
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Impact of the probiotic Bacillus coagulans on loperamide-induced delayed bowel movement in Sprague–Dawley rats†
Joo Hyun
Jang‡
a,
Yeok Boo
Chang‡
 a,
Sang Min
Kim
ab,
Kisoo
Han
c,
Wan-sup
Sim
c,
Ki-Bae
Hong
d,
Hyung Joo
Suh
ab and
Sung Hee
Han
*e
a,
Sang Min
Kim
ab,
Kisoo
Han
c,
Wan-sup
Sim
c,
Ki-Bae
Hong
d,
Hyung Joo
Suh
ab and
Sung Hee
Han
*e
aDepartment of Integrated Biomedical and Life Science, Graduate School, Korea University, Seoul 02841, Republic of Korea
bTransdisciplinary Major in Learning Health Systems, Department of Healthcare Sciences, Graduate School, Korea University, Seoul 02841, Republic of Korea
cNeo Cremar Co., Ltd., Seoul 06142, Republic of Korea
dDepartment of Food Science and Nutrition, Jeju National University, Jeju 63243, Republic of Korea
eInstitute of Human Behavior & Genetics, Korea University, Seoul 02841, Republic of Korea. E-mail: sungheeh@korea.ac.kr; Tel: +82-2-2286-1046
First published on 27th November 2024
Abstract
This study investigated the effects of Bacillus coagulans on alleviating loperamide-induced constipation. To evaluate the efficacy of B. coagulans in Sprague–Dawley (SD) rats, fecal parameters, the intestinal transit rate, and changes in intestinal mucosal cells were measured through histological analysis. Additionally, serotonin levels, water absorption, tight junction-related gene expression, and the cecal short-chain fatty acid (SCFA) content were analyzed. The administration of B. coagulans significantly altered the fecal weight and moisture content and improved gastrointestinal transit in rats with loperamide-induced constipation. Furthermore, B. coagulans supplementation restored the thickness of both muscular and mucosal layers that had been reduced by loperamide and significantly increased the area of intestinal cells, including Cajal and crypt cells. B. coagulans administration upregulated the expression levels of tryptophan hydroxylase and aquaporin genes, which were downregulated by loperamide. As the dose of B. coagulans increased, there was a corresponding upregulation in the expression of tight junction-related genes, including occludin (OCLN), zonula occludens 1 (ZO-1), and claudin 1 (CLDN1). Additionally, the levels of c-kit, AQP 3, and OCLN proteins, which were elevated by loperamide treatment, were reduced with higher concentrations of B. coagulans. Loperamide decreased the acetic acid content; however, high doses of B. coagulans increased it, leading to a significant increase in the total cecal SCFA content. Thus, B. coagulans shows potential as a probiotic for improving constipation.
1. Introduction
Constipation is a prevalent gastrointestinal condition characterized by infrequent or difficult bowel movements, often accompanied by symptoms such as bloating, abdominal pain, and a sensation of incomplete evacuation. It can arise from a variety of factors including a low fiber diet and inadequate fluid intake, both of which affect stool consistency. Other contributing factors include physical inactivity, certain medications, and various health conditions. The management of chronic constipation typically requires dietary adjustments, regular exercise, and the use of fiber supplements, osmotic agents, stool softeners, and over-the-counter laxatives.1,2 Notably, prescription medications can cause various side effects including diarrhea, bloating, gas, nausea, stomach pain, and dependence. Therefore, to improve the quality of life, the effective management of constipation requires sustained lifestyle changes.3Prebiotics, such as lactulose, inulin, and various oligosaccharides, alleviate constipation by promoting the growth and activity of beneficial gut bacteria.4 Probiotics, particularly those from the Lactobacillus or Bifidobacterium genera, are known to increase the frequency of evacuation and stool quality by improving the gastrointestinal transit time.5 Prebiotics, probiotics, and synbiotics alleviate constipation symptoms by increasing the population and diversity of beneficial gut bacteria, thereby promoting colonic muscle contraction and regulating intestinal motility through the production of short-chain fatty acids (SCFAs).6 SCFAs trigger the release of peptide YY, glucagon-like peptide-1, tryptophan 5-hydroxylase, and mucin from enteroendocrine and epithelial cells, thereby affecting intestinal motility and strengthening the epithelial barrier.7–9
Numerous studies have investigated the environmental tolerance and probiotic properties of spore-producing Bacillus coagulans strains. B. coagulans produce enzymes that enhance intestinal excretion and digestion, suppress pathogenic bacterial growth, and interact with the host immune system by modulating metabolites and immune cell function.10,11 Clinical trials have demonstrated that supplementation with B. coagulans improves patients’ quality of life by alleviating abdominal pain, reducing bloating, and modifying stool frequency.12,13 Additionally, B. coagulans has demonstrated potential in managing comorbid major depressive disorder exacerbated by persistent irritable bowel syndrome (IBS) symptoms, as supported by previous clinical study findings.14
Our study aimed to evaluate the effectiveness of B. coagulans in alleviating constipation and to elucidate its specific mechanisms of action using an animal model. To assess the effect of administering B. coagulans on alleviating constipation, the intestinal transit rate, fecal parameters, and intestinal peristalsis-related factors were analyzed in loperamide constipation-induced Sprague–Dawley (SD) rats.
The research investigates Bacillus coagulans, a probiotic known for its ability to improve gut health, focusing on its specific effects in a loperamide-induced constipation model. This highlights its potential as a novel treatment for constipation, distinct from traditional laxatives or other interventions. Constipation is a widespread issue, particularly in chronic cases. The study adds value by exploring a non-pharmacological intervention that could be safer and more sustainable than current treatments, such as laxatives, which often come with side effects or dependency issues.
This research introduces B. coagulans as a promising therapeutic for constipation, enhancing the literature with mechanistic details and suggesting future avenues for human trials.
2. Materials and methods
2.1. Materials
Loperamide was supplied by Sigma-Aldrich (St. Louis, MO, USA). Lactulose, an osmotic laxative, was sourced from JW Pharmaceutical Co., Ltd (Seoul, Korea). B. coagulans GBI-30, 6086 (BC30) was procured from Neo Cremar Co., Ltd (Seoul, Korea).The manufacturing process for Bacillus coagulans GBI-30, 6086 includes fermentation, followed by a recovery phase, and then either spray-drying or freeze-drying. The recovery phase, which involves centrifugation, is designed to collect and concentrate Bacillus coagulans after the fermentation process.
2.2. Animals and experimental design
Six-week-old male SD rats (160–180 g) were supplied by Orient Bio (Seongnam, Republic of Korea). Following a 1-week adaptation period in a controlled environment (23 ± 1 °C, 55 ± 2% relative humidity, and 12-hour light/dark cycle), constipation was induced by administering 5 mg kg−1 of loperamide orally twice a day in all groups except for the normal control (NC) group. The rats were divided into five groups (eight rats per group): NC, control (CON, loperamide only, 5 mg kg−1), positive control (PC, lactulose 2010 mg kg−1 with loperamide), low-dose B. coagulans (BCL, 1 × 107 colony-forming units [CFU] with loperamide), and high-dose B. coagulans (BCH, 2 × 109 CFU with loperamide). All treatments were administered daily at 10 AM for 4 weeks. Sample sizes were determined based on a power analysis to detect statistically significant differences, using 8 animals per group across 5 groups (n = 40 in total). The intestines were harvested immediately after euthanasia.This study was conducted in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health and was approved by the Korea University Institutional Animal Care and Use Committee (KUIACUC-2023-0028). All surgeries were performed under anesthesia, and every effort was made to minimize the suffering of the animals used in this study.
2.3. Analysis of fecal parameters and intestinal transit rate
The fecal parameters and intestinal transit rates were evaluated using the method previously described by Inatomi et al.15 Fecal samples were collected weekly at 11 AM, starting two weeks after the administration of loperamide, lactulose, and/or B. coagulans as required. The samples were dried at 75 °C for 18 hours, and their water content was determined by measuring the weight difference.Before the end of the experiment, the rats were fasted for 12 hours and orally administered a 5% charcoal suspension. Subsequently, the rats were sacrificed within 30 min, and the total intestinal length, including the distance traversed by charcoal, was measured to determine the intestinal transit rate.
2.4. Histological analysis
Hematoxylin–eosin (H&E) staining and immunohistochemistry (IHC) were performed as described in a study by Jang and Kim et al.16 The H&E staining was conducted on sections of the colon from the cecum to the rectum, while the IHC staining was performed using the colon od SD rat. The stained intestinal mucosal cells and interstitial cells of Cajal (ICC) were examined under a light microscope (ZEISS Axiovert S100, Jena, Germany). Crypt and intestinal mucosa cells were examined using an optical microscope with Leica Application Suite software (Leica Microsystem, Heerbrugg, Switzerland).2.5. Quantitative real-time reverse-transcription polymerase chain reaction
The total ribonucleic acid (RNA) was extracted from rat intestines using TRIzol® reagent (Invitrogen, Carlsbad, CA, USA). Genomic deoxyribonucleic acid (DNA) was removed from the RNA samples by treatment them with Q1 RNase-free DNase (Promega, Madison, WI, USA). Subsequently, complementary DNA (cDNA) was synthesized using SuperScript® III (Invitrogen) reverse transcriptase and the 1μg of the resulting cDNA was analyzed through quantitative real-time reverse-transcription polymerase chain reaction (qRT-PCR) using a SYBR Green PCR Master Mix kit (Applied Biosystems, Foster City, CA, USA). The expression levels of each target gene were normalized to the NC group using a previously described ΔΔCt method.172.6. Western blot analysis
Target protein levels were analyzed using the western blotting method described by Jang and Kim et al.16 Intestinal tissue (100 mg) was mixed with lysis buffer to create a homogenate, which was then centrifuged to separate the supernatant (10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g, 10 min, 4 °C). Protein concentrations in the supernatant were determined using a bicinchoninic acid assay kit (Thermo Fisher Scientific, Waltham, MA, USA). The primary antibody (β-actin, C-kit, AQP3, and OCLN, 1:1000) and the secondary antibodies (anti-rabbit IgG, 1:1000) were obtained from Cell Signaling Technology (Beverly, MA, USA). Protein bands were detected using a FluorChem M Fluorescent Western Imaging System (ProteinSimple, Santa Clara, California, USA).
000g, 10 min, 4 °C). Protein concentrations in the supernatant were determined using a bicinchoninic acid assay kit (Thermo Fisher Scientific, Waltham, MA, USA). The primary antibody (β-actin, C-kit, AQP3, and OCLN, 1:1000) and the secondary antibodies (anti-rabbit IgG, 1:1000) were obtained from Cell Signaling Technology (Beverly, MA, USA). Protein bands were detected using a FluorChem M Fluorescent Western Imaging System (ProteinSimple, Santa Clara, California, USA).
2.7. Short-chain fatty acid analysis
SCFAs in the cecum were analyzed following the method described by Kim et al.,18 selected for its proven accuracy and efficacy in the quantification of SCFAs in biological samples. Cecal content (200 mg) was extracted using 2 mL of methanol and was subsequently filtered through a 0.45 μm Millipore filter (Millipore, USA). The resulting extracts were analyzed through gas chromatography (GC, Agilent Technologies, CA, USA) equipped with an HP-FFAP (nitroterephthalic acid modified polyethylene glycol high polarity designed) column (50 m × 0.32 mm × 0.50 μm) and a flame ionization detector. The sample injection volume was set at 1 μL, with inlet and detector temperatures maintained at 200 °C and 240 °C, respectively.2.8. Statistical analysis
The experimental results are expressed as means ± standard errors of the mean (SEM). Statistical analyses were performed using the SPSS software version 24.0 (SPSS Inc., USA). Comparisons among experimental groups were performed using Tukey's multiple-range test after an analysis of variance with the significance level set at a p value of <0.05.3. Results
3.1. Changes in fecal parameters caused by B. coagulans administration in loperamide-treated SD rats
Analysis of the changes in body weight, dietary intake, organ weight, and serum biochemical parameters owing to B. coagulans administration revealed no significant differences between the NC and CON groups (Fig. S1–S3†). However, the number, weight, and water content of fecal pellets were significantly reduced in the CON group compared with those in the NC group (Fig. 1A, B and C; p < 0.01, p < 0.05, and p < 0.001, respectively). Loperamide caused a notable reduction in the gastrointestinal transit ratio compared with the NC group (Fig. 1D, p < 0.01). The combined treatment of lactulose (PC) and a high dose of B. coagulans (BCH, 2.0 × 109 CFU per day) significantly increased the number of fecal pellets compared with the CON group (Fig. 1A, p < 0.01, and p < 0.05, respectively). Both the weight and water content of the fecal pellets were significantly higher in the BCL and BCH groups compared with the CON group (Fig. 1B and C, p < 0.001, respectively). Mice in the PC group did not experience significant improvements in the gastrointestinal transit ratio compared with those in the CON group, while mice in the BCL and BCH groups demonstrated significant enhancements (Fig. 1D, p < 0.001). | ||
| Fig. 1 Effect of Bacillus coagulans on the animal fecal and gastrointestinal transit ratio. (A) Number of fecal pellets, (B) weight of fecal pellets, (C) fecal water contents, and (D) gastrointestinal transit ratio in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (n = 6). The following symbols indicated significance: #p < 0.05, ##p < 0.01, ###p < 0.001 vs. NC, and p < 0.05, p < 0.01, p < 0.001 vs. CON according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose treated, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). | ||
3.2. Changes in intestinal architecture caused by B. coagulans administration in loperamide-treated SD rats
The thickness of the intestinal muscular and mucosal layers significantly decreased in the CON group compared to the NC group (Fig. 2A–D; p < 0.001, p < 0.05). In contrast, the PC group, treated with lactulose, showed a marked increase in both layers relative to the CON group (p < 0.001, p < 0.05). Low and high doses of B. coagulans (BCL, BCH) significantly increased the muscular layer thickness (p < 0.01, p < 0.001, respectively), while the mucosal layer thickness increased significantly in the BCH group (p < 0.05). Loperamide-induced constipation also reduced crypt cell and ICC area, but both were restored in the PC and BCH groups (p < 0.05, p < 0.001) area (Fig. 3D, p < 0.05 and p < 0.001, respectively). | ||
| Fig. 2 Effect of Bacillus coagulans on intestinal muscular (A and B) and mucosal (C and D) layer thickness in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (n = 6). The following symbols indicated significance: #p < 0.05, ###p < 0.001 vs. NC and p < 0.05, p < 0.01, p < 0.001 vs. CON according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). | ||
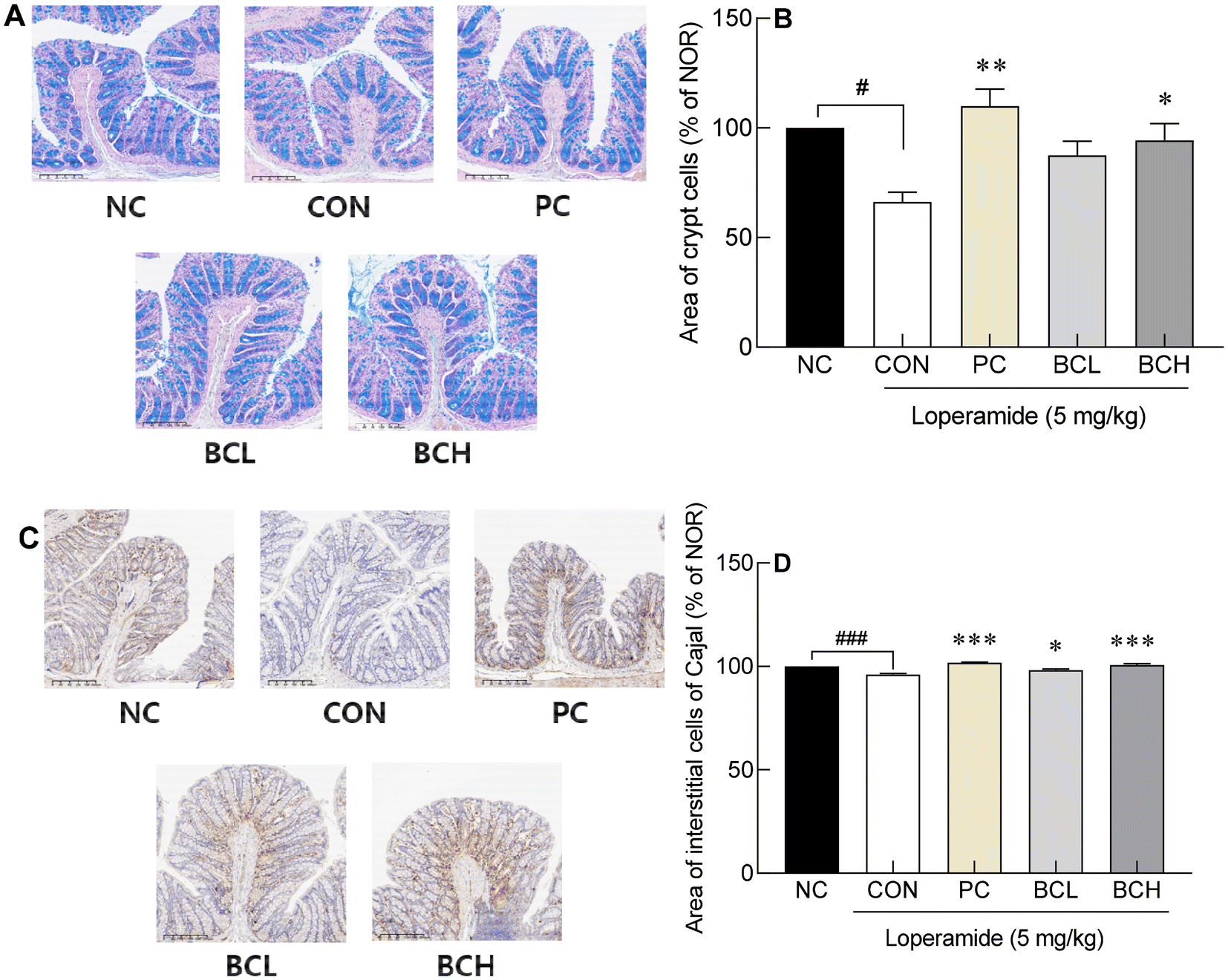 | ||
| Fig. 3 Effect of Bacillus coagulans on the area of crypt cells (A and B) and intestinal cells of Cajal (C and D) in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (n = 6). The following symbols indicated significance: #p < 0.05, ###p < 0.001 vs. NC and p < 0.05, p < 0.01, p < 0.001 vs. CON according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). | ||
3.3. Changes in the expression of serotonin and mucin-related genes caused by B. coagulans administration in loperamide-treated SD rats
Loperamide treatment (CON) significantly reduced serotonin transporter (SERT) expression, crucial for serotonin reuptake, while neither lactulose nor B. coagulans affected its levels (Fig. 4A). Additionally, loperamide suppressed tryptophan hydroxylase (TPH) expression, vital for neurotransmitter synthesis. In contrast, lactulose and B. coagulans increased TPH1 and TPH2 expression, with TPH1 showing a dose-dependent increase (Fig. 4B and C). | ||
| Fig. 4 Effects of Bacillus coagulans on the mRNA expression of (A) SERT, (B) TPH 1, (C) TPH 2, (D) MUC2, and (E) MUC4 in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (n = 6). The following symbols indicated significance: #p < 0.05, ##p < 0.01, ###p < 0.001 vs. NC and p < 0.05, p < 0.01, p < 0.001 vs. CON according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). SERT: serotonin transporter, TPH1: tryptophan hydroxylase. | ||
Loperamide also reduced mucin (MUC) 2 and MUC4 expression, essential for the intestinal mucus barrier (p < 0.001, Fig. 4D and E). Lactulose treatment significantly increased MUC4 expression (p < 0.05, p < 0.001), and B. coagulans elevated both MUC2 and MUC4 levels (Fig. 4E). Notably, MUC2 expression was higher in B. coagulans-treated mice, while MUC4 expression was more significantly elevated by lactulose. Overall, B. coagulans reversed loperamide-induced decreases in TPH and MUC expression.
3.4. Changes in the expression of intestinal mucosal barrier- and tight junction-related genes in loperamide-treated SD rats caused by B. coagulans administration
Absorption and reduced intestinal secretion, resulting in dry stools. Loperamide (CON) upregulated these genes, while lactulose (PC) and B. coagulans (BCL, BCH) significantly downregulated their expression (Fig. 5A–C).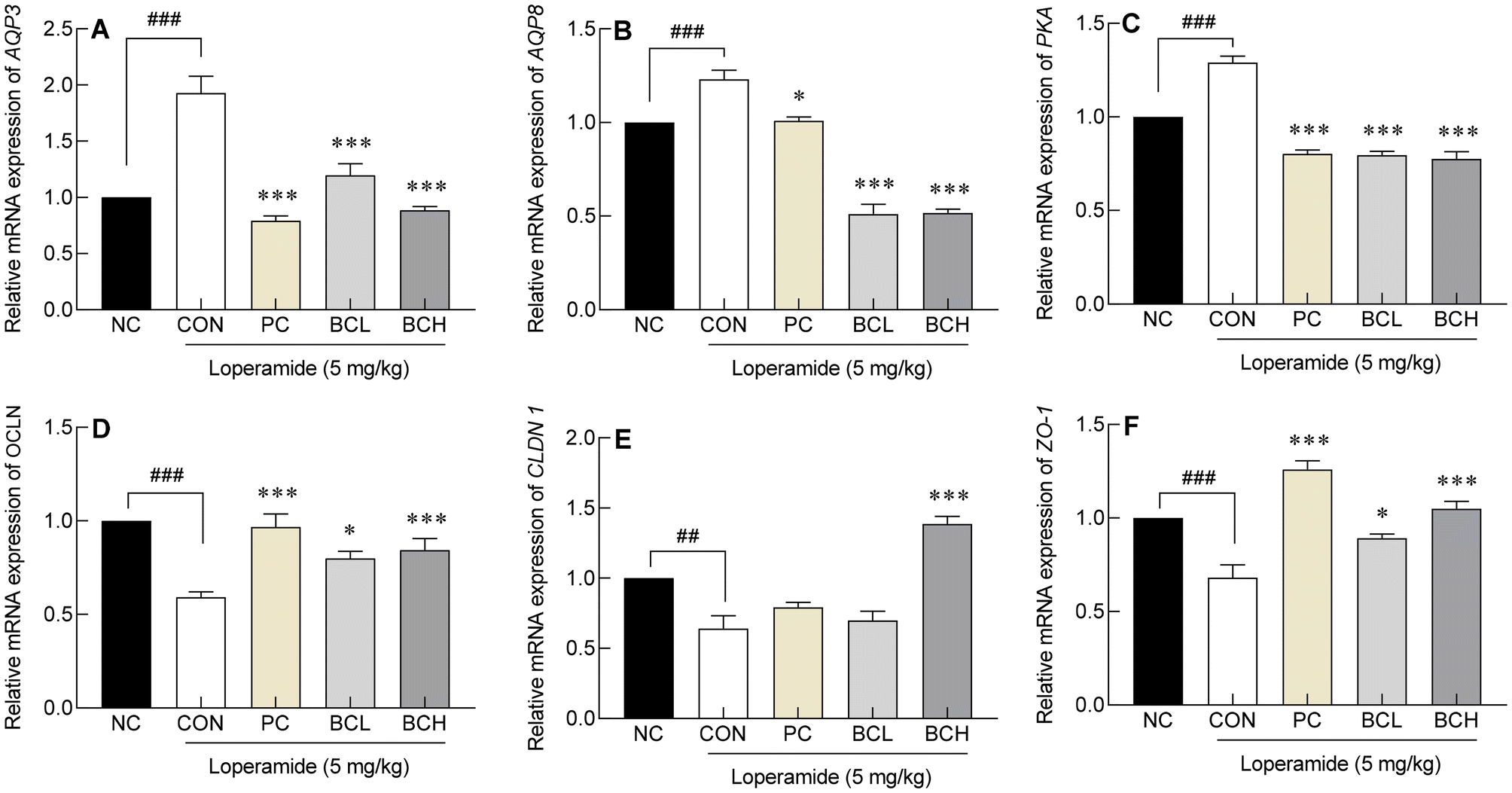 | ||
| Fig. 5 Effects of Bacillus coagulans on the mRNA expression of (A) AQP3, (B) AQP8, (C) PKA, (D) OCLN, (E) CLDN1, and (F) ZO-1 in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (n = 6). The following symbols indicated significance: ###p < 0.001 vs. NC and p < 0.05, p < 0.001 vs. CON according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). MUC: mucin. | ||
Loperamide also decreased the expression of OCLN, CLDN1, and ZO-1, key genes involved in maintaining the intestinal barrier (p < 0.001, p < 0.01, p < 0.001, respectively). B. coagulans restored OCLN and ZO-1 levels in a dose-dependent manner (Fig. 5D and F). While CLDN1 remained unchanged with lactulose (PC) and low-dose B. coagulans (BCL), high-dose B. coagulans (BCH) significantly increased its expression (p < 0.001, Fig. 5E). Overall, B. coagulans restored tight junction integrity and reversed the effects of loperamide on the intestinal mucosal barrier.
3.5. Changes caused by B. coagulans administration in protein levels of loperamide-treated SD rats
Loperamide treatment showed a non-significant trend towards decreasing c-Kit protein levels in the SCF/c-Kit pathway. However, c-Kit levels significantly increased with lactulose (PC) and high-dose B. coagulans (BCH) compared to the CON group (Fig. 6A). Loperamide also significantly increased AQP3 protein levels (p < 0.01, Fig. 6B), consistent with gene expression results. In contrast, both lactulose and B. coagulans significantly reduced AQP3 levels (p < 0.05 and p < 0.001, respectively), with B. coagulans showing a dose-dependent effect. For OCLN protein, the reduction caused by loperamide was significantly reversed by lactulose (p < 0.001), while B. coagulans increased OCLN protein, though the change was not statistically significant (Fig. 6C). | ||
| Fig. 6 Effect of Bacillus coagulans on the protein levels of (A) C-kit, (B) AQP 3, and (C) OCLN in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (n = 6). The following symbols indicated significance: ##p < 0.01 vs. NC and p < 0.05, p < 0.001 vs. CON according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). C-kit: receptor tyrosine kinase protein, AQP 3: aquaporin 3, OCLN: occludin. | ||
3.6. Changes in SCFA content caused by B. coagulans administration in loperamide-treated SD rats
Loperamide-induced constipation significantly reduced total SCFA content, which is crucial for maintaining intestinal barrier integrity and motility (Fig. 7, p < 0.01). Specifically, acetic acid was significantly decreased by loperamide (p < 0.01), while propionic and butyric acid levels were unaffected. Treatment with lactulose (PC) and high-dose B. coagulans (BCH) significantly restored acetic acid levels (p < 0.05 and p < 0.01, respectively), leading to a significant increase in total SCFA content, reversing the effects of loperamide (Fig. 7).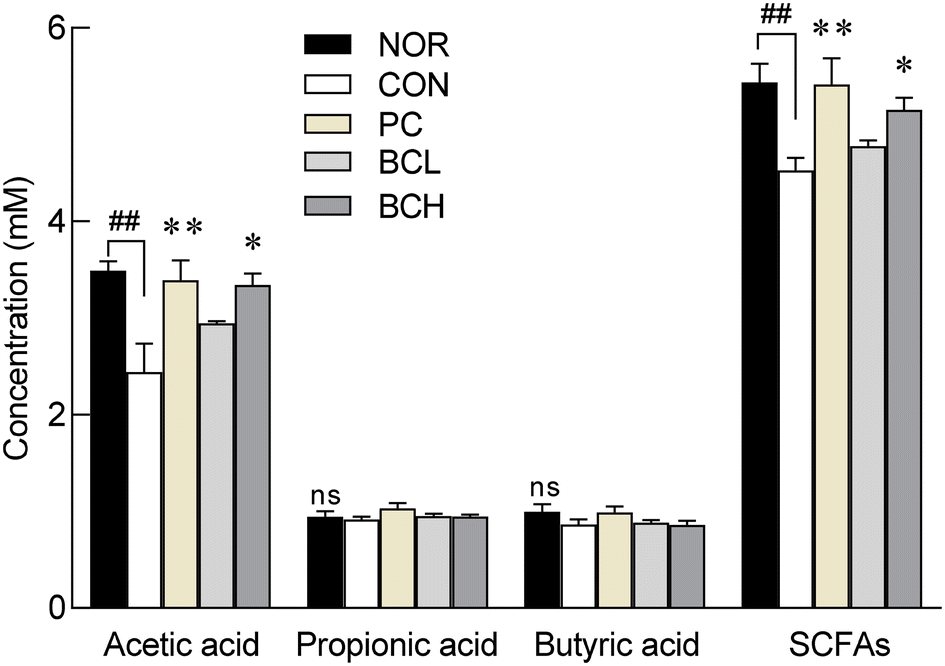 | ||
| Fig. 7 Effects of Bacillus coagulans on SCFAs in SD rats with loperamide-induced constipation. Data are expressed as the means ± standard error of the mean (S.E.M) (n = 6). The following symbols indicated significance: ##p < 0.01 < 0.001 vs. NC and p < 0.05, p < 0.01 vs. CON group according to Tukey's test. NC: normal group; CON: loperamide treated-control group (5 mg kg−1); PC: positive control group (lactulose, 2010 mg kg−1); BCL: low-dose Bacillus coagulans-treated group (1.0 × 107 CFU per day); BCH: high-dose Bacillus coagulans-treated group (2.0 × 109 CFU per day). SCFAs: short chain fatty acids; ns: not significant. | ||
4. Discussion
Constipation, affecting 5%–20% of the general population, is one of the most prevalent gastrointestinal issues. However, its symptoms vary widely among patients and often present as a cluster of related issues. This makes it challenging to objectively define it.1 Factors such as lifestyle, medications, pelvic floor muscle weakness, and changes in the colon or rectal tissues contribute to constipation. Dietary control and appropriate physical activity are essential preventive measures. Recent studies have suggested that the probiotic B. coagulans can alleviate the symptoms of constipation and IBS. However, limited research exists on the molecular and intestinal morphological effects of B. coagulans in relieving constipation.In the present study, we assessed the effects of different doses of B. coagulans on fecal parameters and gastrointestinal transit in rats with loperamide-induced constipation (Fig. 1). Constipation can arise from various causes, with long-term issues often linked to imbalances in intestinal microbiota. Previous studies have reported that probiotics like Lactobacillus acidophilus, Bifidobacterium bifidum, and Clostridium tabificum can enhance gastrointestinal transit in germ-free mice.19 The results of several clinical trials further support the efficacy of probiotics in improving bowel movements and alleviating chronic constipation with benefits often persisting even after treatment discontinuation.20,21 Therefore, in-depth research on the clinical effects of probiotics is essential.
Using H&E and IHC staining, we assessed the impact of B. coagulans on the intestinal architecture in rats with the loperamide-induced constipation (Fig. 2 and 3). Constipation and IBS often disrupt the gastrointestinal barrier owing to changes in the epithelium and mucosal layers, which express mucin glycoproteins. Loperamide-induced constipation typically results in reduced mucus production by crypt epithelial cells and a thinner mucosal surface layer.22 Probiotics like Lactobacillus and Bifidobacterium have enhanced mucus production and thickness in drug-induced constipation models.23 Additionally, these probiotics increased the mucus layer's thickness and prevented defects in models of dextran sulfate sodium (DSS)-induced colitis and obesity.24 Furthermore, while constipation reduces crypt cells and ICCs, thereby slowing intestinal fluid passage and peristalsis,25,26 Wnt/β-catenin signaling promotes stem cell renewal in the crypts, and probiotics boost crypt height, intestinal stem cells, Paneth cells, and goblet cells in the small intestine.27,28
Serotonin (5-hydroxytryptamine or 5-HT) plays a crucial role in regulating intestinal peristalsis and influences muscle contraction and relaxation. It is involved in gastrointestinal motility, secretion, and inflammation in disorders such as IBS and constipation, making it a target for treatment.29,30 Alterations in SERT transcript expression have been linked to different types of IBS, and increased TPH1 expression in enterochromaffin cells influences intestinal motility.31,32 The intake of B. coagulans elevates the expression levels of SERT, TPH1, and TPH2, which were reduced by loperamide treatment (Fig. 4).
In loperamide-treated rats, constipation is associated with a decrease in luminal mucin, which protects the colonic mucosa. However, we demonstrated that the administration of B. coagulans significantly elevated the expression of MUC2 and MUC4 (Fig. 4). Moreover, prebiotics such as sulfated polysaccharides and the probiotic formula VSL#3 enhanced the epithelial mucin production, MUC2 expression, and mucus layer thickness, thereby improving fecal excretion through enhanced intestinal motility.33,34 Additionally, single bacterial strains such as L. plantarum 299v, Escherichia coli Nissle 1917, and L. casei GG increase mucin gene expression in HT-29 and Caco-2 cell lines, with L. plantarum 299v exerting protective effects by modulating the caspase-dependent apoptotic pathway.35–38
B. coagulans administration influenced the expression of intestinal mucosal barrier genes (AQP3, AQP8, and PKA) in the qRT-PCR analysis (Fig. 5). AQPs facilitate water transport and help maintain the intracellular water balance. The abnormal expression of AQPs is linked to constipation, as it can result in excessive water absorption and reduced intestinal fluid secretion.39,40 Loperamide exacerbates constipation by activating the vasoactive intestinal peptide (VIP)/cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA)/aquaporin 3 (AQP3) signaling pathway.41 Additionally, the role of PKA in phosphorylating neuronal nitric oxide synthase and promoting ileal relaxation is associated with slow-transit constipation.42 Prebiotics and probiotics, such as Lactococcus lactis subsp. lactis HFY14 and high specific volume polysaccharides, alleviate constipation symptoms by modulating this signaling pathway.43,44
The expression levels of OCLN, CLDN1, and ZO-1, which are essential components of tight junctions, were significantly elevated in the B. coagulans-treated group compared with those in the loperamide-induced constipation model (Fig. 5). These tight junction proteins, located beneath the mucus layer that protects the intestines, form complexes with cytoplasmic adaptors, such as ZO-1 to maintain the intestinal epithelial barrier.45 In piglets, a synbiotic mix of lactulose and B. coagulans improved the LPS-induced intestinal damage by regulating tight junction protein expression in the jejunum and ileum.46 Similarly, B. coagulans SCC-19 mitigates cadmium-induced damage in common carp by increasing the mRNA levels of tight junction proteins and preserving intestinal barrier function.47
Our results revealed that B. coagulans administration increased the protein levels of C-kit, AQP3, and OCLN, as well as the SCFA content, which were collectively decreased with loperamide treatment (Fig. 6 and 7). c-kit, a receptor involved in gastrointestinal motility, alleviates constipation when its expression is enhanced with B. coagulans.4 The SCF/C-kit signaling pathway influences gastrointestinal muscle activity by regulating the ICC, which are crucial for motility and constipation relief.26 Probiotics alleviate constipation by modifying the gut microbiota, improving transit time, and increasing the SCFA content, which support colonic epithelial cell proliferation and mucosal growth.48 For instance, Bifidobacterium enhances acetate levels, correlating with reduced constipation,49 while B. coagulans X26 boosts SCFA content and improves the intestinal structure in laying hens.50
Here, we demonstrated the effectiveness of B. coagulans in alleviating constipation. Moving forward, we will further investigate the gut microbiome to analyze the role of B. coagulans on the abundance of beneficial gut microbiota and evaluate the various changes it introduces in the gut environment.
Meanwhile, there are several studies that have explored Bacillus coagulans as a potential treatment for constipation in humans. For example, one clinical study investigated the effects of B. coagulans Unique IS2 in adults with functional constipation. The results showed that participants who received the B. coagulans supplement had significant improvements in bowel movements compared to the placebo group, suggesting its potential as a therapeutic agent for managing constipation.51 Another randomized controlled trial demonstrated that B. coagulans SNZ 1969 improved intestinal motility and reduced constipation symptoms by modulating the gut microbiota. These findings point to the probiotic's ability to influence gut health and alleviate constipation symptoms, though further research, particularly involving larger human trials, is needed to establish its long-term efficacy and safety across different populations.
5. Conclusion
B. coagulans alleviates loperamide-induced constipation symptoms. These effects occur through changes in intestinal environment-related factors. In summary, this study demonstrates the efficacy of B. coagulans as a probiotic for managing constipation. B. coagulans influences the secretion of neurotransmitters and glycoproteins, as well as the expression levels of genes and proteins that are crucial for maintaining a balance in the intestinal environment. Additionally, it alleviates constipation symptoms by altering the content of SCFAs in response to the changes in the gut microbiota.Author contributions
Joo Hyun Jang: conceptualization, formal analysis, data curation, writing – original draft, funding acquisition. Yeok Boo Chang: methodology, formal analysis, investigation, writing – original draft. Sang Min Kim: methodology, formal analysis, writing – review & editing. Kisoo Han: methodology, formal analysis. Sim Wan Scu: methodology, formal analysis. Ki-Bae Hong: conceptualization, methodology. Hyung Joo Shu: conceptualization, methodology. Sung Heel Han: conceptualization, methodology, writing – review & editing, supervsion, funding acquisition.Data availability
The data are available from the corresponding author on reasonable request.Conflicts of interest
The authors declare no conflict of interest.References
- A. E. Bharucha and B. E. Lacy, Mechanisms, Evaluation, and Management of Chronic Constipation, Gastroenterology, 2020, 158, 1232–1249 CrossRef CAS.
- A. Sharma, S. S. C. Rao, K. Kearns, K. D. Orleck and S. A. Waldman, Review article: diagnosis, management and patient perspectives of the spectrum of constipation disorders, Aliment. Pharmacol. Ther., 2021, 53, 1250–1267 CrossRef.
- T. Ueki and M. Nakashima, Relationship Between Constipation and Medication, J. UOEH, 2019, 41, 145–151 CrossRef CAS PubMed.
- M. M. Araujo and P. B. Botelho, Probiotics, prebiotics, and synbiotics in chronic constipation: Outstanding aspects to be considered for the current evidence, Front. Nutr., 2022, 9, 935830 CrossRef.
- E. Dimidi, S. M. Scott and K. Whelan, Probiotics and constipation: mechanisms of action, evidence for effectiveness and utilisation by patients and healthcare professionals, Proc. Nutr. Soc., 2020, 79, 147–157 CrossRef.
- K. R. Pandey, S. R. Naik and B. V. Vakil, Probiotics, prebiotics and synbiotics- a review, J. Food Sci. Technol., 2015, 52, 7577–7587 CrossRef CAS.
- A. Psichas, M. L. Sleeth, K. G. Murphy, L. Brooks, G. A. Bewick, A. C. Hanyaloglu, M. A. Ghatei, S. R. Bloom and G. Frost, The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents, Int. J. Obes., 2015, 39, 424–429 CrossRef CAS.
- B. B. Nankova, R. Agarwal, D. F. MacFabe and E. F. La Gamma, Enteric bacterial metabolites propionic and butyric acid modulate gene expression, including CREB-dependent catecholaminergic neurotransmission, in PC12 cells–possible relevance to autism spectrum disorders, PLoS One, 2014, 9, e103740 CrossRef PubMed.
- L. E. Willemsen, M. A. Koetsier, S. J. van Deventer and E. A. van Tol, Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E(1) and E(2) production by intestinal myofibroblasts, Gut, 2003, 52, 1442–1447 CrossRef CAS PubMed.
- D. Keller, S. Verbruggen, H. Cash, S. Farmer and K. Venema, Spores of Bacillus coagulans GBI-30, 6086 show high germination, survival and enzyme activity in a dynamic, computer-controlled in vitro model of the gastrointestinal tract, Benefic. Microbes, 2019, 10, 77–87 CAS.
- T. V. Bomko, T. N. Nosalskaya, T. V. Kabluchko, Y. V. Lisnyak and A. V. Martynov, Immunotropic aspect of the Bacillus coagulans probiotic action, J. Pharm. Pharmacol., 2017, 69, 1033–1040 CrossRef CAS PubMed.
- M. Majeed, K. Nagabhushanam, S. Natarajan, A. Sivakumar, F. Ali, A. Pande, S. Majeed and S. K. Karri, Bacillus coagulans MTCC 5856 supplementation in the management of diarrhea predominant Irritable Bowel Syndrome: a double blind randomized placebo controlled pilot clinical study, Nutr. J., 2016, 15, 21 CrossRef PubMed.
- M. R. Sudha, N. Jayanthi, M. Aasin, R. D. Dhanashri and T. Anirudh, Efficacy of Bacillus coagulans Unique IS2 in treatment of irritable bowel syndrome in children: a double blind, randomised placebo controlled study, Benefic. Microbes, 2018, 9, 563–572 CAS.
- M. Majeed, K. Nagabhushanam, S. Arumugam, S. Majeed and F. Ali, Bacillus coagulans MTCC 5856 for the management of major depression with irritable bowel syndrome: a randomised, double-blind, placebo controlled, multi-centre, pilot clinical study, Food Nutr. Res., 2018, 62, 218 Search PubMed.
- T. Inatomi and M. Honma, Effects of probiotics on loperamide-induced constipation in rats, Sci. Rep., 2021, 11, 24098 CrossRef CAS.
- J. H. Jang, S. M. Kim, H. J. Suh, M. Gim, H. Shin, H. Jang, H. S. Choi, S. H. Han and Y. B. Chang, Lactitol Alleviates Loperamide-Induced Constipation in Sprague Dawley Rats by Regulating Serotonin, Short-Chain Fatty Acids, and Gut Microbiota, Foods, 2024, 13, 2128 CrossRef CAS PubMed.
- K. J. Livak and T. D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method, Methods, 2001, 25, 402–408 CrossRef CAS PubMed.
- B.-K. Kim, I. S. Choi, J. Kim, S. H. Han, H. J. Suh and J.-K. Hwang, Effects of Fermented Milk with Mixed Strains as a Probiotic on the Inhibition of Loperamide-Induced Constipation, Food Sci. Anim. Resour., 2017, 37, 906–916 Search PubMed.
- E. Husebye, P. M. Hellstrom, F. Sundler, J. Chen and T. Midtvedt, Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats, Am. J. Physiol.: Gastrointest. Liver Physiol., 2001, 280, G368–G380 CrossRef CAS PubMed.
- A. Tremblay, J. Auger, Z. Alyousif, S. E. C. Calero, O. Mathieu, D. Rivero-Mendoza, A. Elmaoui, W. J. Dahl and T. A. Tompkins, Total Transit Time and Probiotic Persistence in Healthy Adults: A Pilot Study, J. Neurogastroenterol. Motil., 2023, 29, 218–228 CrossRef PubMed.
- A. Ibarra, M. Latreille-Barbier, Y. Donazzolo, X. Pelletier and A. C. Ouwehand, Effects of 28-day Bifidobacterium animalis subsp. lactis HN019 supplementation on colonic transit time and gastrointestinal symptoms in adults with functional constipation: A double-blind, randomized, placebo-controlled, and dose-ranging trial, Gut Microbes, 2018, 9, 236–251 CrossRef CAS PubMed.
- A. Shimotoyodome, S. Meguro, T. Hase, I. Tokimitsu and T. Sakata, Decreased colonic mucus in rats with loperamide-induced constipation, Comp. Biochem. Physiol. A Mol. Integr. Physiol., 2000, 126, 203–212 CrossRef CAS PubMed.
- E. Dimidi, S. Christodoulides, S. M. Scott and K. Whelan, Mechanisms of Action of Probiotics and the Gastrointestinal Microbiota on Gut Motility and Constipation, Adv. Nutr., 2017, 8, 484–494 CrossRef CAS.
- S. Fernandez-Tome, L. O. Moreno, M. Chaparro and J. P. Gisbert, Gut Microbiota and Dietary Factors as Modulators of the Mucus Layer in Inflammatory Bowel Disease, Int. J. Mol. Sci., 2021, 22, 10224 CrossRef CAS PubMed.
- C. L. He, L. Burgart, L. Wang, J. Pemberton, T. Young-Fadok, J. Szurszewski and G. Farrugia, Decreased interstitial cell of cajal volume in patients with slow-transit constipation, Gastroenterology, 2000, 118, 14–21 CrossRef CAS PubMed.
- T. Wedel, J. Spiegler, S. Soellner, U. J. Roblick, T. H. Schiedeck, H. P. Bruch and H. J. Krammer, Enteric nerves and interstitial cells of Cajal are altered in patients with slow-transit constipation and megacolon, Gastroenterology, 2002, 123, 1459–1467 CrossRef PubMed.
- H. Clevers and R. Nusse, Wnt/beta-catenin signaling and disease, Cell, 2012, 149, 1192–1205 CrossRef CAS PubMed.
- Y. S. Lee, T. Y. Kim, Y. Kim, S. H. Lee, S. Kim, S. W. Kang, J. Y. Yang, I. J. Baek, Y. H. Sung, Y. Y. Park, S. W. Hwang, E. Oh, S. Kim, S. Liu, N. Kamada, N. Gao and M. N. Kweon, Microbiota-Derived Lactate Accelerates Intestinal Stem-Cell-Mediated Epithelial Development, Cell Host Microbe, 2018, 24, 833–846 CrossRef CAS.
- M. Camilleri, Serotonin in the gastrointestinal tract, Curr. Opin. Endocrinol., Diabetes Obes., 2009, 16, 53–59 CrossRef CAS PubMed.
- I. S. Vahora, N. Tsouklidis, R. Kumar, R. Soni and S. Khan, How Serotonin Level Fluctuation Affects the Effectiveness of Treatment in Irritable Bowel Syndrome, Cureus, 2020, 12, e9871 Search PubMed.
- D. J. Walther and M. Bader, A unique central tryptophan hydroxylase isoform, Biochem. Pharmacol., 2003, 66, 1673–1680 CrossRef CAS PubMed.
- J. M. Yano, K. Yu, G. P. Donaldson, G. G. Shastri, P. Ann, L. Ma, C. R. Nagler, R. F. Ismagilov, S. K. Mazmanian and E. Y. Hsiao, Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis, Cell, 2015, 161, 264–276 CrossRef CAS PubMed.
- A. Shimotoyodome, S. Meguro, T. Hase, I. Tokimitsu and T. Sakata, Sulfated polysaccharides, but not cellulose, increase colonic mucus in rats with loperamide-induced constipation, Dig. Dis Sci., 2001, 46, 1482–1489 CrossRef CAS.
- C. Caballero-Franco, K. Keller, C. De Simone and K. Chadee, The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells, Am. J. Physiol.: Gastrointest. Liver Physiol., 2007, 292, G315–G322 CrossRef CAS PubMed.
- H. Hardy, J. Harris, E. Lyon, J. Beal and A. D. Foey, Probiotics, prebiotics and immunomodulation of gut mucosal defences: homeostasis and immunopathology, Nutrients, 2013, 5, 1869–1912 CrossRef CAS.
- M. M. Hafez, Upregulation of Intestinal Mucin Expression by the Probiotic Bacterium E. coli Nissle 1917, Probiotics Antimicrob. Proteins, 2012, 4, 67–77 CrossRef CAS PubMed.
- A. F. Mattar, D. H. Teitelbaum, R. A. Drongowski, F. Yongyi, C. M. Harmon and A. G. Coran, Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model, Pediatr. Surg. Int., 2002, 18, 586–590 CrossRef CAS.
- N. S. Dykstra, L. Hyde, A. MacKenzie and D. R. Mack, Lactobacillus plantarum 299v Prevents Caspase-Dependent Apoptosis In Vitro, Probiotics Antimicrob. Proteins, 2011, 3, 21–26 CrossRef PubMed.
- G. P. Nagaraju, R. Basha, B. Rajitha, O. B. Alese, A. Alam, S. Pattnaik and B. El-Rayes, Aquaporins: Their role in gastrointestinal malignancies, Cancer Lett., 2016, 373, 12–18 CrossRef CAS PubMed.
- S. Zhu, J. Ran, B. Yang and Z. Mei, Aquaporins in Digestive System, Adv. Exp. Med. Biol., 2017, 969, 123–130 CrossRef CAS.
- J. Y. Zhang, B. Li, N. X. Gao, H. K. Li, X. Y. Cui, H. Q. Jiang, S. Y. Tang, C. Y. Jin and J. L. Tian, Preventive effect of kiwi berry on loperamide-induced constipation, Food Sci. Hum. Wellness, 2024, 13, 1410–1421 CrossRef CAS.
- D. D. Guerra, R. Bok, R. A. Lorca and K. J. Hurt, Protein kinase A facilitates relaxation of mouse ileum via phosphorylation of neuronal nitric oxide synthase, Br. J. Pharmacol., 2020, 177, 2765–2778 CrossRef CAS.
- Q. Tan, J. Hu, Y. Zhou, Y. Wan, C. Zhang, X. Liu, X. Long, F. Tan and X. Zhao, Inhibitory Effect of Lactococcus lactis subsp. lactis HFY14 on Diphenoxylate-Induced Constipation in Mice by Regulating the VIP-cAMP-PKA-AQP3 Signaling Pathway, Drug Des., Dev. Ther., 2021, 15, 1971–1980 CrossRef.
- L. Cong, L. W. Duan, W. P. Su, S. Hao and D. F. Li, Efficacy of High Specific Volume Polysaccharide - A New Type of Dietary Fiber - On Molecular Mechanism of Intestinal Water Metabolism in Rats With Constipation, Med. Sci. Monit., 2019, 25, 5028–5035 CrossRef CAS PubMed.
- J. K. Wang, W. Wei, D. Y. Zhao, H. F. Wang, Y. L. Zhang, J. P. Lei and S. K. Yao, Intestinal mucosal barrier in functional constipation: Dose it change?, World J. Clin. Cases, 2022, 10, 6385–6398 CrossRef PubMed.
- W. Zheng, Z. Zhao, Y. Yang, L. Ding and W. Yao, The synbiotic mixture of lactulose and Bacillus coagulans protects intestinal barrier dysfunction and apoptosis in weaned piglets challenged with lipopolysaccharide, J. Anim. Sci. Biotechnol., 2023, 14, 80 CrossRef CAS PubMed.
- X. Chang, M. Kang, Y. Shen, L. Yun, G. Yang, L. Zhu, X. Meng, J. Zhang and X. Su, Bacillus coagulans SCC-19 maintains intestinal health in cadmium-exposed common carp (Cyprinus carpio L.) by strengthening the gut barriers, relieving oxidative stress and modulating the intestinal microflora, Ecotoxicol. Environ. Saf., 2021, 228, 112977 CrossRef CAS.
- S. Zhang, R. Wang, D. Li, L. Zhao and L. Zhu, Role of gut microbiota in functional constipation, Gastroenterol. Rep., 2021, 9, 392–401 CrossRef PubMed.
- L. Wang, L. Hu, Q. Xu, T. Jiang, S. Fang, G. Wang, J. Zhao, H. Zhang and W. Chen, Bifidobacteria exert species-specific effects on constipation in BALB/c mice, Food Funct., 2017, 8, 3587–3600 RSC.
- L. Xu, Y. Zhou, Z. Zhan, W. Zhang, D. Fu, R. Zhao and X. Chen, Research Note: Effects of Bacillus coagulans X26 on the production performance, intestinal structure, short-chain fatty acids and flora composition of laying hens during the peak laying period, Poult. Sci., 2022, 101, 101835 CrossRef CAS PubMed.
- R. S. Madempudi, J. Neelamraju, J. J. Ahire, S. K. Gupta and V. K. Shukla, Bacillus coagulans Unique IS2 in constipation: a double-blind, placebo-controlled study, Probiotics Antimicrob. Proteins, 2020, 12, 335–342 CrossRef.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4fo04237a |
| ‡ These authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2025 |