
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceMilk peptides found in human jejunum induce enteroendocrine hormone secretion and inhibit DPP-IV†
Santiaga María
Vivanco-Maroto‡
 a,
Cristina
Gómez-Marín‡
ab,
Isidra
Recio
a and
Beatriz
Miralles
*a
a,
Cristina
Gómez-Marín‡
ab,
Isidra
Recio
a and
Beatriz
Miralles
*a
aInstituto de Investigación en Ciencias de la Alimentación, CIAL (CSIC-UAM, CEI UAM+CSIC), Nicolás Cabrera, 9, 28049 Madrid, Spain. E-mail: beatriz.miralles@csic.es
bEscuela de Doctorado, Universidad Autónoma de Madrid, C. Francisco Tomás y Valiente, 2, 28049 Madrid, Spain
First published on 29th May 2025
Abstract
The strong effect of protein digestion products on gastrointestinal hormone release is well recognized. However, little is known about the specific characteristics of hormone inducing peptides. A detailed analysis of food-derived peptides remaining in the human intestinal lumen after protein ingestion would constitute a practical strategy for the targeted identification of hormone inducing and DPP-IV inhibitory peptides. In this study, in vivo gastrointestinal resistant peptides derived from casein and whey protein were evaluated in epithelial intestinal cells. The secretion of cholecystokinin (CCK) and glucagon-like peptide-1 (GLP-1) was evaluated in the STC-1 enteroendocrine cell line and the DPP-IV inhibitory potential was assayed in situ using the Caco-2 cell line. Hydrophobic residues at N-terminal positions were crucial for the secretagogue and inhibitory activities, while the presence of multiple glutamic acid residues was shown to be a key trait for CCK secretion. The results appointed the sequence 126TPEVDDEALEKFDK138 from β-lactoglobulin as a strong CCK inducer. Additionally, 94KILDKVGINYWL105, derived from α-lactalbumin, not only promoted GLP-1 secretion but also demonstrated significant DPP-IV inhibitory activity. These findings provide new insights into the functional potential of food-derived peptides, offering promising therapeutic avenues for regulating gastrointestinal hormones and improving metabolic health.
1. Introduction
Dietary proteins, beyond their nutritional role, elicit a range of metabolic effects both in the short and long term, including enhanced thermogenesis, reduced energy intake, and improved insulin sensitivity.1 Among the factors mediating these metabolic effects, hormones secreted by the gut endocrine cells play a major role in the normal postprandial physiology, as part of the gut–brain–pancreas axis.2 Protein digestion products are known to activate enteroendocrine cells through specific sensing receptors and sodium-coupled transporters. Various receptors involved in the detection of protein digestion products throughout the mouse and human gut epithelium have been described. Concretely, receptor GPR93, which is specialized in peptide sensing, shows high expression levels in the mid-intestine, peaking in the distal jejunum.3Some studies using the enteroendocrine cell line STC-1 have shown that cholecystokinin (CCK) was maximally induced by peptides and to a lower extent by free amino acids or intact protein.4,5 More recently, further evidence has been added on the prevailing role of food-derived peptides in the stimulating effects for both CCK and glucagon-like peptide-1 (GLP-1). Thus, whey-derived peptides longer than five amino acids were more effective than di- and tri-peptides on CCK secretion6 and a casein digest elicited a strong GLP-1 response as compared with the same product subjected to complete hydrolysis.7 A number of peptide sequences derived from food proteins i.e. milk proteins,8 bovine haemoglobin,9 egg white ovalbumin,10 tilapia proteins,11 soy β-conglicinin,12 barley13 or buckwheat14 have demonstrated a secretagogue effect on the enteroendocrine cell lines STC-1 and GLUTag, being capable of inducing the release of CCK or GLP-1.
Concerning the characteristics of hormone inducing peptides, some authors have pointed out that peptide sequences with more than five free amino acids and aromatic residues were potent inducers of CCK secretion.6,9 Likewise, we have previously shown that the peptide fraction in the 500–1500 Da range from egg white protein digests was mainly responsible for the GLP-1 secretory effect.10 In order to elucidate the structure–function relationship, some studies have looked at the net charge and hydrophobicity of the peptides. Hence, peptides with hydrophobicity scores between 13 and 27 and isoelectric pH between 6 and 7.9 were reported to present a strong secretory effect on GLP-1.6 Other authors have linked hydrophobicity with enhanced GLP-1 secretion. Komatsu et al.8 suggested that high hydrophobicity scores would contribute to an efficient membrane permeability without the involvement of the peptide sensing mechanisms that have been described for L cells, namely, the peptide transporter 1 and calcium-sensing receptor.15 Lastly, a number of studies have looked at the role of specific amino acids within the sequence. Some kokumi-active peptides containing γ-glutamyl residues have been shown to stimulate CCK and GLP-1 secretion in STC-1 cells by the activation of the calcium-sensing receptor.16 Glutamic acid/glutamine-containing peptides have been recently postulated as important for the reduced fasting blood glucose in db/db diabetic-like mice produced by a casein hydrolysate. The intake of the hydrolysate increased the hepatic metabolites rich in glutamic acid, which acted as substrates for the Krebs cycle, enhancing hepatic glucose consumption.17
On the other hand, the incretin GLP-1 has a short half-life in the organism. Immunohistochemical studies revealed the conversion of GLP-1 (7–36) amide into the inactive (9–36) form by the enzyme dipeptidyl peptidase IV (DPP-IV) in the brush border epithelium as well as in the capillary endothelium supplying the L cells of the porcine intestine.18 Thus, peptide sequences acting as DPP-IV inhibitors are expected to extend the half-life of this incretin. Examples of peptides with DPP-IV inhibitory action exerting glycaemic control through an increase in plasma GLP-1 are available.19–21 Different food-derived peptides capable of inhibiting DPP-IV have been described from very diverse protein sources, with the sizes ranging from 2 to 15 amino acids.22 The study of the position of selected amino acid residues in the peptide sequence and the side-chain properties of known inhibitors has allowed the proposal of some descriptors related to the inhibitory potency. Branched chain amino acids at the N-terminal position and proline at the second N-terminal position are the most prominent examples.23 However, other amino acid residues such as tryptophan appear to play a relevant role. Tryptophan-rich oligopeptides from milk and fish sources have shown inhibitory activity against DPP-IV and high binding affinities to the DPP-IV enzyme by molecular docking.21,24,25 Further experiments have linked these sequences with decreased postprandial blood glucose levels in humans.21 The structure–function features can be used in in silico approaches to predict and facilitate the enzymatic release of these DPP-IV inhibitors.26 However, evidence of these peptides to survive gastrointestinal digestion is still small.
Once the chyme enters the small intestine, the polypeptides are cleaved into oligopeptides and amino acids by proteases of different specificity. The products of endoproteases such as trypsin, chymotrypsin and elastase are further attacked by exopeptidases which remove amino acids from one or the other end of the chain. It has been reported that, in humans, the breakdown products of protein digestion comprise 30% free amino acids and 70% oligopeptides between 2 and 8 residues.27 Although some of them are further hydrolysed by aminopeptidases from the brush-border membranes to free amino acids and di- or tripeptides, there is a great likelihood of resistant sequences to be sensed by specialized receptors in enteroendocrine cells. Thus, a detailed analysis of food-derived peptides identified in the gastrointestinal tract after ingestion would constitute a practical strategy for the targeted identification of hormone inducing and DPP-IV inhibitory peptides. In this study, peptides previously identified in human jejunum aspirates after the intake of milk proteins (casein or whey) with diverse amino acid arrangements were selected based on their occurrence in at least three volunteers.28 The objective was to identify those sequences able to induce CCK and/or GLP-1 release in the enteroendocrine cell line STC-1 and to determine their capacity to inhibit the DPP-IV enzyme in situ using Caco-2 cells.
2. Materials and methods
2.1. Samples/synthetic peptides
Peptide sequences derived from β-casein (6LNVPGEIVE14, 7NVPGEIVE14, 8VPGEIVE14, 9PGEIVE14, 10GEIVE14, 81PVVVPPFLQPE91, 82VVVPPFLQPE91, 85PPFLQPEV92, 89QPEV92, 172LPVPQ176, 173PVPQ176), β-lactoglobulin (β-Lg) (41VYVEELKPTPEGDLEIL57, 94VLDTDYK100, 107MENSAEPEQS116, 125TPEVDDEALEKFDK138) and α-lactalbumin (α-La) (19GGVSLPEWV27, 80FLDDDLTDD88, 94KILDKVGINYWL105, 95ILDKV99, 96LDKVG100) previously identified in human jejunum after casein or whey ingestion28 were chemically synthesized by CSBio Ltd (Shanghai, China). The purity of the peptides was verified by elemental analysis and reverse phase high-performance liquid chromatography coupled with mass spectrometry (LC-MS). The sequence length, theoretical isoelectric point and hydrophobicity of the peptides are given in Table 1.| Protein | Sequence | M w average | Sequence length | Hydrophobicity | Isoelectric point |
|---|---|---|---|---|---|
| β-Casein | 6LNVPGEIVE14 | 969.11 | 9 | 24.97 | 3.1 |
| β-Casein | 7NVPGEIVE14 | 855.95 | 8 | 19.25 | 3.1 |
| β-Casein | 8VPGEIVE14 | 684.79 | 7 | 15.67 | 3.1 |
| β-Casein | 9PGEIVE14 | 642.71 | 6 | 11.65 | 3.1 |
| β-Casein | 10GEIVE14 | 545.59 | 5 | 9.32 | 3.1 |
| β-Casein | 81PVVVPPFLQPE91 | 1221.48 | 11 | 34.09 | 3.3 |
| β-Casein | 82VVVPPFLQPE91 | 1124.36 | 10 | 32.39 | 3.3 |
| β-Casein | 85PPFLQPEV92 | 926.09 | 8 | 27.64 | 3.3 |
| β-Casein | 89QPEV92 | 471.51 | 4 | 2.83 | 3.3 |
| β-Casein | 172LPVPQ176 | 552.68 | 5 | 11.06 | 6.0 |
| β-Casein | 173PVPQ176 | 439.52 | 4 | 3.36 | 6.0 |
| β-Lactoglobulin | 41VYVEELKPTPEGDLEIL57 | 1944.23 | 17 | 41.24 | 3.6 |
| β-Lactoglobulin | 94VLDTDYK100 | 852.95 | 7 | 12.62 | 3.9 |
| β-Lactoglobulin | 107MENSAEPEQS116 | 1121.15 | 10 | 6.59 | 3.0 |
| β-Lactoglobulin | 125TPEVDDEALEKFDK138 | 1635.76 | 14 | 25.40 | 3.8 |
| α-Lactalbumin | 94KILDKVGINYWL105 | 1461.78 | 12 | 38.52 | 9.7 |
| α-Lactalbumin | 95ILDKV99 | 586.74 | 5 | 14.77 | 6.8 |
| α-Lactalbumin | 96LDKVG100 | 530.63 | 5 | 7.87 | 6.8 |
| α-Lactalbumin | 19GGVSLPEWV27 | 943.08 | 9 | 31.47 | 3.3 |
| α-Lactalbumin | 80FLDDDLTDD88 | 1068.07 | 9 | 24.02 | 2.7 |
2.2. Cell assays
STC-1 cells were supplied by the American Type Culture Collection (ATCC CRL3254, Manassas, VA, USA). They were cultured in Dulbecco's modified Eagle's medium (DMEM, Merck, Darmstadt, Germany) containing 4.5 g L−1 glucose and 5 mM L-glutamine supplemented with 100 U mL−1 penicillin, 100 mg L−1 streptomycin, 0.25 μg mL−1 amphotericin (Cytiva, HyClone, Utah) and 10% fetal bovine serum (ATCC, Manassas, VA, USA). Caco-2 cells were supplied by ATCC and cultured in DMEM supplemented with 100 U mL−1 penicillin, 100 mg L−1 streptomycin, 0.25 μg mL−1 amphotericin, 10% fetal bovine serum (Merck, Darmstadt, Germany), and 1% of non-essential amino acids (Merck, Darmstadt, Germany). Both cell lines were grown at 37 °C under a 5% CO2 humidified atmosphere, and once they reached 80% confluence, they were trypsinized and seeded according to each cell study requirement.![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :10; v:v (AlamarBlue™ Cell Viability Reagent, Thermo Fisher Scientific, Waltham, MA, USA) followed by 1 h of incubation. Fluorescence was measured at 590/530 nm excitation/emission wavelengths. HEPES buffer and Triton™ X-100 diluted 1:30; v:v were used as the negative and positive control, respectively. Cell viability experiments were performed in triplicate as well as the fluorescence measurement.
:10; v:v (AlamarBlue™ Cell Viability Reagent, Thermo Fisher Scientific, Waltham, MA, USA) followed by 1 h of incubation. Fluorescence was measured at 590/530 nm excitation/emission wavelengths. HEPES buffer and Triton™ X-100 diluted 1:30; v:v were used as the negative and positive control, respectively. Cell viability experiments were performed in triplicate as well as the fluorescence measurement.
Statistical analysis
The intracellular calcium concentration was compared to the control and within samples by performing statistical analysis using one-way ANOVA with Tukey's post-hoc test for pairwise comparisons. CCK and GLP-1 release was compared to the control and within samples for both concentrations using one-way ANOVA with Tukey's post-hoc test. DPP-IV inhibition was compared within samples and between using one-way ANOVA with Tukey's post-hoc test. The results were considered significant if p < 0.05. GraphPad Prism version 8.0.2 for Windows (GraphPad Software, San Diego, CA, USA) was used.3. Results
The assayed peptides were selected among the large number of peptides previously identified in the human jejunum,28 namely, 362 distinct sequences after casein and 313 distinct sequences after whey ingestion. From them, 185 sequences had been found in at least 3 subjects. Eleven peptides from β-casein, four from β-Lg and five from α-La with common amino acid motifs were selected. When this was not possible, the occurrence of aromatic and hydrophobic amino acids in combination with varying hydrophobicity or isoelectric points has been used as the selection criterion. Table 1 shows the protein fragment, molecular weight, sequence length, hydrophobicity and isoelectric point of the assayed peptides. Cell viability in the enteroendocrine cell line STC-1 was tested and none of the synthetic peptides affected cell viability at the assayed concentrations (ESI Fig. 1†).3.1. Intracellular calcium mobilization of milk peptides identified in human jejunum samples
Prior to the hormone secretion assays, intracellular calcium mobilization was examined in STC-1 cells in the presence of selected peptide sequences, and intracellular calcium was expressed as (RFUmax − RFUmin)/RFUmin. The sequence 41VYVEELKPTPEGDLEIL57 could not be assayed due to its difficulty in dissolving at 10 mM of the necessary stock to reach 1 mM in the well. All β-casein-derived peptides (Fig. 1A–C) increased significantly the intracellular calcium levels after exposure, except fragment β-casein 7NVPGEIVE14. Greater calcium mobilization was observed for β-casein peptide sequences 9PGEIVE14, 85PPFLQPEV92, and 173PVPQ175. Regarding whey-derived peptides (Fig. 1D–F), all samples showed increased intracellular calcium levels except fragment α-La 96LDKVG100.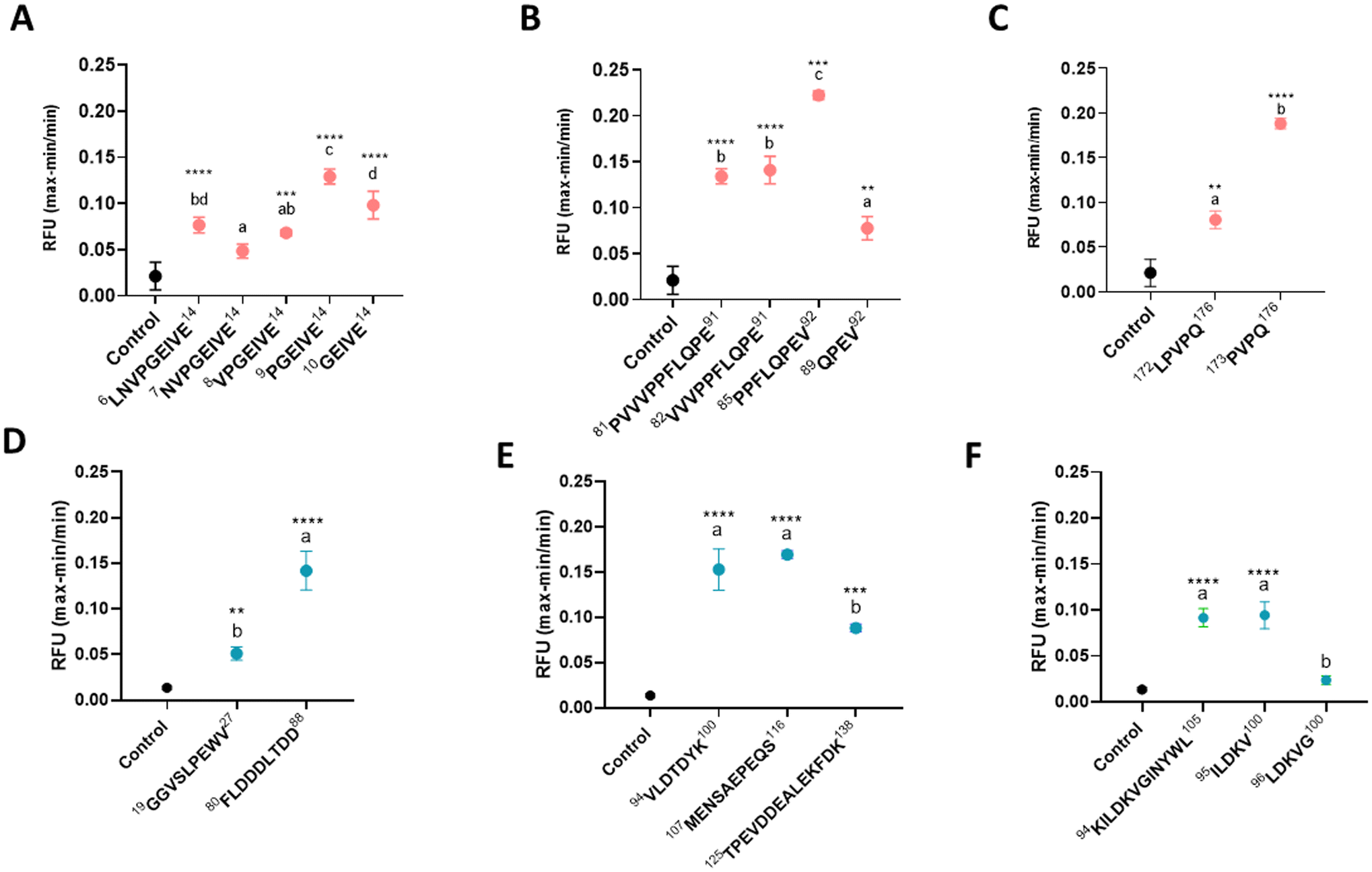 | ||
| Fig. 1 Changes in the intracellular calcium concentration in STC-1 cells exposed to synthetic peptides. Intracellular calcium concentrations were measured by fluorescence. Cells were exposed to synthetic peptides at 1 mM in HBSS buffer containing HEPES and glucose (control), in triplicate. Intracellular calcium was expressed in relative fluorescence units (RFU) as (RFUmax − RFUmin)/RFUmin after exposure to casein (A–C) and whey protein derived peptides (D–F) at 0.5 mM. Error bars indicate SEM (n = 3). Statistical significance compared with the control is indicated (one-way ANOVA with Tukey's post hoc test) by *p < 0.05 and **p < 0.01. Different letters denote statistically significant differences (p < 0.05) between different peptides. | ||
3.2. CCK and GLP-1 release in response to casein and whey peptides in STC-1 cell culture
Fig. 2 shows the CCK levels induced by STC-1 cells in response to peptides assayed at 1 and 0.25 mM. The β-casein peptide sequences 6LNVPGEIVE14, 7NVPEGEIVE14, and 8VPGEIVE14 reached the highest levels of CCK, with values approximately 20-fold the control, showing that loss of the amino acids 6L and 7N at the N-terminal end did not produce alterations in CCK secretion. The subsequent loss of the amino acids 8V and 9P resulted in approximately twice less secretion than that of the longer forms, but still showed significant secretion (Fig. 2A). In other groups from casein-derived peptides, CCK release was moderate, about two-fold the control levels, with significant CCK hormone levels only at the 1 mM concentration as compared to the control. In the β-casein-derived peptide 81PVVVPPFLQPE91 and related forms, the N-terminal loss of 81P and various valine residues slightly reduced the observed levels of CCK. Regarding 85PPFLQPEV92 and the comprised sequence 89QPEV92, the short form increased the response (Fig. 2B). In the β-casein peptide 172LPVPQ176 the loss of 172L increased the CCK release of the resulting fragment, as another example of induction augmented with a lower peptide size (Fig. 2C). Concerning the whey-derived peptides (Fig. 2D–F), all the assayed sequences were able to induce CCK secretion, except the β-Lg 107MENSAEPEQS116 sequence (Fig. 2D). A fragment from β-Lg 125TPEVDDEALEKFDK138 displayed the highest secretion of this hormone (40-fold the control), being the most potent inducer among all the forms tested, although β-Lg 41VYVEELKPTPEGDLEIL57 also showed a remarkable inducing power with 15-fold the control CCK release. These two sequences have in common a relatively long size (14 and 17 amino acids, respectively), an acidic isoelectric point with multiple negative residues and elevated hydrophobicity. By contrast, the α-La-derived sequences showed moderate CCK secretion (Fig. 2E and F). | ||
| Fig. 2 CCK secretion after 2 h of incubation of STC-1 cells with synthetic peptides identified in human jejunum after taking a solution of casein (A–C) or whey (D–F). Synthetic peptides were tested at 1 mM and 0.25 mM. CCK secretion was determined by ELISA. Cell experiments were performed in triplicate, followed by technical duplicates. Error bars indicate SEM (n = 3). Statistical significance compared with the control (C) (one-way ANOVA with Tukey's post hoc test) is indicated by *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001. Statistical significance (p < 0.05) in the comparison between different samples is indicated by different letters. | ||
GLP-1 release was enhanced upon incubation of STC-1 cells with all the studied sequences at the highest concentration (Fig. 3). Remarkably, high values (over 5-fold the control) were observed for all β-casein related peptides derived from 6LNVPGEIVE14, except 7NVPGEIVE14 (Fig. 3A). A less potent response was observed with the β-casein 81PVVVPPFLQPE91 and related forms, all of which achieved levels between 2- and 3-fold the control release, except the 82–91 form (Fig. 3B). In the case of the related forms 172LPVPQ176 and 173PVPQ176, the second one showed a more pronounced GLP-1 release, similar to the behaviour found for CCK secretion (Fig. 3C). Among the whey-derived peptides, the sequence with the highest hormone secretion was the α-La fragment 94KILDKVGINYWL105 that produced 15-fold the control release (ca. 800 pM of GLP-1) (Fig. 3F). Its derived form, 96ILDKV100, induced 10-fold GLP-1 release while 96LDKVG100 further reduced the secretory activity. Other β-Lg or α-La derived peptides assayed reached levels of GLP-1 release between 2- and 5-fold the level of the control secretion (Fig. 3D and E).
 | ||
| Fig. 3 GLP-1 secretion after 2 h of incubation of STC-1 cells with synthetic peptides identified in human jejunum after taking a solution of casein (A–C) or whey (D–F). Synthetic peptides were tested at 1 mM and 0.25 mM. GLP-1 secretion was determined by ELISA. Cell experiments were performed in triplicate, followed by technical duplicates. Error bars indicate SEM (n = 3). Statistical significance compared with the control (C) (one-way ANOVA with Tukey's post hoc test) is indicated by *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001. Statistical significance (p < 0.05) in the comparison between different samples is indicated by different letters. | ||
3.3. DPP-IV inhibitory effect of casein and whey peptides in Caco-2 cell culture
The DPP-IV inhibitory effect of the assayed peptides was evaluated in cells using 2 mM, 0.2 mM, and 0.02 mM peptide concentrations in the Caco-2 cell culture (Fig. 4). Several peptides were shown to strongly inhibit the DPP-IV activity, namely 172LPVPQ176 from β-casein, 107MENSAEPQS116 from β-Lg, and 80FLDDDLTDD88 and α-La 125TPEVDDEALEKFDK138 from α-La, inhibiting more than 90% of the enzyme activity and the higher assayed concentration. Other sequences with notable DPP-IV inhibition were β-casein 6VPGEIVE14 (88%), 85PPFLQPEV92 (83%), and 89QPEV92 (86%). Moreover, the inhibitory effect was assayed in four β-casomorphin-7 related forms which were also found to be resistant to in vivo digestion and described as CCK and GLP-1 inducers in our previous study.7 Two of these sequences, β-casein 60YPFPGPI66 and 60YPFPGPG64, showed 88% DPP-IV inhibition (ESI, Fig S2†).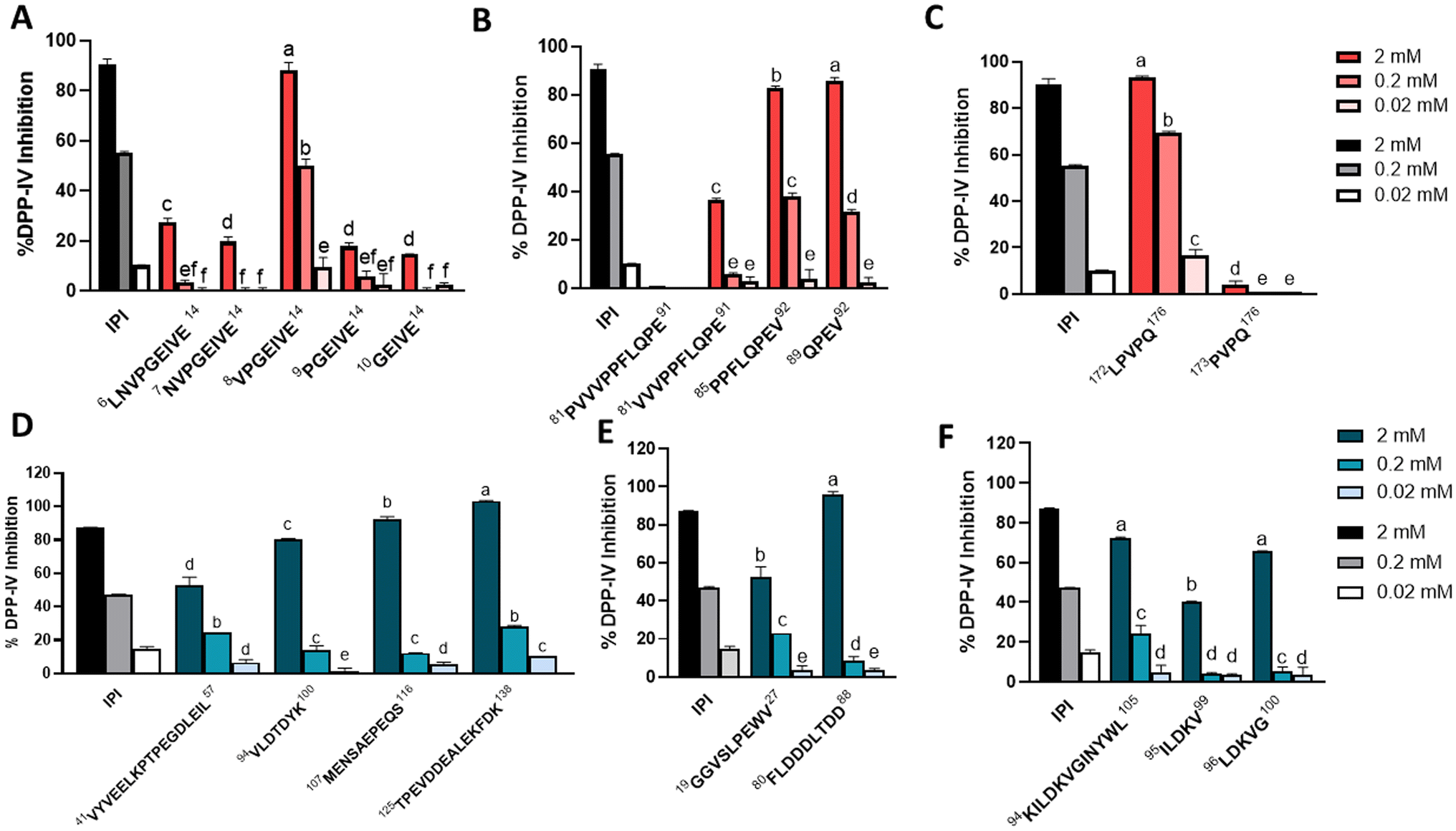 | ||
| Fig. 4 DDP-IV enzyme inhibition in the presence of casein peptides (A–C) and whey peptides (D–F) at different peptide concentrations (2.00, 0.20, and 0.02 mM) using Caco-2 cells. Error bars indicate SEM (n = 3). Statistical significance compared between different samples is indicated by different letters (one-way ANOVA with Tukey's post hoc test). IPI is the positive control. | ||
4. Discussion
There is still limited knowledge regarding the specific characteristics of anorexigenic hormone-inducing peptides. An essential requirement to induce signalling in the intestinal epithelium is to be resistant to the digestive enzymes, a prerequisite that was fulfilled by the assayed sequences as they had been identified in human jejunum aspirates after casein or whey protein intake. In addition to that, peptide features developed in various studies for the induction of CCK and GLP-1 by enteroendocrine cells include size, net charge, hydrophobicity, or position in the chain of selected amino acid residues.In our work, we have evaluated the response of sequences with lengths ranging from 5 to 17 amino acids and isoelectric points throughout a wide pH range, between 2.7 and 9.7. The highest hormone release has been observed for sequences that displayed high hydrophobicity, over 10 GRAVY units, regardless of the isoelectric point, a peptide feature that had been previously been linked to elevated hormone secretion in vitro.8
In some cases, a greater hydrophobic character was linked to a higher secretory effect, especially for GLP-1. Thus, the 95ILDKV99 fragment of α-La exhibited a stronger GLP-1 secretory effect compared to the related 96LDKVG100 fragment with the first displaying a hydrophobicity value twice as the second. This can also be supported by low intracellular calcium mobilization for the last sequence. Interestingly, the sequence 94KILDKVGINYWL105 comprising them displayed the highest GLP-1 inducing activity with four-fold theoretical hydrophobicity. All these fragments share the occurrence of the hydrophobic leucine and isoleucine as N-terminal amino acids, although the last form displays the basic residue lysine at the N-terminal. By contrast, the β-casein fragment 89QPEV92 showed similar GLP-1 induction to its related sequences but displayed markedly lower hydrophobicity. This points to the contribution of other descriptors.
In terms of peptide length, a previous study by our group has shown an increase in GLP-1 secretion with decreasing size of peptides related to the opioid sequence β-casomorphin (β-casein 60–66) in which the 60YPFPG64 motif, which includes the aromatic amino acids tyrosine and phenylalanine, was conserved.7 The size did not correlate with the CCK inducing activity in these β-casomorphin-related fragments, although the smaller size fragment, with five amino acids, sharply decreased CCK induction. Interestingly, in the sequences of β-casein retaining the 10GEIVE14 motif, the CCK induction trend points again to higher sensing with a size over seven amino acid residues. It should be noted that the maximal CCK response in this study has been observed for peptides with 14 and 17 residues. CCK is known to be induced by amino acids such as phenylalanine.32 This hormone is also induced by a 35 amino acid luminal cholecystokinin releasing factor in the intestine and direct cellular effects have been shown in STC-1 cells in a calcium dependent manner.33 Long food-derived peptides have similarly shown activity on CCK secretion. Thus, intraduodenal infusion of the 13 amino acid fragment 51–63 from β-conglycinin, VRIRLLQRFNKRS, inhibited food intake and raised the portal plasma CCK concentration in Sprague-Dawley rats. In the study of the mechanism, the authors remarked that this sequence strongly bound the brush border membrane and acted as a substrate of trypsin in the lumen due to multiple nonadjacent arginine residues in the peptide structure.12
Alternative sequence features such as the occurrence and location in the peptide chain of the amino acid residues, glutamic acid, and tryptophan have been assessed in the observed results. The above-mentioned group of β-casein fragments that shared the motif 10GEIVE14 can be considered an outstanding set, in terms of both CCK and GLP-1 secretion. They present as common traits two glutamic acid amino acid residues in the peptide chain. Other potent secretors revealed in this study, especially for CCK secretion, are β-Lg 41VYVEELKPTPEGDLEIL57 and 125TPEVDDEALEKFDK138, also with multiple glutamic acid residues. Other CCK inducers having survived gastrointestinal digestion with glutamic acid in the peptide chain derive from buckwheat (PAFKEEHL) or egg white (VLLPDEVSGL).10,14 Moreover, some di- and tri-peptides containing either one or two consecutive γ-glutamyl residues have been shown to stimulate CCK and GLP-1 secretion in STC-1 cells.16 These peptides share with the present studied sequences a high resistance to gastrointestinal digestion through their unique γ-bond.34
As for tryptophan, the behaviour of α-La 94KILDKVGINYWL105 in relation to GLP-1 secretion is striking. It shows a very potent inducing activity while none of the related forms, 95ILDKV99 and 96LDKVG100, deprived of tryptophan, reaches the values observed for the parent sequence, which points to a prominent role of this aromatic residue. It is worth mentioning that tryptophan has been also found in other peptides with potent hormonal secretagogue effects. For instance, a seven amino acid long sequence showing N-terminal tryptophan, the lysozyme derived peptide123 WIRGCRL,129 has previously shown a potent GLP-1 secretory activity.30 It is interesting to note that tryptophan in the penultimate C-terminal can also have a hormone secreting role, as the strongly inducing sequence 94KILDKVGINYWL105 reveals. Another outstanding sequence, fragment 19GGVSLPEWV27 from α-La, contains both tryptophan and glutamic acid at adjacent positions. It is remarkable that other surviving peptides in the human aspirates related to this sequence displayed at least one glutamic acid residue, such as fragments 17GYGGVSLPE25 or 19GGVSLPEW26. The well-known metabolic effects of whey proteins35 could be linked with the profile of surviving peptides at the intestinal level, with a prominent presence of glutamic acid in peptides, as previously observed.28
The assayed sequences have shown highly varying values of inhibition of the DPP-IV enzyme in the model system with Caco-2, ranging from no inhibition in the case of fragment 81PVVVPPFLQPE91 from β-casein to complete inhibition for β-Lg 125TPEVDDEALEKFDK138. One of the structural features important for this inhibitory activity is the presence of proline at the second N-terminal position, and some of the peptides with the greatest effect, such as the β-casein derivatives 8VPGEIVE14, 85PPFLQPEV92, 89QPEV92 or 172LPVPQ176, or the β-Lg derivative 126TPEVDDEALEKFDK138, showed this feature. There have been some studies aimed at identifying the structure–activity relationship from diverse peptides displaying DPP-IV inhibitory activity. In addition to proline at position 2, Tulipano and co-workers drew a consensus in the primary structure of inhibitory peptides that included a branched-chain amino acid or an aromatic amino acid at the first position.23 Hence, QSAR models linking DPP-IV inhibitory activity and peptide descriptors suggest a greater inhibition with reduced hydrophilicity for the two N-terminal amino acids.36
In the present study, we have determined the DPP-IV inhibitory activity of four β-casomorphin-related sequences, of which our group had previously reported hormone-inducing activity.7 Two of these sequences with 90% inhibition of DPP-IV showed the aromatic amino acid tyrosine at the first position in addition to proline at the second N-terminal position. Also, other sequences determined in this study as DPP-IV inhibitory showed the previously mentioned N-terminal features, such as β-Lg 94VLDTDYK100 with the branched-chain amino acids valine and leucine, or the α-La sequence 80FLDDDLTDD88 with the aromatic amino acid phenylalanine at the N-terminal position. In short, hydrophobic and aromatic residues seem to play a crucial role in the DPP-IV inhibitory activity. Hence, the presence of hydrophobic amino acids such as leucine, phenylalanine, and methionine has been reported to facilitate various structural and interactional factors that enhance binding stability with the enzyme.36 The position of tryptophan in the peptide chain has been previously studied in the analysis of short peptide forms with anti-diabetic potential. In an overview of 55 food-derived dipeptides and tripeptides with DPP-IV or α-glucosidase inhibitory activity, analysis of the amino acid composition revealed that tryptophan was the most frequently occurring amino acid residue, followed by arginine. 72% of these oligopeptides had tryptophan at the N-terminus, while 25% displayed the same amino acid at the C-terminus.37
Another issue that would merit attention is the concurrence of CCK or GLP-1 hormone-inducing activity and DPP-IV inhibitory activity. In the family of β-casein peptides sharing the 10GEIVE14 motif, with an important CCK and GLP-1 secretory capacity, only fragment 8VPGEIVE14 exerted a relevant inhibitory activity, in accordance with the structural feature of proline at the second N-terminal position. Moreover, this sequence fulfilled another of the previously highlighted criteria, namely the presence of a branched amino acid at the first N-terminal position.23 There were great differences in the inhibitory activity of the DPP-IV enzyme for overlapping sequences, such as the β-casein peptide 172LPVPQ176 showing 93% inhibition compared to 4% in 173PVPQ176. The presence of the branched amino acid at the N-terminal position together with the presence of proline at the second N-terminal position is clearly related to the higher activity of 172LPVPQ176. A moderate inducing activity on both CCK or GLP-1 was shown for these sequences, although induction was higher for the short one, a peptide with null DPP-IV inhibitory activity. A similar case is the higher CCK and GLP-1 activity of α-La 95ILDKV99 than that of the overlapping 96LDKVG100 where greater DPP-IV inhibition is observed in the less active sequence.
There are several mechanisms by which peptides derived from dietary proteins can act as glucoregulatory compounds in the organism. Peptides can be involved in carbohydrate digestion, insulin secretion and function, glucose uptake, adipose tissue modification, and enteroendocrine hormone release.38 Among the proposed mechanisms, the modulation of the endogenous intestinal hormone secretion is a highly promising strategy in obesity and type-2 diabetes39 while the resistance to DPP-IV is known to prolong the metabolic stability of incretins in vivo.18 In this study, milk peptides resistant to gastrointestinal digestion in humans have been assayed. The hydrophobic nature of the N-terminal end of the peptides has shown a crucial role in both the secretion of GLP-1 and the DPP-IV inhibitory activity, with the aromatic and branched chain residues at the N-terminal position being highly determinant, and proline at the second position key for the last activity. However, the hormone inducing and DPP-IV-inhibiting activities were not necessarily connected. While in the assayed sequences, peptide length, hydrophobicity or charge could not be unequivocally linked to the activity, the occurrence of tryptophan or multiple glutamic acid residues in the peptide chain has been shown to be a key trait. In particular, the α-La derived sequence 94KILDKVGINYWL105 displays an outstanding GLP-1 inducing activity together with a notable DPP-IV inhibitory activity, and the β-Lg peptide 125TPEVDDEALEKFDK138 strongly induces CCK and fully inhibits DPP-IV. This points to both sequences as promising candidates for in vivo studies. The findings on hormone-inducing combined with the DPP-IV inhibitory activity are expected to shed light on the molecular characteristics associated with the metabolic signalling peptides of dietary origin.
Author contributions
Santiaga María Vivanco-Maroto: investigation, formal analysis, and writing – original draft. Cristina Gómez Marín: investigation, formal analysis, and writing – original draft. Beatriz Miralles: conceptualization, funding acquisition, supervision, and writing – review & editing. Isidra Recio: conceptualization, funding acquisition, supervision, and writing – review & editing.Data availability
All data used for this study appear in the illustrated figures, and the raw data will promptly be made available upon request.Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.Acknowledgements
This work has received financial support from project PID2023-146542OB-I00, funded by MCIN/AEI/10.13039/501100011033 from the Spanish Ministry of Science and Innovation (MICINN). SMVM and CGM are the recipients of the contracts PIE 202470E195 and PIE 202270E126 from CSIC.References
- Z. Dehnavi, H. Barghchi, A. J. Esfehani, M. Barati, Z. Khorasanchi, F. Farsi, A. N. Ostad, G. Ranjbar, R. Rezvani, M. R. Gorgani and M. Safarian, Animal and Plant-Based Proteins Have Different Postprandial Effects on Energy Expenditure, Glycemia, Insulinemia, and Lipemia: A Review of Controlled Clinical Trials, Food Sci. Nutr., 2023, 11(8), 4398–4408, DOI:10.1002/fsn3.3417.
- F. M. Gribble and F. Reimann, Function and Mechanisms of Enteroendocrine Cells and Gut Hormones in Metabolism, Nat. Rev. Endocrinol., 2019, 15(4), 226–237, DOI:10.1038/s41574-019-0168-8.
- E. L. Symonds, M. Peiris, A. J. Page, B. Chia, H. Dogra, A. Masding, V. Galanakis, M. Atiba, D. Bulmer, R. L. Young and L. A. Blackshaw, Mechanisms of Activation of Mouse and Human Enteroendocrine Cells by Nutrients, Gut, 2015, 64(4), 618–626, DOI:10.1136/gutjnl-2014-306834.
- M. Cordier-Bussat, C. Bernard, S. Haouche, C. Roche, J. Abello, J. A. Chayvialle and J. C. Cuber, Peptones Stimulate Cholecystokinin Secretion and Gene Transcription in the Intestinal Cell Line STC-I, Endocrinology, 1997, 138(3), 1137–1144, DOI:10.1210/endo.138.3.5023.
- B. Cudennec, M. Fouchereau-Peron, F. Ferry, E. Duclos and R. Ravallec, In Vitro and in Vivo Evidence for a Satiating Effect of Fish Protein Hydrolysate Obtained from Blue Whiting (Micromesistius Poutassou) Muscle, J. Funct. Foods, 2012, 4(1), 271–277, DOI:10.1016/j.jff.2011.12.003.
- G. Tulipano, L. Faggi, A. Cacciamali and A. M. Caroli, Whey Protein-Derived Peptide Sensing by Enteroendocrine Cells Compared with Osteoblast-like Cells: Role of Peptide Length and Peptide Composition, Focussing on Products of β-Lactoglobulin Hydrolysis, Int. Dairy J., 2017, 72, 55–62, DOI:10.1016/j.idairyj.2017.04.004.
- S. M. Vivanco-Maroto, V. Gallo, B. Miralles and I. Recio, CCK and GLP-1 Response on Enteroendocrine Cells of Semi-Dynamic Digests of Hydrolyzed and Intact Casein, Food Res. Int., 2023, 171, 113047, DOI:10.1016/j.foodres.2023.113047.
- Y. Komatsu, Y. Wada, H. Izumi, T. Shimizu, Y. Takeda, T. Hira and H. Hara, Casein Materials Show Different Digestion Patterns Using an in Vitro Gastrointestinal Model and Different Release of Glucagon-like Peptide-1 by Enteroendocrine GLUTag Cells, Food Chem., 2019, 277, 423–431, DOI:10.1016/j.foodchem.2018.10.123.
- J. Caron, B. Cudennec, D. Domenger, Y. Belguesmia, C. Flahaut, M. Kouach, J. Lesage, J. F. Goossens, P. Dhulster and R. Ravallec, Simulated GI Digestion of Dietary Protein: Release of New Bioactive Peptides Involved in Gut Hormone Secretion, Food Res. Int., 2016, 89, 382–390, DOI:10.1016/j.foodres.2016.08.033.
- M. Santos-Hernández, L. Amigo and I. Recio, Induction of CCK and GLP-1 Release in Enteroendocrine Cells by Egg White Peptides Generated during Gastrointestinal Digestion, Food Chem., 2020, 329, 127188, DOI:10.1016/j.foodchem.2020.127188.
- S. Theysgeur, B. Cudennec, B. Deracinois, C. Perrin, I. Guiller, A. Lepoudère, C. Flahaut and R. Ravallec, New Bioactive Peptides Identified from a Tilapia Byproduct Hydrolysate Exerting Effects on DPP-IV Activity and Intestinal Hormones Regulation after Canine Gastrointestinal Simulated Digestion, Molecules, 2021, 26(1), 136, DOI:10.3390/molecules26010136.
- T. Nishi, H. Hara, K. Asano and F. Tomita, The Soybean β-Conglycinin β 51–63 Fragment Suppresses Appetite by Stimulating Cholecystokinin Release in Rats, J. Nutr., 2003, 133(8), 2537–2542, DOI:10.1093/jn/133.8.2537.
- K. Huang, Q. Wang, H. Song, H. Cao and X. Guan, Gastrointestinal Digestion of Highland Barley Protein: Identification and Characterization of Novel Bioactive Peptides Involved in Gut Cholecystokinin Secretion, J. Sci. Food Agric., 2023, 103(15), 7869–7876, DOI:10.1002/jsfa.12870.
- H. Song, Q. Wang, Z. Shao, X. Wang, H. Cao, K. Huang, Q. Sun, Z. Sun and X. Guan, In Vitro Gastrointestinal Digestion of Buckwheat (Fagopyrum Esculentum Moench) Protein: Release and Structural Characteristics of Novel Bioactive Peptides Stimulating Gut Cholecystokinin Secretion, Food Funct., 2023, 14(16), 7469–7477, 10.1039/D3FO01951A.
- E. Diakogiannaki, R. Pais, G. Tolhurst, H. E. Parker, J. Horscroft, B. Rauscher, T. Zietek, H. Daniel, F. M. Gribble and F. Reimann, Oligopeptides Stimulate Glucagon-like Peptide-1 Secretion in Mice through Proton-Coupled Uptake and the Calcium-Sensing Receptor, Diabetologia, 2013, 56(12), 2688–2696, DOI:10.1007/s00125-013-3037-3.
- J. Yang, W. Bai, X. Zeng and C. Cui, γ-[Glu](N=1,2)-Phe/-Met/-Val Stimulates Gastrointestinal Hormone (CCK and GLP-1) Secretion by Activating the Calcium-Sensing Receptor, Food Funct., 2019, 10(7), 4071–4080, 10.1039/C9FO00313D.
- C. Wang, R. Xu, C. C. Udenigwe, L. Lin, L. Zheng and M. Zhao, Exploration of the Fasting Hypoglycemic Mechanism of Casein Hydrolysate Enriched with Glu/Gln and Glu/Gln-Containing Peptides in Db/Db Diabetic-like Mice Using Multiomics Analysis, J. Agric. Food Chem., 2025, 73(3), 1902–1916, DOI:10.1021/acs.jafc.4c07689.
- L. Hansen, C. F. Deacon, C. Ørskov and J. J. Holst, Glucagon-Like Peptide-1-(7–36)Amide Is Transformed to Glucagon-Like Peptide-1-(9–36)Amide by Dipeptidyl Peptidase IV in the Capillaries Supplying the L Cells of the Porcine Intestine1, Endocrinology, 1999, 140(11), 5356–5363, DOI:10.1210/endo.140.11.7143.
- H. Uenishi, T. Kabuki, Y. Seto, A. Serizawa and H. Nakajima, Isolation and Identification of Casein-Derived Dipeptidyl-Peptidase 4 (DPP-4)-Inhibitory Peptide LPQNIPPL from Gouda-Type Cheese and Its Effect on Plasma Glucose in Rats, Int. Dairy J., 2012, 22(1), 24–30, DOI:10.1016/j.idairyj.2011.08.002.
- C. H. Hsieh, T. Y. Wang, C. C. Hung, M. C. Chen and K. C. Hsu, Improvement of Glycemic Control in Streptozotocin-Induced Diabetic Rats by Atlantic Salmon Skin Gelatin Hydrolysate as the Dipeptidyl-Peptidase IV Inhibitor, Food Funct., 2015, 6(6), 1887–1892, 10.1039/C5FO00124B.
- Y. Fujiwara, E. Seki, J. Susuki, T. Yamane and I. Ohkubo, Effect of Katsuodashi on Postprandial Blood Glucose Levels and Identification of Dipeptidyl Peptidase IV Inhibitory Peptides among Its Components, Clin Nutr Open Sci., 2021, 39, 1–10, DOI:10.1016/j.nutos.2021.08.002.
- M. Zhang, L. Zhu, G. Wu, T. Liu, X. Qi and H. Zhang, Food-Derived Dipeptidyl Peptidase IV Inhibitory Peptides: Production, Identification, Structure-Activity Relationship, and Their Potential Role in Glycemic Regulation, Crit. Rev. Food Sci. Nutr., 2024, 64(8), 2053–2075, DOI:10.1080/10408398.2022.2120454.
- G. Tulipano, L. Faggi, A. Nardone, D. Cocchi and A. M. Caroli, Characterisation of the Potential of β-Lactoglobulin and α-Lactalbumin as Sources of Bioactive Peptides Affecting Incretin Function: In Silico and in Vitro Comparative Studies, Int. Dairy J., 2015, 48, 66–72, DOI:10.1016/j.idairyj.2015.01.008.
- E. Seki, A. Yamamoto, Y. Fujiwara, T. Yamane, H. Satsu and I. Ohkubo, Dipeptidyl Peptidase-IV Inhibitory Activity of Katsuobushi-Derived Peptides in Caco-2 Cell Assay and Oral Glucose Tolerance Test in ICR Mice, J. Agric. Food Chem., 2020, 68(23), 6355–6367, DOI:10.1021/acs.jafc.0c01942.
- C. Elisha, P. Bhagwat and S. Pillai, In Silico and in Vitro Analysis of Dipeptidyl Peptidase-IV and Angiotensin-Converting Enzyme Inhibitory Peptides Derived from Milk Lactoferrin, Int. Dairy J., 2025, 160, 106092, DOI:10.1016/j.idairyj.2024.106092.
- R. J. FitzGerald, M. Cermeño, M. Khalesi, T. Kleekayai and M. Amigo-Benavent, Application of in Silico Approaches for the Generation of Milk Protein-Derived Bioactive Peptides, J. Funct. Foods, 2020, 64, 103636, DOI:10.1016/j.jff.2019.103636.
- B. E. Goodman, Insights into Digestion and Absorption of Major Nutrients in Humans, Am. J. Physiol.: Adv. Physiol. Educ., 2010, 34(2), 44–53, DOI:10.1152/advan.00094.2009.
- J. Sanchón, S. Fernández-Tomé, B. Miralles, B. Hernández-Ledesma, D. Tomé, C. Gaudichon and I. Recio, Protein Degradation and Peptide Release from Milk Proteins in Human Jejunum. Comparison with in Vitro Gastrointestinal Simulation, Food Chem., 2018, 239, 486–494, DOI:10.1016/j.foodchem.2017.06.134.
- M. Santos-Hernández, D. Tomé, C. Gaudichon and I. Recio, Stimulation of CCK and GLP-1 Secretion and Expression in STC-1 Cells by Human Jejunal Contents and in Vitro Gastrointestinal Digests from Casein and Whey Proteins, Food Funct., 2018, 9(9), 4702–4713, 10.1039/c8fo01059e.
- M. Santos-Hernández, S. M. Vivanco-Maroto, B. Miralles and I. Recio, Food Peptides as Inducers of CCK and GLP-1 Secretion and GPCRs Involved in Enteroendocrine Cell Signalling, Food Chem., 2023, 402, 134225, DOI:10.1016/j.foodchem.2022.134225.
- M. Santos-Hernández, M. Cermeño, I. Recio and R. J. FitzGerald, In Vitro Dipeptidyl Peptidase IV Inhibitory Activity and in Situ Insulinotropic Activity of Milk and Egg White Protein Digests, Food Funct., 2021, 12(24), 12372–12380, 10.1039/D1FO00641J.
- T. Hira, S. Nakajima, Y. Eto and H. Hara, Calcium-Sensing Receptor Mediates Phenylalanine-Induced Cholecystokinin Secretion in Enteroendocrine STC-1 Cells, FEBS J., 2008, 275(18), 4620–4626, DOI:10.1111/j.1742-4658.2008.06604.x.
- Y. Wang, V. Prpic, G. M. Green, J. R. Reeve and R. A. Liddle, Luminal CCK-Releasing Factor Stimulates CCK Release from Human Intestinal Endocrine and STC-1 Cells, Am. J. Physiol.: Gastrointest. Liver Physiol., 2002, 282(1), G16–22, DOI:10.1152/ajpgi.2002.282.1.g16.
- S. Guha and K. Majumder, Comprehensive Review of γ-Glutamyl Peptides (γ-GPs) and Their Effect on Inflammation Concerning Cardiovascular Health, J. Agric. Food Chem., 2022, 70(26), 7851–7870, DOI:10.1021/acs.jafc.2c01712.
- O. Power-Grant, C. Bruen, L. Brennan, L. Giblin, P. Jakeman and R. J. FitzGerald, In Vitro Bioactive Properties of Intact and Enzymatically Hydrolysed Whey Protein: Targeting the Enteroinsular Axis, Food Funct., 2015, 6(3), 972–980, 10.1039/c4fo00983e.
- A. B. Nongonierma and R. J. FitzGerald, Structure Activity Relationship Modelling of Milk Protein-Derived Peptides with Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Activity, Peptides, 2016, 79, 1–7, DOI:10.1016/j.peptides.2016.03.005.
- J. Wu, C. Cui and D. Sun-Waterhouse, Food-Derived Trp-Rich Oligopeptides: Production, Bioactivities, and Structure-Function Relationship, Trends Food Sci. Technol., 2024, 143, 104247, DOI:10.1016/j.tifs.2023.104247.
- F. Jahandideh, S. L. Bourque and J. Wu, A Comprehensive Review on the Glucoregulatory Properties of Food-Derived Bioactive Peptides, Food Chem.:X, 2022, 13, 100222, DOI:10.1016/j.fochx.2022.100222.
- M. Santos-Hernández, F. Reimann and F. M. Gribble, Cellular Mechanisms of Incretin Hormone Secretion, J. Mol. Endocrinol., 2024, 72(4), e230112, DOI:10.1530/JME-23-0112.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5fo01394a |
| ‡ These authors contributed equally to this paper. |
| This journal is © The Royal Society of Chemistry 2025 |