
Adsorption behavior of carbon dots on La3+ and the multiple effects on the growth of mung bean seedlings under La3+ stress†
Xinanbei
Liu
abc,
Xianfei
Niu
ac,
Yinshuai
Tian
d,
Yue
Jiang
ac,
Cheng
Cheng
ac,
Ting
Wang
ac,
Yiran
Sun
 ac,
Fang
Chen
ac and
Ying
Xu
*ac
ac,
Fang
Chen
ac and
Ying
Xu
*ac
aKey Laboratory of Bio-Resources and Eco-Environment of Ministry of Education, College of Life Sciences, Sichuan University, Chengdu 610064, P.R. China. E-mail: xuying@scu.edu.cn; Tel: +86 (028) 8541 7281
bCollege of Resources and Environment, Baoshan University, Baoshan 678000, P.R.China
cNational and Local Joint Engineering Laboratory for Energy Plant Bio-oil Production and Application, Chengdu 610064, China
dSchool of Landscape and Ecological Engineering, Hebei University of Engineering, Handan, P.R.China
First published on 19th October 2024
Abstract
The excessive use of rare earth elements (REEs) as micro-fertilizers is harmful to agricultural production and the environment. This study explored the potential application of carbon dots (CDs) to mitigate the effects of La contamination. The results indicate that the CDs based on citric acid (C-CDs) can adsorb La3+ in an aqueous solution system through surface carboxyl and pyrrolic-N. While no significant alteration in the total La content within mung bean seedlings was observed, the presence of C-CDs induced the conversion of La into an inactive form within the body and significantly affected the chemical form and distribution of La in the plant body. However, with increased concentrations, C-CDs do not effectively improve growth inhibition of the seedlings under La stress but exacerbate it. This may be relevant to the peroxidation damage and excess extracellular precipitates. RNA-seq results showed stronger cell wall-related synthesis under C-CD and La co-treatment than in La treatment, which indicated the important role of the cell wall in this process. Although many issues remain to be addressed, this study demonstrates that C-CDs possess distinct advantages in remediating soil La contamination without significantly impeding the plant's La absorption, thus exhibiting considerable potential for agricultural application.
Environmental significance“Hormesis” effects show that high lanthanum (La) concentrations adversely affect plants. The widespread use of REEs and phosphate fertilizers, plus global warming, raises La levels in farmland, which is harmful to agriculture. Compared to the traditional approach of isolating heavy metals from the external environment, the carbon quantum dot treatment used in this study provides a novel strategy for excessive agricultural REEs. The two major findings in this study are: a) C-CDs promoted the in vivo accumulation of La precipitates in plants without significantly impeding their translocation into the plant body and b) C-CDs altered the chemical form and subcellular distribution of La in vivo which is helpful to alleviate the growth stress caused by excessive REEs at the appropriate concentration, and the C-CDs exacerbated the growth inhibition of plants with the increased La3+ which may be relevant to the cell wall response and peroxidation. This method enables plants to enrich excessive La while maintaining their normal growth. |
1 Introduction
The increasing use of rare earth elements (REEs) in agriculture and industry has raised concerns about their environmental impact. Among the REEs, cerium (Ce) and lanthanum (La) are the most commonly used elements in agriculture.1–3 Much evidence has accumulated in support of the view that small amounts of REEs may favor many aspects of plant development and growth, including root growth, yield formation, photosynthesis efficiency, antioxidative enzyme activities, photochemical efficiency, mineral nutrition, phytohormonal balance, and stress tolerance.3–8 However, plants have the capability to bioaccumulate rare earth elements (REEs) stemming from diverse anthropogenic activities, encompassing REE mining and refining operations, and the discharge of industrial residues containing REEs, as well as agricultural practices.9–13 Thus, with continuous accumulation, unfavorable levels of soil REEs in local areas are inevitable. Farming practices, being a pivotal contributor to rare earth element (REE) contamination in soil, are exerting a significant influence on the soil's REE content.14–16 For example, REE fertilizers and phosphate-based fertilizers contain REE impurities that are continuously emitted to the soil.15,17 These kinds of REEs are mainly in soluble forms, which are much more soluble and reactive than the average soil pool.1,2,18 In addition, REE bioaccumulation in soil may be enhanced in a warmer world.19–21 Globally reported concentrations spanning a broad spectrum of REEs range from below 15.9 mg kg−1 to as high as 249.1 mg kg−1, with an average of 6.6–50 mg kg−1 La2O3 in soil per kg.14,15,22 As their “hormesis” effects, high concentrations of REEs exhibit negative impacts such as the inhibition of Ca ion channels, modifications of ultrastructures, oxidative stress, regulation of endogenous hormones, or micro-interference with functional proteins on the plants.2,23–27 The presence of excessive REEs in soil also poses adverse effects on the fauna residing within, exemplified by the significant inhibition of growth and reproduction in nematodes at La3+ concentrations exceeding 10 μmol L−1.28 Furthermore, concentrations of lanthanum oxide (La2O3) surpassing 500 mg kg−1 in soil have been shown to induce pronounced neurotoxic effects on earthworms, a crucial component of soil ecosystems.29 Even more alarming, the incorporation of rare earth elements (REEs) into the human food chain, facilitated by plants and animals through multifarious pathways, poses significant threats to human health.14,30,31However, current methods for heavy metal-contaminated soils may not be suitable for minor REE-contaminated soils. Soil leaching is one of the most common methods in the treatment of heavy metal-contaminated soil. Leaching agents such as low-molecular organic ligands showed good capacity for REE-contaminated soil remediation.32,33 However, it must be noted that the subsequent acidity change and the leaching of nutrients and minerals are unfavorable for the cropland and cultivation.34 Compared to leaching, phytoremediation is deemed as an ideal, green, and cost-effective remediation technology to restore REE-contaminated soils to arable farmland,35–37 but the generation of highly contaminated plant materials and potential allelopathy will be the new problem.38,39
Carbon quantum dots (CDs) are a selective solution compared to the above methods. As a new type of carbon nanomaterial, CDs have many excellent properties including good biocompatibility and water solubility, easy functionalization, and low bio-toxicity due to their non-toxic carbon backbone.40,41 In agriculture, CDs have attracted interest owing to their advantages in improving the growth and production of plants.42 Attributed to the abundant active groups on the surface, CDs could latch metal ions by electric adsorption. For instance, CDs could alleviate the toxicity of Cd to plants by adsorbing the environment Cd ions and preventing their accumulation in plant bodies.43,44 However, barely a few studies exist on the remediation of minor agricultural REE-contaminated soils by CDs, therefore we conducted a study to contribute to the body of knowledge in this field.
Furthermore, to explore the mechanisms involved, paving the way for developing effective strategies for finding solutions, further studies were conducted. The La adsorption capacity of CDs was determined, and the morphology of the precipitation which formed from CDs and La (CDs–La) was observed using an electron microscope. Furthermore, the binding site, La chemical form, and alteration of subcellular distribution were analyzed for the follow-up mechanism study with the RNA-seq results.
2 Materials and methods
2.1. Synthesis and characterization of C-CDs
Citric acid-based CDs (C-CDs) were synthesized according to a previously published method with minor modifications.45 In brief, 3 g of urea and 3 g of citric acid were dissolved in 10 mL of deionized water. The above mixed solution was heated in a 750 W microwave oven for 5 min. The resulting brown solid was kept in a 60 °C oven for 1 h to remove small molecule substances. After that, the product was transferred to a 50 mL centrifuge tube with deionized water and centrifuged (3000 rpm, 10 min) to remove the aggregated particles. The resulting brown supernatant was purified using a 14![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 Da molecular weight dialysis bag, and the final solution was subsequently dried to obtain the C-CD powder. Afterward, the obtained C-CDs were redispersed in deionized water with different concentrations.
000 Da molecular weight dialysis bag, and the final solution was subsequently dried to obtain the C-CD powder. Afterward, the obtained C-CDs were redispersed in deionized water with different concentrations.
The size, distribution, and morphology of the synthetic C-CDs were verified by transmission electron microscopy (TEM) using a Tecnai G2 F20 S-TWIN transmission electron microscope (FEI, US). The C-CD particle distribution was determined by measuring approximately 100 C-CD particles using Image J software (Rawak Software Inc., Germany). The florescence spectra were acquired using an Infinite 200 PRO multimode reader (Tecan, Männedorf, Switzerland).
2.2. Morphology and surface analysis of the CD–La composite
After mixing C-CDs with LaCl3 solution, the morphology of CD–La particles formed in the aqueous system and their corresponding sedimentation behavior were observed. For the solution system used for morphology observation, the final concentrations of C-CDs and LaCl3 were 0.01 mg ml−1 and 30 μmol L−1, respectively. The above samples were left to stand for 2 hours after mixing to ensure the complete reaction, and the morphology of the hydrodynamic CDs–La was verified by TEM, using an FEI Tecnai G2 F20 transmission electron microscope (FEI, USA). The hydrodynamic diameter of the CDs-La was measured using a ZEN3690 Zeta Sizer (Malvern, UK).And for the solution system used to observe sedimentation behavior, the final concentrations of C-CDs and LaCl3 were adjusted to 1 mg ml−1 and 600 μmol L−1, respectively. Observation commenced from the initial mixing of the samples, with the endpoint defined as the complete accumulation of precipitates at the bottom and no visible precipitation in the supernatant. Photographs were taken and records were made every 10 minutes until the endpoint was reached. And then the above C-CDs and CD–La precipitates were collected by centrifugation (1000 rpm, 5 min), and the precipitates were dried for subsequent experiments. The scanning electron microscopy (SEM) observation of C-CDs and CD–La precipitates was performed using a JSM7500F scanning electron microscope (JEOL, Japan). And the Fourier transform infrared (FT-IR) spectra were taken on an IR trace-100 FT-IR spectrophotometer (Shimadzu, Japan). The X-ray photoelectron spectroscopy (XPS) spectra were obtained by employing an AXIS ULTRA DLD X-ray photoelectron spectroscope (AXIS, Kratos).
Additionally, to verify the generality of C-CDs' adsorption for La towards other commonly used light rare earth elements (LREEs) in agriculture, such as Ce, Pr, and Nd, experiments were also conducted by mixing 1 mg mL−1 C-CDs with 600 μmol L−1 Ce, Pr, and Nd, individually, and observing their sedimentation phenomena.
2.3. Experimental design for plant cultivation
Treatment with 150 μmol L−1 La3+ significantly inhibits the growth of mung bean seedlings.46,47 And the influence of 0.01 mg mL−1 C-CDs on plants exhibits a discernible growth promotion effect (Fig. S2†), prompting us to elevate the C-CD concentration to 0.05 mg mL−1 to facilitate a more profound exploration of the intricate interactions that occur between C-CDs and La under higher concentration conditions. The experimental design for plant cultivation is briefly as follows: mung bean (Vigna radiata (Linn.) Wilczek; Chuanyulv 1) seeds of similar size were divided into three groups randomly based on the different treatments: soaked in a 50 ml flask with water (CK), C-CDs (0.05 mg mL−1), and C-CDs (0.05 mg mL−1) with LaCl3 (150 μmol L−1), respectively. The seeds were grown at 25 °C without illumination for 3 days. After 3 days, the beakers containing mung bean seeds were placed in a constant light incubator at 25 °C for 2 days. About 5 days, mung bean sprouts were harvested for measurements. The fresh weight, total length, root length and stem length were measured. The root materials were collected for further experiments, and some of them were frozen with liquid nitrogen immediately and stored at −80 °C for subsequent RNA-seq and qRT-PCR.2.4. Determination of malondialdehyde (MDA), superoxide dismutase (SOD), peroxidase (POD), H2O2, superoxide anion and La contents
After harvesting, the roots were collected, the MDA, SOD, and POD contents of mung bean were analyzed using a corresponding kit (Jian Cheng, China), the H2O2 content was analyzed using a DAB staining kit (Solarbio, China), and the superoxide anion content was analyzed using a dihydroethidium (DHE) fluorescence probe kit (Beyotime, China).For the La content, the root, stem and leaf tissues of mung bean were divided and placed in a drying oven at 60 °C for 48 h. The tissues were separately digested in 3 mL of concentrated HNO3 at 100 °C for 1 h. After cooling the solution to room temperature, 0.5 mL of 30% H2O2 was added and mixed uniformly, and the temperature was raised again to 100 °C for half an hour. The La content in the root and leaf tissues was analyzed using inductively coupled plasma-atomic emission spectrometry (ICP-AES, Agilent 5110 (OES), USA).
2.5. Observations of the ultrastructure and La distribution pattern in vivo
The root samples treated with the method in section 2.3 were collected from mung bean sprouts after harvesting and were soaked in 5% glutaraldehyde. The tissues underwent step-by-step dehydration processes and were placed into an embedding agent. Ultrathin cross sections of the tissues were imaged using an FEI Tecnai Spirit transmission electron microscope (FEI, United States), operated at 200 kV. TEM mapping was performed using a JEM2800F transmission electron microscope (TEM, JEOL, Japan).The determination methods for the distribution of different chemical forms and subcellular compartments of La were adjusted based on the methods described by Lai et al.,48,49 as summarized below: the root samples treated with the method in section 2.3 were dried and ground into powder (passing through a 100 mesh sieve) for later use. 0.1 g of powder was weighed and treated with 80% ethanol (F1), ultra-pure water (F2), 1 M NaCl (F3), or 2% acetic acid (F4), respectively. The remaining solid fractions were digested using the wet digestion method according to NY/T 1938-2010 (F5). Each treatment solution was filtered through a 0.45 μm membrane and the La content was determined using ICP-OES. The treatment of C-CDs–La was the same as above. Additionally, 0.3 g of root samples treated with the method in section 2.3 were ground in liquid nitrogen and mixed with a solution containing 0.25 mM sucrose, 50 mM Tris-HCl (pH 7.5), and 1 mM mercaptoethanol. After being shaken well and incubated at 4 °C for 10 minutes, they were centrifuged at 1000g for 15 minutes at 4 °C. The supernatant and precipitate were digested according to NY/T 1938-2010 and the La content was determined using ICP-OES; the La contents of the supernatant and precipitate corresponded to intracellular and cell wall La, respectively.
2.6. RNA extraction and sequencing
Total RNA was extracted from the tissue using a TRIzol® reagent (Plant RNA Purification Reagent for plant tissue, USA) according to the manufacturer's instructions (Invitrogen), and genomic DNA was removed using DNase I (TaKara, USA). The samples with an RNA integrity number (RIN) ≥8 underwent subsequent analysis. The library construction and sequencing were performed at Shanghai Majorbio Bio-pharm Biotechnology Co., Ltd. (Majorbio, China) according to the manufacturer's instructions (Illumina, CA). The libraries were then sequenced with an Illumina NovaSeq 6000 sequencer (2 × 150 bp read length). A detailed description of the method is provided in the ESI† (Document S1). The transcriptome sequencing and analysis were conducted by Majorbio Bio-pharm Biotechnology Co., Ltd. (Shanghai, China). Each treatment group consisted of three independent biological replicates.2.7. Differential expression analysis and functional enrichment
To identify DEGs (differential expression genes) between two different samples/groups, the expression level of each gene was calculated according to the transcript per million read (TPM) method. RSEM (https://deweylab.biostat.wisc.edu/rsem/)50 was used to quantify gene abundances. Essentially, differential expression analysis was performed using DESeq2.51 DEGs with log2 (foldchange) ≥1 and P-adjust ≤0.05 were considered to be significantly different expressed genes. In addition, functional-enrichment analysis including GO (Gene Ontology, https://www.geneontology.org) and KEGG (Kyoto Encyclopedia of Genes and Genomes, https://www.genome.jp/kegg/) was performed to identify which DEGs were significantly enriched in GO terms and metabolic pathways at P-adjust ≤0.05 compared with the whole-transcriptome background. GO functional enrichment and KEGG pathway analysis were carried out with Goatools (https://github.com/tanghaibao/Goatools) and KOBAS (https://kobas.cbi.pku.edu.cn/home.do).52
To validate the RNA-seq results, 10 peroxidase-related DEGs were randomly selected for qPCR analysis. The specific primers of candidate genes were designed using primer 5.0 (Table S1†). According to the operating instructions of the qPCR kit (AceQ qPCR SYBR Green Master Mix, Vazyme), the PCR reaction system was 10 μL, and the amplification process was first denaturation at 95 °C for 20 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 20 s. The relative expression levels of the candidate genes normalized to UBI were calculated using the 2−ΔΔCt method. Each treatment group consisted of three independent biological replicates. The qPCR results showed that the relative expression level of candidate genes was different from that of RNA-seq, but their expression patterns were consistent (up-regulated, down-regulated), indicating the reliability of transcriptome data (Table S2; Fig. S1†).
Raw sequence reads have been uploaded to the NCBI database (BioProject accession: PRJNA956858). A visual analysis of the metabolic pathways of the DEGs was performed using Mapman software (version 3.6.0RC1, https://mapman.gabipd.org/web/guest/mapman-version-3.6.0).
2.8. Statistical analysis
Each indicator was measured three times repeatedly, and the values were expressed as means ± SD (standard deviation). The statistical analysis of experimental data was performed with one-way ANOVA based on Duncan's multiple comparison (p ≤ 0.05) in IBM SPSS version 22 (IBM, USA).3 Results
3.1. C-CDs could adsorb La3+ precipitates in an aqueous system
Citric acid-based CDs (C-CDs) were synthesized from citric acid and urea using the microwave method (Fig. 1A). The prepared C-CD aqueous solution is light yellow under natural light conditions and emits clear blue-green fluorescence under UV excitation (Fig. 1B-b). The C-CD particles were approximately spherical and evenly dispersed in the aqueous solution. Their diameters ranged from 0.8 nm to 3.2 nm. The particles with diameters of 1 nm to 2 nm accounted for 77.3% (Fig. 1A and B). No large particle impurities or obvious agglomerates were found in the field of view.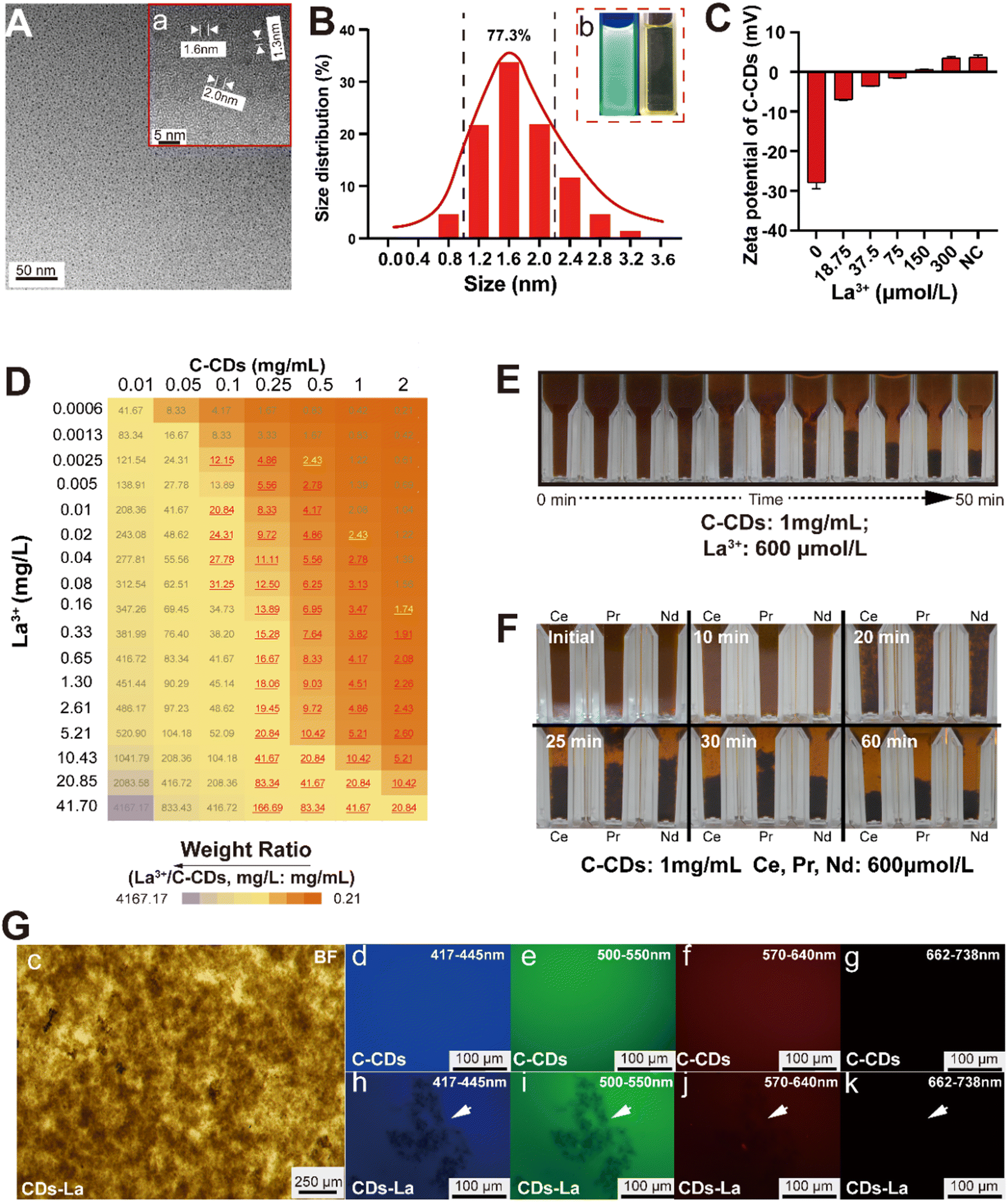 | ||
| Fig. 1 The precipitation of C-CDs and La. Note: A: TEM image of C-CDs; a: magnification box of C-CDs; B: particle size statistics of C-CDs; b: C-CDs exposed to UV light (left) and nature light (right). C: The effects of different concentrations of La cations on the zeta-potential of C-CDs; D: precipitation under different concentrations of C-CDs and La3+, the number in the heat-map means the weight ratio of La3+ and C-CDs, the red number means that precipitates could be observed, the gray number means no visible precipitation, and the yellow number means precipitation formation time more than 24 h; E: the precipitation process of C-CDs and La; F: the precipitation process of C-CDs and Ce, Pr, and Nd, respectively; G: morphology of CDs–La (c) and the fluorescence observation of CDs (d–g) and CDs–La (h–k) under different wavelength coverages. | ||
Due to the significant difference with C-CDs, precipitates which formed from C-CDs and La3+ (CDs–La) were further observed by electron microscopy. Through microscopic observation, it could be observed that CD–La particles were mainly composed of particles of varying sizes, which appeared golden yellow or brown under the microscope (Fig. 1c). Under a fluorescence microscope, CDs–La lost the fluorescence properties of C-CDs, and the aggregated CD–La particles did not show color under various excitation conditions. In the fluorescence field, they were clearly distinguishable black aggregates (Fig. 1h–k).
3.2. C-CDs could enter cells and enhance the precipitation of La
When the concentrations of C-CDs and La3+ were both low (0.01 mg mL−1 C-CDs, 30 μmol L−1 La3+), the CD–La mixture solution was still clear, and no visible precipitation could be detected with the naked eye. However, the formed CD–La complex can still be observed using the transmission electron microscope. The complex was loose, porous flocculent with dark nucleus-like structures among them. Their diameters were less than 300 nm and mainly distributed between 100 and 200 nm (Fig. 2A–C).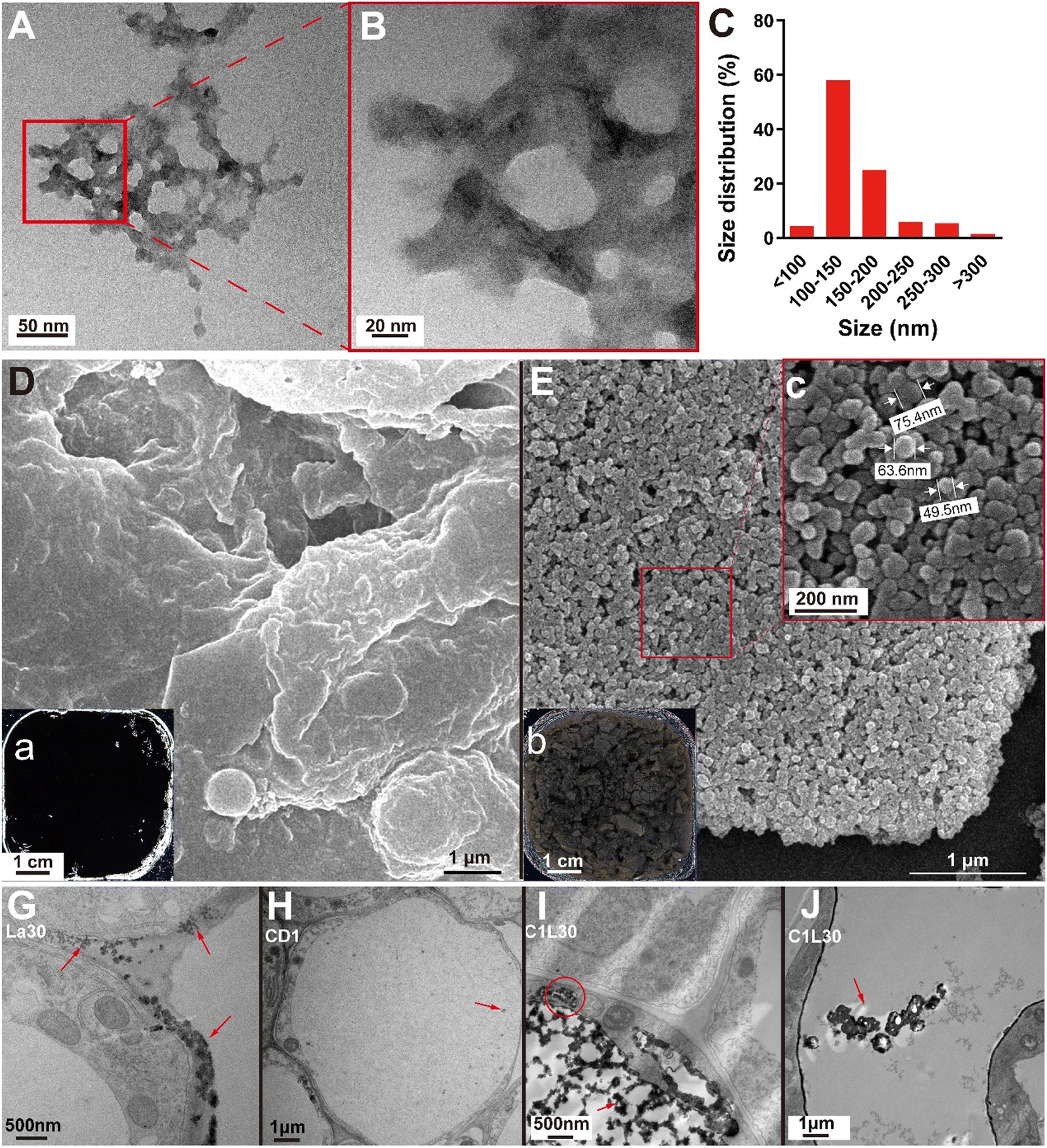 | ||
| Fig. 2 Electron microscopy images of C-CDs and the CD–La complex. Note: the SEM image with a local image (A and B) and size distribution (C) of the CD–La complex; the TEM image of the C-CD precipitates (D), CD–La complex precipitates (E) and the spherical shape and diameters of particles (c); a and b exhibit the different appearance between dried C-CD precipitates and CD–La complex precipitates. G–J show the precipitates under the root ultrastructure observation, the seedlings were treated with 30 μmol L−1 LaCl3 (G), 0.01 mg mL−1 C-CDs (H), and 0.01 mg mL−1 C-CDs +30 μmol L−1 LaCl3 (I and J), respectively. The red arrows indicate intracellular precipitation; the red arrow indicates the precipitate position, and the red circle in I indicates the observed vesicle structures. | ||
The morphology of the dried CD–La complexes was also significantly different from that of the dried C-CD complexes. The former were relatively loose gray mud-like substances (Fig. 2b), while the latter were black precipitates with a dense and smooth surface (Fig. 2a). Under scanning electron microscopy, the microstructure of CD–La precipitates was composed of circular or quasi circular particles smaller than 100 nm, with a loose surface and numerous pores (Fig. 2E), forming a significant difference from the dense C-CD precipitates (Fig. 2D).
The pores of plant cell walls are generally between 5 and 20 nm,53 suggesting that the CD–La complexes formed outside the cell are difficult to freely diffuse into the cell body. However, we found that some precipitates existed in the roots of C-CD and La co-treated seedlings (Fig. 2I), which are distinct from La precipitates (Fig. 2G) and C-CD precipitates (Fig. 2H). The cross-linked structure of the precipitate in the C-CD and La co-treated group is similar to that observed in vitro before (Fig. 2A), and a bigger precipitate in micro-size could be observed intracellularly. Therefore, one potential explanation is that the adsorption between C-CDs and La3+ does not only occur in vitro, but also in vivo; meanwhile, further adsorption may occur among the CD–La precipitates after entering the cells.
3.3. Carboxyl and pyrrolic-N groups on the surface of C-CDs might play key roles in the adsorption of La3+
To learn more details about the interaction between C-CDs and La3+, the surface functional groups of C-CDs and CDs–La were further characterized by FT-IR and X-ray photoelectron spectroscopy (XPS) methods (Fig. 3). The FT-IR spectrum of C-CDs (Fig. 3A) showed a broad absorption band at 3750–3300 cm−1, which is generally considered to be caused by the stretching vibrations of N–H and O–H. The peak at 3000–2700 cm−1 corresponds to the stretching vibrations of CH– bonds. The peak at 1900–1650 cm−1 corresponds to the vibrations of COO– bonds. The absorption bands at 1460–1350 cm−1 were assigned to the stretching bending vibrations of CH and the bands at 1360–1020 cm−1 were attributed to the stretching vibrations of C–N. The peak detected at 1200–1000 cm−1 represents the C–O stretching vibration.45 Compared to that of C-CDs, the FTIR spectrum of the CDs–La contained representative C-CD absorption bands of the coupled higher absorption bands at 2975 cm−1, 2927 cm−1, 1049 cm−1, and 881 cm−1, which were ascribed to the stretching vibrations of the La–O bonds.54,55 These results demonstrated that La3+ was doped into C-CDs primarily by the formation of La–O bonds.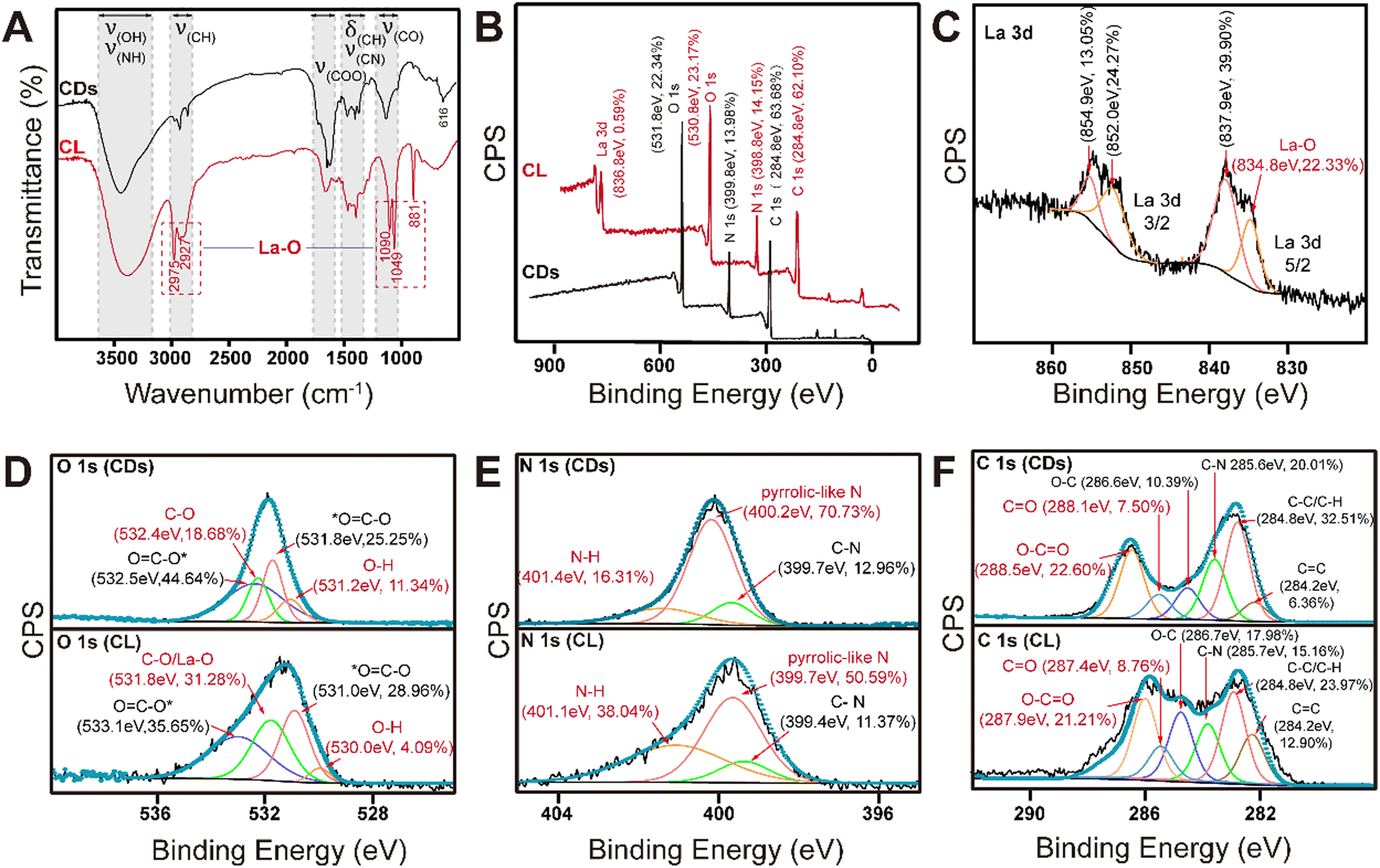 | ||
| Fig. 3 FT-IR and XPS analysis of C-CDs and the CD–La complex. Note: A: the FT-IR spectra of C-CDs (CDs, black) and CDs–La (CL, red); B: the XPS survey spectra of C-CDs (black) and CDs–La (red); C: the La 3d high resolution spectrum of CDs–La; D–F: the O1s, N 1s, and C 1s high resolution spectra of C-CDs and CDs–La. | ||
According to XPS spectroscopy results, there are three peaks corresponding to C (ca. 284.8 eV), N (ca. 399.8 eV) and O (ca. 531.8 eV) on the surface of C-CDs. Their composition ratio is 63.68%:13.98%:22.34%. Meanwhile, the CDs–La were composed of 62.10% C (ca. 284.8 eV), 14.15% N (ca. 398.8 eV), 23.17% O (ca. 530.8 eV), and 0.59% La (ca. 836.8 eV). Subsequently, the O 1 s XPS spectrum of C-CDs (Fig. 3D) could be deconvoluted into peaks at ca. 532.5 eV, ca. 532.4 eV, ca. 531.8 eV, and ca. 531.2 eV, suggesting the existence of O![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) C–O*, C–O, *OC–O, and O–H. The deconvoluted N 1s XPS spectrum of C-CDs (Fig. 3E) showed peaks at 401.40 eV, 400.20 eV, and 399.70 eV which corresponded to the nitrogen atom of N–H, pyrrolic-like N, and C–N, respectively. The C 1s XPS spectrum of C-CDs (Fig. 3F) illustrated six components corresponding to OC–O at ca. 288.50 eV, CO at ca. 288.10 eV, C–O at ca. 286.6 eV, C–N at ca. 285.60 eV, C–C/C–H at ca. 284.80 eV, and CC at ca. 284.2.56–58
C–O*, C–O, *OC–O, and O–H. The deconvoluted N 1s XPS spectrum of C-CDs (Fig. 3E) showed peaks at 401.40 eV, 400.20 eV, and 399.70 eV which corresponded to the nitrogen atom of N–H, pyrrolic-like N, and C–N, respectively. The C 1s XPS spectrum of C-CDs (Fig. 3F) illustrated six components corresponding to OC–O at ca. 288.50 eV, CO at ca. 288.10 eV, C–O at ca. 286.6 eV, C–N at ca. 285.60 eV, C–C/C–H at ca. 284.80 eV, and CC at ca. 284.2.56–58
The XPS spectrum of the CDs–La contained similar components to that of C-CDs, but the difference was mainly in the binding energy and component proportion. Compared to the C-CDs, the O 1s XPS spectrum of the CDs–La showed a stronger peak at ca. 531.8 eV. Combined with the weaker O–H peak at ca. 530.0 eV, the peak at ca. 531.8 eV could be ascribed to C–O and also corresponds to La–O.59 Similarly, the N 1s XPS spectrum of the CDs–La showed a stronger peak at ca. 401.1 eV (N–H) and a weaker peak at ca. 399.7 eV (pyrrolic-like N). And the C 1s XPS spectrum of the CDs–La indicated the migration of O–CO and CO to higher binding energies.
Furthermore, as a result of a substituent, peaks at binding energies of ca. 851.4 eV (2/3) and ca. 837.9 eV (5/2) were observed in the La 3d XPS spectrum of CDs–CL (Fig. 3C). These results demonstrated that the La3+ cation could bond to the surface groups of C-CDs and the COOH group and pyrrolic-like N should be the primarily substituent position. And the enhanced La3+ adsorption ability of C-CDs can be attributed to the formation of La–O bonds between the carboxylic groups and pyrrolic-like nitrogen. The plant response to heavy metal ion entry commonly involves combination with phosphate. As depicted in Fig. 4, C-CDs not only exhibit significant adsorption of La3+ in aqueous solution (Fig. 4A), but also demonstrate superior adsorption capacity in the presence of phosphate, surpassing even citric acid (Fig. 4B).
 | ||
| Fig. 4 Adsorption of La by C-CDs, citric acid, and phosphate. Note: A: adsorption of La3+ with different concentrations of C-CDs, the concentration of La3+ is 40 μmol L−1. B: Relative La content in the supernatant before and after treatments; C: relative P content in the supernatant before and after treatment. CLP: 0.05 mg mL−1 sodium citrate + 1.200 mM LaCl3 + 25 mM NaH2PO4; CDLP: 0.05 mg mL−1 C-CDs + 1.200 mM LaCl3 + 25 mM NaH2PO4; LP: 1.200 mM LaCl3 + 25 mM NaH2PO4. Different uppercase letters (a, b, c) indicate significant differences between different treatment groups, and “ns” represents no significant difference (P < 0.05). | ||
3.4. C-CDs altered the chemical form and subcellular distribution of La in vivo
The adsorption capacity based on the carboxyl group and pyrrole nitrogen can also be reflected under higher concentration conditions. As shown in Fig. 5, precipitates in the roots of mung bean were observed both in La150 and C5L150 groups, and the morphology observation of precipitates in vivo was carried out by TEM. In many morphology characteristics, the precipitates in La150 and C5L150 are similar; they are basically needle-shaped crystallites, have a relatively dense core, and are mainly in a grained form, in or adjoining the cell wall or dispersed in the intercellular space (Fig. 5A and B, a and b). However, the TEM-mapping shows that the La and P contents of precipitates in the La150 and C5L150 groups were distinct, which indicated the different compositions of precipitates in the two groups (Fig. 5c–h). Further confirmations were performed by ICP-OES; the chemical form of La was distinct between the La150 and C5L150 groups (Fig. 5D). Compared to the CD–La precipitates in vitro, the precipitates in vivo contained a significant portion of La in the residual form, which may be attributed to the plant in vivo precipitation mechanism, such as phosphates. Totally, the ratio of La in less active forms was significantly lower than that in the La150 group, for instance, the La in the residual form was 16.4% and 21.6% in the La150 and C5L150 groups, respectively. In contrast, the ratio of La in active forms in the C5L150 group was significantly lower than that in the La150 group, for instance, the La in the inorganic state in the C5L150 group was 0.7%, and that in the La150 group was 5.6% (Fig. 5D). And subsequent La distribution analysis further revealed the different precipitation behavior between the two groups in vivo. In the C5L150 group, the La distributed intracellularly was 13.9% and that extracellularly was 86.1%, and those in the La150 group were 25.5% and 74.5%, respectively. And a further reduction was observed when an endocytosis inhibitor (tyrphostin A23) existed (Fig. 5E). In addition, the C-CD treatment has a distinct characteristic, that is, it does not affect the total content of La in plants (Fig. 5C and S3†). | ||
| Fig. 5 Precipitate morphology in La150 (A, a) and C5L150 groups (B, b), TEM mapping of precipitates in La150 (c–e) and C5L150 groups (f–h), total La content (C), chemical form (D) and subcellular distribution (E) of La in the seedling roots of La150 and C5L150 groups. Note: materials in CK, CD5, La150, and C5L150 groups were treated with water, 0.05 mg mL−1 C-CDs, 150 μmol mL−1 LaCl3, and 0.05 mg mL−1 C-CDs + 150 μmol mL−1 LaCl3, respectively. Different uppercase letters (a, b, c) indicate significant differences between different treatment groups, and “ns” represents no significant difference (P < 0.05). F1–F5 in D mean inorganic metals (F-ethanol), water-soluble metals of organic acid complexes (F-dH2O), pectate-integrated and protein-integrated metals (F-NaCl), phosphate-sequestered metals (F-HAc), and insoluble metals (F-residue), respectively. | ||
3.5. C-CDs altered the growth state of mung bean seedlings under lanthanum stress
Rare earth elements have a typical “hormesis” effect, for instance, Ce can promote the growth of Arabidopsis thaliana at concentrations below 50 μmol L−1, while above this concentration, a gradually increasing inhibitory effect is observed.60 In this study, the 150 μmol L−1 La treatment concentration also significantly inhibited the growth of mung bean seedlings. As shown in Fig. 6A, the morphology of mung bean seedlings was significantly different between the four groups; the growth of seedlings and roots in La150 (150 μmol L−1 LaCl3 treatment) and C5L150 (150 μmol L−1 LaCl3 + 0.05 mg ml−1 C-CD treatment) was significantly worse than that in the control and CD5 groups (0.05 mg ml−1 C-CD treatment). Compared to the control, the total length and root length number of mung bean seedlings in the La150 group decreased by 16.5% and 23.1%, respectively. And these decreases were 31.3%, and 32.4% in the C5L150 group, respectively (Fig. 6C and D). In terms of morphological results, C-CDs did not alleviate the growth stress of seedlings caused by excessive La, but even partly aggravate the stress.
 | ||
| Fig. 6 Effect of C-CD and La treatments on the morphology (A), H2O2 and superoxide anion levels (B), total length (C), root length (D), and MDA (E), CAT (F), SOD (G), and POD (H) levels of mung bean seedlings. Note: the seedlings of each group were treated with water (CK), 0.05 mg mL−1 C-CDs (CD5), 150 μmol L−1 La (La150), and 0.05 mg mL−1 C-CDs + 150 μmol L−1 La (C5L150), respectively. The H2O2 and superoxide anion levels shown in b were determined using the DAB staining method and a DHE fluorescence probe, respectively. Different uppercase letters (a, b, c) indicate significant differences between different treatment groups, and “ns” represents no significant difference (P < 0.05). | ||
3.6. Analysis based on the RNA-seq
Relevant studies have shown that carbon dots can affect plant growth at the level of gene regulation through related genes such as in vivo peroxidation and heavy metal transport.61 Additionally, in this study, both carbon dots and La exhibited the characteristic of root accumulation, which had a significant impact on the root growth of plants. Therefore, to confirm the above inference and make a clearer understanding of the mechanisms on how C-CDs affect La3+in vivo, RNA-seq was performed based on the mung bean root materials from previous experiments. By PCA analysis, the La150 and C5L150 groups could be distinguished from the CD5 and control groups clearly. Principal component 1 explained 48.79% of the differences among all the samples, and these differences were primarily induced by La treatments (Fig. 7A). | ||
| Fig. 7 PCA analysis (A) of transcriptome, counts of upregulated and downregulated DEGs (B), summary of genes expressed in each treatment group and genes co-expressed (C), and GO and KEGG functional notes of the La150 and C5L150 groups (D). Note: materials in CK, C-CD, La, and CL groups were treated with water, 0.05 mg mL−1 C-CDs, 150 μmol mL−1 LaCl3, and 0.05 mg mL−1 C-CDs + 150 μmol mL−1 LaCl3, respectively. The pathways highlighted in red indicate that there were no significant changes in this pathway within the other group. | ||
The counts of upregulated and downregulated DEGs in each comparison are shown in Fig. 7B. A total of 438 upregulated and 402 downregulated DEGs were identified in the samples of the CD5 group, and these numbers in the La150 and C5L150 groups were 1313 and 1609, and 1653 and 1531, respectively. The summary of genes expressed in each treatment group and genes co-expressed is shown in Fig. 7C. A total of 106 genes were expressed commonly, with 154, 469, and 738 genes shared between the CD5, La150, and C5L150 groups, respectively. Besides that, a total of 1082 genes were expressed commonly between the La150 and C5L150 groups, respectively (Fig. 7C). Compared with the La150 and C5L150 groups, the total DEGs in the CD5 group were significantly lower, combined with the similar seedling morphology to the control group; thus we mainly compared the difference between the La150 and C5L150 groups (Fig. S3 and S4†).
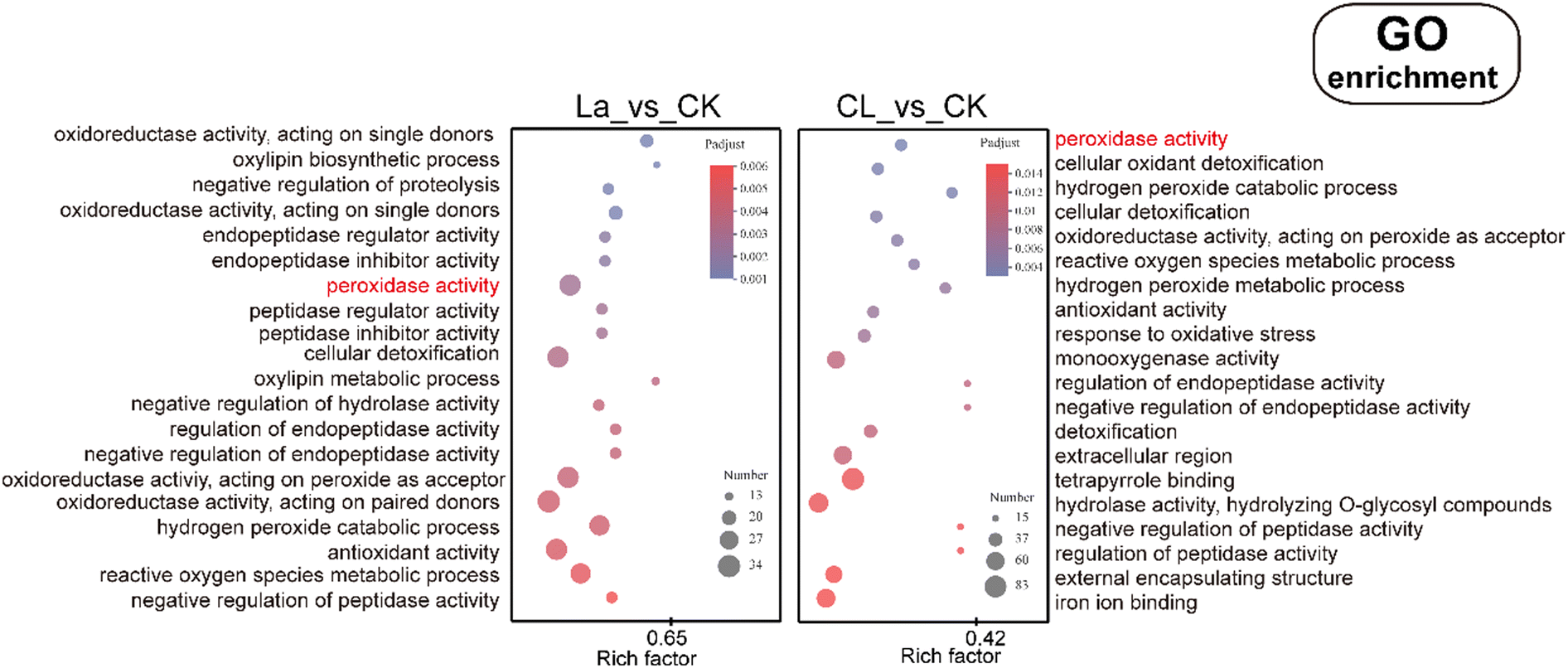 | ||
| Fig. 8 GO enrichment of DEGs in the La and CL groups. | ||
Through further screening, 17 and 16 DEGs related to peroxidase were obtained in the La150 and C5L150 groups, respectively. The up-regulation numbers were both 10, and the down-regulation numbers were 7 and 6 in the La150 and C5L150 groups, respectively. Although the total number of DEGs and the number of up-regulation and down-regulation are basically the same, the genes involved are quite different. Among the DEGs related to peroxidase in the two groups, 11 genes are the same, and there is no significant difference in their expression (Table 1). For different DEGs, there are two peroxidase-related DEGs expressed in the apoplast or cell wall in La150, and three expressed in the endoplasmic reticulum, plasma membrane, nucleus or cytoplasm, respectively, while all peroxidase-related DEGs in C5L150 are expressed on the membrane system (1 plasma membrane, 3 endomembrane system). In general, the results of GO analysis reflect the different peroxidation statuses of the two groups of plants, and the coexistence of similarity and difference in peroxidase-related DEGs can correspond to some extent with the previous results (Fig. 6).
| ID | Function | La_vs_CK | CL_vs_CK | Location |
|---|---|---|---|---|
| rna11354 | Peroxidase activity, heme binding | −7.69 | −7.56 | Endomembrane system |
| rna15138 | Peroxidase activity, heme binding | −6.85 | −6.72 | Endomembrane system |
| rna15139 | Peroxidase activity, heme binding | −6.56 | −6.43 | Endomembrane system |
| rna17727 | Protein binding, peroxidase activity | 2.52 | 3.29 | Apoplast, cell wall |
| rna17731 | Protein binding, peroxidase activity | 3.27 | 3.75 | Apoplast, cell wall |
| rna44057 | Protein binding, peroxidase activity | 2.59 | 2.81 | Apoplast, cell wall |
| rna19869 | Peroxidase activity, heme binding | 3.46 | 3.94 | Endomembrane system |
| rna26850 | Peroxidase activity, heme binding | −4.52 | −4.56 | Endomembrane system |
| rna26851 | Peroxidase activity, heme binding | 3.79 | 3.68 | Endomembrane system |
| rna43733 | Protein binding, peroxidase activity | −2.83 | −3.99 | Apoplast, cell wall |
| gene28761 | Protein binding, peroxidase activity | 4.99 | 4.82 | Apoplast, cell wall |
| rna45349 | Peroxidase activity, heme binding | −7.52 | −4.44 | Endomembrane system |
| rna28652 | Peroxidase activity | −6.87 | — | Endoplasmic reticulum |
| rna24654 | Peroxidase activity, heme binding | −2.59 | — | Nucleus, cytoplasm |
| rna35222 | Peroxidase activity, heme binding | 2.51 | — | Plasma membrane |
| rna17730 | Protein binding, peroxidase activity | −2.72 | — | Apoplast, cell wall |
| rna17214 | Protein binding, peroxidase activity | −4.34 | — | Apoplast, cell wall |
| rna35220 | Peroxidase activity, heme binding | — | −2.76 | Plasma membrane |
| rna39145 | Peroxidase activity | — | −3.63 | Endomembrane system |
| rna45348 | Peroxidase activity, heme binding | — | −3.16 | Endomembrane system |
| rna45674 | Peroxidase activity, heme binding | — | −2.76 | Endomembrane system |
The pathway enrichment results were similar in the La150 and C5L150 groups (Fig. 9A). The most enriched pathways in both groups are phenylpropanoid biosynthesis, which also involves linoleic acid biosynthesis, isoflavonoid biosynthesis, MAPK signaling, and cutin and wax biosynthesis. Phenylpropanoid biosynthesis is directly related to lignin synthesis, and lignin synthesis is also connected with flavonoid synthesis. The MAPK signaling pathway is an important conserved signaling pathway in plant stress responses. Linoleic acid metabolism is not only directly related to stress metabolism, but also directly related to the structure and function of cell membranes. Therefore, from the results of KEGG analysis, it can be inferred that both groups of plants have produced stress responses, and cell wall-related metabolism plays an important role in this process.
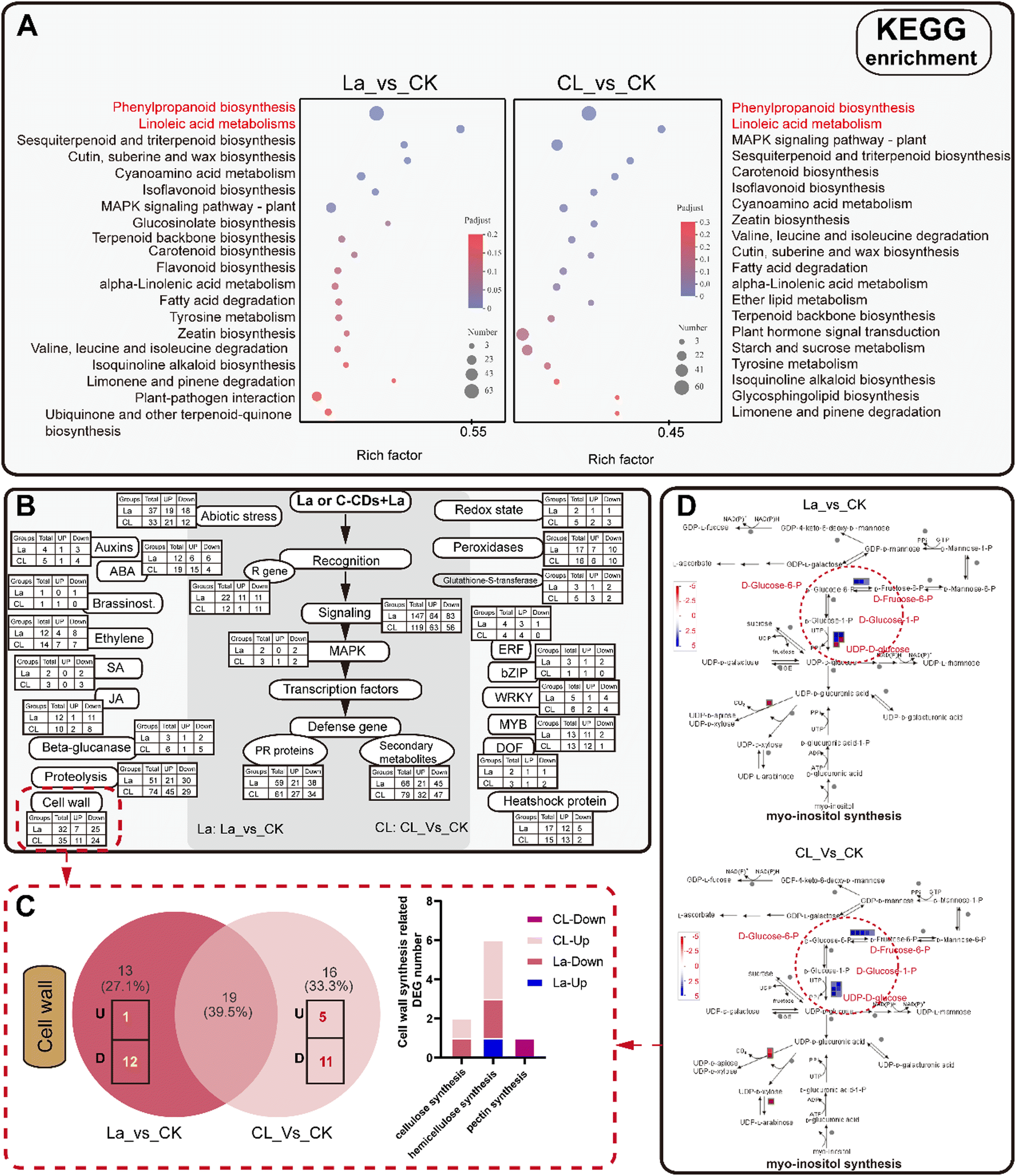 | ||
| Fig. 9 Results of KEGG enrichment (A) and the analysis of stress response-related genes (B), indicating the different expression of cell wall-related genes (C) in the La and CL groups, including the myo-inositol-synthesis related genes (D). Note: La and CL (C-CDs + La) mean seedlings were treated with 150 μmol L−1 LaCl3 and 0.05 mg mL−1 C-CDs + 150 μmol L−1 LaCl3, respectively. “UP”, “U” or blue boxes in A–C mean upregulated, and “down”, “D” or red boxes mean downregulated. | ||
Further analyses were conducted using Mapman software (version 3.6.0RC1). Results show that abundant DEGs are distributed in stress response-related pathways, including the redox state, peroxidase, signaling, abiotic stress, PR proteins, secondary metabolites, proteolysis, cell wall, and defense-related transcription factors (Fig. 9A). Significant changes in cell wall-related genes were found in the La150 and C5L150 groups (Fig. 9B, red dotted box). Unlike the results of KEGG analysis, the numbers of cell wall-related DEGs in both groups are similar, but further comparison reveals significant differences in the involved genes and their expression (Fig. 9C).
As shown in Fig. 9C, the number of cell wall-related DEGs (differential expressed genes) is 32 in the La150 group, including 7 up-regulated DEGs and 25 down-regulated DEGs, and that number in the C5L150 group is 35, including 11 up-regulated DEGs and 24 down-regulated DEGs, respectively. Within those DEGs, 19 genes were differentially expressed in the two groups. The main difference existed in the other uncommon DEGs, such as cellulose synthesis, hemicellulose synthesis, and pectin synthesis related genes. Cellulose, hemicellulose, and pectin are the main components of the cell wall, and the synthesis-related DEGs of these components in C5L150 were mainly upregulated, which in La150 were mainly downregulated. Meanwhile, enhanced cell wall precursor (myo-inositol) synthesis in the seedlings of the C5L150 group was revealed based on the greater number of relevant up-regulated genes compared to that in the La150 group (Fig. 9D). These results indicated a more frequent cell wall synthesis which may respond to the increased stress on the cell wall in the C5L150 group, compared to the La150 group.
 | ||
| Fig. 10 The root cell morphology of La150 (A; a1 and a2 are under different magnifications) and C5L150 groups (B, b1 and b2 are under different magnifications). Note: the seedlings of each group were treated with 150 μmol L−1 La (La150) and 0.05 mg mL−1 C-CDs + 150 μmol L−1 La (C5L150), respectively. | ||
4 Discussion
4.1. The possible reasons for the effects of C-CDs on the growth status of mung bean seedlings under La stress
Combining the above results, we can find similarities and differences between the La150 and C5L150 groups. From the morphology perspective, both plants in the La150 and C5L150 groups have undergone significant growth inhibition. The growth inhibition observed in the C5L150 group seems more severe, its causes are rather complex, and the positive effects of C-CDs could be observed in this study, but overall they tended to exacerbate growth stress. Although the exact reasons for the aggregation of growth inhibition of mung bean seedlings by C-CDs under high concentration conditions are not fully understood, they are at least related to two reasons: 1) peroxidation damage and 2) the cell wall status.The impact of C-CDs on the peroxidation status under La stress may be linked to the reduction of active La in vivo after C-CD treatment. Entry of heavy metal ions into cells will lead to a series of negative impacts, for example the generation of reactive oxygen species (ROS), resulting in oxidative damage to lipids, proteins and nucleic acids.63 A previous study indicated that La also leads to the accumulation of ROS in roots.64 In general, REE cations taken up by plant cells form complexes with organic compounds and are immobilized during transport by chemical precipitation (mainly phosphate particles).65–67 C-CDs could enhance this precipitation process (Fig. 4). In the C5L150 group, La in insoluble and phosphate-sequestered forms was 10% more than that in the La150 group. In contrast, La present in the inorganic form, water soluble form of organic acid complexes, and pectate-integrated and protein-integrated forms was significantly decreased in the C5L150 group compared to the La150 group (Fig. 5D).
Besides that, the subcellular distribution ratio of intracellular and extracellular La in the La150 group is approximately 1:3, while that in the C5L150 group is nearly 1:6. When the endocytosis inhibitor (tyrphostin A23) was added, this ratio was further increased to 1:14 (Fig. 5E). Therefore, the cell wall outside the cytoplasm blocked the majority of La, and the reduction of intracellular La is beneficial for reducing the La triggered ROS response.
In summary, C-CDs displayed good capacity to adsorb La3+ and alter the chemical state and subcellular distribution of La, reducing the peroxidation pressure caused by La and potentially alleviating peroxidation levels in plants. Therefore, the formation of a different peroxidation status in the C5L150 group compared to the La150 group may be related to other factors. One possible explanation is that C-CDs themselves can lead to a decrease in peroxidase activity in plants. This effect is not significant when C-CDs are applied alone, but the inhibitory effect may be significantly magnified under La stress conditions.
To the cell wall, another influential factor may be related to the precipitation between C-CDs and La3+. The cell wall contains abundant negatively charged sites for bonding La3+, and the high valence of REE ions leads to a high affinity of REEs with plant cell walls.32,66 Accumulation of precipitates in vivo seems to be a better choice to avoid the disadvantages of aggregates in vitro such as the release of contaminants.69 For example, the subcellular distribution and chemical forms of heavy metals such as Cd in plants directly affected the toxicity of these elements in plants.48 However, as the primary adherence site of intercellular precipitates, the increased intercellular precipitates may generate more stress to the cell wall. Compared to the La150 group, genes included the synthesis of the cell wall precursor myo-inositol, and the synthesis of the main cell wall structure components cellulose and hemicellulose was enhanced in the C5L150 group (Fig. 9C and D). And those stronger responses of the cell wall may lead to differences in the stele cell morphology between the two groups.
Previous studies have shown that CDs can alleviate heavy metal stress by adsorbing heavy metal ions and forming precipitates in vitro to prevent heavy metals from entering the plant.43,44,70,71 C-CDs used in this study also showed good capacity to adsorb La cations based on the surface groups, particularly COOH and pyrrolic-like N groups. The increase in the ratio of La present in insoluble and phosphate-sequestered forms in the C5L150 group led to a reduction in the active La in vivo. Besides that, in the C5L150 group, by binding to a negative site on the cell wall or forming precipitates with C-CDs and phosphates, more La was held back extracellularly (included within the cell wall) than that in the La150 group. The cell wall is the main accumulation site of heavy metals and also serves as the direct contact region of intercellular precipitates.72 In general, the pores of the plant cell wall are mainly in the range of 5–20 nm,53 but the particle size of the CD–La floc in this study was larger than that. For instance, the particles even under the conditions of 0.01 mg mL−1 C-CDs and 30 μmol L−1 La3+ were distributed in a range of 100–300 nm, the diameter of the dried subunit of CD–La precipitates was approximately 100 nm, and the needle-shaped precipitates observed in the seedlings of the C5L150 group were also significantly larger than the pores of the plant cell wall. Moreover, it seems that further adsorption could occur among the CD–La precipitates, assembling into bigger precipitates in micron-size (Fig. 2J). Hence, the size of flocs and precipitates in vitro significantly limited their entry into cells. Consequently, the CD–La precipitates in vivo predominantly resided in an intercellular space, adhering to the cell wall. As described in this study, C-CDs significantly altered the subcellular distribution ratio of intracellular and extracellular La. This also means there are more precipitates that existed extracellularly. This could be the reason for the more active expression of cell wall synthesis-related genes in the C5L150 group. Meanwhile, the cell wall plays a crucial role in abiotic stress perception and transmission. Therefore, it is one of the reasons that numerous functional pathways related to signal transduction appear in the results of KEGG analysis.
In conclusion, while the specific reasons for the exacerbation of growth inhibition of mung bean seedlings by C-CDs under La stress remain unclear, the difference in the peroxide state and extracellular precipitates played a crucial role. And it is evident that C-CDs have the potential to modulate the peroxidation status and cell wall metabolism, which may contribute to the different responses in the growth and development regulation at the gene level (Fig. S2†), ultimately resulting in the observed growth phenotypes.
4.2. The potential use in agriculture and environmental governance: C-CDs did not affect the total amount of La in the plant
One interesting finding in this study is that the precipitates mainly existed in vivo and there was no significant alteration in the total La content after C-CD treatment (Fig. 5C and S6†). Forming precipitates and accumulating in vivo can be a useful strategy in some specific situations, such as biomineralization or phytoremediation, especially the minor REE contamination caused by agricultural production or global warming.73,74 REE fertilizers and phosphate-based fertilizers contain trace levels of REE impurities that are continuously emitted to the soil.15,17 These kinds of REEs are primarily in soluble forms, which are much more soluble and reactive than the average soil pool.1,2,18 Due to the “hormesis effect”, trace La3+ favors the plant growth.75 And the results show that C-CDs have good capacity to promote the growth of mung seedlings and alleviate the growth stress caused by excessive La3+ or Ce3+ at the appropriate concentration (Fig. S7 and S8†), and similar results were observed in maize and soybean (Fig. S9†). C-CD treatment allowed soil La to be removed with the plant during the planting process in a moderate manner, leaving room for the stimulation effect of La3+ on plants. Additionally, C-CDs have many advantages for agriculture application. C-CDs as low biotoxicity materials can decompose spontaneously in the soil, and the part ingested by animals can be excreted through urine.40,45 The C-CDs applied in this study were mainly accumulated in the plant roots, which facilitates further treatment (Fig. S3†).Obviously, the corresponding negative effects described in this study may pose obstacles to the further application of this method. With the C-CD treatment, the cell wall outside the cytoplasm could block the majority of La, and intracellular La reduction could alleviate the ROS response, but the negative effects of C-CDs will be amplified with increasing treatment concentration, and the cell wall may be further stimulated due to extracellular La and the relevant precipitates. Furthermore, excessively high concentrations of C-CDs can also exert notable negative impacts on plant growth (Fig. S10†). Therefore, how to avoid excessive accumulation and optimize the application conditions of C-CDs or other similar CDs needs to be addressed in subsequent studies and should be carefully considered in practical applications.
5 Conclusion
The objective of this research was to evaluate the La3+ adsorption capacity of C-CDs both in vitro and in vivo. Although there are some issues in the application of C-CDs, this method still has many advantages that can be exploited. The results showed that C-CD treatment effectively adsorbed La3+, which was mainly attributed to the surface carboxyl and pyrrolic-N groups. The adsorption between C-CDs and La3+ had little impact on the total La content in plants. C-CDs significantly altered the chemical form and subcellular distribution of La3+, and the precipitates formed by C-CDs and La3+ were abundant in the intercellular space. C-CDs exacerbating growth inhibition may be relevant to the peroxidation damage and excess extracellular precipitates. Although the mechanisms require further investigation, this study provides a potential solution to dealing with minor REE contamination caused by agricultural activities under the background of global warming and preliminary examination of the negative effects of C-CD treatment. Some cautions are also provided for the agricultural application of small particle size nanomaterials based on the study of C-CDs.Abbreviations
| REEs | Rare earth elements |
| CDs | Carbon dots |
| C-CDs | Citric acid-based CDs |
| La | Lanthanum |
| Ce | Cerium |
| DEGs | Differential expression genes |
| CDs–La | Precipitates formed from C-CDs and La3+ |
| DAB | Diaminobenzidine |
| DHE | Dihydroethidium |
Data availability
The data that support the findings of this study are available from the corresponding author, Ying Xu, upon reasonable request.Author contributions
Xinanbei Liu: data curation; methodology; writing – original draft; funding acquisition. Xianfei Niu: data curation; methodology; Yinshuai Tian: formal analysis, conceptualization; Yue Jian: data curation; Cheng; data curation. Ting Wang; data curation. Yiran Sun: data curation; Ying Xu: conceptualization; writing – review & editing. Fang Chen: conceptualization; funding acquisition.Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.Acknowledgements
This work was supported by the Special Project for New Transgenic Technologies and Methods of Ministry of Agriculture (no. 2016ZX08010001-010), the Yunnan Provincial Education Office Scientific Research Foundation Projects (no. 2023J1121), the Doctoral Starting Foundation of Baoshan University (no. BSKY202303), and the Baoshan University National Research and Cultivation Fund Project (no. BYPY202303).References
- Z. Hu, H. Richter, G. Sparovek and E. Schnug, Physiological and Biochemical Effects of Rare Earth Elements on Plants and Their Agricultural Significance: A Review, J. Plant Nutr., 2004, 27(1), 183–220 CrossRef CAS.
- G. Tyler, Rare earth elements in soil and plant systems - A review, Plant Soil, 2004, 267(1), 191–206 CrossRef CAS.
- W. Cui, M. Kamran and Q. Song, et al., Lanthanum chloride improves maize grain yield by promoting photosynthetic characteristics, antioxidants enzymes and endogenous hormone at reproductive stages, J. Rare Earths, 2019, 37(07), 111–120 CrossRef.
- F. Hong, L. Wang and L. Chao, Study of lanthanum on seed germination and growth of rice, Biol. Trace Elem. Res., 2003, 94(3), 273–286 CrossRef CAS PubMed.
- Y. F. Xie, X. L. Cai and W. L. Liu, et al., Effects of lanthanum nitrate on growth and chlorophyll fluorescence characteristics of Alternanthera philoxeroides under perchlorate stress, J. Rare Earths, 2013, 31(8), 823–829 CrossRef CAS.
- H. Lian, C. Qin and C. Zhang, et al., Foliar-applied lanthanum chloride increases growth and phosphorus acquisition in phosphorus-limited adzuki bean seedlings, Plant Soil, 2019, 442, 385–399, DOI:10.1007/s11104-019-04197-5.
- H. Luo, Y. Chen and L. He, et al., Lanthanum (La) improves growth, yield formation and 2-acetyl-1-pyrroline biosynthesis in aromatic rice (Oryza sativa L.), BMC Plant Biol., 2021, 21, 233, DOI:10.1186/s12870-021-03006-5.
- E. D. F. W. Smith and C. J. Asher, Effects of Lanthanum and Cerium on the Growth and Mineral Nutrition of Corn and Mungbean, Ann. Bot., 2008, 101(7), 971–982 CrossRef.
- O. Wiche and H. Heilmeier, Germanium (Ge) and rare earth element (REE) accumulation in selected energy crops cultivated on two different soils, Miner. Eng., 2016, 92, 208–215 CrossRef CAS.
- C. R. M. Rao, A. Sahuquillo and J. F. Lopez-Sanchez, Comparison of single and sequential extraction procedures for the study of rare earth elements remobilisation in different types of soils, Anal. Chim. Acta, 2010, 662(2), 128–136 CrossRef CAS PubMed.
- Z. Hao, Y. Li, H. Li, B. Wei, X. Liao and T. Liang, et al., Levels of rare earth elements, heavy metals and uranium in a population living in Baiyun Obo, Inner Mongolia, China: A pilot study, Chemosphere, 2015, 128(128), 161–170 CrossRef CAS PubMed.
- L. Wang, T. Liang, Q. Zhang and K. Li, Rare earth element components in atmospheric particulates in the Bayan Obo mine region, Environ. Res., 2014, 131, 64–70 CrossRef CAS.
- B. Koen, T. J. Peter and B. Bart, et al., Recycling of rare earths: a critical review, J. Cleaner Prod., 2013, 51(14), 1–22, DOI:10.1016/j.jclepro.2012.12.037.
- M. Adeel, Y. Lee and M. Zain, et al., Cryptic footprints of rare earth elements on natural resources and living organisms, Environ. Int., 2019, 127, 785–800 CrossRef CAS PubMed.
- P. J. Thomas, D. Carpenter, C. Boutin and J. E. Allison, Rare earth elements (REEs): Effects on germination and growth of selected crop and native plant species, Chemosphere, 2014, 96(2), 57–66 CrossRef CAS PubMed.
- D. S. Todorovsky, N. L. Minkova and D. P. Bakalova, Effect of the application of superphosphate on rare earths' content in the soil, Sci. Total Environ., 1997, 203(1), 13–16 CrossRef CAS.
- A. V. Val'kov, S. I. Stepanov, V. V. Sergievskii and A. M. Chekmarev, Monazite raw material for the production of highly effective fertilizers, Theor. Found. Chem. Eng., 2010, 44(4), 497–499 CrossRef.
- P. Xin, D. Li and P. An, Application of rare-earth elements in the agriculture of China and its environmental behavior in soil, Environ. Sci. Pollut. Res., 2002, 9(2), 143–148 CrossRef PubMed.
- C.-J. Zhang, J.-P. Shen and Y.-F. Sun, et al., Interactive effects of multiple climate change factors on ammonia oxidizers and denitrifiers in a temperate steppe, FEMS Microbiol. Ecol., 2017, 93(4), fix037, DOI:10.1093/femsec/fix037.
- M. Lukac and D. L. Godbold, Soil Ecology in Northern Forests: A Belowground View of a Changing World, Cambridge University Press, 2011 Search PubMed.
- L. Kou, W. Chen and L. Jiang, et al., Simulated nitrogen deposition affects stoichiometry of multiple elements in resource-acquiring plant organs in a seasonally dry subtropical forest, Sci. Total Environ., 2018, 624, 611–620 CrossRef CAS PubMed.
- J. Li, R. A. Verweij and C. A. M. van Gestel, et al., Lanthanum toxicity to five different species of soil invertebrates in relation to availability in soil, Chemosphere, 2018, 193, 412–420 CrossRef CAS.
- K. Yuriko, I. Takashi, K. Kazuhiko, Y. Orito and K. Hiroyuki, Characterisation of lanthanum toxicity for root growth of Arabidopsis thaliana from the aspect of natural genetic variation, Funct. Plant Biol., 2007, 34(11), 984–994 CrossRef.
- L. Yangyang, S. Liangliang, Z. Ping, W. Jinpeng, W. Ruling and X. Jin, Lanthanum Inhibits Primary Root Growth by Repressing Auxin Carrier Abundances in Arabidopsis, Front. Plant Sci., 2017, 8, 1661 CrossRef PubMed.
- C. D. Oliveira, S. J. Ramos, J. O. Siqueira, Quin V. Fa, E. M. De Castro and D. C. Amaral, et al., Bioaccumulation and effects of lanthanum on growth and mitotic index in soybean plants, Ecotoxicol. Environ. Saf., 2015, 122, 136–144 CrossRef.
- L. Wang, J. He, Q. Yang, X. Li, H. Wei and D. Chen, et al., A preliminary study on the effects of lanthanum (III) on plant vitronectin-like protein and its toxicological basis, Ecotoxicol. Environ. Saf., 2017, 145, 227 CrossRef CAS.
- E. He and H. Qiu, Lanthanum and cerium disrupt similar biological pathways and interact synergistically in Triticum aestivum as revealed by metabolomic profiling and quantitative modeling, J. Hazard. Mater., 2021, 127831 Search PubMed.
- H. Zhang, X. He, W. Bai, X. Guo, Z. Zhang and Z. Chai, et al., Ecotoxicological assessment of lanthanum with Caenorhabditis elegans in liquid medium, Metallomics, 2010, 2(12), 806–810 CrossRef CAS PubMed.
- M. Adeel, N. Shakoor, T. Hussain, I. Azeem and J. Rinklebe, Bio-interaction of nano and bulk Lanthanum and Ytterbium oxides in soil system: Biochemical, genetic, and histopathological effects on Eisenia fetida, J. Hazard. Mater., 2021, 415, 125574, DOI:10.1016/j.jhazmat.2021.125574.
- D. J. Naczynski, M. C. Tan, M. Zevon, B. Wall, J. Kohl and A. Kulesa, et al., Rare-earth-doped biological composites as in vivo shortwave infrared reporters, Nat. Commun., 2013, 4, 1345–1346 CrossRef.
- X. He, T. Yuan, X. Jiang, H. Yang and C. L. Zheng, Effects of contaminated surface water and groundwater from a rare earth mining area on the Biology and the Physiology of Sprague-Dawley rats, Sci. Total Environ., 2020, 761(543), 144123 Search PubMed.
- D. Shiming, T. Liang, Z. Chaosheng, Y. Juncai and Z. Zili, Accumulation and fractionation of rare earth elements (REEs) in wheat: controlled by phosphate precipitation, cell wall absorption and solution complexation, J. Exp. Bot., 2005, 56(420), 2765–2775 CrossRef PubMed.
- A. T. Lima and L. Ottosen, Recovering rare earth elements from contaminated soils: Critical overview of current remediation technologies, Chemosphere, 2020, 265, 129–163, DOI:10.1016/j.chemosphere.2020.129163.
- H. Tang, W. Shuai, X. Wang and Y. Liu, Extraction of rare earth elements from a contaminated cropland soil using nitric acid, citric acid, and EDTA, Environ. Technol., 2016, 38(16), 1980–1986, DOI:10.1080/09593330.2016.1244563.
- R. Chaney and M. Malik, et al., Phytoremediation of soil metals, Curr. Opin. Biotechnol., 1997, 8(3), 279–284, DOI:10.1016/S0958-1669(97)80004-3.
- N. Canha, M. C. Freitas, H. M. Anawar, I. Dionísio, H. M. Dung and C. Pinto-Gomes, et al., Characterization and phytoremediation of abandoned contaminated mining area in Portugal by INAA, J. Radioanal. Nucl. Chem., 2010, 286(2), 577–582 CrossRef CAS.
- L. Zhou, Z. Li and W. Liu, et al., Restoration of rare earth mine areas: organic amendments and phytoremediation, Environ. Sci. Pollut. Res., 2015, 22, 17151–17160 CrossRef CAS PubMed.
- Z. Wei, B. Gao, K. Y. Cheng, A. H. Kaksonen, S. D. Kolev, J. W. C. Wong and J. Cui, Exploring the use of Dicranopteris pedata ash as a rare earth fertilizer to Ipomoea aquatica Forsskal, J. Hazard. Mater., 2020, 400, 123207 CrossRef CAS PubMed.
- L. C. Davide, G. Soares and L. F. Viccini, Mutagenic effects due to allelopathic action of fern (Gleicheniaceae) extracts, Allelopathy J., 2008, 22(1), 143–152 Search PubMed.
- K. Bhattacharya, S. P. Mukherjee, A. Gallud, S. C. Burkert and B. Fadeel, Biological interactions of carbon-based nanomaterials: From coronation to degradation, Nanomedicine, 2016, 12(2), 333–351 CrossRef CAS.
- M. Arnab, M. Sanghamitra, A. D. Servin, P. Luca, D. O. Parkash and J. C. White, Carbon Nanomaterials in Agriculture: A Critical Review, Front. Plant Sci., 2016, 7, 1770, DOI:10.3389/fpls.2016.00172.
- Y. Li, X. Xu, Y. Wu, J. Zhuang, X. Zhang and H. Zhang, et al., A review on the effects of carbon dots in plant systems, Mater. Chem. Front., 2020, 4, 437 RSC.
- L. Xiao, H. Guo, S. Wang, J. Li, Y. Wang and B. Xing, Carbon dots alleviate the toxicity of cadmium ions (Cd2+) toward wheat seedlings, Environ. Sci.: Nano, 2019, 6, 1493–1506 RSC.
- J. Li, L. Xiao, Y. Cheng, Y. Cheng and L. Ding, Applications of carbon quantum dots to alleviate Cd2+ phytotoxicity in Citrus maxima seedlings, Chemosphere, 2019, 236, 124385 CrossRef CAS PubMed.
- S. Qu, X. Wang and Q. Lu, A Biocompatible Fluorescent Ink Based on Water-Soluble Luminescent Carbon Nanodots, Angew. Chem., Int. Ed., 2012, 51(49), 12215–12218 CrossRef CAS.
- E. Diatloff, F. W. Smith and C. J. Asher, Effects of lanthanum and cerium on the growth and mineral nutrition of corn and mungbean, Ann. Bot., 2008, 7, 101 Search PubMed.
- E. Diatloff, C. J. Asher and F. W. Smith, The Effects of Rare Earth Elements on the Growth and Nutrition of Plants, 1998 Search PubMed.
- J. L. Lai, Z. W. Liu, L. I. Chen and X. G. Luo, Analysis of accumulation and phytotoxicity mechanism of uranium and cadmium in two sweet potato cultivars, J. Hazard. Mater., 2020, 409, 124997 CrossRef.
- F. Dai, G. Luo, Z. Li, X. Wei and C. Tang, Physiological and transcriptomic analyses of mulberry (Morus atropurpurea) response to cadmium stress, Ecotoxicol. Environ. Saf., 2020, 205, 111298 CrossRef CAS.
- C. N. Dewey and L. Bo, RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome, BMC Bioinf., 2011, 12(1), 323 CrossRef.
- M. I. Love, W. Huber and S. Anders, Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2, Genome Biol., 2014, 15(12), 550 CrossRef.
- X. Chen, X. Mao, J. Huang, D. Yang, J. Wu and S. Dong, et al., KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases, Nucleic Acids Res., 2011, 39, 316–322 CrossRef.
- N. Carpita, D. Sabularse, D. Montezinos and D. P. Delmer, Determination of the Pore Size of Cell Walls of Living Plant Cells, Science, 1979, 205(4411), 1144–1147 CrossRef CAS.
- Z. Zhang, X. E. Verykios, S. M. Macdonald and S. Affrossman, Comparative Study of Carbon Dioxide Reforming of Methane to Synthesis Gas over Ni/La 2 O 3 and Conventional Nickel-Based Catalysts, J. Phys. Chem., 1996, 100(2), 744–754 CrossRef CAS.
- S. Hui, Y. Ding, J. Duan, Q. Zhang and X. Zheng, Transesterification of sunflower oil to biodiesel on ZrO2 supported La2O3 catalyst, Bioresour. Technol., 2009, 101(3), 953–958 Search PubMed.
- X. Chen, Q. Jin, L. Wu, C. H. Tung and X. Tang, Synthesis and Unique Photoluminescence Properties of Nitrogen-Rich Quantum Dots and Their Applications, Angew. Chem., 2014, 126(46), 12750–12755, DOI:10.1002/ange.201408422.
- R. A. Xin, A. Qq, A. Xq, X. A. Wei, A. Sl and L. B. Lei, et al., Water-soluble pillar[6]arene functionalized nitrogen-doped carbon quantum dots with excellent supramolecular recognition capability and superior electrochemical sensing performance towards TNT - ScienceDirect, Sens. Actuators, B, 2018, 257, 362–371 CrossRef.
- X. Jia, J. Li and E. Wang, One-pot green synthesis of optically pH-sensitive carbon dots with upconversion luminescence, Nanoscale, 2012, 4, 5572–5575 RSC.
- L. Kubelková, J. Nováková and K. Nedomová, Reactivity of surface species on zeolites in methanol conversion, J. Catal., 1990, 124(2), 441–450 CrossRef.
- X. Wang, Y. Lin, D. Liu, X. U. Hengjian, T. Liu and F. Zhao, Cerium toxicity, uptake and translocation in Arabidopsis thaliana seedling, J. Rare Earths, 2012, 30(6), 579–585 CrossRef CAS.
- E. He and H. Qiu, Lanthanum and cerium disrupt similar biological pathways and interact synergistically in Triticum aestivum as revealed by metabolomic profiling and quantitative modeling, J. Hazard. Mater., 2022, 426, 127831 CrossRef CAS.
- R. Tenhaken, Cell wall remodeling under abiotic stress, Front. Plant Sci., 2015, 5, 771 Search PubMed.
- S. S. Sharma and K. J. Dietz, The relationship between metal toxicity and cellular redox imbalance, Trends Plant Sci., 2009, 14(1), 43–50 CrossRef CAS.
- L. Yang-Yang, W. Ru-Ling, Z. Ping, S. Liang-Liang and X. Jin, Involvement of reactive oxygen species in lanthanum-induced inhibition of primary root growth, J. Exp. Bot., 2016, 21, 6149–6159 Search PubMed.
- L. Taiz and E. Zeiger, Secondary metabolites and plant defense, Plant Physiol., 2006, 4, 283–308 Search PubMed.
- L. Brioschi, M. Steinmann, E. Lucot, M. C. Pierret, P. Stille and J. Prunier, Transfer of rare earth elements (REE) from natural soil to plant systems: implications for the environmental availability of anthropogenic REE, Plant Soil, 2013, 366(1–2), 143–163 CrossRef CAS.
- C. J. Sanderson, The uptake of a polyvalent cation and its distribution in the root apices of Allium cepa: Tracer and autoradiographic studies, Planta, 1969, 89(2), 136–154 CrossRef.
- D. He, B. Xia, Q. Zhou, L. Wang and X. Huang, Rare earth elements regulate the endocytosis and DNA methylation in root cells of Arabidopsis thaliana, Chemosphere, 2019, 227, 522–532 CrossRef CAS PubMed.
- L. Wang, J. Rinklebe, F. M. G. Tack and D. Hou, A review of green remediation strategies for heavy metal contaminated soil, Soil Use Manage., 2021, 37(4), 936–963 CrossRef.
- J. R. Cumming and G. J. Taylor, Mechanisms of metal tolerance in plants: physiological adaptations for exclusion of metal ions from the cytoplasm, Plant Biol., 1990, 12, 329–356 Search PubMed.
- M. Jiang, S. Dai, B. Wang, Z. Xie, J. Li and L. Wang, et al., Gold nanoparticles synthesized using melatonin suppress cadmium uptake and alleviate its toxicity in rice, 2021.
- T. M. Mwamba, L. Li, R. A. Gill, F. Islam, A. Nawaz and B. Ali, et al., Differential subcellular distribution and chemical forms of cadmium and copper in Brassica napus, Ecotoxicol. Environ. Saf., 2016, 134(1), 239–249 CrossRef CAS.
- Y. Zhang, L. Wang, X. Liu, C. Cao, J. Yao and Z. Ma, et al., Enhancing La(III) biosorption and biomineralization with Micromonospora saelicesensis: Involvement of phosphorus and formation of monazite nano-minerals, Sci. Total Environ., 2024, 914, 169851 CrossRef CAS.
- S. Yao and B. Zhou, Enhancing phytoremediation of cadmium and arsenic in alkaline soil by Miscanthus sinensis: A study on the synergistic effect of endophytic fungi and biochar, Sci. Total Environ., 2024, 923, 171458 CrossRef CAS.
- E. Agathokleous, M. Kitao and E. J. Calabrese, The rare earth element (REE) lanthanum (La) induces hormesis in plants, Environ. Pollut., 2018, 238, 1044–1047 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4en00530a |
| This journal is © The Royal Society of Chemistry 2025 |