
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Chiral sensing combined with nuclease activity assay to track Cas9 dynamics in solution: ROA and CPL study†
Monika
Halat
 *a,
Magdalena
Klimek-Chodacka
a,
Agnieszka
Domagała
bc,
Grzegorz
Zając
b,
Tomasz
Oleszkiewicz
a,
Josef
Kapitán
d and
Rafal
Baranski
a
*a,
Magdalena
Klimek-Chodacka
a,
Agnieszka
Domagała
bc,
Grzegorz
Zając
b,
Tomasz
Oleszkiewicz
a,
Josef
Kapitán
d and
Rafal
Baranski
a
aDepartment of Plant Biology and Biotechnology, Faculty of Biotechnology and Horticulture, University of Agriculture in Krakow, AL. Mickiewicza 21, 31-120 Krakow, Poland. E-mail: m.halat@urk.edu.pl
bJagiellonian Centre for Experimental Therapeutics (JCET), Bobrzynskiego 14, 30-348 Krakow, Poland
cDoctoral School of Exact and Natural Sciences, Jagiellonian University, Prof. S. Łojasiewicza 11, 30-348 Krakow, Poland
dDepartment of Optics, Palacký University Olomouc, 17. listopadu 12, 77146, Olomouc, Czech Republic
First published on 15th April 2025
Abstract
Chiroptical studies of the SpyCas9 protein are extremely rare. Nondestructive methods are needed to characterize its active ribonucleoprotein form. Using Raman optical activity (ROA) and circularly polarized luminescence (CPL), we present a new approach to detect key biomolecules involved in CRISPR-Cas technology while preserving their original nucleolytic activity.
Since 2012, the use of Streptococcus pyogenes SpyCas9 protein has rapidly revolutionized biological sciences1 as a primary element of site-directed mutagenesis2 (gene scissors or molecular scalpel technology utilizing CRISPR-Cas system), derived from the adaptive immune response in microbes. Given the growing interest in exploiting the CRISPR-Cas mechanism, there is an urgent need for efficient detection of active and stable ribonucleoprotein complexes (RNP) enabling targeting and cutting of any predefined DNA region under physiological conditions. For this reason, Raman optical activity (ROA) was applied and complemented by nuclease activity assay, including the induction of DNA cleavage in vitro3 and in living cells,4 and the determination of the size of the obtained FAM-labelled DNA fragments using capillary electrophoresis (see Table S2 and Fig. S1, S2 of the ESI†).
ROA enables the study of protein structure in aqueous solution,5 which is a significant advantage over the more popular X-ray crystallography,6 and, unlike NMR spectroscopy, directly reflects rapid conformational changes.7 By recording a small difference in the intensity of Raman scattered light for the right and left circular polarization (IR − IL),8 ROA provides detailed insight into the protein geometry, such as secondary and tertiary elements,9 backbone hydration,10 side-chain conformation,11 and structural arrangements present in the unfolded state (polyproline II helix, PPII).12 It is more sensitive to 3D structure than electronic circular dichroism (ECD) by reporting chirality associated with vibrational transitions of molecules.2 ROA of proteins has evolved over the years – from simple polypeptides measured in the 1990s13 to the more complex targets studied today, such as glycoproteins,14 photoreceptor proteins,15 blood plasma,16 and amyloid fibrils.17 Moreover, the ROA effect can be successfully combined with circularly polarized luminescence (CPL), which is known as the ROA-CPL approach.18 This method requires the use of luminescent probes, e.g. Eu(III) complexes, whose chirality is induced by interactions with optically active components, such as peptides,19 proteins20 or DNA.21 Matching the energy of the ROA excitation line (532 nm) to the electronic transition of the Eu(III)-based probe gives rise to an enhanced CPL signal sensitive to the local chiral environment, here the SpyCas9 nuclease and its RNP complex (Fig. 1).
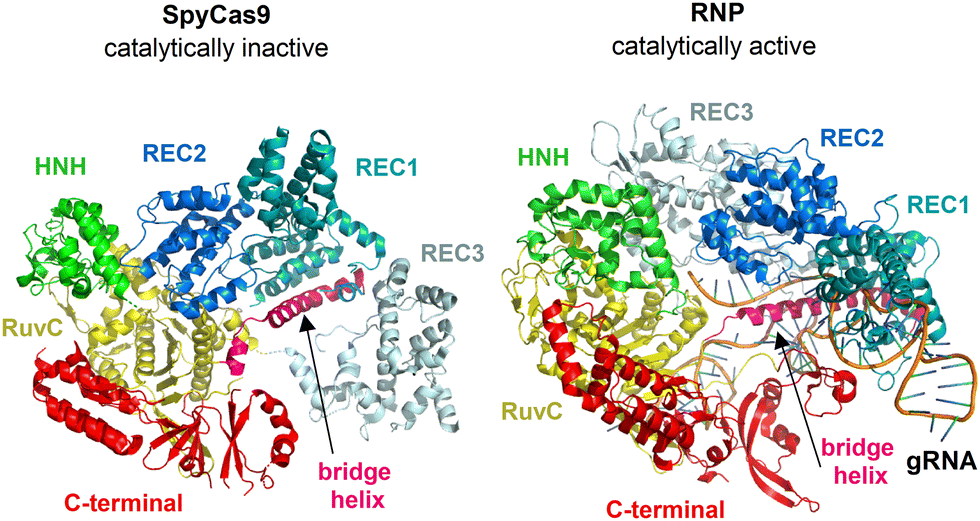 | ||
| Fig. 1 Overall structure of SpyCas9 (PDB ID: 4CMP)22a and RNP (PDB ID: 4ZT0)22b shown in ribbon representation. | ||
Therefore, we present the first, to our knowledge, ROA study combined with Eu(III)-CPL probing identifying major changes in the geometry of SpyCas9 enzyme during its catalytic activation, i.e. upon binding of a short oligonucleotide guide RNA (gRNA). Finally, we propose two biologically neutral ROA-CPL sensors: [Eu(DPA)3]3− and EuEDTA−, intended to detect initially inactive and activated forms of the SpyCas9 in solution, without the need for further genetic testing.
As shown in Fig. 2, ROA spectra of the SpyCas9 (blue) and its RNP (red) expose chiral signatures that can be directly assigned to a proper secondary structure. The first one is the amide I couplet originating from the C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O stretching vibrations of the peptide bond. In both cases the couplet is centered at ∼1650 cm−1 with the −/+ pattern of bands, indicating the right-handed α-helix.5a,9a,9d Crystallographic data22 confirms that the SpyCas9 is a bi-lobed α-rich nuclease (49.6%)23 featuring also mixed α/β domains (Fig. 1). The same order of bands is also observed for β-proteins, but it is typically shifted by ∼10 cm−1 towards higher wavenumbers.5a,9a,9d Spectral variations in the amide I range may signify some rearrangements in a protein structure. However, both ROA couplets reveal similar intensities and frequencies, regardless of whether the gRNA is attached to SpyCas9 or not.
O stretching vibrations of the peptide bond. In both cases the couplet is centered at ∼1650 cm−1 with the −/+ pattern of bands, indicating the right-handed α-helix.5a,9a,9d Crystallographic data22 confirms that the SpyCas9 is a bi-lobed α-rich nuclease (49.6%)23 featuring also mixed α/β domains (Fig. 1). The same order of bands is also observed for β-proteins, but it is typically shifted by ∼10 cm−1 towards higher wavenumbers.5a,9a,9d Spectral variations in the amide I range may signify some rearrangements in a protein structure. However, both ROA couplets reveal similar intensities and frequencies, regardless of whether the gRNA is attached to SpyCas9 or not.
 | ||
| Fig. 2 ROA spectra of SpyCas9 protein (blue) and its active RNP complex (red). Asterisks were assigned to the buffer bands (see Fig. S3, S4 and Table S3, ESI†). | ||
The most noticeable ROA differences induced by the active RNP formation arise from the extended amide III range: ∼1230–1350 cm−1, dominated by the coupled N–H and Cα–H deformations (Fig. 2, grey background). The −/+/+ pattern observed at 1255/1296/1346 cm−1 in the SpyCas9 spectrum is very sensitive to change in the α-helical conformation.10c,11a,12a There is a decrease in the intensity of the +1296 cm−1 band, which is practically invisible in the RNP profile. This marker strongly depends on the backbone geometry, defined by the two torsion angles ϕ and ψ, which can be affected by intramolecular hydrogen bonding or interaction with the solvent.10b,10c,12a ROA simulations proved that the + intensity at ∼1300 cm−1 collapses for structures with ϕ, ψ values in the bottom right of the α-helical region of the Ramachandran plot (ϕ, ψ = −59°, −44°; ϕ, ψ = −55°, −53°) or with increasing conformational freedom.10c,12a The ROA signal at +1346 cm−1 is more resistant to different ϕ, ψ combinations and is considered a robust signature of the regular α-helix (ϕ, ψ = −66°, −41°).10c,12a However, its intensity may be lower when the classical α-structure unfolds into an extended PPII formation (ϕ, ψ = −75°, 145°).5a,12a Since the ROA +1319 cm−1 band is characteristic of the PPII element and occurs in the RNP spectrum with a clearly less intense +1348 cm−1 band, such interconversion is possible. Comparing the intensity ratios of the 1319/1348 cm−1 bands before and after nuclease activation, it appears that the PPII content slightly increases upon gRNA binding. The PPII hydration manner makes the structure very flexible and highly exposed to the solvent.24 Typically, PPIIs participate in numerous protein–protein and protein–nucleic acid interactions, acting as peptide ligands or forming structural binding motifs. It follows, that PPII helices may have a significant role during the SpyCas9 → RNP transition. In fact, when gRNA associates, the α-helical lobe of SpyCas9 (REC lobe, Fig. 1) undergoes serious conformational alternations.22a,25 The number of internal hydrogen bonds decreases reducing the folding of the protein structure – the percentage of helical elements drops by a total of 5% in favor of a rise in dynamic residues (up to 20% in the REC2 domain).25b Hence, the sensitivity of ROA to distinguish two positive amide modes with Cα–H bending perpendicular (∼1300 cm−1) or parallel (∼1345 cm−1) to the Cα–N bond, is extremely valuable in tracking structural heterogeneity of α-helical sequences in solution.10b,c,12a This is a great advantage over the ECD study, which only showed a general decrease in SpCas9 α-helicity upon gRNA binding.23
The negative part of the amide III range is strongly dependent on the orientation of the protein side chains.11 The nuclease bridge helix (Fig. 1) contains eight positively charged arginine (Arg) residues that are crucial for establishing contact with the negatively charged gRNA backbone.25b,26 The shift down in wavenumbers of the −1255 → −1246 cm−1 band (Fig. 2) may be attributed to the different Arg rotamers,11a determined, by the side-chain dihedral angles: χ1 = 180°(trans) → χ1 = −60°(gauche−). The average impute of hydrophobic residues to the amide III profile should also be taken into account,11a since their side chains exposure expands after RNP complexation.25b The spectral variation observed within the second negative amide III mode (−1230 → −1222 cm−1) may be related to rearrangements in domains containing β-motifs (Fig. 1, RuvC and C-terminal parts22a,25). In early ROA experiments, the ROA feature at ∼1220 cm−1 was proposed as a characteristic of the β-turn moiety.5a,9d However, recent calculations have canceled this assignment as it may be affected by the conformation of other secondary structures.11b Nevertheless. upon interaction with gRNA, the amount of β-sheets decreases and β-turns increases by about several %,25b which may generate the down-shift of this ROA band.
An alternative method to confirm successful RNP complexation is the ROA-CPL approach using two non-chiral probes: [Eu(DPA)3]3− and EuEDTA− (Fig. 3). The Raman and ROA spectra are dominated by the total luminescence (TL) and CPL bands, respectively, assigned to selected electronic transitions of Eu(III).27 Vibrational bands of studied molecules are invisible due to their low concentrations (6.1 μM). The vibrational bands of the probes are also not recognizable under the 532 nm excitation line.28 Each achiral, racemic Eu(III)-complex (2 mM) generates a characteristic spectroscopic response under the chiral influence of SpyCas9, gRNA, and RNP (chirality induction phenomenon), without changing their catalytic activity (see Fig. S1 and Table S2, ESI†). Therefore, ROA-CPL spectra are much more informative and allow for detecting differences between those species, while Raman-TL spectra reveal identical profiles, perfectly matching the spectrum of a single Eu(III)-probe.
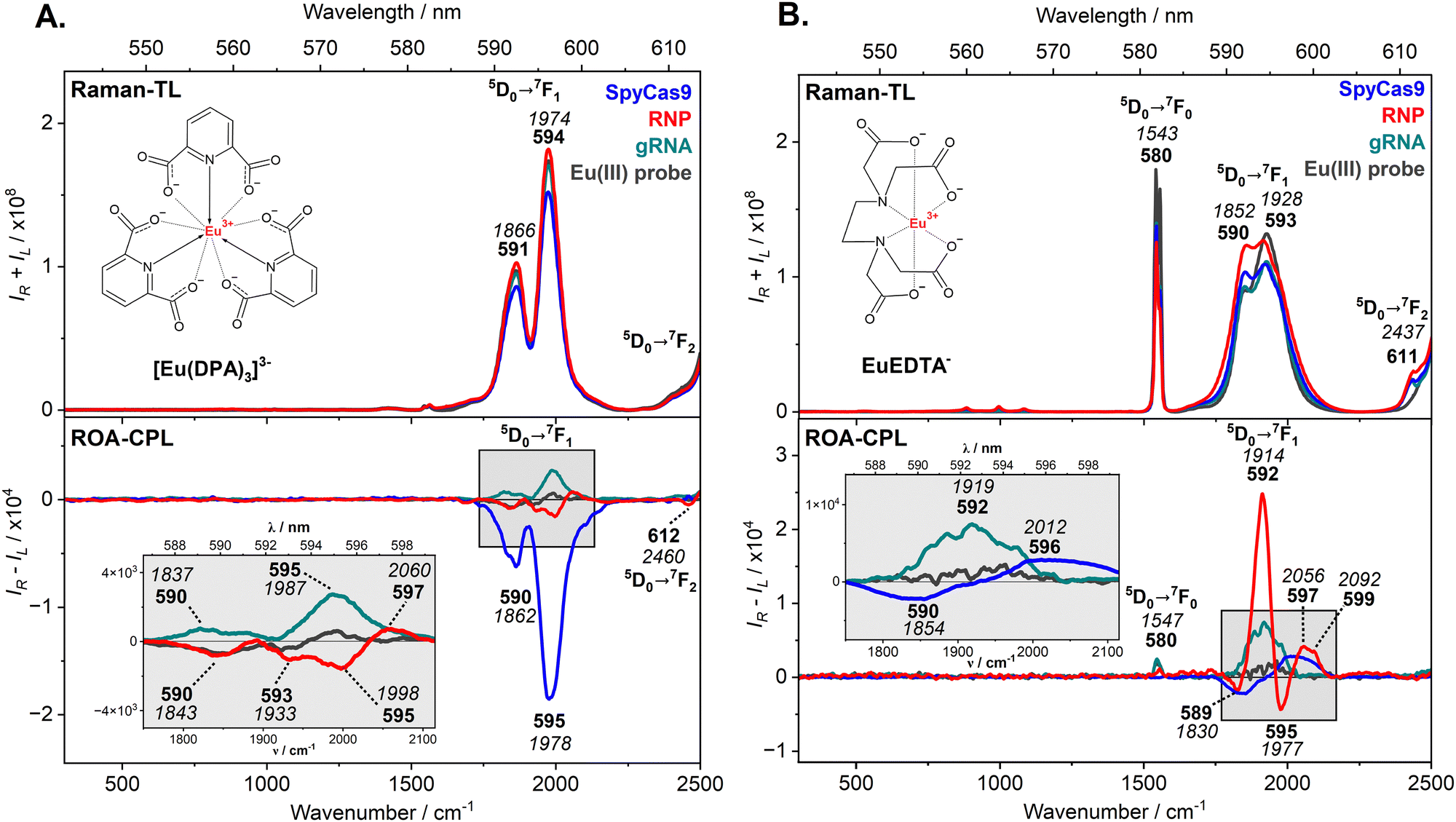 | ||
| Fig. 3 Raman-TL and ROA-CPL spectra of SpyCas9 (blue), RNP (red), and gRNA (green), combined with [Eu(DPA)3]3− (panel A) and EuEDTA− (panel B), and of the free Eu(III)-probe (dark grey). Luminescent bands were expressed in nm (bold) and cm−1 (italic) scales. | ||
As presented in Fig. 3A, the most intense TL and CPL bands are associated with the 5D0 → 7F1 transition (590–595 nm). The 5D0 → 7F0 transition (∼580 nm) is barely seen, because it is forbidden by the selection rules for high-symmetry complexes such as [Eu(DPA)3]3− (D3 symmetry).27b Next, the 5D0 → 7F2 transition (over 613 nm) is cut off at the very beginning, due to the limited spectral range. The CPL patterns induced by the SpyCas9 protein, gRNA, and their active RNP form differ in terms of signs and the number of bands, due to different chirality and interaction sites of the measured species, which differently disturb the enantiomeric equilibrium (Λ ↔ Δ) of the [Eu(DPA)3]3− probe (Pfeifer effect).20b,29 Their individual CPL response can also be quantified by the circular intensity difference (CID values, Table S5, ESI†), i.e. the ratio of ROA-CPL (IR − IL) to Raman-TL (IR + IL) spectra. The negative CPL profile of the SpyCas9 nuclease suggests that one of the [Eu(DPA)3]3− enantiomers predominates in the solution. Adequately, CID values for these bands are relatively high (∼10−4) compared to those calculated for gRNA and RNP (∼10−6–10−5). The [Eu(DPA)3]3− probe is more efficient in detecting the SpyCas9 structure without destroying its dedicated nucleolytic activity (Table S2 and Fig. S1, ESI†). Low-intensity CPL bands for gRNA and RNP are just above the ROA detection limit, thus, it is recommended to use another Eu(III)-probe to easily distinguish them in solution.
The EuEDTA− complex possesses three vacancies in the inner coordination sphere,30 so it behaves differently under the influence of chiral agents, which not only disturb its enantiomeric equilibrium (Λ ↔ Δ) but can also bind to its central Eu(III) ion. Fig. 3B shows the Raman-TL and ROA-CPL spectra of SpyCas9, gRNA, and RNP probed with the EuEDTA−, where the luminescent signals of the sensor are dominant. Again, all TL profiles are quite similar and only the ROA spectra provide very distinctive CPL bands in the range of 5D0 → 7F1 transition (589–599 nm). The most intense CPL response was registered for the RNP form suggesting the strongest interaction to the EuEDTA−. Its bisignate pattern of multiple CPL bands is characterized by high CID ratios (∼10−4, Table S5, ESI†), up to 100 times higher in comparison to the previous [Eu(DPA)3]3− probing. The EuEDTA− complex sensitively recognizes the RNP complex at very low concentrations (6.1 μM), maintaining the programmed catalytic activity (Table S2 and Fig. S1, ESI†). SpyCas9 and gRNA treated separately with EuEDTA− do not generate as strong CPL response as their active combination (CID values ∼10−6–10−5). However, their CPL patterns look different, the −/+ couplet for SpyCas9 and the single positive band for gRNA are observed.
In summary, we employed ROA combined with Eu(III)-CPL probing as an effective approach to identify structural rearrangements in the SpyCas9 geometry upon gRNA binding in solution. Through an advanced activity assay, we confirmed the harmlessness of ROA and the neutral nature of Eu(III)-based probes regarding the original properties of the CRISPR-related biomolecules. The most informative ROA region was the amide III range, which revealed changes in the conformation of the α-helical backbone, an increase in PPII content, and the orientation of Arg side chains. The obtained CPL patterns were distinctive, enabling clear differentiation between inactive and active nuclease forms. [Eu(DPA)3]3− exhibited a stronger affinity for SpyCas9, while EuEDTA− was more suitable for RNP detection. We found EuEDTA− to be a promising sensor for tracking catalytically active RNPs at low concentrations (<6.1 μM) without the need for complex genetic assays.
Funding was provided by the National Science Centre (NCN) under grants no. 2021/40/C/ST4/00190 to M.H. (SpyCas9 purchase, gRNA and RNPs synthesis, ROA/CPL measurements and analysis, nuclease activity assay) and 2019/35/B/ST4/04161 to G.Z. (providing Eu(III)-based probes, ROA/CPL analysis).
Data availability
All data are available in the main text or the ESI.† Data are also available upon request to the corresponding author.Conflicts of interest
The authors declare no competing interests.Notes and references
- M. Jinek, K. Chylinski, I. Fonfara, M. Hauer, J. A. Doudna and E. Charpentier, Science, 2012, 337, 816–821 CrossRef CAS PubMed.
- C. Zhang, Z. Huang, Z. Li, J. Hu, R. Liu and Y. Lv, TrAC, Trends Anal. Chem., 2024, 170, 117431 CrossRef CAS.
- M. Klimek-Chodacka, T. Oleszkiewicz, L. G. Lowder, Y. Qi and R. Baranski, Plant Cell Rep., 2018, 37, 575–586 CrossRef CAS PubMed.
- M. Klimek-Chodacka, M. Gieniec and R. Baranski, Int. J. Mol. Sci., 2021, 22, 10740 CrossRef CAS PubMed.
- (a) T. Fujisawa, Encyclopedia of Analytical Chemistry: Applications, Theory and Instrumentation, John Wiley & Sons, 2020, 1–19 Search PubMed; (b) F. Zhu, N. W. Isaacs, L. Hecht and L. D. Barron, Structure, 2005, 13, pp. 1409–1419 CrossRef PubMed.
- (a) M. L. Pusey, Z.-J. Liu, W. Tempel, J. Praissman, D. Lin, B. C. Wang, J. A. Gavira and J. D. Ng, Prog. Biophys. Mol. Biol., 2005, 88, 359–386 CrossRef CAS PubMed; (b) K. R. Acharya and M. D. Lloyd, Trends Pharmacol. Sci., 2005, 26, 10–14 CrossRef CAS PubMed.
- (a) M. Buděšínský, P. Daněček, L. Bednárová, J. Kapitán, V. Baumruk and P. Bouř, J. Phys. Chem. A, 2008, 112, 8633–8640 CrossRef PubMed; (b) S. C. Sue, C. F. Chang, Y. T. Huang, C. Y. Chou and T. Huang, Phys. A: Stat. Mech. Appl., 2005, 350, 12–27 CrossRef CAS.
- L. D. Barron, L. Hecht, I. H. McColl and E. W. Blanch, Mol. Phys., 2004, 102, 731–744 CrossRef CAS.
- (a) C. Mensch, A. Konijnenberg, R. Van Elzen, A.-M. Lambeir, F. Sobott and C. Johannessen, J. Raman Spectrosc., 2017, 48, 910–918 CrossRef CAS; (b) J. Kessler, J. Kapitán and P. Bouř, J. Phys. Chem. Lett., 2015, 6, 3314–3319 CrossRef CAS; (c) A. N. L. Batista, J. M. Batista, V. S. Bolzani, M. Furlan and E. W. Blanch, Phys. Chem. Chem. Phys., 2013, 15, 20147–20152 RSC; (d) L. D. Barron, L. Hecht, E. W. Blanch and A. F. Bell, Prog. Biophys. Mol. Biol., 2000, 73, 1–49 CrossRef CAS PubMed.
- (a) S. Yamamoto and F. Kimura, Phys. Chem. Chem. Phys., 2022, 24, 3191–3199 RSC; (b) C. Mensch, P. Bultinck and C. Johannessen, Phys. Chem. Chem. Phys., 2019, 21, 1988–2005 RSC; (c) C. Mensch, L. D. Barron and C. Johannessen, Phys. Chem. Chem. Phys., 2016, 18, 31757–31768 RSC.
- (a) C. Mensch and C. Johannessen, ChemPhysChem, 2019, 20, 42–54 CrossRef CAS PubMed; (b) C. Mensch and C. Johannessen, ChemPhysChem, 2018, 19, 3134–3143 CrossRef CAS PubMed.
- (a) C. Mensch, P. Bultinck and C. Johannessen, ACS Omega, 2018, 3, 12944–12955 CrossRef CAS PubMed; (b) C. Johannessen, J. Kapitán, H. Collet, A. Commeyras, L. Hecht and L. D. Barron, Biomacromolecules, 2009, 10, 1662–1664 CrossRef CAS PubMed; (c) C. D. Syme, E. W. Blanch, C. Holt, R. Jakes, M. Goedert, L. Hecht and L. D. Barron, Eur. J. Biochem., 2002, 269, 148–156 CrossRef CAS PubMed.
- L. D. Barron, Z. Q. Wen and L. Hecht, J. Am. Chem. Soc., 1992, 114, 784–786 CrossRef CAS.
- L. Ashton, P. D. A. Pudney, E. W. Blanch and G. E. Yakubov, Adv. Colloid Interface Sci., 2013, 199–200, 66–77 CrossRef CAS PubMed.
- J. Matsuo, T. Kikukawa, T. Fujisawa, W. D. Hoff and M. Unno, J. Phys. Chem. Lett., 2020, 11, 8579–8584 CrossRef CAS PubMed.
- O. Vrtělka, K. Králová, M. Fousková, L. Habartová, P. Hříbek, P. Urbánek and V. Setnička, Analyst, 2023, 148, 2793–2800 RSC.
- (a) A. Kolodziejczyk, L. A. Nafie, A. Wajda and A. Kaczor, Chem. Commun., 2023, 59, 10793–10796 RSC; (b) A. Kurochka, J. Průša, J. Kessler, J. Kapitán and P. Bouř, Phys. Chem. Chem. Phys., 2021, 23, 16635–16645 RSC.
- T. Wu, Phys. Chem. Chem. Phys., 2022, 24, 15672–15686 RSC.
- A. Domagała, S. Buda, M. Baranska and G. Zając, Spectrochim. Acta A Mol. Biomol. Spectrosc., 2025, 324, 124995 CrossRef PubMed.
- (a) M. Krupová, J. Kapitán and P. Bouř, ACS Omega, 2019, 4, 1265–1271 CrossRef PubMed; (b) T. Wu, J. Kessler and P. Bouř, Phys. Chem. Chem. Phys., 2016, 18, 23803–23811 RSC.
- T. Wu, P. Bouř and V. Andrushchenko, Sci. Rep., 2019, 9, 1068 CrossRef PubMed.
- (a) M. Jinek, F. Jiang, D. W. Taylor, S. H. Sternberg, E. Kaya, E. Ma, C. Anders, M. Hauer, K. Zhou, S. Lin, M. Kaplan, A. T. Iavarone, E. Charpentier, E. Nogales and J. A. Doudna, Science, 2014, 343, 1247997 CrossRef PubMed; (b) F. Jiang, K. Zhou, L. Ma, S. Gressel and J. A. Doudna, Science, 2015, 348, 1477–1481 CrossRef CAS PubMed; (c) H. Nishimasu, F. A. Ran, P. D. Hsu, S. Konermann, S. I. Shehata, N. Dohmae, R. Ishitani, F. Zhang and O. Nureki, Cell, 2014, 156, 935–949 CrossRef CAS PubMed.
- M. Halat, M. Klimek-Chodacka, J. Orleanska, M. Baranska and R. Baranski, Int. J. Mol. Sci., 2021, 22, 1–14 Search PubMed.
- (a) A. A. Adzhubei, M. J. E. Sternberg and A. A. Makarov, J. Mol. Biol., 2013, 425, 2100–2132 CrossRef CAS PubMed; (b) P. Kumar and M. Bansal, J. Struct. Biol., 2016, 196, 414–425 CrossRef CAS PubMed.
- (a) F. Jiang and J. A. Doudna, Annu. Rev. Biophys., 2017, 46, 505–529 CrossRef CAS PubMed; (b) G. Panda and A. Ray, Comput. Struct. Biotechnol. J., 2022, 20, 4172–4184 CrossRef CAS PubMed.
- M. Bratovič, I. Fonfara, K. Chylinski, E. J. C. Gálvez, T. J. Sullivan, S. Boerno, B. Timmermann, M. Boettcher and E. Charpentier, Nat. Chem. Biol., 2020, 16, 587–595 CrossRef PubMed.
- (a) A. T. Frawley, R. Pal and D. Parker, Chem. Commun., 2016, 52, 13349–13352 RSC; (b) K. Binnemans, Coord. Chem. Rev., 2015, 295, 1–45 CrossRef CAS.
- T. Wu, P. Bouř, T. Fujisawa and M. Unno, Adv. Sci., 2024, 11, 2305521 CrossRef CAS PubMed.
- (a) P. Gawryszewska, J. Legendziewicz, Z. Ciunik, N. Esfandiari, G. Muller, C. Piguet, M. Cantuel and J. P. Riehl, Chirality, 2006, 18, 406–412 CrossRef CAS PubMed; (b) S. Wu, G. L. Hilmes and J. P. Riehl, J. Phys. Chem., 1989, 93, 2307–2310 CrossRef CAS.
- (a) M. S. Thomsen, P. R. Nawrocki, N. Kofod and T. J. Sørensen, Eur. J. Inorg. Chem., 2022, e202200334 CrossRef CAS; (b) T. Wu, J. Průša, J. Kessler, M. Dračínský, J. Valenta and P. Bouř, Anal. Chem., 2016, 88(8), 8878–8885 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5cc00971e |
| This journal is © The Royal Society of Chemistry 2025 |