and covers a dynamic range from 1 to 108 CFU mL−1. It provides accurate sample testing within 45 minutes.42 This μPAD-based system offers a rapid and highly sensitive method for detecting S. typhi, with significant potential for applications in microbiology and diagnostic fields. Nonetheless, the system faces limitations due to the distinct steps involved in its operation. Future endeavors could explore integrating DNA extraction, amplification, and Cas12a trans-cleavage into a unified, one-step process, or enhancing automation to streamline operations and enable easier deployment in field settings. Artificial intelligence has reportedly been used to develop a machine-learning model that can identify bacteria based on recorded SERS spectra.45
Integrating microfluidic systems with fluorescence technology has markedly enhanced the sensitivity and efficiency of bacterial detection. Xing et al. developed a one-step, no-clean microfluidic biosensor platform that interfaces with smartphones, enabling the simultaneous detection of multiple foodborne bacterial targets through the detection of single-stranded DNA. This technology relies on fluorescence resonance energy transfer between graphene oxide, coated on the microfluidic chip, and fluorescent molecule-modified bacterial DNA. A smartphone-based fluorescence detector captures the fluorescence recovery process, facilitating real-time detection. Demonstrating optimal analytical performance, the platform can detect four bacterial species—Staphylococcus aureus, Listeria monocytogenes, Escherichia coli, and Salmonella enteritidis—within 5 minutes, with detection limits of 0.17, 0.18, 0.27, and 0.17 nM, respectively.46 The chip is portable and easy to store, though image analysis requires a computer. Future developments will optimise applications based on the established algorithm, likely enhancing its utility in various practical settings. In a separate study, Wan et al. explored the use of semiconducting polymers in photodynamic therapy, a process in which compounds produce reactive oxygen species when exposed to light in the presence of oxygen, effectively eliminating microorganisms. Their research focused on semiconducting polymers incorporating antibiotic units, which label specific bacteria via fluorescence imaging and eradicate them through photokilling47 selectively. This study presented a straightforward approach to creating bioconjugated probes for bacterial identification while establishing a robust platform for pathogen photokilling. The findings hold significant promise for various applications, including antibacterial treatments, bacterial diagnostics, and real-time image interpretation, thereby expanding the potential uses of semiconducting polymers in both clinical and research settings.
The aptamer sensor incorporates microfluidic channels and electrodes directly fabricated on cotton threads, enhanced with molybdenum disulfide nanosheets to boost sensitivity. With a detection limit of up to 5.74 CFU mL−1,49 the sensor offers advantages such as cost-effectiveness, heightened sensitivity, specificity, and accuracy, all while overcoming the constraints of many traditional microfluidic devices reliant on external pumps. Future advancements may concentrate on optimizing the pairing of bioreceptors with viable nanomaterials to enhance sensitivity, enable multiplex detection, and explore other enhancements. Hou et al. developed a method for the continuous flow isolation of Salmonella from large-volume samples using self-assembled magnetic nanoparticle chains for efficient bacterial capture within 1 hour. The detection is performed within a microfluidic chip with a thin-film Ag/AgCl reference electrode array. The quantity of target bacteria can be determined by measuring the resistance of the catalyst. The method achieves a detection limit of 10 CFU mL−1.50 Modifying the antibodies can broaden the capability to detect additional foodborne pathogens, consequently enhancing food safety measures.
Additionally, a recent study introduced a microfluidic-based electrochemical device for the rapid and sensitive detection of E. coli. This device facilitates bacterial growth under ambient atmospheric conditions without an incubator, allowing for simultaneous electrochemical monitoring. It features three embedded electrodes created using screen-printed carbon paste, with the working electrode reinforced by graphitized mesoporous carbon. Cyclic voltammetry responses were used to assess E. coli concentration and growth within the device. The system can specifically detect bacterial infections with a detection limit of 0.35 CFU mL−1.51 The developed sensing platform exhibits high sensitivity towards E. coli and reliability in assessing the resistance profiles and efficacy of various antibiotics.
By integrating various bacterial detection methods, researchers can leverage the complementary advantages of each technique. This approach addresses the limitations inherent in relying on a single detection method, thereby enhancing overall sensitivity and effectiveness. Combining different techniques mitigates individual methods' shortcomings and significantly improves the detection process.
2.3 Point-of-care testing for pathogen detection
Recent advancements in point-of-care testing (POCT) have significantly enhanced the speed and convenience of pathogen detection directly at the care site (Table 1). We have selected common epidemic pathogens as the focus of POCT research, which will help us to respond immediately and reduce the medical burden. Some of them are important pathogens of gastrointestinal tract infections or urinary tract infections, and respiratory tract infections, including some drug-resistant pathogens.| Pathogen bacteria | Method | Limit of detection | Time | Portable device | Ref. |
|---|---|---|---|---|---|
| E. coli O157:H7 | Color-photo | 50 CFU mL−1 | 20 min | Smartphone | 52 |
| S. typhimurium | Color-photo | 16 CFU mL−1 | 30 min | Smartphone | 53 |
| E. coli K12, S. typhimurium, and S. aureus | Fluorescence photo | 10 CFU mL−1 for E. coli and S. typhimurium and 100 CFU mL−1 for S. aureus | <10 min | Smartphone | 54 |
| E. coli O157:H7 | Digital result | 62![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 CFU mL−1 000 CFU mL−1 |
15 min | Personal glucose meters | 55 |
| Salmonella | Color intensity | 63 CFU mL−1 | 30 min | Smartphone | 56 |
| E. coli | Fluorescence photo | 240 CFU mL−1 | <25 min | Smartphone | 57 |
| P. aeruginosa, S. typhimurium, V. parahaemolyticus, V. vulnificus, S. iniae and V. alginolyticus | Fluorescence signals | 200 cells per μL | — | Centrifugal microfludics | 58 |
| MTB DNA | Temperature | 39 nmol L−1 (or 0.58 μg mL−1) | Within 2 h | Thermometer | 59 |
| Streptococcus pneumoniae | Loop-mediated isothermal amplification | 20 fg μL−1 | — | Smartphone | 60 |
Rapidly screening foodborne pathogens is a critical challenge in preventing and managing gastrointestinal tract infections. Significant advancements have been made in developing simplified, rapid, and highly sensitive detection methods in recent years. One notable innovation is a microfluidic biosensor that utilizes gold nanoparticle aggregation and smartphone imaging to detect Escherichia coli O157:H7 (E. coli O157:H7) (Fig. 4A). This system exploits the unique optical properties of gold nanoparticles (AuNPs) to detect varying concentrations of the bacteria, with color changes monitored through a smartphone application.52 The biosensor demonstrates exceptional specificity and sensitivity, making it a promising tool for accurate pathogen detection. While the biosensor offers advantages such as simplicity, high integration, and cost-effectiveness, there is room for improvement in the response efficacy. One potential enhancement is using a smaller mixer to improve the efficiency and reduce the detection time. In a separate advancement, Guo et al. developed a portable biosensor that integrates magnetic nanoparticle-based immune separation, nanocluster signal amplification, and smartphone image analysis for the rapid and sensitive detection of Salmonella Typhi (S. typhi).53 This system leverages magnetic nanoparticles with smartphone-based image analysis techniques, significantly enhancing detection capabilities. The biosensor offers a good linear detection range and demonstrates considerable potential for further development, highlighting its promise as a portable, efficient, and highly sensitive diagnostic tool.
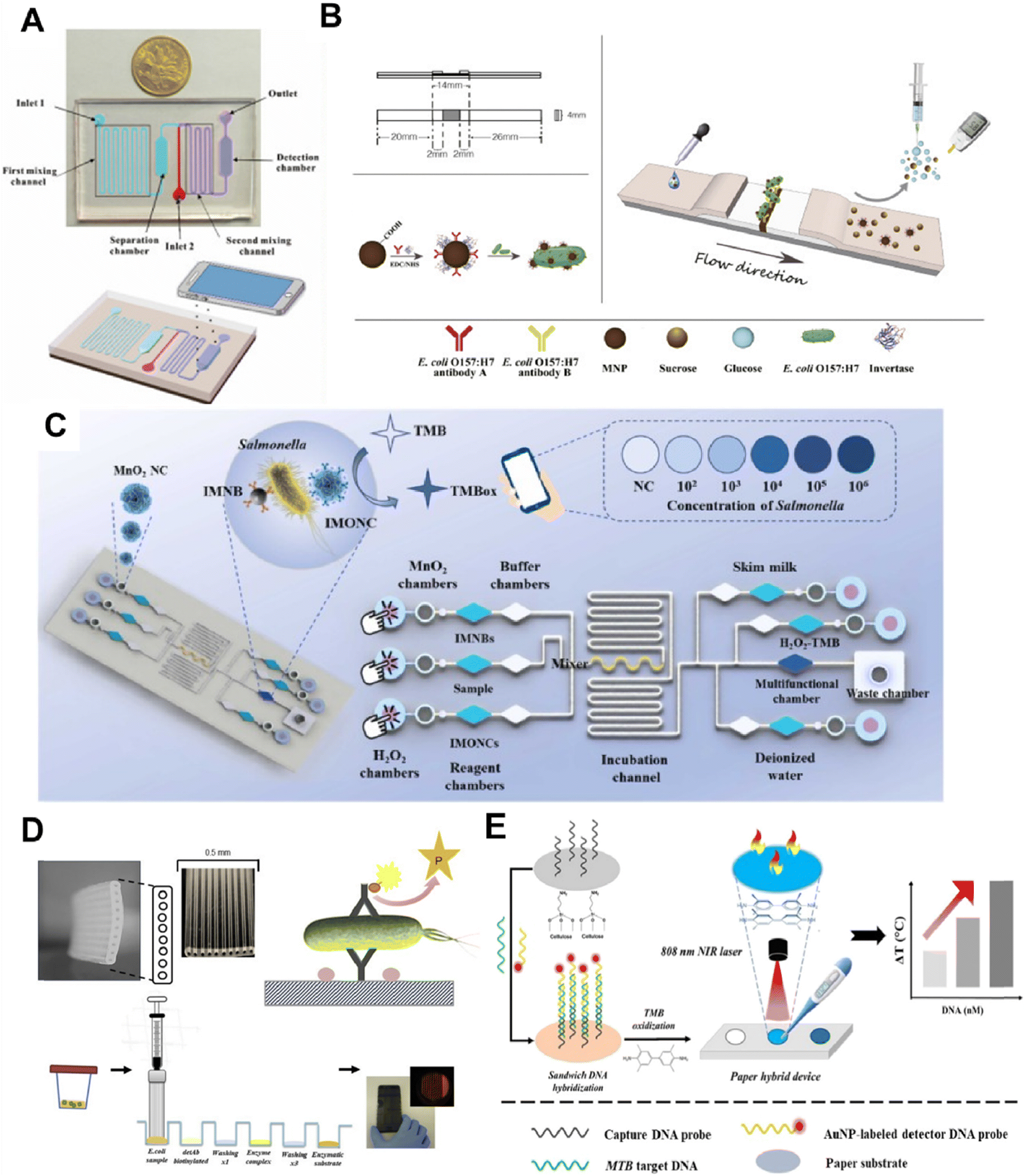 | ||
| Fig. 4 Method of POCT for pathogen bacteria detection. A) the proposed colourimetric biosensor for rapid detection of E. coli O157:H7 based on gold nanoparticle aggregation and smartphone imaging. The image is reproduced from ref. 52 with permission from Elsevier, copyright 2019. B) A portable and quantitative point-of-care monitoring system for E. coli O157:H7 using a personal glucose meter is achieved through an immunochromatographic assay. The image is reproduced from ref. 55 with permission from Elsevier, copyright 2018. C) Principle of the microfluidic biosensor for POCT of Salmonella. The image is reproduced from ref. 56 with permission from Elsevier, copyright 2023. D) The microcapillary platform was utilised to develop a quantitative fluorescent immunoassay for E. coli detection. The image is reproduced from ref. 57 with permission from Elsevier, copyright 2019. E) The working principle of the photothermal biosensing of MTB target DNA on a paper hybrid device. The image is reproduced from a ref. 59 licensed by CC BY 3.0. | ||
Kim et al. introduced a paper-based microfluidic chip utilizing protamine as a biological receptor for bacteria. Fluorescent particles conjugated with protamine are sequentially loaded into the chip and interact with bacterial membranes, leading to particle aggregation.54 Low-cost smartphone fluorescence microscopy is employed to image and quantify the bacterial presence. It is worth mentioning that the biosensor demonstrated excellent specificity even in the presence of environmental pollutants. A recent study introduced a portable quantitative immunochromatographic assay for detecting E. coli O157:H7 using a personal blood glucose meter as a readout (Fig. 4B)—carboxyl-coated Fe3O4 nanoparticles, conjugated with invertase and anti-E. coli antibodies produce glucose through sucrose hydrolysis by enzyme invertase, which is then detected by the glucose meter.55 This method offers a qualitative and quantitative diagnostic tool with practical clinical applications. The biosensor has the advantages of portability, low cost, and wide availability; however, further improvements in detection limits and data stability may be needed. Lastly, Wang et al. explored a novel application of bifunctional manganese dioxide nanoclusters in a power-free biosensor for Salmonella POCT (Fig. 4C). This biosensor, analyzed using a specialized smartphone application, quantitatively detects Salmonella at 63 CFU mL−1 within 30 minutes, all on a single microfluidic chip without additional equipment.56 This advancement represents a significant step in point-of-care testing for bacterial pathogens, providing a simple and efficient solution. The biosensor utilizes manganese dioxide nanoclusters to significantly improve the sensitivity of the sensor and greatly reduce the complexity of the operation.
Urinary tract infections are among the most common bacterial infections and pose a significant healthcare burden. The development and utilization of POCT enable swift diagnostics, potentially curbing antibiotic over-prescription and the rise of drug-resistant strains. A smartphone-based optical microfluidic fluorescent sandwich immunoassay is reported capable of quantifying E. coli in buffer and synthetic urine within 25 minutes (Fig. 4D). This system features a 10-well microfluidic strip with an open fluid design, allowing significant sample volumes to pass through antibody-coated microcapillaries. The device achieves a detection limit of 240 CFU mL−1 and holds the potential for rapid diagnosis of E. coli infections in urinary tract infections.57 This study leverages a microfluidic device coupled with a smartphone camera, offering simplicity, stability, and reproducibility. Future endeavors will involve validating with clinical samples and progressing towards power-free reagent loading. Zhang et al. presented a manual centrifugal microfluidic platform integrating nucleic acid purification, loop-mediated isothermal amplification, and fluorescent signal detection into a single disk.58 This platform enables the simultaneous detection of six pathogenic bacteria with high-speed rotation, making it suitable for nucleic acid detection in remote settings without electricity. It was verified to have good sensitivity and specificity for pathogens in artificial urine.
Tuberculosis is a potentially serious infectious disease that primarily affects the lungs. Zhou et al. developed an affordable photothermal biosensing method for quantifying MTB DNA using a thermometer integrated into a paper mixing device (Fig. 4E). DNA capture probes are immobilized on paper and DNA hybridization. The subsequent gold nanoparticles catalyze an oxidation reaction of 3,3′,5,5′-tetramethylbenzidine (TMB), serving as a photothermal agent under 808 nm laser irradiation for photothermal biosensing.59 This novel method offers a cost-effective and miniaturized solution for nucleic acid quantification, which is ideal for point-of-care settings. This photothermal biosensing platform has broad potential for application. Streptococcus pneumoniae is the most common pathogen associated with community-acquired pneumonia requiring hospitalization. Wang et al. introduced a multifunctional microfluidic chip platform with integrated loop-mediated isothermal amplification for rapid detection of Streptococcus pneumoniae.60 The portable device features a built-in heating module, a Bluetooth Wi-Fi camera module, and a dedicated smartphone application interface to communicate.
In the preceding section, we focused on exploring the potential of microfluidic chips for bacterial detection, highlighting their promising development prospects. The following section focuses on microfluidic models of bacteria-driven inflammatory infections. By identifying the pathogens responsible for these infections, we examine the complex inflammatory responses they trigger and the subsequent therapeutic approaches critical for addressing bacterial infections across various diseases. This comprehensive approach not only enhances readers' understanding of the diverse applications of microfluidic chips but also underscores the potential of microfluidics to advance our knowledge of inflammation and drive the development of innovative strategies for bacterial detection and treatment.
3. Microfluidic-based organ-on-a-chip models
Microorganisms trigger infections, while inflammation is a biological response of the organism to the presence of these pathogens.61 Pathogenic bacteria invade human tissues, causing infections. This infection, in turn, induces inflammation as the host's immune system reacts to the bacteria and their toxins to combat the infection. It is important to highlight that bacterial infections are not only a direct cause of illness but also a key driver of inflammation.62 Therefore, modeling inflammation in the context of bacterial infection is essential for understanding the full scope of the immune response and disease progression. Microfluidic-based organ-on-a-chip (OOC) models represent a significant advancement in disease modeling, offering a miniaturized, precise platform to replicate human tissue environments and study pathogen–host interactions. These models enable detailed analysis of cellular responses and infection mechanisms, providing more accurate and controlled systems for research and therapeutic development.3.1 Conventional methods for studying infectious diseases
Conventional methods for studying infectious diseases have long relied on in vitro cell cultures, animal models, and clinical observations to understand pathogen behavior and host responses.63In vitro cell cultures offer a controlled environment to investigate cellular interactions with pathogens, assess cytotoxicity, and evaluate potential therapeutic agents.64 These systems, however, often need to catch up in replicating the complex physiological conditions of human tissues and may need to capture the multifaceted nature of infection processes fully. Animal models have been instrumental in bridging this gap by providing a more holistic view of disease progression and immune responses.65,66 They allow for studying infection dynamics in a whole-organism context, offering insights into systemic effects and long-term outcomes. However, animal models have ethical considerations, variability, and limitations in translating findings directly to human health.67 Simultaneously, the U.S. Food and Drug Administration acknowledged the constraints of animal research by enacting the Modernization Act 2.0. This act abolished the mandate stipulating that all drugs must undergo testing in animal models.68Clinical observations and trials provide essential data on disease prevalence, progression, and treatment efficacy in human populations.69,70 Despite their invaluable contribution, these studies are often limited by variability in patient responses and the complexity of real-world conditions. These conventional methods have advanced our understanding of infectious diseases and sophisticated, controlled, human-relevant models. Recent innovations in microfluidics and OOC technologies are emerging as powerful tools to address these limitations, offering more precise and reproducible systems to study pathogen-host interactions and develop targeted interventions.71
3.2 Advances in microfluidics technology in bacteria infection disease model
Advances in microfluidics technology have significantly transformed the landscape of disease modeling.72 Developing sophisticated OOC systems has enabled researchers to recreate and investigate complex biological environments with unprecedented precision. These microfluidic devices integrate various cell types and extracellular matrices, offering detailed insights into bacterial interactions at critical sites. These innovations in microfluidics technology offer more accurate and physiologically relevant models for bacterial infection studies and pave the way for new therapeutic strategies to address complex diseases, ultimately advancing health and improving patient outcomes.73 | ||
| Fig. 5 A) Fetal membrane (FMi) and placenta (PLA) feto-maternal interface OOC design. The image is reproduced from a ref. 72 licensed by CC BY 4.0. B) The FMi-OOC device with the integrated media reservoir. The image is reproduced from ref. 75 with permission from the Royal Society of Chemistry. C) Schematic image of the VCD-OOC with different cell culture chambers represented by different colors and connected by an array of microchannels. The image is reproduced from ref. 76 with permission from John Wiley and Sons, copyright 2022. D) Schematic top (left) and cross-sectional views (right) of the dual channel microfluidic organ chip lined by human cervical epithelium interfaced across an ECM-coated porous membrane with human cervical fibroblasts. The image is reproduced from ref. 77, licensed by CC BY 4.0. | ||
Microfluidic OOC models provide critical insights into reproductive infections and inflammatory responses, advancing maternal–fetal health research. A study introduced a six-chamber vaginal-cervical-decidua (VCD)-OOC model to mimic the female reproductive tract during pregnancy. This model features compartments with vaginal epithelial cells, cervical epithelial and stromal cells, and decidua cells, replicating the reproductive environment76 (Fig. 5C). The cervix is the gateway to the fetal–maternal interface, represented by the decidua in the FMi-OOC. An ascending infection with Ureaplasma parvum (U. parvum) was induced in the VCD-OOC, leading to minimal inflammation. However, a combined infection with U. parvum and LPS resulted in significant inflammation in the VCD-OOC and FMi-OOC cells. The study highlighted the protective role of the cervical epithelial layer against ascending infections and emphasized the need to include immune cells and endocrine factors for a comprehensive understanding. These findings shed light on U. parvum's limited immunogenicity and its implications for preterm birth, underscoring the importance of studying pathogen interactions in ascending infections for reproductive research and drug testing in pregnancy-related infections. Advanced OOC models enhance understanding reproductive infections and inflammation, especially concerning preterm birth. A human cervix mucosa OOC model was designed to study cervical physiology and potential treatments for women's health. This model replicates the human cervix epithelium-stromal interface using a commercially available microfluidic chip featuring two microchannels with a porous membrane.77 Human primary cervical epithelial cells and stromal fibroblasts were cultured on opposite sides (Fig. 5D). The model was infected with healthy or dysbiotic microbial communities, showing differences in immune responses and barrier function. The study highlighted how Lactobacillus crispatus benefits cervical health compared to Gardnerella vaginalis. This model offers a relevant system for studying cervical physiology and host–microbiome interactions, aiding in developing therapeutic interventions and advancing women's health research.
The human placenta is essential in reproductive processes, acting as the primary interface for maternal–fetal nutrient exchange.78 However, studying human placental pathology is fraught with challenges due to the difficulties in accessing and analyzing placental tissues, which poses significant experimental limitations.79–81
A study developed a microengineering model of the human placental barrier on a chip to study inflammation responses to bacterial infection (E. coli).82 The device mimics the placental interface, housing co-cultures of trophoblast and endothelial cells (Fig. 6A). Through perfusion, it replicates circulatory flows for targeted inflammatory studies. Complex interactions include increased cytokine secretion post-infection by trophoblasts and maternal macrophage adhesion, revealing insights into fetal inflammatory response syndrome. This model mirrors clinical scenarios, offering an understanding of placental dysfunctions and preterm birth. Enhancements using pluripotent stem cell-derived placental cells could deepen insights into reproductive health. Microengineering models on chips study inflammatory responses in pregnancy components, advancing reproductive health understanding. A study created a model of the human amniotic membrane with human induced pluripotent stem cells (hiPSCs) on a chip to examine amniotic tissue's inflammatory response to bacteria.83 The device creates a conducive microenvironment for amniotic differentiation, showing functional impairment after E. coli exposure and mimicking clinical infection symptoms (Fig. 6B). This chip model provides insights into early pregnancy inflammation, offering potential in embryology and reproductive medicine. It effectively mirrors inflammatory responses to bacterial infections, uncovering issues like cell apoptosis and junction disruptions, aiding the study of pregnancy complications and drug responses. Microengineering models in reproductive medicine enable deep investigations into physiological interactions. A study focused on a placenta-on-a-chip model to examine the impact of Placental Malaria (PM) on fetal–maternal nutrient exchange.84 In the model, trophoblast and endothelial cells are co-cultured on opposite sides of an extracellular matrix gel in a microfluidic system, creating a barrier that mimics the maternal–fetal interface in placental villi (Fig. 6C). By simulating infected erythrocytes adhering to trophoblast cells via chondroitin sulfate A (CSA), characteristic of placental malaria, the model revealed how these cells impede glucose transport across the placental barrier. This insight enhances understanding of PM pathogenesis and supports the development of potential therapies. The study highlights the importance of fluid mechanics in maintaining vascular cell characteristics and facilitating nutrient exchange at the maternal–fetal interface.
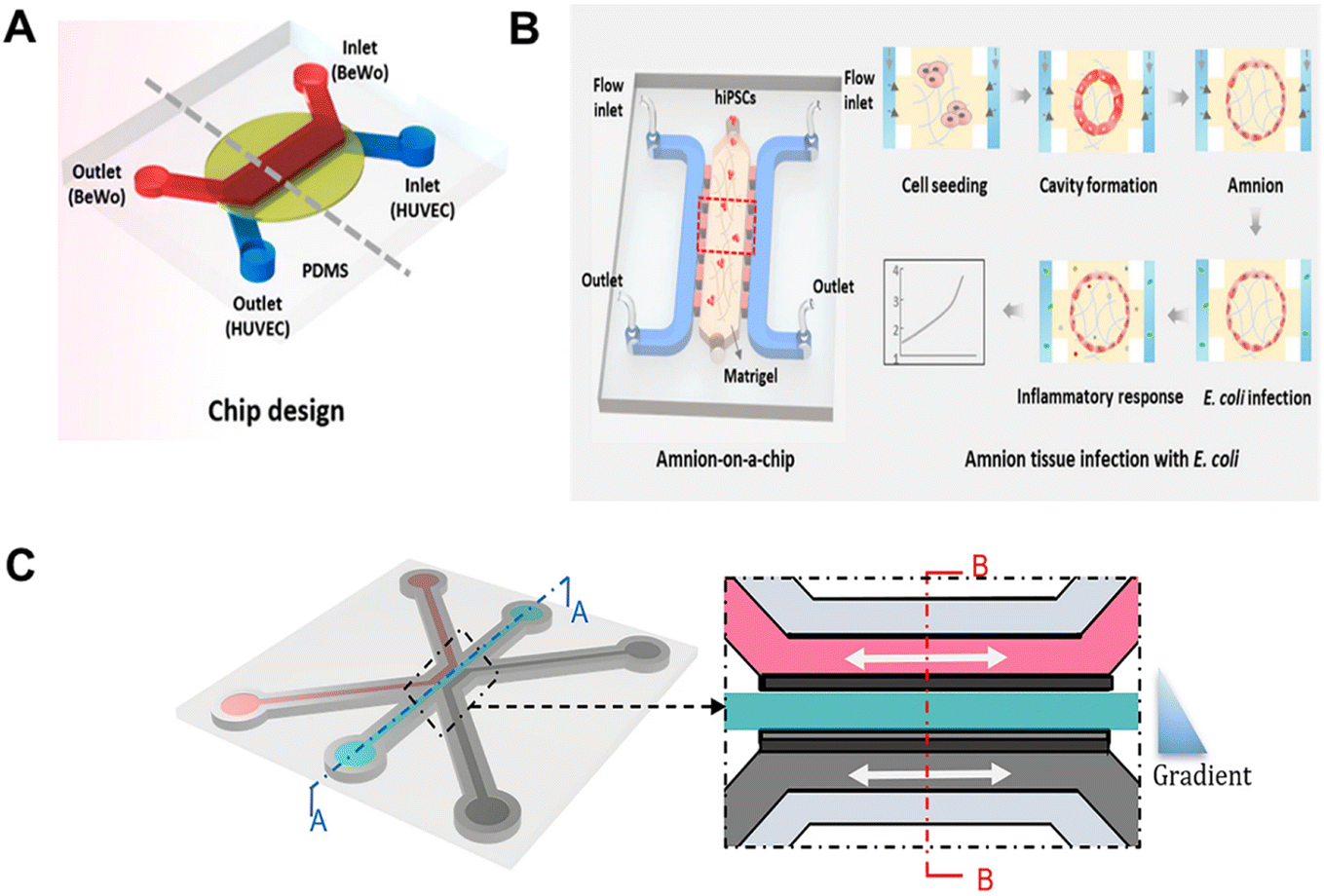 | ||
| Fig. 6 A) Design and assembly of the multilayered chip mimicking the human placental barrier in vitro. The image is reproduced from ref. 82 with permission from the American Chemical Society, copyright 2018. B) Schematic depiction of the amnion-on-a-chip device for investigating amniotic inflammatory responses under bacterial exposure. The image is reproduced from ref. 83 with permission from the American Chemical Society, copyright 2020. C) Overview of the method for modeling placenta malaria in the 3-lane OrganoPlate platform. The image is reproduced from a ref. 84 licensed by CC BY 4.0. | ||
 | ||
| Fig. 7 A) Conceptual diagram of the HuMiX model for the representative co-culture of human epithelial cells with gastrointestinal microbiota. The image is reproduced from a ref. 88 licensed by CC BY 4.0. B) Microsystem for gut-on-a-chip. Three-dimensional schematics of the device show the structure of individual components and the sealing method. The image is reproduced from a ref. 89 licensed by CC BY 4.0. C) Schematic of the fabrication process for 1-chamber and 2-chamber devices. The image is reproduced from ref. 90 with permission from John Wiley and Sons, copyright 2021. D) A gut-on-a-chip microfluidic device to mimic the human intestinal microenvironment undergoing injury and inflammation and to study interactions between cultured microbiome and human intestinal epithelial cells in an organ-like context.91 E) 3D structure diagram of the gut chip microdevice with relevant oxygen gradients. The zoomed-in area is a brightfield image of the cross-section of the chip. The image is reproduced from ref. 92 with permission from the Royal Society of Chemistry. F) Microfabrication of the EMI axis's human gut-on-a-chip and microfluidic cultures. The image is reproduced from ref. 93, licensed by CC BY 4.0. G) Analysis of co-cultivation of Caco-2 cells on a chip with specific anaerobic bacteria F. prausnitzii. The image is reproduced from ref. 94 with permission from the Royal Society of Chemistry. | ||
The intestinal environment supports epithelial cells aerobically and microbiota anaerobically. Existing experimental models struggle to recreate this dual oxygen environment without external anaerobic chambers. A novel microfluidic device was developed to co-culture intestinal epithelial and bacterial cells (Bifidobacterium adolescentis) in controlled, partitioned oxygen conditions.90 The device comprises two chambers separated by a porous membrane: an upper chamber and a lower chamber (Fig. 7C). Oxygen levels were validated using microsensors, Caco-2 cell immunostaining, and genetically encoded bacteria growth monitoring. This model offers precise oxygen control for cells/bacteria and a mathematical model for oxygen consumption rates. It simplifies studying oxygen dynamics in the gut and cell–bacteria interactions. Advanced microfluidic models have transformed biological research, particularly in studying complex physiological interactions. A human gut-on-a-chip microdevice investigated gut microbiota interactions, emphasizing gut health, inflammation, and cell–bacteria dynamics. The study employed this device to co-culture multiple symbiotic microorganisms with live human intestinal epithelial cells, assessing the impact of gut microbiota, inflammatory cells, and mechanical deformations on intestinal bacterial overgrowth and inflammation.91 The microfluidic device featured three parallel hollow microchannels, enabling the formation of villi-like structures that mimic in vivo intestinal morphology (Fig. 7D). Integrating various cell types and mechanical deformations revealed novel insights into gut inflammation, bacterial overgrowth, and barrier function. The findings underscore the potential of microfluidic gut-on-a-chip devices to model human gut diseases and enhance understanding of intestinal pathophysiology. Probiotics show promise in treating intestinal inflammation like inflammatory bowel disease (IBD).
Bifidobacterium bifidum (B. bifidum) supplementation demonstrates potential in managing and treating IBD, with a focus on understanding its mechanisms. The study proposes a cost-effective method using numerical simulations to design a dual-channel gut-on-a-chip model with controlled oxygen gradients.92 The microfluidic gut chip device features two parallel microchannels separated by a flexible porous membrane. The upper channel is supplied with a hypoxic culture medium, while the lower channel receives an aerobic medium, establishing relevant oxygen gradients within the device (Fig. 7E). In the study, IBD symptoms were induced in the chip to assess the effects of B. bifidum on the intestinal epithelial barrier. Results suggest that B. bifidum can enhance barrier stability, offering the potential to treat intestinal inflammation. This research showcases the potential of organ-on-a-chip models for understanding and treating conditions like IBD.
Advanced microphysiological models are crucial for studying human immune responses to gastrointestinal inflammation. Like the gut-on-a-chip device, these models mimic the complex interactions within the epithelial–microbiome–immune (EMI) axis under healthy and inflammatory conditions.93 The gut-on-a-chip microfluidic devices, fabricated using soft lithography, feature a central cell culture chamber with upper and lower microchannels and bilateral vacuum chambers assembled stepwise (Fig. 7F). These models offer insights into inflammatory factors' roles in immune activation and barrier dysfunction by integrating flow, mechanical stretching, and a physiological environment with gut microbiota and immune cells. Such microengineering immune models can potentially advance precision medicine through customizable, patient-specific chips.
Researchers have developed a microfluidic-based gut-on-a-chip model, the depression-on-gut-chip (DoGC), to explore depression and gut microbiota interactions in a controlled environment. This model integrates gut microbiota from individuals with depression, enabling the study of long-term interactions between human intestinal cells and gut bacteria.94 Initially, the model incorporated the anaerobic bacterium Faecalibacterium prausnitzii and established a preliminary co-culture with oxygen-consuming Caco-2 cells (Fig. 7G). By introducing representative gut microbiota linked to depression, the model mimics depression-related physiological characteristics like impaired intestinal barriers, chronic inflammation, and altered neurotransmitter levels. Metabolomic analysis showed changes indicative of depression. The DoGC is a robust tool for investigating the role of gut microbiota in depression, providing insights into its pathophysiology and supporting personalized medicine.
The technology of microfluidic platforms could present the multicellular three-dimensional (3D) cultures by the co-cultivation of different cell types and microorganisms in specific spatial distributions and proportions, resulting in organ models exhibiting structural and functional characteristics.97,98 Deng et al. designed a 3D cancer biofilm microfluidic device, known as the in vitro platform, to analyze the effects of intratumoral and extratumoral bacteria to investigate tumor progression influenced by bacterial interactions using uropathogenic Escherichia coli and bladder cancer cell clusters.99 This device explores the impact of intratumoral bacteria (IB) and extratumoral bacteria (EB) on cancer progression, focusing on the human bladder carcinoma cell line UMUC-3. Unlike conventional bacterial models such as flow chamber biofilm assays, this microfluidic design aims to replicate biofilms' in vivo 3D structure through a microwell array. The device has three layers, featuring four inlets for bacterial and cell culture in the top layer. This platform facilitates comparative analysis of bacterial patterns on cancer cell behaviour, revealing that IB and EB promote closer cell–cell contacts within tumour clusters, though only IB reduced cancer cell viability. The presence of EB was associated with the formation of extracellular DNA-based biofilms and a significant increase in cancer stem cell counts, which are linked to metastasis and poor patient outcomes. The findings underscore the potential of this 3D microfluidic device to advance our understanding of bacterial influences on tumor promotion and suggest avenues for future research into the efficacy of antibacterial agents in cancer treatment. (Fig. 8A) The typical microfluidic chips provide the potential insight for studying the pathogenesis of cancer with bacterial infection and for drug screening,100 which could be obvious and quick to understand the relationship between bacteria and cells. Various biological components could be involved in studying different interactions and reflecting the effectiveness of clinical and new potential drugs.101,102 Observing long-term growth and quantifying the penetration of multiple cancer therapies are conducted to evaluate the drug efficacy and mimic the cell responses under drug application in the microenvironment.103 Developing various microfluidic platforms could help build up another convenient pathway to discover and study cancer therapies in the future.104
 | ||
| Fig. 8 A) A schematic diagram of the microfluidic device consisted of three layers, with four inlets for bacteria culturing within the top layer. This image is reproduced from ref. 99 with permission from Elsevier, copyright 2021. B) Conceptual diagram of the microfluidic device to study the interaction between bacteria and colonocytes in the colorectal cancer microenvironment. This image is reproduced from a ref. 105 licensed by CC BY 3.0. C) Schematic diagram of the microfluidic platform for investigating bacterial chemotaxis mechanism. This image is reproduced from a ref. 106 licensed by CC BY 4.0. | ||
On the other hand, the carcinogenic mechanisms of gastrointestinal (GI) tract microbiota in colorectal cancer (CRC) have been extensively studied;107 however, there is a notable scarcity of platforms that effectively mimic the human colorectal microenvironment for in-depth investigations of pathogenic bacteria and colorectal microbiota interactions. Traditional in vitro co-culture assays often need to restore the complexity of bacteria–colonocyte interactions seen in vivo accurately.108 This leads to an underestimation of the role of pathogens and microbial interactions in colorectal carcinogenesis.109
A newly designed co-culture microfluidic device featuring colonocyte spheroids and colorectal microbiota has been developed to address existing gaps in the colorectal cancer (CRC)105 study. This advanced microfluidic model consists of four stacked layers, each serving a distinct function in the co-culture process. By more accurately replicating the in vivo tumor microenvironment, the device enables the compartmentalization of various cellular and microbial components, facilitating the study of complex interactions between colonocytes and microbiota in the context of CRC. The model effectively supports CRC tumor formation and associated alterations, as evidenced by the expression of cancer-related colonocyte genes, analyzed within a three-dimensional matrigel scaffold. Proof-of-concept experiments demonstrate the model's utility in studying CRC, with assessments of cell proliferation and epithelial-to-mesenchymal transition (EMT) conducted through viability tests and quantitative PCR. Further investigations incorporating pathogenic bacteria, such as Fusobacterium nucleatum (Fn), under various culturing conditions, including aerobic and anaerobic environments, reveal that these bacteria can activate inflammatory and mitogenic transcriptional programs in colonocytes. This model offers significant potential for advancing our understanding of CRC pathogenesis and the role of microbiota in tumour development.
This co-culture microfluidic model presents a promising platform for exploring novel microbial interactions within the colorectal cancer (CRC) microenvironment and elucidating the complex relationships between colonocytes and microbiota (Fig. 8B). The CRC co-culturing microfluidic chip shares several design features with the model developed by Deng et al., such as 3D multilayer structures and the simulation of interactions between cancer cells and bacteria within the tumor microenvironment.99 However, the CRC model described here specifically focuses on investigating the infectious interactions between CRC cells and pathogenic bacteria. In contrast, the model designed by Deng et al. not only studies the biological aspects of the tumor microenvironment but also incorporates multi-drug screening, providing insights into the effectiveness of clinical drug therapies.
In addition to replicating the bacterial-tumor microenvironment in a three-dimensional context for deeper investigation, microfluidic devices offer valuable insights into existing therapeutic shortcomings and facilitate the development of novel cancer treatments.110 Bacterial-mediated cancer therapy, a newly emerging technique, leverages bacterial strains' unique properties and mechanisms to target tumors specifically. Researchers have designed and fabricated a microfluidic platform to illustrate the chemotaxis mechanism underlying bacterial targeting of cancer cells.106 This device is constructed to enable the simultaneous three-dimensional co-culture of various cell lines within separate chambers, allowing the study of interactions between different cell types in a controlled environment. By simulating chemotactic gradients between cancer cells and bacteria, the platform establishes consistent concentration gradients of biochemical compounds in a central channel through diffusion via microchannels.
Additionally, fluorescence intensity from green fluorescent protein-encoding bacteria allows for real-time tracking of bacterial movements and behaviors in response to these chemotactic gradients. This approach is powerful for discovering lung cancer diagnostic markers and identifying specific biochemical molecules' roles in preferential bacterial chemotaxis. The multi-chambered microfluidic device provides critical insights into bacteria–cell interactions and potential therapeutic strategies, offering a reproducible and controlled platform for advancing cancer treatment research (Fig. 8C). This platform mimics directions differently than the other two microfluidic chips mentioned before. It mainly describes the precise control over various parameters that affect the changing of the human microenvironment, which closely resembles in vivo conditions for the human body. The design of this model is to observe and analyse the relationships between cells and bacteria in cancer progressions through biological and chemical aspects. Accurately modelling organ physiological functions and behaviours provides another direction for future in vitro research in cancer progression studies. The microfluidic platforms could also involve multiple cell types co-culturing with bacterial infection under a stable chemical gradient, i.e., chemotactic preferential migration of bacteria on different cell types.111,112 The bacterial effect of targeting and changing tumour phenotypes is studied for clinical therapy's actual mechanism. The controlled ideal environment created by the microfluidic chip could ideally investigate the chemotactic behaviour of bacteria and cancer cells.
Compared to traditional two-dimensional cell cultures and animal models, organ-on-a-chip systems could mimic the realistic human microenvironment in a more precise parameter.113,114 Different environmental parameters could control the chips, offering valuable insights into cellular behaviour and interactions with other components through numerous signal pathways.115 Various biological and physiological phenomena could be presented and accurately reflect the actual effects of drugs on the human body.116 This type of microfluidic platform facilitates the identification of suitable drug candidates with potential therapeutic benefits and fewer side effects. Therefore, the microfluidic platform is a possible tool for studying abundant detailed physiological interactions and could even closely reflect clinical phenomena in the following development.
In contrast, organ-on-a-chip models present a complementary approach devoid of these constraints. Sharma et al. present a bladder-on-a-chip model that co-cultivates a well-characterized human bladder epithelial cell line with bladder microvascular endothelial cells in a device structure, enabling both cell types to encounter urine and nutrient media, respectively. Their findings reveal that neutrophil migration into infection sites on the epithelial side triggers neutrophil extracellular trap formation, while intracellular flora protects bacteria against antibiotic clearance.120 Strategies targeting the eradication of intracellular flora are thus pivotal for therapeutic effectiveness.
Pneumonia stands as the most severe inflammatory occurrence in the lower respiratory tract. Acute respiratory diseases pose a significant medical challenge, especially in immunocompromised individuals.121 Pneumonia can stem from various microorganisms, with S. aureus being recognized as a primary cause of severe invasive infections.122 Studies investigating microbial lung infections have heavily leaned on animal models like mice. Nonetheless, notable variations exist between mice and humans in lung anatomy, physiology, and immune responses.123 Deinhardt-Emmer et al. developed a human alveolar model featuring an immune-responsive cell interface linking the vascular endothelium with the epithelial layer exposed to the air. Their research showed that flow conditions and the presence of macrophages enhanced barrier function, and disruption of this barrier led to significant endothelial cell harm. This model facilitated the observation of the spatial and temporal dissemination of S. aureus, enabling the characterization of infection-induced structural and functional changes.124 The alveolar chip provides a promising platform for delving into the mechanisms governing host–pathogen interactions and pinpointing molecular and cellular targets for novel therapeutic approaches in pneumonia treatment. In another study, an alveolar chip was utilized to investigate the direct impact of lung surfactant during the initial stages of infection. Thacker et al. engineered a model of early TB infection incorporating channels to mimic air and blood circulation. The absence of surfactant resulted in the swift and unregulated proliferation of bacteria within macrophages and alveolar epithelial cells.125 These discoveries offer insights into why individuals such as smokers and the elderly, who have compromised surfactant function, face an elevated risk of developing active TB.
The highly perfused liver plays a critical role in filtering blood-borne infections. About 80% of tissue-resident macrophages, known as Kupffer cells, reside in the liver.126 Positioned in the liver sinusoids, Kupffer cells are at the forefront of encountering pathogens that enter the liver, actively engaging in the early immune response against these invaders.127 Rennert et al. established an in vitro human liver model to study interactions between pathogens and key liver cell types. This liver-chip setup integrates endothelial cells, hepatocytes, and macrophages, enabling a comprehensive examination of pathogen–cell interactions within the liver. The microfluidic perfusion chip, resembling a microscope slide, features endothelial cells on top of the membrane receiving perfused cell culture medium, hepatocytes on the opposite side, and macrophages at the interface between endothelial cells and hepatocytes.128S. aureus in planktonic form is introduced into the cell culture medium perfused within the vascular space on the endothelial side of the liver-chip model. Results indicated a notable increase in surviving intracellular bacteria within M2-polarized macrophages 48 hours post-infection.129 Future investigations utilizing advanced microphysiological models aim to provide deeper insights into S. aureus adaptation to environmental stimuli and its mechanisms for persistence and dissemination within the human liver. Gröger et al. found that monocyte-induced inflammation-related hepatocyte dysfunction was restored in another study. Liver dysfunction often marks the onset of multi-organ failure associated with sepsis. Their biochip employs a suspended porous membrane as a substrate for human umbilical vein endothelial cells, which are co-cultured with primary macrophages to replicate the immunoregulatory roles of Kupffer cells.130 This model replicates inflammation-induced liver dysfunction at a cellular level and enables the study of monocyte-mediated tissue restoration and immune tolerance mechanisms.
4. Challenges and future directions for microfluidic models in bacteria detection and inflammatory modeling
Microfluidic models have emerged as transformative tools, particularly in the context of bacteria detection and inflammation-driven diseases. These platforms offer unprecedented control over the cellular microenvironment, enabling researchers to mimic the complex interactions during inflammatory responses. It also paves the way for home and POCT development, which is critical to curbing any pathogen outbreak. Despite significant advancements, several challenges still need to be addressed, limiting the widespread application of microfluidic models in bacteria detection and inflammatory modeling. Addressing these challenges is crucial for advancing the field and unlocking the full potential of these technologies in clinical and research settings. This section explores the key challenges and outlines potential future directions for microfluidic in bacteria detection and inflammation-driven diseases.4.1 Advanced tissue engineering and organoid integration
One of the primary challenges in developing microfluidic models for inflammatory detection is accurately reproducing the physiological complexity of human tissues and organs. Inflammation is a highly dynamic process involving multiple cell types, signaling molecules, and extracellular matrix components.131 Traditional microfluidic models often fail to replicate this complexity, as they may need more cellular diversity and capture the intricate signaling networks that drive inflammation. Additionally, the static nature of many microfluidic systems does not account for the temporal and spatial variations in inflammatory responses, limiting the ability to model chronic or evolving inflammatory conditions.Recent advancements in organoid-based models have garnered significant attention, mainly due to their high scalability and ability to mimic human organs' physiological complexity better. Organoids, which are miniature, self-organizing tissue cultures that replicate the architecture and function of organs, offer a promising approach for creating more physiologically relevant models of inflammation.132 These culture systems provide a more accessible and cost-effective alternative to traditional OOC models, making them valuable tools for studying diseases, particularly those difficult to model in animals.133 The scalability of organoids and their ability to replicate the multicellular interactions of inflamed tissues has fueled their growing popularity in microfluidic research. For example, recent work by researchers utilizing microfluidics for spheroid-based research134 has shown the potential of integrating spheroids with microfluidic systems, leading to commercial ventures such as Okomera and significant investment from major companies like Roche.
Future microfluidic models should incorporate advanced tissue engineering techniques, including organoids, tumor spheroids, and 3D bioprinting, to overcome the limitations of reproducing physiological complexity.135 3D bioprinting, for instance, can create more intricate tissue structures within microfluidic devices, allowing for studying inflammation in a more realistic context.136 The integration of organoids into microfluidic systems enhances the mimicking of organ-specific functions, offering exciting prospects for precision medicine and personalized disease treatment. The combination of organoids and OOC holds significant potential for advancing disease modeling, as the strengths of each approach can complement one another in replicating complex disease mechanisms and improving therapeutic efficacy.
4.2 High-throughput and automated systems
Another significant challenge is the scalability of microfluidic models. While these systems excel in creating highly controlled microenvironments, they often do so at the expense of throughput. The fabrication of microfluidic devices can be labor-intensive and requires specialized equipment, making it difficult to produce large quantities of devices for high-throughput screening. This limitation is particularly problematic when studying inflammation across various conditions or simultaneously testing multiple therapeutic agents. Furthermore, the small-scale microfluidic models have rewarded biotechnology researchers with vast amounts of data but not necessarily the ability to analyze complex data effectively. The significant opportunity presented by the amalgamation of microfluidics and deep learning in tackling biotechnological hurdles has yet to be fully realised.137,138Scalifully realistic models for high-throughput applications are essential for their broader research and drug development adoption. Advances in microfabrication techniques, such as soft lithography and injection molding, can facilitate the production of large quantities of microfluidic devices at lower costs. Additionally, automation and integration with robotics can enhance the throughput of microfluidic experiments, allowing for the rapid screening of multiple conditions or therapeutic agents. Developing modular microfluidic systems, where different components can be easily exchanged or combined, will also contribute to increased scalability and flexibility in experimental design.139
4.3 Immune-on-a-chip models
A critical aspect of inflammation is the immune system's involvement, which orchestrates the response to injury or infection.140 However, integrating immune components into microfluidic models presents several challenges. Immune cells are highly sensitive to their microenvironment, and maintaining their viability and functionality within a microfluidic system can be difficult. Additionally, the immune response is influenced by a myriad of factors, including the presence of pathogens, cytokines, and other signaling molecules.141 Replicating these conditions in a microfluidic device requires precise control over multiple variables, which can be technically challenging and may result in models that must fully capture the immune response's complexity.Developing immune-on-a-chip models represents a promising direction for integrating immune components into microfluidic systems. These models aim to replicate the interactions between immune cells and other tissue types within a controlled microenvironment, allowing for studying immune-mediated inflammatory responses. Researchers can better understand the immune system's role in inflammation by incorporating lymphoid structures, cytokine gradients, and pathogen exposure into microfluidic devices.142 Immune-on-a-chip models could also be used to investigate the effects of immunotherapies or to model autoimmune diseases, where the immune system plays a central role in driving inflammation.
4.4 Material innovations
The materials used to fabricate microfluidic devices can present challenges in inflammation modeling. PDMS is commonly chosen for its biocompatibility and ease of fabrication. However, one of the major drawbacks of PDMS is its high cost per chip, particularly in comparison to thermoplastics. This concern has been highlighted in recent literature. Polycarbonate, a thermoplastic material, offers a cost-effective alternative to PDMS, providing benefits such as low enzyme adsorption, thermal stability, optical transparency, and compatibility with scalable production processes.143 Another limitation of PDMS is its propensity to absorb small molecules, which can alter the concentration of inflammatory mediators and affect the model's accuracy.144 PDMS exhibits hydrophobic properties, a characteristic that frequently constrains its utility in biological sample solutions. Moreover, PDMS tends to swell when exposed to specific reagents.Conversely, numerous endeavors have been undertaken to render PDMS surfaces hydrophilic and resistant to protein adsorption. However, these approaches also face constraints such as challenges in reproducibility, durability, chemical stability, mechanical degradation, and the inability to sustain hydrophilic surfaces over extended periods.145 Potential solutions include using hydrogels as a matrix to improve the functionality and feasibility of PDMS microfluidic chips,146 but they bring new challenges in manufacturing complexity and device durability.
Developing new materials for microfluidic devices will be crucial for overcoming the limitations of current systems. Materials that more closely mimic the mechanical properties of human tissues, such as hydrogels with tunable stiffness, can improve the physiological relevance of microfluidic models. Additionally, using materials with low absorption properties can reduce the loss of small molecules and enhance the accuracy of inflammatory detection. Advances in biomaterial science, including developing stimuli-responsive materials that can change their properties in response to environmental cues, offer exciting possibilities for creating more dynamic and responsive microfluidic models.
4.5 Integration of real-time sensing technologies
Incorporating real-time sensing and data collection into microfluidic models is challenging. Developing integrated sensors that can continuously monitor parameters such as cytokine levels, cell migration, and changes in tissue architecture is essential for capturing the full spectrum of inflammatory responses. However, integrating such sensors into microfluidic systems requires sensor technology and device engineering advances.Advances in microelectronics and nanotechnology have led to the development of miniaturized sensors capable of detecting a wide range of biological signals, including cytokine levels, pH changes, and oxygen concentration.147 These sensors can be integrated into microfluidic devices to continuously monitor inflammatory processes, enabling researchers to track the progression of inflammation in real time. Furthermore, non-invasive imaging techniques, such as fluorescence microscopy or optical coherence tomography, can complement these sensors by providing detailed information on cellular and tissue-level changes during inflammation. Smartphones have been widely used for inspection in microfluidic systems with built-in cameras and internal microprocessors, which can perform image processing without external computers and share real-time results when necessary.148
4.6 Clinical translation
In contemplating the future of tests about POCT clinical diagnostics, it remains a crucial necessity to emphasize fundamental research and its application in clinical settings, leveraging advanced technologies like nanotechnology and biotechnology. A symbiotic relationship between frontline medical institutions and academia is imperative to expedite the translation of these innovations into clinical practice. Sustained collaboration is vital among various stakeholders, including researchers, technologists, end users (such as physicians and healthcare professionals), and business partners (like marketing specialists and investors) to effectively introduce microfluidic tools and techniques to the market cohesively and cooperatively. Clinical trials should encompass proof-of-concept studies while considering scalability for successful commercialization and optimal performance outcomes.5. Conclusion
We aim to showcase the utilization of microfluidic chips in both bacterial detection and models of bacteria-driven inflammatory infections. After identifying the pathogens responsible for infections within the body, we then elucidate the inflammatory responses and subsequent treatment protocols following bacterial infections across various diseases. This comprehensive approach will provide readers with a thorough understanding of the diverse applications of microfluidic chips. Microfluidic models are promising for advancing our understanding of inflammation and developing new bacteria detection and treatment strategies. However, several challenges remain, including the need to better replicate physiological complexity, improve scalability, integrate immune components, overcome material limitations, and enhance real-time monitoring capabilities. Addressing these challenges will require interdisciplinary collaboration and continued innovation in materials science, tissue engineering, and sensor technology. By focusing on these areas, future research can unlock the full potential of microfluidic models, paving the way for more effective therapies and diagnostic tools in the fight against bacteria-associated inflammatory diseases.Abbreviations
| TB | Tuberculosis |
| PCR | Polymerase chain reaction |
| EIA | Enzyme-linked immunoassay |
| ELFA | Enzyme-linked fluorescence assay |
| ELISA | Enzyme-linked immunosorbent assay |
| IMB | Immunomagnetic beads |
| S. aureus | Staphylococcus aureus |
| MNPs | Magnetic nanoparticles |
| μPAD | Microfluidic paper-based analysis device |
| SERS | Surface-enhanced Raman scattering |
| S. typhi | Salmonella typhimurium |
| POCT | Point-of-care testing |
| AuNPs | Gold nanoparticles |
| TMB | 3,3′,5,5′-Tetramethylbenzidine |
| ABX-GMFETs | Graphene micropatterned field-effect transistor |
| MTB | Mycobacterium tuberculosis |
| OOC | Organ-on-a-chip |
| FMi–PLA-OOC | Fetal membrane/placenta-deciduous interface OOC |
| FMi-OOC | Feto-maternal interface |
| LPS | Lipopolysaccharide |
| VCD-OOC | Vagina-cervix-decidua OOC |
| FMi–PLA-OOC | Fetal membrane/placenta (fetal)-decidua (maternal) interface organ-on-a-chip |
| eIL-10 | Interleukin-10 |
| FMi | Feto-maternal interface |
| U. parvum | Ureaplasma parvum |
| ECM | Extracellular matrix |
| hiPSCs | Human induced pluripotent stem cells |
| PM | Placental malaria |
| CSA | Chondroitin sulfate A |
| HuMiX | Human-microbial crosstalk |
| TEER | Trans-epithelial electrical resistance |
| LGG | Lactobacillus rhamnosus GG |
| IBD | Inflammatory bowel disease |
| EMI | Epithelial–microbiome–immune |
| DoGC | Depression-on-gut-chip |
| F. prausnitzii | Faecalibacterium prausnitzii |
| IB | Intratumoral bacteria |
| EB | Extratumoral bacteria |
| GI | Gastrointestinal |
| CRC | Colorectal cancer |
| Fn | Fusobacterium nucleatum |
| PDMS | Polydimethylsiloxane |
Data availability
No primary research results, new data, software or code have been included as part of this review.Author contributions
B. L. K. conceived the manuscript, J. Z., Y. T. F., C. Y. F., H. J. H., W. L., and B. L. K. wrote and revised the manuscript. The author(s) read and approved the final manuscript.Conflicts of interest
The authors declare that they have no competing interests.Acknowledgements
This work was supported by the City University of Hong Kong (7006082, 7020073, 9609332, 9609333, 9678292, 7020002), the Research Grants Council (RGC) (9048206, 8799020), the Hong Kong Center for Cerebro-Cardiovascular Health Engineering (COCHE), Innovation and Technology Commission (PRP/001/22FX), the Hetao Shenzhen-Hong Kong Science and Technology Innovation Cooperation Zone Shenzhen Park Project (HZQB-KCZYZ-2021017), Science Research Project of Hebei Education Department (QN2025325), Department of Education of Hebei Province (QN2025325), Hebei University Medical Science (2024B07), and the Education Bureau Gifted Education Programme (3030780).References
- E. Roy, A. Nagar, S. Chaudhary and S. Pal, ChemistrySelect, 2020, 5, 722–735 CrossRef CAS.
- W. Li, K. Dong, J. Ren and X. Qu, Am. Ethnol., 2016, 128, 8181–8185 Search PubMed.
- A. H. Havelaar, M. D. Kirk, P. R. Torgerson, H. J. Gibb, T. Hald, R. J. Lake, N. Praet, D. C. Bellinger, N. R. De Silva and N. Gargouri, PLoS Med., 2015, 12, e1001923 CrossRef PubMed.
- D. M. Monack, A. Mueller and S. Falkow, Nat. Rev. Microbiol., 2004, 2, 747–765 CrossRef CAS PubMed.
- C. de Martel, D. Georges, F. Bray, J. Ferlay and G. M. Clifford, Lancet Global Health, 2020, 8, e180–e190 CrossRef PubMed.
- L. Yang, A. Li, Y. Wang and Y. Zhang, Signal Transduction Targeted Ther., 2023, 8, 35 CrossRef CAS PubMed.
- X. Ma, W. Ding, C. Wang, H. Wu, X. Tian, M. Lyu and S. Wang, Sens. Actuators, B, 2021, 331, 129422 CrossRef CAS.
- J. Zhang, J. Huang and F. He, Biosens. Bioelectron., 2019, 138, 111322 CrossRef CAS PubMed.
- C. Fleischmann, A. Scherag, N. K. Adhikari, C. S. Hartog, T. Tsaganos, P. Schlattmann, D. C. Angus and K. Reinhart, Am. J. Respir. Crit. Care Med., 2016, 193, 259–272 Search PubMed.
- R. Gao, X. Liu, Z. Xiong, G. Wang and L. Ai, Food Res. Int., 2024, 114767 CrossRef CAS PubMed.
- S. C. Davies, T. Fowler, J. Watson, D. M. Livermore and D. Walker, Lancet, 2013, 381, 1606–1609 CrossRef PubMed.
- B. Strommenger, C. Kettlitz, G. Werner and W. Witte, J. Clin. Microbiol., 2003, 41, 4089–4094 Search PubMed.
- O. Lazcka, F. J. Del Campo and F. X. Munoz, Biosens. Bioelectron., 2007, 22, 1205–1217 CrossRef CAS PubMed.
- S. Yang and R. E. Rothman, Lancet Infect. Dis., 2004, 4, 337–348 CrossRef CAS PubMed.
- B. A. Slominski and P. S. Lee, in Practical Handbook of Microbiology, CRC Press, 2021, pp. 19–36 Search PubMed.
- T. Di Febo, M. Schirone, P. Visciano, O. Portanti, G. Armillotta, T. Persiani, E. Di Giannatale, M. Tittarelli and M. Luciani, Food Anal. Methods, 2019, 12, 322–330 CrossRef.
- J.-C. Lagier, S. Edouard, I. Pagnier, O. Mediannikov, M. Drancourt and D. Raoult, Clin. Microbiol. Rev., 2015, 28, 208–236 CrossRef CAS PubMed.
- Y. Zhao, Y. Shang, Z. Wang, Z. Wang, J. Xie, H. Zhai, Z. Huang, Y. Wang, Q. Wu and Y. Ding, TrAC, Trends Anal. Chem., 2024, 117736 CrossRef CAS.
- D. M. Silva and L. Domingues, Ecotoxicol. Environ. Saf., 2015, 113, 400–411 CrossRef PubMed.
- N. Amino and Y. Hidaka, Nihon Rinsho, 1995, 53, 2107–2111 CAS.
- J. M. Van Emon, Immunoassay and other bioanalytical techniques, CRC Press, 2016 Search PubMed.
- T.-H. Liu, S.-S. Cheng, H.-L. You, M. S. Lee and G.-B. Lee, Analyst, 2019, 144, 1210–1222 RSC.
- Y. Wen, Y. Tan, L. Zhao, X. Lv, L. Lin, D. Liang and L. Wang, Microchem. J., 2022, 178, 107348 CrossRef CAS.
- L. F. Alonzo, T. C. Hinkley, A. Miller, R. Calderon, S. Garing, J. Williford, N. Clute-Reinig, E. Spencer, M. Friend and D. Madan, Lab Chip, 2022, 22, 2155–2164 RSC.
- C. Nogueira, J. Sousa, R. Afonso, D. Santos, P. Rodrigues, L. Albuquerque, A. Faustino, S. Abalde-Cela, P. Freitas and S. Cardoso, Sens. Actuators, B, 2024, 404, 135175 CrossRef CAS.
- J. Zhan, L. Wang, Y. Zhu, H. Gao, Y. Chen, J. Chen, Y. Jia, J. He, Z. Fang and Y. Zhu, ACS Appl. Mater. Interfaces, 2018, 10, 35830–35837 CrossRef CAS PubMed.
- K. H. Kim, S. J. Park, C. S. Park, S. E. Seo, J. Lee, J. Kim, S. H. Lee, S. Lee, J.-S. Kim and C.-M. Ryu, Biosens. Bioelectron., 2020, 167, 112514 CrossRef CAS PubMed.
- C.-H. Wang, J.-R. Chang, S.-C. Hung, H.-Y. Dou and G.-B. Lee, Sens. Actuators, B, 2022, 365, 131968 CrossRef CAS.
- B. Koskella and S. Meaden, Viruses, 2013, 5, 806–823 CrossRef PubMed.
- S. P. Costa, C. R. Caneira, V. Chu, P. P. Freitas, J. P. Conde and C. M. Carvalho, Sens. Actuators, B, 2023, 376, 132917 CrossRef CAS.
- S. G. Dastider, S. Barizuddin, N. S. Yuksek, M. Dweik and M. F. Almasri, J. Sens., 2015, 2015, 293461 Search PubMed.
- J. Shrestha, S. R. Bazaz, L. Ding, S. Vasilescu, S. Idrees, B. Söderström, P. M. Hansbro, M. Ghadiri and M. E. Warkiani, Lab Chip, 2023, 23, 146–156 RSC.
- C.-H. Chang, C.-L. Wang and B.-R. Li, Biosens. Bioelectron., 2023, 236, 115403 CrossRef CAS PubMed.
- J. Liao, J. Ren, H. Wei, R. H. Lam, S. L. Chua and B. L. Khoo, Biosens. Bioelectron., 2021, 191, 113412 CrossRef CAS PubMed.
- Y.-H. Nam, S. Lee, S.-K. Lee, J.-H. Kim and J.-H. Park, Sens. Actuators, B, 2024, 404, 135202 CrossRef CAS.
- D. Liu, Y. Zhu, N. Li, Y. Lu, J. Cheng and Y. Xu, Sens. Actuators, B, 2020, 310, 127834 CrossRef CAS.
- I. Michael, D. Kim, O. Gulenko, S. Kumar, S. Kumar, J. Clara, D. Y. Ki, J. Park, H. Y. Jeong and T. S. Kim, Nat. Biomed. Eng., 2020, 4, 591–600 CrossRef CAS PubMed.
- J. Liao, S. Zou, Y. Deng, Y. Jiang, S. Chua and B. Khoo, Chem. Eng. J., 2023, 453, 139595 Search PubMed.
- Q. Shang, Y. Su, Y. Liang, W. Lai, J. Jiang, H. Wu and C. Zhang, Anal. Bioanal. Chem., 2020, 412, 3787–3797 Search PubMed.
- W. Wu, B. T. T. Nguyen, P. Y. Liu, G. Cai, S. Feng, Y. Shi, B. Zhang, Y. Hong, R. Yu and X. Zhou, Biosens. Bioelectron., 2022, 215, 114594 CrossRef CAS PubMed.
- A. Cui, L. Dong, Y. Hou, X. Mu, Y. Sun, H. Wang, X. Zhong and G. Shan, Biosens. Bioelectron., 2024, 116320 CrossRef CAS PubMed.
- J. Zhuang, Z. Zhao, K. Lian, L. Yin, J. Wang, S. Man, G. Liu and L. Ma, Biosens. Bioelectron., 2022, 207, 114167 CrossRef CAS PubMed.
- C.-H. Wang, J.-J. Wu and G.-B. Lee, Sens. Actuators, B, 2019, 284, 395–402 CrossRef CAS.
- Ö. Baltekin, A. Boucharin, E. Tano, D. I. Andersson and J. Elf, Proc. Natl. Acad. Sci. U. S. A., 2017, 114, 9170–9175 CrossRef PubMed.
- W. Liu, J.-W. Tang, J.-W. Lyu, J.-J. Wang, Y.-C. Pan, X.-Y. Shi, Q.-H. Liu, X. Zhang, B. Gu and L. Wang, Microbiol. Spectrum, 2022, 10, e02409–e02421 CAS.
- G. Xing, N. Li, H. Lin, Y. Shang, Q. Pu and J.-M. Lin, Talanta, 2023, 253, 123980 Search PubMed.
- Y. Wan, L. Zheng, Y. Sun and D. Zhang, J. Mater. Chem. B, 2014, 2, 4818–4825 Search PubMed.
- J. Jiang, H. Wu, Y. Su, Y. Liang, B. Shu and C. Zhang, Anal. Chem., 2020, 92, 7708–7716 CrossRef CAS PubMed.
- H. Jiang, Z. Sun, Q. Guo and X. Weng, Biosens. Bioelectron., 2021, 182, 113191 CrossRef CAS PubMed.
- Y. Hou, W. Tang, W. Qi, X. Guo and J. Lin, Biosens. Bioelectron., 2020, 157, 112160 CrossRef CAS PubMed.
- S. Fande, K. Amreen, D. Sriram and S. Goel, Anal. Chim. Acta, 2023, 1237, 340591 Search PubMed.
- L. Zheng, G. Cai, S. Wang, M. Liao, Y. Li and J. Lin, Biosens. Bioelectron., 2019, 124, 143–149 Search PubMed.
- R. Guo, S. Wang, F. Huang, Q. Chen, Y. Li, M. Liao and J. Lin, Sens. Actuators, B, 2019, 284, 134–139 Search PubMed.
- S. Kim, A. Romero-Lozano, D. S. Hwang and J.-Y. Yoon, J. Hazard. Mater., 2021, 413, 125338 Search PubMed.
- H. Huang, G. Zhao and W. Dou, Biosens. Bioelectron., 2018, 107, 266–271 Search PubMed.
- L. Wang, X. Huo, F. Jiang, X. Xi, Y. Li and J. Lin, Sens. Actuators, B, 2023, 393, 134242 Search PubMed.
- I. P. Alves and N. M. Reis, Biosens. Bioelectron., 2019, 145, 111624 CrossRef CAS PubMed.
- L. Zhang, F. Tian, C. Liu, Q. Feng, T. Ma, Z. Zhao, T. Li, X. Jiang and J. Sun, Lab Chip, 2018, 18, 610–619 RSC.
- W. Zhou, J. Sun and X. Li, Anal. Chem., 2020, 92, 14830–14837 CrossRef CAS PubMed.
- H. Wang, Z. Ma, J. Qin, Z. Shen, Q. Liu, X. Chen, H. Wang, Z. An, W. Liu and M. Li, Biosens. Bioelectron., 2019, 126, 373–380 CrossRef CAS PubMed.
- A. Signore, EJNMMI Res., 2013, 3, 8 Search PubMed.
- A. Signore and A. W. Glaudemans, Ann. Nucl. Med., 2011, 25, 681–700 Search PubMed.
- Y. López Hernández, D. Yero, J. M. Pinos-Rodríguez and I. Gibert, Front. Microbiol., 2015, 6, 38 Search PubMed.
- Y. Fu, Y. Deng, J. Zhang, S. L. Chua and B. L. Khoo, Acta Biomater., 2023, 168, 333–345 CrossRef CAS PubMed.
- S. Sarkar and M. T. Heise, Clin. Ther., 2019, 41, 1912–1922 CrossRef CAS PubMed.
- I. C. Herron, T. R. Laws and M. Nelson, Front. Cell. Infect. Microbiol., 2024, 14, 1340017 CrossRef CAS PubMed.
- A. Abrhaley, M. Giday and A. Hailu, BMC Med. Res. Methodol., 2024, 24, 211 Search PubMed.
- J. J. Han, Artif. Organs, 2023, 47, 449–450 CrossRef PubMed.
- W. G. Rosen, R. C. Mohs and K. L. Davis, Am. J. Psychiatry, 1984, 141, 1356–1364 CrossRef CAS PubMed.
- D. Galasko, D. Bennett, M. Sano, C. Ernesto, R. Thomas, M. Grundman and S. Ferris, Alzheimer Dis. Assoc. Disord., 1997, 11, 33–39 Search PubMed.
- F. S. Gazzaniga, D. M. Camacho, M. Wu, M. F. S. Palazzo, A. L. Dinis, F. N. Grafton, M. J. Cartwright, M. Super, D. L. Kasper and D. E. Ingber, Front. Cell. Infect. Microbiol., 2021, 11, 638014 CrossRef CAS PubMed.
- M. Safarzadeh, L. S. Richardson, A. K. Kammala, A. Mosebarger, M. Bettayeb, S. Kim, P. Y. Lam, E. Radnaa, A. Han and R. Menon, J. Extracell. Vesicles, 2024, 3, 100035 CrossRef PubMed.
- G. Mahajan, E. Doherty, T. To, A. Sutherland, J. Grant, A. Junaid, A. Gulati, N. LoGrande, Z. Izadifar and S. S. Timilsina, Microbiome, 2022, 10, 201 Search PubMed.
- R. Romero, J. Espinoza, J. P. Kusanovic, F. Gotsch, S. Hassan, O. Erez, T. Chaiworapongsa and M. Mazor, BJOG, 2006, 113, 17–42 Search PubMed.
- L. S. Richardson, S. Kim, A. Han and R. Menon, Lab Chip, 2020, 20, 4486–4501 Search PubMed.
- O. A. G. Tantengco, L. S. Richardson, E. Radnaa, A. K. Kammala, S. Kim, P. M. B. Medina, A. Han and R. Menon, FASEB J., 2022, 36, e22551 CrossRef CAS PubMed.
- Z. Izadifar, J. Cotton, S. Chen, V. Horvath, A. Stejskalova, A. Gulati, N. T. LoGrande, B. Budnik, S. Shahriar, E. R. Doherty, Y. Xie, T. To, S. E. Gilpin, A. M. Sesay, G. Goyal, C. B. Lebrilla and D. E. Ingber, Nat. Commun., 2024, 15, 4578 Search PubMed.
- T. Cindrova-Davies and A. N. Sferruzzi-Perri, Semin. Cell Dev. Biol., 2022, 131, 66–77 CrossRef CAS PubMed.
- J. Hutson, F. Garcia-Bournissen, A. Davis and G. Koren, Clin. Pharmacol. Ther., 2011, 90, 67–76 Search PubMed.
- K. E. Madsen, C. Ø. Mariager, C. S. Duvald, E. S. S. Hansen, L. B. Bertelsen, M. Pedersen, L. H. Pedersen, N. Uldbjerg and C. Laustsen, Tomography, 2019, 5, 333 CrossRef PubMed.
- F. Soydemir, S. Kuruvilla, M. Brown, W. Dunn, P. Day, I. P. Crocker, P. N. Baker, C. P. Sibley and P. Brownbill, Lab. Invest., 2011, 91, 181–189 CrossRef CAS PubMed.
- Y. Zhu, F. Yin, H. Wang, L. Wang, J. Yuan and J. Qin, ACS Biomater. Sci. Eng., 2018, 4, 3356–3363 CrossRef CAS PubMed.
- F. Yin, Y. Zhu, H. Wang, Y. Wang, D. Li and J. Qin, ACS Biomater. Sci. Eng., 2020, 6, 4644–4652 CrossRef CAS PubMed.
- B. Mosavati, A. Oleinikov and E. Du, Sci. Rep., 2022, 12, 15278 CrossRef CAS PubMed.
- K. Nishino, A. Nishida, R. Inoue, Y. Kawada, M. Ohno, S. Sakai, O. Inatomi, S. Bamba, M. Sugimoto and M. Kawahara, J. Gastroenterol., 2018, 53, 95–106 CrossRef PubMed.
- F. Karlsson, V. Tremaroli, J. Nielsen and F. Bäckhed, Diabetes, 2013, 62, 3341–3349 CrossRef CAS PubMed.
- Z. Jie, H. Xia, S.-L. Zhong, Q. Feng, S. Li, S. Liang, H. Zhong, Z. Liu, Y. Gao and H. Zhao, Nat. Commun., 2017, 8, 845 CrossRef PubMed.
- P. Shah, J. V. Fritz, E. Glaab, M. S. Desai, K. Greenhalgh, A. Frachet, M. Niegowska, M. Estes, C. Jäger, C. Seguin-Devaux, F. Zenhausern and P. Wilmes, Nat. Commun., 2016, 7, 11535 Search PubMed.
- W. Zhao, Y. Yao, T. Zhang, H. Lu, X. Zhang, L. Zhao, X. Chen, J. Zhu, G. Sui and W. Zhao, Front. Bioeng. Biotechnol., 2022, 10, 1035647 CrossRef PubMed.
- C. Wang, T. Dang, J. Baste, A. Anil Joshi and A. Bhushan, FASEB J., 2021, 35, e21291 CAS.
- H. J. Kim, H. Li, J. J. Collins and D. E. Ingber, Proc. Natl. Acad. Sci. U. S. A., 2016, 113, E7–E15 Search PubMed.
- J. Liu, R. Lu, X. Zheng, W. Hou, X. Wu, H. Zhao, G. Wang and T. Tian, Biomater. Sci., 2023, 11, 2504–2517 RSC.
- Y. M. Ambrosini, W. Shin, S. Min and H. J. Kim, Immune Netw., 2020, 20, e13 CrossRef PubMed.
- W. Wang, Y. Liu, Z. Yao, D. Chen, Y. Tang, J. Cui, J. Zhang, H. Liu and Z. Hao, Lab Chip, 2024, 24, 2537–2550 RSC.
- M. T.-Q. Duong, Y. Qin, S.-H. You and J.-J. Min, Exp. Mol. Med., 2019, 51, 1–15 Search PubMed.
- A. Ozcelikkale, K. Shin, V. Noe-Kim, B. D. Elzey, Z. Dong, J.-T. Zhang, K. Kim, I. C. Kwon, K. Park and B. Han, J. Controlled Release, 2017, 266, 129–139 CrossRef CAS PubMed.
- S. Khatun, T. Appidi and A. K. Rengan, Curr. Res. Microb. Sci., 2021, 2, 100078 CAS.
- A. Fu, B. Yao, T. Dong, Y. Chen, J. Yao, Y. Liu, H. Li, H. Bai, X. Liu and Y. Zhang, Cell, 2022, 185, 1356–1372.e1326 CrossRef CAS PubMed.
- Y. Deng, S. Y. Liu, S. L. Chua and B. L. Khoo, Biosens. Bioelectron., 2021, 180, 113113 CrossRef CAS PubMed.
- S. Uthaman, S. Zheng, J. Han, Y. J. Choi, S. Cho, V. D. Nguyen, J. O. Park, S. H. Park, J. J. Min and S. Park, Adv. Healthcare Mater., 2016, 5, 288–295 Search PubMed.
- C. L. Walsh, B. M. Babin, R. W. Kasinskas, J. A. Foster, M. J. McGarry and N. S. Forbes, Lab Chip, 2009, 9, 545–554 RSC.
- B. Subia, U. R. Dahiya, S. Mishra, J. Ayache, G. V. Casquillas, D. Caballero, R. L. Reis and S. C. Kundu, J. Controlled Release, 2021, 331, 103–120 Search PubMed.
- Y. B. Kang, T. R. Sodunke, J. Lamontagne, J. Cirillo, C. Rajiv, M. J. Bouchard and M. Noh, Biotechnol. Bioeng., 2015, 112, 2571–2582 Search PubMed.
- K.-H. Nam, A. S. Smith, S. Lone, S. Kwon and D.-H. Kim, J. Lab. Autom., 2015, 20, 201–215 CrossRef CAS PubMed.
- D. Penarete-Acosta, R. Stading, L. Emerson, M. Horn, S. Chakraborty, A. Han and A. Jayaraman, Lab Chip, 2024, 24, 3690–3703 RSC.
- J. Song, Y. Zhang, C. Zhang, X. Du, Z. Guo, Y. Kuang, Y. Wang, P. Wu, K. Zou and L. Zou, Sci. Rep., 2018, 8, 6394 Search PubMed.
- A. Gagnaire, B. Nadel, D. Raoult, J. Neefjes and J.-P. Gorvel, Nat. Rev. Microbiol., 2017, 15, 109–128 CrossRef CAS PubMed.
- D. Penarete-Acosta, R. Stading, L. Emerson, M. Horn, S. Chakraborty, A. Han and A. Jayaraman, Lab Chip, 2024, 24, 3690–3703 RSC.
- A. Burmeister and A. Grünberger, Curr. Opin. Biotechnol., 2020, 62, 106–115 CrossRef CAS PubMed.
- T. Liu, C. Li, H. Li, S. Zeng, J. Qin and B. Lin, Electrophoresis, 2009, 30, 4285–4291 CrossRef CAS PubMed.
- J. W. Hong, S. Song and J. H. Shin, Lab Chip, 2013, 13, 3033–3040 RSC.
- A. Skardal, M. Devarasetty, S. Soker and A. R. Hall, Biofabrication, 2015, 7, 031001 CrossRef PubMed.
- J. H. Sung and M. L. Shuler, Lab Chip, 2009, 9, 1385–1394 RSC.
- W. Tan and T. A. Desai, Biomaterials, 2004, 25, 1355–1364 CrossRef CAS PubMed.
- A. P. Wong, R. Perez-Castillejos, J. C. Love and G. M. Whitesides, Biomaterials, 2008, 29, 1853–1861 CrossRef CAS PubMed.
- M. S. Kim, J. H. Yeon and J.-K. Park, Biomed. Microdevices, 2007, 9, 25–34 Search PubMed.
- B. Foxman, Nat. Rev. Urol., 2010, 7, 653–660 CrossRef PubMed.
- H. Horsley, D. Dharmasena, J. Malone-Lee and J. L. Rohn, Sci. Rep., 2018, 8, 1238 CrossRef PubMed.
- Y. C. Smith, K. K. Grande, S. B. Rasmussen and A. D. O'Brien, Infect. Immun., 2006, 74, 750–757 CrossRef CAS PubMed.
- K. Sharma, N. Dhar, V. V. Thacker, T. M. Simonet, F. Signorino-Gelo, G. W. Knott and J. D. McKinney, eLife, 2021, 10, e66481 CrossRef CAS PubMed.
- E. Azoulay, D. Mokart, A. Kouatchet, A. Demoule and V. Lemiale, Lancet Respir. Med., 2019, 7, 173–186 CrossRef PubMed.
- S. Deinhardt-Emmer, S. Sachse, J. Geraci, C. Fischer, A. Kwetkat, K. Dawczynski, L. Tuchscherr and B. Löffler, J. Hosp. Infect., 2018, 100, 309–315 CrossRef CAS PubMed.
- D. Masopust, C. P. Sivula and S. C. Jameson, J. Immunol., 2017, 199, 383–388 CrossRef CAS PubMed.
- S. Deinhardt-Emmer, K. Rennert, E. Schicke, Z. Cseresnyés, M. Windolph, S. Nietzsche, R. Heller, F. Siwczak, K. F. Haupt and S. Carlstedt, Biofabrication, 2020, 12, 025012 Search PubMed.
- V. V. Thacker, N. Dhar, K. Sharma, R. Barrile, K. Karalis and J. D. McKinney, eLife, 2020, 9, e59961 CrossRef CAS PubMed.
- L. J. Dixon, M. Barnes, H. Tang, M. T. Pritchard and L. E. Nagy, Compr. Physiol., 2013, 3, 785 CrossRef PubMed.
- M. Bilzer, F. Roggel and A. L. Gerbes, Liver Int., 2006, 26, 1175–1186 CrossRef CAS PubMed.
- K. Rennert, S. Steinborn, M. Gröger, B. Ungerböck, A.-M. Jank, J. Ehgartner, S. Nietzsche, J. Dinger, M. Kiehntopf and H. Funke, Biomaterials, 2015, 71, 119–131 CrossRef CAS PubMed.
- F. Siwczak, Z. Cseresnyes, M. I. A. Hassan, K. O. Aina, S. Carlstedt, A. Sigmund, M. Groger, B. G. Surewaard, O. Werz and M. T. Figge, Biomaterials, 2022, 287, 121632 CrossRef CAS PubMed.
- M. Gröger, K. Rennert, B. Giszas, E. Weiß, J. Dinger, H. Funke, M. Kiehntopf, F. T. Peters, A. Lupp and M. Bauer, Sci. Rep., 2016, 6, 1–16 CrossRef PubMed.
- G. G. Vaday and O. Lider, J. Leukocyte Biol., 2000, 67, 149–159 Search PubMed.
- M. Hofer and M. P. Lutolf, Nat. Rev. Mater., 2021, 6, 402–420 CrossRef CAS PubMed.
- E. Jackson and E. H. Lu, Integr. Biol., 2016, 8, 672–683 Search PubMed.
- V. Bonnet, E. Maikranz, M. Madec, N. Vertti-Quintero, C. Cuche, M. Mastrogiovanni, A. Alcover, V. Di Bartolo and C. N. Baroud, Proc. Natl. Acad. Sci. U. S. A., 2024, 121, e2316500121 CrossRef PubMed.
- M. Filippi, T. Buchner, O. Yasa, S. Weirich and R. K. Katzschmann, Adv. Mater., 2022, 34, 2108427 Search PubMed.
- C. G. van Dijk, M. M. Brandt, N. Poulis, J. Anten, M. van der Moolen, L. Kramer, E. F. Homburg, L. Louzao-Martinez, J. Pei and M. M. Krebber, Lab Chip, 2020, 20, 1827–1844 RSC.
- J. Riordon, D. Sovilj, S. Sanner, D. Sinton and E. W. Young, Trends Biotechnol., 2019, 37, 310–324 Search PubMed.
- H. Hua, Y. Deng, J. Zhang, X. Zhou, T. Zhang and B. L. Khoo, Biosens. Bioelectron., 2024, 253, 116086 CrossRef CAS PubMed.
- S. Dekker, W. Buesink, M. Blom, M. Alessio, N. Verplanck, M. Hihoud, C. Dehan, W. César, A. Le Nel and A. Van Den Berg, Sens. Actuators, B, 2018, 272, 468–478 CrossRef CAS.
- O. P. Kulkarni, J. Lichtnekert, H.-J. Anders and S. R. Mulay, Mediators Inflammation, 2016, 2016, 2856213 Search PubMed.
- S. Parlato, G. Grisanti, G. Sinibaldi, G. Peruzzi, C. M. Casciola and L. Gabriele, Lab Chip, 2021, 21, 234–253 Search PubMed.
- M. J. Rupar, H. Hanson, S. Rogers, B. Botlick, S. Trimmer and J. J. Hickman, Lab Chip, 2024, 24, 3604–3625 RSC.
- W. Postek, N. Pacocha and P. Garstecki, Lab Chip, 2022, 22, 3637–3662 RSC.
- M. W. Toepke and D. J. Beebe, Lab Chip, 2006, 6, 1484–1486 RSC.
- I. Miranda, A. Souza, P. Sousa, J. Ribeiro, E. M. Castanheira, R. Lima and G. Minas, J. Funct. Biomater., 2021, 13, 2 CrossRef PubMed.
- Y. Shin, S. Han, J. S. Jeon, K. Yamamoto, I. K. Zervantonakis, R. Sudo, R. D. Kamm and S. Chung, Nat. Protoc., 2012, 7, 1247–1259 CrossRef CAS PubMed.
- A. T. Young, K. R. Rivera, P. D. Erb and M. A. Daniele, ACS Sens., 2019, 4, 1454–1464 CrossRef CAS PubMed.
- J. Zhang, W. Li, Q. Huang, Y. Fu, Y. Liu, X. Luo, S. Zou, S. L. Chua, S. Leung and B. L. Khoo, Sens. Actuators, B, 2023, 395, 134485 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2025 |