
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceDevelopment and evaluation of a microfluidic human testicular tissue chip: a novel in vitro platform for reproductive biology and pharmacology studies†
Jiaming
Shen‡
a,
Xinlong
Wang‡
 a,
Chenghua
Yang
a,
Guanyu
Ren
a,
Lei
Wang
a,
Shuguang
Piao
a,
Boyang
Zhang
a,
Weihao
Sun
a,
Xie
Ge
b,
Jun
Jing
b,
Yijian
Xiang
b,
Zhaowanyue
He
b,
Linhui
Wang
*b,
Bing
Yao
*b and
Zhiyong
Liu
*a
a,
Chenghua
Yang
a,
Guanyu
Ren
a,
Lei
Wang
a,
Shuguang
Piao
a,
Boyang
Zhang
a,
Weihao
Sun
a,
Xie
Ge
b,
Jun
Jing
b,
Yijian
Xiang
b,
Zhaowanyue
He
b,
Linhui
Wang
*b,
Bing
Yao
*b and
Zhiyong
Liu
*a
aDepartment of Urology, Changhai Hospital, Naval Medical University, Shanghai, China. E-mail: medlzhy@aliyun.com
bDepartment of Reproductive Medicine, Nanjing Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University, Nanjing, China
First published on 17th January 2025
Abstract
Organ-on-a-chip culture systems using human organ tissues provide invaluable preclinical insights into systemic functions in vitro. This study aimed to develop a novel human testicular tissue chip within a microfluidic device employing computer-aided design software and photolithography technology. Polydimethylsiloxane was used as the primary material to ensure marked gas permeability and no biotoxicity, enabling effective mimicry of the in vivo testicular microenvironment. This biochip preserved the structural integrity and cellular composition of human testicular tissue, as well as part of its functionality, over an extended period in vitro. Moreover, compared to traditional static culture methods, it more effectively maintained tissue viability and endocrine function. The chip maintained cellular components, histological morphology, and an ultrastructure similar to those in vivo. Notably, the addition of gonadotropins to the human testis tissue on the chip resulted in consistent and steady in vitro production of testosterone and inhibin B. Additionally, the chip displayed sensitivity to the reproductive toxicity of the chemotherapeutic drug busulfan. The results demonstrate the successful establishment of a novel human testicular tissue chip culture system, providing a novel in vitro approach enabling the exploration of human reproductive biology, reproductive pharmacology, toxicology, individual diagnosis, and treatment strategies.
Introduction
Male factor infertility affects 9–16% of all men,1 contributing to 40–50% of all cases of infertility worldwide.2 Testicular cells are typically used to investigate male reproductive biology;3 however, they cannot mimic the testicular microenvironment owing to the lack of complex cell-to-cell communication that is vital for testicular function. Animal models can provide direct evidence related to infertility; however, species-related differences in the pharmacokinetics of compounds may affect data interpretation conclusions.4 Therefore, a more accurate human-representative system is required to model drug-candidate compounds in the male reproduction system.Organ-on-a-chip culture systems have the advantage of recapitulating many important cell niches in vitro compared to cells in a 2D culture. Organ chips based on a microfluidic device (MFD) contain hollow microchannels lined by human-derived cells. They recapitulate organ-specific tissue–tissue interfaces and physical microenvironments (including fluid flow and air–liquid interfaces) that are necessary to mimic human organ-level pathophysiology.5 In recent years, microfluidic technology has been applied to multiple organ-chip culture systems, such as the human lung alveolus, human ovaries, human brain, rat liver slices, and mouse testicular tissue.6–10 In addition, the MFD can closely simulate the physiological structure and microenvironment of the organ through the design of suitable microchannels and incorporate advanced biosensors with high sensitivity and potential for rapid responses to microenvironmental signals and organ function indicators.11 Recently, more haploid cells were found in the MFD chip of mouse testicular organoids compared with plates via fluorescence-activated cell sorting.12 In addition, this study established that under the same medium composition, the haploid gametogenesis efficiency of a dynamic culture with a microfluidic chip was nearly double that of a static culture. Meanwhile, multi-organ-chip co-cultures of human liver and testis organoids were maintained by the MFD in a steady state for a minimum of 1 week, and the co-cultures reproduced specific natural and drug-induced liver–testis systemic interactions.13 These studies enable the implementation of MFD technologies in future therapeutic strategies for pre-pubertal male fertility preservation and adults with maturation arrest and serve as a platform for drug and toxin testing. However, reports on the use of the MFD during in vitro culture of human testicular tissues are lacking.
Polydimethylsiloxane (PDMS) is characterised by gas permeability,14 no biotoxicity,15 and high plasticity, serving as the main material of the MFD since 1990.16 PDMS is a suitable culture condition for the long-term maintenance of mouse testis tissue in vitro.10,17 In the present study, we manufactured a human testis-on-a-chip (hToC) with PDMS for a human-derived seminiferous tubule culture in vitro. Pillars were designed around the tissue space in the MFD to allow better penetration of nutrients from the flowing medium into the tissues while reducing damage from the fluid shear force. The appropriate gaps between these pillars allow the nutrients to slowly penetrate the tissue. Meanwhile, the PDMS material has good air permeability that leverages the mimicry of the organ-specific tissue–tissue interfaces and physical microenvironments, including fluid flow and air–liquid interfaces.5,14
In this study, we first placed human seminiferous tubules on the chip, whereafter their structure, activity, and functions were maintained and evaluated over two weeks. The tissue on the chip exhibited stable secretion levels of testosterone (T) and inhibin B following regulation by gonadotropin. Moreover, the chip has been proved to be a sensitive platform for reproductive pharmacology and toxicology testing with the chemotherapy drug busulfan (a drug known for its reproductive toxicity).18 This in vitro culture system is valuable in basic studies to reveal the detailed mechanisms behind male reproduction and has significant clinical application prospects to prevent and rescue male infertility. In summary, the human testicular tissue chip has the potential to be an in vitro pharmacology and toxicology platform for fertility rescue after chemotherapy in adolescent male patients with tumours and to screen toxic reproductive compounds.19,20
Experimental section
Chip fabrication
The chip is comprised a PDMS layer, a glass layer, and upper- and lower-layer fixtures. The PDMS layer is the most crucial component of the chip. Its elaborate structure was designed using computer-aided design software, and the corresponding patterns of the structure were etched on a photomask using photolithography technology. A negative photoresist material (MicroChem, China, SU-82100) was then poured onto a 4-inch wafer, and the coating was rotated to achieve a thickness of 250 μm required for the microchannel. Next, it was pre-baked in two stages: 65 °C and then 95 °C. The time for each stage depends on the thickness of the SU-8 photoresist layer. The material was treated with ultraviolet light through a photomask for a few seconds to cure the channel pattern and then baked in two stages as described above to complete the curing reaction. Finally, the unexposed part was dissolved in SU-8 by incubating for 10 min in SU-8 degumming solution (MicroChem) and then rinsing for 3 min in isopropanol (HUSHI, China, 40064360) to form a master to produce the PDMS layer. The production of each device involved mixing the PDMS prepolymer and curing agent (Maito, USA, RTV615) at a ratio of 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 w/w and pouring it on the master. The PDMS was cured in a drying oven at 85 °C for 30 to 40 min, followed by peeling off the master and cutting to obtain the PDMS layer (Fig. 1).
:1 w/w and pouring it on the master. The PDMS was cured in a drying oven at 85 °C for 30 to 40 min, followed by peeling off the master and cutting to obtain the PDMS layer (Fig. 1).
 | ||
| Fig. 1 Preparation of the polydimethylsiloxane (PDMS) layer. | ||
The glass layer was provided by Citoglas (China). The upper- and lower-layer fixtures were made of acrylic (Mitsubishi, Japan) that can fix the PDMS and glass layers in the middle. The upper fixture carried a 15 ml centrifugal tube (BIOFILL, China) as a liquid storage tank. The inlet and outlet were connected to bacterial filters (Deutschland, Millex-GP, SLGP033RB) to filter the air and balance the air pressure. The chips were sterilised with ethylene oxide gas and stored in a sterilisation bag.
Tissue sources
All tissue samples were obtained with informed consent from the patient, and approval was obtained from the Ethics Committee of Changhai Hospital of Shanghai (No. CHEC2021-086). All sample information is recorded in Table S1.†Culture of testis tissue and experimental grouping
Part of the human testis sample received by the laboratory was placed in 4% paraformaldehyde for control experiments. According to the standard gas–liquid interphase method,21 testis tissue fragments, 1–3 mm in diameter, were placed on an agarose gel half-soaked in medium. The remaining samples were cut into 900 × 1000 μm micropieces for the culture on the chips. The samples were set in the tissue space of the PDMS layer, immediately bonded to the glass layer, and secured using upper and lower fixtures. The basic medium (BM) for the tissue culture was Dulbecco's modified Eagle's medium (DMEM, Gibco, C11995500BT) supplemented with 10% KnockOut™ Serum Replacement (KSR) (Gibco, 12618013), and 1% penicillin–streptomycin solution (Gibco, 15140-122). The flow rate of the culture medium was 1 ml per day.For the experimental grouping, fresh tissue was used as the control, and the experimental tissue was cultured after 2 weeks to verify the maintenance effect of the chip culture system on human testicular tissue in vitro. The experiment to verify the reactivity of testis tissue on the chip to gonadotropin regulation used the tissue cultured in BM as a control group, and the tissue cultured in BM with 20 mIU ml−1 recombinant follicle-stimulating hormone (rFSH) (GLPBIO, GP21254) and 1 ng ml−1 recombinant luteinising hormone (rLH) (GLPBIO, GP21254, and GC39581) was used as the experimental group (gonadotropin regulation group). The tissues cultured in BM with a concentration of 1 × 10−4 M busulfan for 24 h were used for the experiment, and the tissues cultured in BM without busulfan served as the control to explore the potential of the testicular tissue on a chip as a pharmacological and toxicological platform.18 Samples were obtained from the same patient for the control and experimental groups in each experiment.
Histological examination
Human testicular tissues and fresh tissues were harvested for analysis after two weeks of culture. Then, the tissues on the chip were fixed in 4% paraformaldehyde for 24 h at 25 °C, dehydrated through a graded series of alcohol: 50, 70, 80, 95, and 3 × 100%, embedded in paraffin wax, and cut at 5 μm thickness using a microtome. The slides were stained with haematoxylin and eosin (H&E), dehydrated through a graded series of alcohol washes, and soaked in xylene for 10 min. The slides were mounted using neutral gum. Finally, the H&E-stained slides were visualised under a NanoZoomer s60 light microscope (Hamamatsu, Japan, C13210-01).Western blotting
The tissue on the chip was cleaved, and the total protein was extracted using a mixture of RIPA (pc101, elegant enzyme) and a protease inhibitor (grf101, elegant enzyme). The lysate was centrifuged at 10000 rcf for 10 min at 4 °C, and the supernatant was obtained. Protein concentration was determined using a bicinchoninic acid protein assay kit (zj101, Elegant enzyme). The samples were separated using 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis and transferred onto a polyvinylidene difluoride (PVDF) membrane. After sealing with 5% bovine serum albumin for 1 h, the membrane was washed with tris-buffered saline containing Tween (TBST) three times (7 min each time). Primary antibodies (Table S2†), diluted in TBST, were added to the PVDF membranes and incubated overnight at 4 °C. The membranes were washed with TBST thrice (7 min each time), and the secondary antibody coupled with horse radish peroxidase (HRP) diluted in TBST was added to the PVDF membranes and incubated at 25 °C for 1 h. The membranes were washed thrice with TBST (10 min each time), and the immune response was detected through enhanced chemiluminescence. Protein expression levels were standardised using β-actin. The proteins were visualised using Blodtight Western Chemiluminescent HRP Substrate (Bioworld, Visalia, CA, USA), and images were captured using a Tanon-5200 Chemiluminescent Imaging System (Tanon Science and Technology, Co., Ltd., Shanghai, China). The intensities of bands were quantified using ImageJ version 1.53t software (National Institutes of Health, MD, USA).
Immunofluorescence
The slides loaded with tissue sections were processed in 1% Triton-X for 20 min. The nonspecific adhesion site was blocked for 60 min at 25 °C using 3% bovine serum albumin. Then, the primary antibodies were incubated overnight at 4 °C, and the secondary antibodies connected by AlexaFluor488 were incubated at 25 °C for 1 h. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) and incubated for 10 min at 25 °C in the dark. The stained sample was irradiated using a fluorescence exciter to produce fluorescence, and the image was captured using a fluorescence microscope (Leica, Germany, MICA). The primary antibodies were anti-DDX4 (Abcam, ab13840), anti-SOX9 (Abcam, ab185966), anti-STAR (Proteintech, 12225-1-AP), anti-PLZF (Santa Cruz, sc-28319), and anti-STRA8 (Abcam, ab217380). DDX4, PLZF, STRA8, SOX9, and STAR are markers of germ cells, spermatogonia cells, secondary spermatocyte cells, Sertoli cells, and Leydig cells, respectively. In order to perform quantitative analysis of cells, the quantities of DDX4+, PLZF+, STRA8, and SOX9+ cells in each cross-section of the testis were tallied in five randomly selected fields per section. For each group, analysis was conducted on a minimum of three sections obtained from separate experiments.Transmission electron microscopy (TEM)
Testicular tissues were collected and fixed with 0.1 M phosphate buffer containing 2.5% glutaraldehyde (Servicebio Wuhan, China) and then sequentially stained with 1% osmium tetroxide, 2% uranyl acetate solution, and lead citrate (all from Servicebio). Embedded samples were sectioned and visualised using a transmission electron microscope (Hitachi, Tokyo, Japan). Observations of fresh human testicular tissue and human testicular subject tissue were subjected to two weeks of culture on the chip.BTB integrity assay
The integrity of the BTB was assessed in vitro using a biotin tracer, followed by analysis with fluorescence microscopy. Seminiferous tubules were incubated with EZ-Link Sulfo-NHS-LC-biotin (10 mg mL−1 in PBS/0.01 M CaCl2, Thermo Fisher, Waltham, MA) according to the manufacturer's instructions, ensuring that the ends of the tubules were not submerged. The samples were incubated at 37 °C in the dark for 30 minutes. The treated tubules were then cryosectioned into 10 μm sections. After fixation and blocking following standard protocols, the testicular cryosections were incubated with Alexa Fluor 568-conjugated streptavidin and DAPI at room temperature for 2 hours. Finally, biotin was visualized using fluorescence microscopy.Hormone level assay
BM containing rFSH (20 mIU ml−1) and rLH (1 ng ml−1) was used for the culture of both chip and statically cultured testicular tissues, while BM without rFSH (20 mIU ml−1) and rLH (1 ng ml−1) was used for the chip-based culture as a control. The culture medium (2 ml) was collected every two days. The concentration was measured using ELISA kits for inhibin B (Cloud-Clone Corp., CCA760Mu) and T (Nanjing Jiancheng Bio-Engineering Institute, A019-2-1). The reagents for each kit were prepared according to the manufacturer's instructions.Flow cytometry
Testicular tissue was dissociated into single cells using DNase I (20 mg mL−1) (Worthington, LS002139), collagenase type I (100 mg mL−1) (Worthington, LS004194), and trypsin-EDTA (0.25%). Staining was performed according to the instructions of the cell cycle staining kit (Lianke Bio, CCS012). A cell suspension with a concentration of 1 × 106 was prepared by adding 1 mL of DNA staining solution and 10 μL of permeabilization solution, followed by vortexing for 5–10 seconds. The samples were incubated in the dark at 25 °C for 30 minutes. Flow cytometry (Miltenyi MACSQuant 10 Analyzer) detection was performed using the lowest sample injection speed, and the results were further analyzed using FlowJo 10.4.Terminal deoxynucleotidyl transferase dUTP nick end labelling (TUNEL) staining
Cell apoptosis was evaluated through TUNEL staining. Tissues were cultured for two weeks, fixed in 4% paraformaldehyde overnight, washed with PBS, dehydrated with ethanol, and embedded in paraffin wax. Five micrometre-thick sections were prepared and rehydrated for TUNEL staining. The sections were incubated with proteinase K (20 μg ml−1) for 15 min at 25 °C. Subsequently, sections were washed thrice with PBS and incubated with 50 μl of the TUNEL mix (Beyotime, 45 μl of fluorescein-dUTP and 5 μl of terminal deoxynucleotidyl transferase [TdT]) for 60 min at 37 °C. They were then incubated with DAPI for 10 min at 25 °C. Images were captured using a MICA fluorescence microscope (Leica Microsystems). Fresh tissues were processed in the same way as the control group. The ratio of TUNEL+ cells was obtained by manually counting at least three fields of view.Tissue LIVE/DEAD assay
The tissue was cultured in the chip for 14 days according to the manufacturer's scheme (LIVE/DEAD activity/cytotoxicity kit for mammalian cells, Thermo Fisher Scientific, USA, L3224). Human testicular tissue cultured for 14 days and fresh human testicular tissue in the control group were labelled with calcein-AM and ethidium homodimer-1, and the dye solution was made by adding 5 μl calcein-AM and 20 μl ethidium homodimer-1 to 10 ml DPBS. The 20× objective lens (10× oculars) image was automatically combined into a wide-field image using MICA imaging (Leica). The same method was used to photograph the fresh tissue.Statistical analysis
All experiments were independently replicated at least three times unless otherwise indicated, and data were presented as the mean ± one standard deviation around the mean (SD). Statistical significance was analysed using the t-test (P < 0.05 indicated significance). All graphs were constructed and statistical analyses performed using GraphPad Prism 9.5.0 (GraphPad Software Inc., La Jolla, CA, USA).Results
Design and application of the chip
The chip was mainly comprised of a PDMS layer and a glass layer in the middle, and it was fixed through two fixtures on the top and bottom (Fig. 2A). The main structures of the chip include microchannels and tissue spaces, both of which are designed on the PDMS layer. The upper fixture included a medium reservoir and a medium outlet. The medium is driven by hydrostatic pressure, flows horizontally through the microchannels on both sides of the tissue space, and spreads to the tissue through the gap between the pillars to provide nutrients; oxygen permeates into the tissue through the PDMS material (Fig. 2B). All elaborate structures (including microchannels, tissue space, and medium inlets/outlets) are designed on the PDMS layer. The depth of the microchannel and tissue space in the PDMS layer was designed to be 250 μm to accommodate the diameter of human seminiferous tubules (Fig. 2B). The estimated distance for nutrient diffusion to a sufficient level of supply is approximately 400–500 μm for testicular tissue.17 Therefore, the width of the tissue space was designed to be 900 μm. The microchannels on both sides of the tissue space each had a width of 550 μm (Fig. 2B) and were separated from the tissue space by pillars. The pillars around the tissue space (150 × 150 × 250 μm) were lined at intervals of 100 μm (Fig. 2B). The medium flowing in the chip requires appropriate resistance to ensure a slow and sustained flow;21,22 therefore, the outlet microchannel is designed as a rectangle with a cross-section of 125 × 100 μm and a length of 20 mm (Fig. 2B).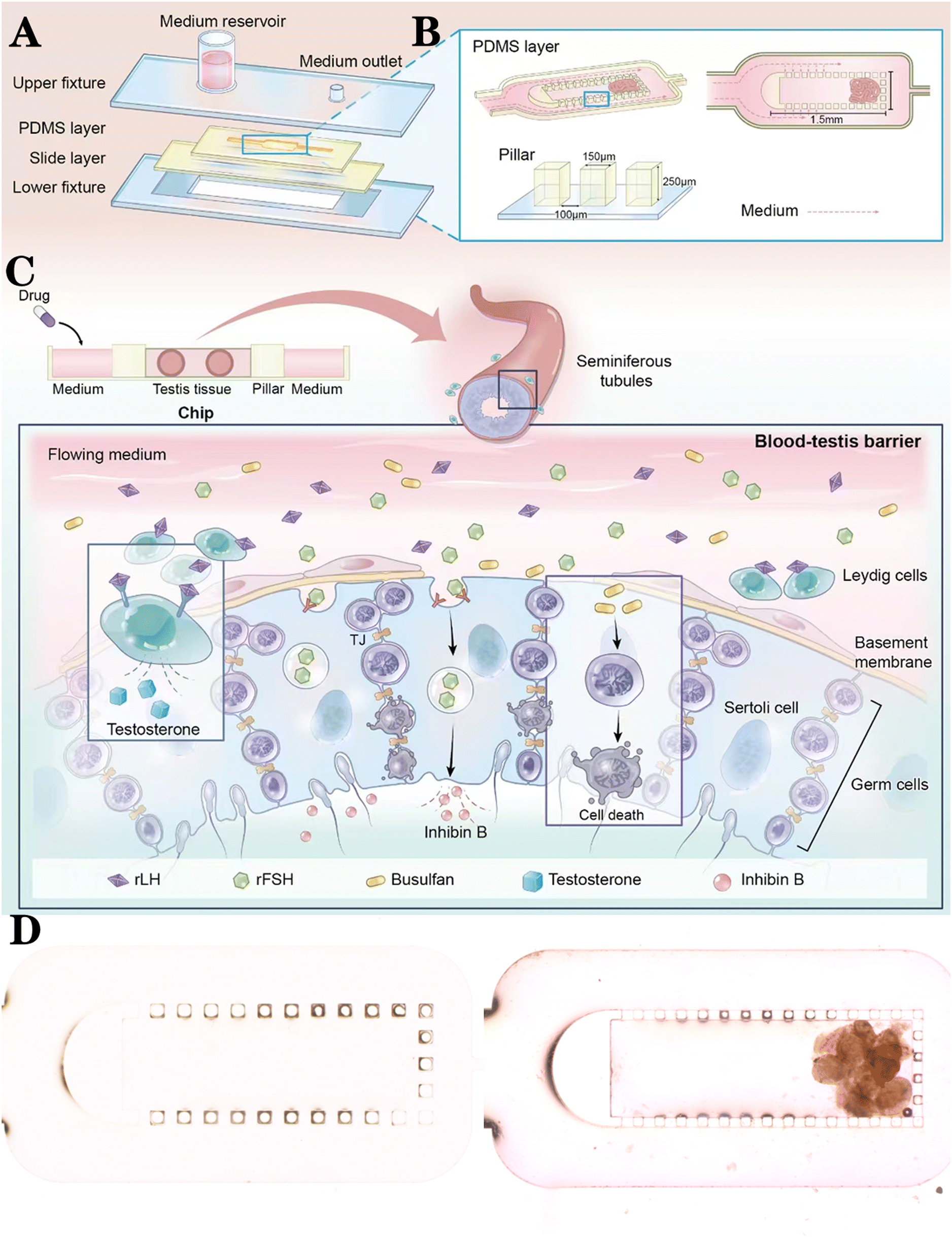 | ||
| Fig. 2 Chip design and use. (A) Each part of the chip. (B) Design of the tissue space, pillars, and microchannels. (C) Cross-section of the chip and a schematic diagram of the pharmacological and toxicological model. (D) Empty chip (left) and chip loaded with human testis tissue (right). Scale bar: 500 μm. | ||
We introduced drugs to the flowing medium, allowing them to gradually infiltrate the testicular tissue and cells (Fig. 2C). For instance, gonadotropins entered the tissue space from the flowing culture medium through osmosis and acted on the testicular tissue. Recombinant luteinising hormone stimulated Leydig cells in the testicular stroma, promoting T production, while rFSH targeted Sertoli cells in the seminiferous tubules, stimulating inhibin B production (Fig. 2C). Additionally, reproductive toxicity drugs like busulfan were used to eliminate germ cells through the flowing culture medium, creating corresponding toxicological injury models (Fig. 2C).
The daily volume of the collected culture medium and the initial hydrostatic pressure were consistent with the function curve of the mathematical model (Table 1).
| Statistics of the medium flow simulation | |||||||
|---|---|---|---|---|---|---|---|
| Sample | Daily liquid collection (ml) | ||||||
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 | |
| 1 | 2.49 | 2.43 | 2.37 | 2.41 | 2.36 | 2.45 | 2.48 |
| 2 | 1.01 | 1.03 | 0.95 | 1 | 0.99 | 0.98 | 1.07 |
| 3 | 0.53 | 0.51 | 0.55 | 0.43 | 0.5 | 0.49 | 0.52 |
| Day 8 | Day 9 | Day 10 | Day 11 | Day 12 | Day 13 | Day 14 | |
|---|---|---|---|---|---|---|---|
| Initial liquid level height of sample 1: 20 mm. Initial liquid level height of sample 2: 8 mm. Initial liquid level height of sample 3: 4 mm. | |||||||
| 1 | 2.43 | 2.38 | 2.38 | 2.37 | 2.44 | 2.41 | 2.5 |
| 2 | 1.01 | 1.08 | 1.11 | 1.08 | 0.92 | 0.97 | 0.99 |
| 3 | 0.44 | 0.51 | 0.49 | 0.5 | 0.51 | 0.54 | 0.52 |

|
|||||||
Comparison of the chip culture system and static culture
Maintaining tissue morphology and structure is critical for evaluating tissue culture. The human testis tissue on the chip was photographed several times during two weeks of in vitro culture using an inverted microscope (Fig. 3A). The morphological structure and size of the testicular tissue did not significantly change during the in vitro culture period. HE staining results showed that after two weeks of culture, both the chip-based and static culture groups maintained a structure and morphology of human testicular tissue similar to that in vivo. | ||
| Fig. 3 Tissue morphology, quality, and endocrine function during chip-based culture. (A) Morphology of testis tissue on the chip under a microscope. Scale bar: 500 μm. (B) Haematoxylin and eosin (H&E) staining of fresh (left), statically cultured for 14 days (center), and chip-cultured for 14 days testicular tissue (right). Scale bar: 100 μm. (C) Representative images show immunofluorescence analysis of germ cell marker DDX4 (yellow), Leydig cell marker Star (purple), and Sertoli cell marker SOX9 (red) and their statistical analysis (D). Scale bar: 100 μm. (E) Inhibin B and testosterone levels were measured at different time points (2nd–14th groups); data represented as mean ± SD (*p < 0.05). Apoptotic cells (TUNEL+) are shown in green, and statistical results are presented (F, *p < 0.05, **p < 0.01). | ||
Germ cells, Sertoli cells, and Leydig cells are the three main types of cells in testicular tissue. Analysis of fresh tissue and tissue cultured for two weeks indicates that the germ cells and Sertoli cells in the statically cultured testicular tissue are slightly lower than in the other two groups, but the difference is not statistically significant (Fig. 3C and D). Hormones are crucial indicators for evaluating the endocrine function of the testes. The levels of testosterone and inhibin B in the culture medium were measured, and the results indicated that testicular tissue cultured on the chip secreted higher levels of testosterone and inhibin B (Fig. 3E). Apoptosis in these groups was further assessed by TUNEL staining (Fig. 3F). Quantitative analysis of fluorescent cell counts showed an apoptotic cell rate of 4.23% in the fresh group, 7.57% in the static group, and 5.17% in the chip group. No statistically significant difference was observed between these two groups (Fig. 3F).
Assessment of cell composition, viability, and function of chip culture system
Germ, Sertoli, and Leydig cells are crucial in the testis. The analysis of 2 weekthe chip-based culture tissue showed that 37.53 ± 2.65% of cells expressed DDX4 (indicative of germ cells), 23.63 ± 3.49% expressed SOX9 (indicative of Sertoli cells), and cells expressed STAR in the interstitial cells (indicative of Leydig cells). Meanwhile, 41.47 ± 1.56% of cells expressed DDX4, and 23.07 ± 4.18% expressed SOX9 in the interstitial cells before culture (Fig. 4A–C). Immunofluorescence staining results showed that the proportion of germ cells, Sertoli cells, and Leydig cells did not significantly change in testis tissue samples after 2 weeks of chip-based culture. The ultrastructure of seminiferous tubules (such as the blood–testis barrier and mitochondria) plays an important role in maintaining the testis microenvironment. The testis tissue maintained the same complete blood–testis barrier and normal mitochondrial morphology after 14 days of in vitro chip-based culture, according to TEM (Fig. 4D). Immunofluorescence biotin permeation experiments also confirmed that both groups maintained intact BTB (Fig. 4E). | ||
| Fig. 4 Composition, activity, and function of testis tissue cultured on the chip. Representative images (A–C) show immunofluorescence analysis of germ cell marker DDX4 (yellow), Leydig cell marker STAR (purple), and Sertoli cell marker SOX9 (red) in the experimental (right) and control groups (left). Scale bars: (A and C) 100 μm, (B) 50 μm. Statistical results indicate germ cells (DDX4+) (A) and Sertoli cells (SOX9+) (B) in fresh and chip groups (*p < 0.05). Transmission electron microscopy (TEM) results of fresh (left) and chip (right) groups. Red arrows: blood–testis barrier (BTB); purple arrows: SSC; blue arrows: spermatocyte; yellow arrows: SC. Scale bar: 20 μm (low) scale bar: 2 μm (high) (D). Seminiferous tubules surrounded by fluorescent biotin (E). Bright field images (F) depict living (green) and dead (red) cells. Scale bar: 250 μm. Apoptotic cells (TUNEL+) are shown in green, and statistical results are presented (G, *p < 0.05). Western blot (H) showing DDX4 and SYCP3 levels. Grey value ratio analysis is shown in l (*p < 0.05). Flow cytometry of the cell cycle in fresh and chip groups (I). | ||
The entire live/dead cells in these two groups were determined using the polyanionic dye calcein and EthD-1 probe, respectively (Fig. 4F). These dyes produce strong and uniform green fluorescence in living cells and bright red fluorescence in dead cells. Almost all tissues showed strong green fluorescence before and after culture (Fig. 4F). The apoptosis of cells in these groups were further evaluated through TUNEL staining (Fig. 4G). Quantitative analysis of the fluorescent cell count showed that the ratio of apoptotic cells of the chip group was 5.53% and that of the fresh group was 4.16%. There was no statistically significant difference between these two groups (Fig. 4G). These two results showed that the human testis tissue chip culture system can maintain the survival rate of the cultured tissue for up to 2 weeks in vitro.
Meiosis and hormone endocrine function are important physiological functions of testicular tissue. The germ cell marker DDX4 and synaptonemal complex protein 3 (SYCP3) were assessed through western blotting (Fig. 4H). Statistical analysis showed that there was no significant difference in the expression levels of these two markers between these two groups (Fig. 4H). Flow cytometry analysis of the cell cycle showed that chip-based culture maintained the presence of haploid and tetraploid cells in human testicular tissue (Fig. 4I).
Exploring the culture system as a platform for pharmacology and toxicology
Human testis tissues were cultured on chips with 1 × 10−4 M busulfan for 24 h to verify the potential of human testis tissue chips as a platform for reproductive toxicology research.18 The testis tissue of the chip + bus group showed a loss of cells in the seminiferous tubules and a decrease in the thickness of the seminiferous epithelium compared with the chip group, according to H&E staining (Fig. 5A). Furthermore, immunofluorescence assays showed that the germ cell (DDX4+), spermatogonia stem cells (SSC, PLZF+), and differentiated spermatogonia and pre-meiotic cells (STRA8+) in the chip + bus group were markedly decreased (Fig. 5B). On the contrary, the number of Sertoli cells (SOX9+) and Leydig (STAR) cells was not significantly affected (Fig. 5B). Testis tissue exhibited significant germ cell damage following busulfan treatment. Finally, the apoptosis rate of the experimental group (52.80 ± 3.03%) was significantly higher than that of the control group (8.83 ± 1.17%) according to TUNEL staining (Fig. 5C).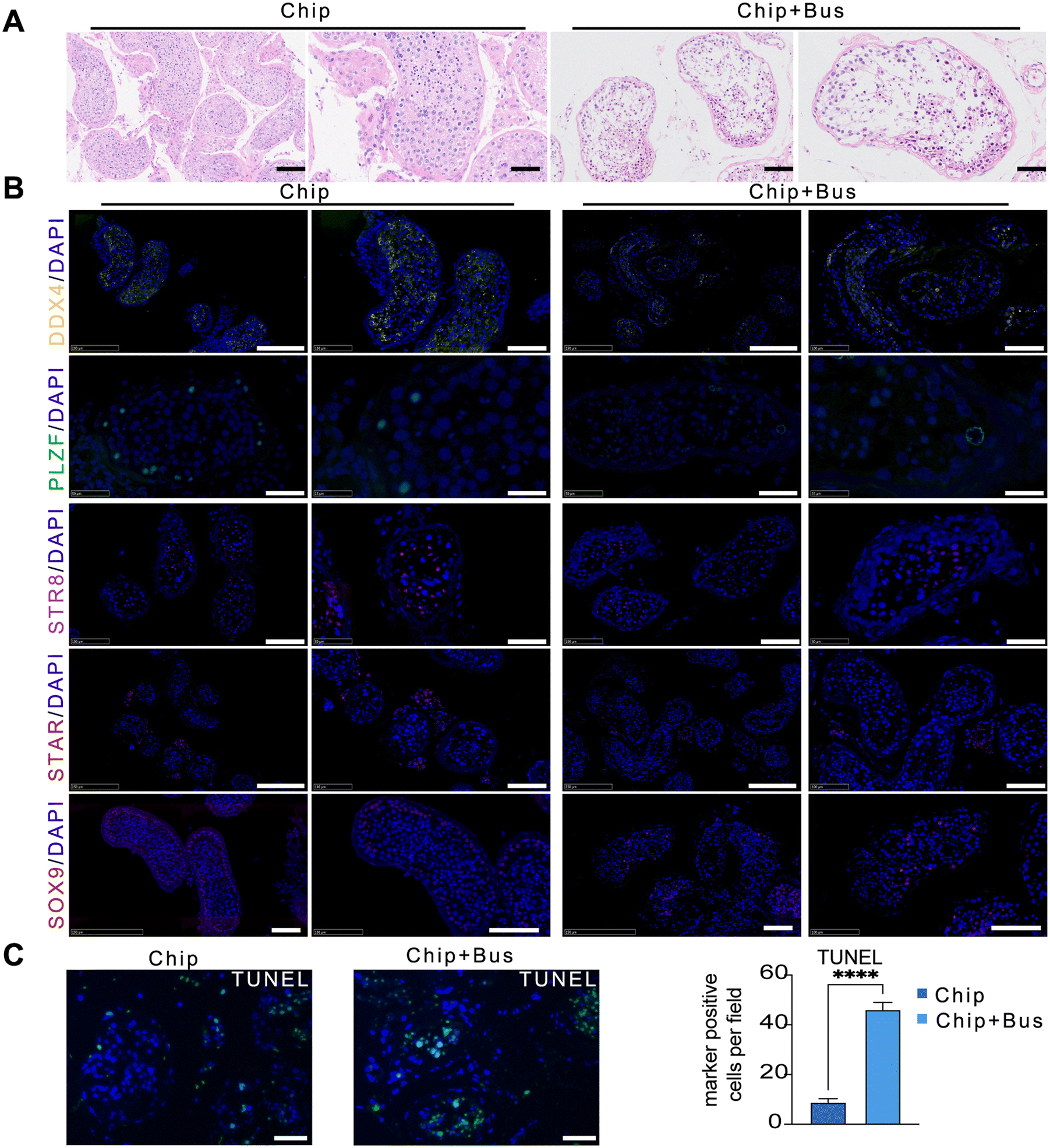 | ||
| Fig. 5 Human testis tissue chip as a platform of reproductive toxicology. (A) Representative H&E staining image of the chip group (left) and chip + bus group (right). Scale bar: 100 μm. (B) Representative images of DDX4 (scale bar: 250 μm and 100 μm), PLZF (scale bar: 50 μm and 25 μm), STRA8 (scale bar: 100 μm and 50 μm), STAR (scale bar: 250 μm and 100 μm), and SOX9 (scale bar: 100 μm and 100 μm) of the chip group (left) and chip + bus group (right). (C) Representative TUNEL staining images of the control group (left) and the experimental group (right). Statistical results of the percentage of apoptotic cells in the control and experimental groups. Data represent the mean ± SD (*p < 0.05, **p < 0.01, ****p < 0.0001). | ||
Discussion
Organ chips can simulate the in vivo environment owing to advances in 3D printing technology and biomaterial technology. In recent years, an increasing number of researchers have attempted to use organ chips to construct novel tissue culture systems in vitro. Interestingly, organ chips constructed with MFDs to culture mouse testis tissue induce meiosis of spermatogonia stem cells to a greater degree than traditional gas–liquid interface methods.16 Therefore, we referred to the mouse testis chip of Yamanaka et al. and designed a novel microfluidic chip device more suitable for human testis tissue and cultured human testis tissue in vitro for up to two weeks.10Initially, we aimed to maintain the activity of tissue cells in vitro for an extended period of time. This mainly depends on sufficient oxygen and nutrient supply. Therefore, we referred to the design of other microfluidic chips and continued to use PDMS as the main material of the chip to ensure that the tissue obtains sufficient oxygen.22 KSR has a variety of activities that stimulate the proliferation and differentiation of stem cells and germ cells;23 therefore, we added 10% KSR to the medium of the chip. In addition, mouse testis tissue chip-based culture requires a 360 μl mg−1 per day medium flow rate to provide sufficient nutrition to maintain its activity and function.22 Therefore, we recorded several sets of data between different initial hydrostatic pressures and flow rates (Table 1) to analyse the functional relationship between them to better control the flow rate of the culture medium in the human testis tissue chip. The diameter of human testicular seminiferous tubules is thicker than that of mouse seminiferous tubules, with a maximum diameter of approximately 250 μm.24 Therefore, the tissue space in this study was set to 250 μm. Moreover, the width of the tissue space was set to 900 μm as the penetration distance of the flowing medium from both sides of the tissue space into the tissue was limited to approximately 400–500 μm.
This design considers the nutritional supply factors and physiological characteristics of thicker human seminiferous tubules. A circle of pillars was designed around the tissue space to reduce the direct impact of the flowing medium on the tissue, reduce the fluid shear force, and maintain the appropriate permeability of nutrients in the medium on both sides of the tissue space to the cultured tissue. At the same time, the tissue can be well fixed in the tissue space for observation. Subsequently, we designed upper and lower fixtures to prevent the operator from directly contacting the inside of the chip while observing it, reduce the possibility of contaminating the tissue, and reduce the pressure generated by local contact to protect the tissue cultured on the chip. Compared with other PDMS chip plasma bonding fixation methods,9 the above-mentioned fixtures enable repeated loading and unloading, so that the tissue cultured on the chip can be taken out for weighing and other operations.
Endocrine function is a critical physiological function of the testes. Leydig cells and Sertoli cells can specifically produce T and inhibin B, respectively; these functions are inseparable from the support of gonadotropins.24 Therefore, we added appropriate concentrations of rLH and rFSH to the medium to maintain the hormone secretion function of the tissue in vitro.25 The culture medium flowing out of the chip was collected to ensure that hormone secretion in the chip was highly stable but not too rapid. The flow rate of the culture medium will dilute the hormone and decrease its concentration. In contrast, a medium flow rate that is too slow can lead to excessive hormone concentration and reduced nutrient supply. Fortunately, we found that the levels of T and inhibin B secreted by the human testis tissue chip were close to the normal reference range of human T (12–35 nmol L−1) and inhibin B (94–327 pg ml−1) in clinical examination when maintaining the flow rate of the culture medium at 1 ml per day.26
Static culture is a classic method for testicular tissue culture.21 Therefore, in this study, we performed multiple comparisons between static culture and chip-based culture. Kanbar27 suggested that static culture is superior to chip-based culture. However, other researchers have presented differing views, with their results indicating that chip-based perfusion culture of testicular tissue can produce more differentiated spermatocytes and spermatids.12 Our study demonstrated that chip-based culture has greater advantages in maintaining the endocrine function and cellular viability of testicular tissue. We believe that the differences in results between laboratories could be due to various factors, such as flow rate of the culture medium, chip material, and fabrication processes.
Endocrine function, meiosis, and spermatogenesis are important physiological functions of the testis. SYCP3 is one of the best markers of early meiosis in vertebrates.28 Our results showed that SYCP3 levels did not significantly change before and after culture. Flow cytometry analysis of the cell cycle showed that chip-based culture maintained the presence of haploid and tetraploid cells in human testicular tissue. This suggested that the human testis tissue chip culture system can maintain the function of early meiosis in human testis tissue in vitro. Organ-chip culture methods were used to induce neonatal mouse testis tissue to produce sperm cells in vitro.17 However, we were unable to carry out similar studies due to ethical problems and restrictions on sample collection. We hope to have future opportunities to conduct further research to help adolescent leukaemia patients that need chemotherapy to preserve testicular tissue and induce spermatogenesis when needed.
We are interested in utilising the chip as an in vitro pharmacology and toxicology platform, similar to many organoid platforms.25 A total of approximately 121145 children and adolescents in China were diagnosed with cancer between January 2018 and December 2020.29 Leukaemia was the most common cancer, accounting for nearly 30% of all diagnosed cases in children aged 1–4. Chemotherapy drugs for the treatment of leukaemia often cause permanent reproductive damage to men, and the construction of corresponding in vitro pharmacological and toxicological models is of great significance to develop therapeutic drugs to preserve the fertility of children with leukaemia. Hence, busulfan, a conventional chemotherapy drug known for its reproductive toxicity and frequently utilised to create male infertility mouse models for chronic myelogenous leukaemia treatment,30 was employed in this study to assess the feasibility of the human testis tissue chip culture system as an in vitro pharmacological and toxicological platform. The testicular tissue on the chip showed very sensitive reproductive toxicity to busulfan by evaluating the number of germ cells and apoptosis. The apoptosis rate of germ cells was higher than that in the static culture at the same concentration and administration time.30 This may be due to the effect of the flowing medium.
The human testis tissue chip also has advantages compared to the classical method of testicular tissue culture in vitro. Testis tissue fragments with a diameter of approximately 3 mm can be cultured in vitro on an agarose gel,31 which is a classical method of testis tissue culture in vitro. In comparison, our designed human testis tissue chip requires a much smaller sample volume since it is less than 900 μm wide, no more than 250 μm thick, and approximately 0.9–1.5 mm in length. This provides better sample utilisation and more promising clinical application prospects. Testicular sperm aspiration is a commonly used technique in the clinical diagnosis and treatment of male infertility. It is widely used owing to its small size and convenience, but it is also limited by the trace testis tissue sample size. Therefore, it can only be used to evaluate whether patients have sperm or obtain a small amount of sperm for intracytoplasmic sperm injection treatment.32 The utilisation of human testis tissue chip technology is anticipated to analyse residual tissue samples from testicular sperm aspiration comprehensively. This approach enables a detailed assessment of the physiological and pathological conditions of various cells within patients' testicular tissues. It can also provide patients with a platform of reproductive pharmacology with personalised diagnosis and treatment value and provide a novel in vitro research tool for reproductive medicine researchers.
Conclusions
The human testis tissue chip was developed as a new tool to cultivate human testis tissue in vitro. Its advantages include high sample utilisation, a strong visualisation effect, and a stable and good maintenance effect on the morphology and function of cultured tissue. Furthermore, it can serve as a sensitive in vitro pharmacological and toxicological platform. These characteristics will make it easier for clinical transformation and play a role in the fields of individual diagnosis and treatment of male reproduction, research of male reproductive physiology, male reproductive pharmacology, and toxicology platforms.Data availability
The data supporting this article have been included as part of the ESI.†Author contributions
JMS: conceptualisation; data curation, writing – original draft, writing – review & editing. XLW: conceptualisation; data curation, writing – original draft, writing – review & editing. CHY: supervision. GYR: resources. LW: resources. SGP: resources. BYZ: methodology. WHS: methodology. XG: writing – review & editing. JJ: methodology. YJX: validation. ZWYH: validation. LHW: supervision and funding acquisition. BY: supervision and funding acquisition. ZYL: project administration, funding acquisition, writing – review & editing, and supervision. All authors read and approved the final manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by the Establishment of a Male Fertility Protection System (No. 21JSZ04), the Establishment and Application of Testicular Biochips (No. 23JSZ02), and the National Natural Science Foundation of China (No. U22A20277).Notes and references
- C. L. R. Barratt, et al. , Hum. Reprod. Update, 2017, 23, 660–680 CrossRef PubMed.
- N. Kumar and A. K. Singh, J. Hum. Reprod. Sci., 2015, 8, 191–196 CrossRef PubMed.
- B. M. Gadella and M. A. Ferraz, Reprod. Domest. Anim., 2015, 50(Suppl 2), 61–70 CrossRef PubMed.
- A. Skardal, et al. , Drug Discovery Today, 2016, 21, 1399–1411 CrossRef CAS PubMed.
- S. N. Bhatia and D. E. Ingber, Nat. Biotechnol., 2014, 32, 760–772 CrossRef CAS PubMed.
- A. Jain, et al. , Clin. Pharmacol. Ther., 2018, 103, 332–340 CrossRef CAS PubMed.
- B. Sadeghzadeh Oskouei, et al. , Cell J., 2021, 23, 32–39 Search PubMed.
- S. Palma-Florez, et al. , J. Nanobiotechnol., 2023, 21, 115 CrossRef CAS PubMed.
- P. M. van Midwoud, et al. , Biotechnol. Bioeng., 2010, 105, 184–194 CrossRef CAS PubMed.
- H. Yamanaka, et al. , Biochem. Biophys. Res. Commun., 2018, 500, 885–891 CrossRef CAS PubMed.
- M. Liu, et al. , Biosens. Bioelectron., 2022, 218, 114758 CrossRef CAS PubMed.
- A. AbuMadighem, et al. , Biofabrication, 2022, 14, 3 CrossRef PubMed.
- Y. Baert, et al. , Hum. Reprod., 2020, 35, 1029–1044 CrossRef CAS PubMed.
- L. Xu, et al. , Lab Chip, 2015, 15, 3962–3979 RSC.
- N. Mohd Fuad, et al. , Micromachines, 2018, 9, 116 CrossRef PubMed.
- E. Berthier, et al. , Lab Chip, 2012, 12, 1224–1237 RSC.
- M. Komeya, et al. , Sci. Rep., 2016, 6, 21472 CrossRef CAS PubMed.
- X. Zhang, et al. , Free Radical Biol. Med., 2019, 137, 74–86 CrossRef CAS PubMed.
- G. A. Kanakis and D. G. Goulis, Hormones, 2015, 14, 598–614 Search PubMed.
- X. Zhao, et al. , Toxicology, 2020, 440, 152489 CrossRef CAS PubMed.
- O. A. Trowell, et al. , Exp. Cell Res., 1959, 16(1), 118–147 CrossRef CAS PubMed.
- M. Komeya, et al. , Sci. Rep., 2017, 7, 15459 CrossRef PubMed.
- J. Zhang, et al. , Gen. Comp. Endocrinol., 2014, 205, 121–132 CrossRef CAS PubMed.
- D. Ježek, et al., Normal Morphology of the Human Testis and Epididymis, in Atlas on the Human Testis, ed. D. Ježek, Springer, London, 2013 Search PubMed.
- Y. Yang, et al. , Biofabrication, 2022, 15, 1 Search PubMed.
- G. A. Kanakis, et al. , Maturitas, 2019, 125, 41–44 CrossRef CAS PubMed.
- M. Kanbar, et al. , Front. Physiol., 2022, 13, 884122 CrossRef PubMed.
- X. Zhang, et al. , Nat. Commun., 2021, 12, 1758 CrossRef CAS PubMed.
- X. Ni, et al. , Lancet, 2022, 400, 1020–1032 CrossRef PubMed.
- L. Zhao, et al. , Reprod. Toxicol., 2023, 115, 1–7 CrossRef CAS PubMed.
- O. A. Trowell, Exp. Cell Res., 1959, 16, 118–147 CrossRef CAS PubMed.
- P. Rubino, et al. , Hum. Reprod. Update, 2016, 22, 194–227 Search PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4lc00780h |
| ‡ Co-first authors. |
| This journal is © The Royal Society of Chemistry 2025 |