
The recent advancements in protein nanoparticles for immunotherapy
Miaomiao
Xu
 a,
Siyuan
Wei
a,
Lifan
Duan
a,
Yifan
Ji
b,
Xiaofan
Han
b,
Qipeng
Sun
b and
Lixing
Weng
*a
a,
Siyuan
Wei
a,
Lifan
Duan
a,
Yifan
Ji
b,
Xiaofan
Han
b,
Qipeng
Sun
b and
Lixing
Weng
*a
aState Key Laboratory of Organic Electronics and Information Displays & Jiangsu Key Laboratory for Biosensors, Institute of Advanced Materials (IAM), Nanjing University of Posts and Telecommunications, Nanjing 210023, China. E-mail: lxweng@njupt.edu.cn
bPortland Institute, Nanjing University of Posts and Telecommunications, Nanjing 210023, China
First published on 30th May 2024
Abstract
In recent years, the advancement of nanoparticle-based immunotherapy has introduced an innovative strategy for combatting diseases. Compared with other types of nanoparticles, protein nanoparticles have obtained substantial attention owing to their remarkable biocompatibility, biodegradability, ease of modification, and finely designed spatial structures. Nature provides several protein nanoparticle platforms, including viral capsids, ferritin, and albumin, which hold significant potential for disease treatment. These naturally occurring protein nanoparticles not only serve as effective drug delivery platforms but also augment antigen delivery and targeting capabilities through techniques like genetic modification and covalent conjugation. Motivated by nature's originality and driven by progress in computational methodologies, scientists have crafted numerous protein nanoparticles with intricate assembly structures, showing significant potential in the development of multivalent vaccines. Consequently, both naturally occurring and de novo designed protein nanoparticles are anticipated to enhance the effectiveness of immunotherapy. This review consolidates the advancements in protein nanoparticles for immunotherapy across diseases including cancer and other diseases like influenza, pneumonia, and hepatitis.
1. Introduction
Immunotherapy constitutes a therapeutic strategy meticulously formulated to proficiently stimulate the organism's immune system, effectively addressing a range of diseases, notably cancer, influenza, and pneumonia.1–5 For instance, cancer immunotherapy has garnered prominence in recent decades by rectifying deficiencies inherent in conventional cancer treatment methods, such as pronounced cytotoxicity, limited control over metastatic cancer cells, and elevated recurrence rates.6–10 However, the current efficacy of immunotherapy for diverse diseases is hindered by factors such as insufficient infiltration of immune-suppressive cells, inadequate targetability, and inefficient antigen presentation.11,12The emergence of nanoparticles (NPs), facilitated by advancements in nanotechnology, introduces a novel approach for augmenting the effectiveness of immunotherapy.11–15 Their distinctive size, spanning 1–1000 nm, enhances the permeability and retention (EPR) effects, facilitating capture by antigen-presenting cells (APCs).13,16 Furthermore, the unique modifiability of NPs allows for decoration with targeted molecules, thereby reducing off-target effects in immunotherapy.17 Moreover, due to their increased capacity for loading adjuvants and antigens, NPs can serve as efficacious platforms in immunotherapy to enhance its overall efficiency.18,19
In immunotherapy, NPs are typically categorized based on their building blocks, including inorganic compounds, lipids, polymers, and proteins.11,13,17,20 NPs constructed from metal and inorganic materials have been extensively employed to enhance the pharmaceutical properties of anti-cancer drugs. Certain NPs, such as mesoporous silica and gold nanospheres, have played a role in immunogenic cell death (ICD)-mediated cancer immunotherapy.21,22 However, concerns persist regarding the biological toxicity and limited biodegradability of these inorganic NPs.23,24 Lipids, derived from hydrogenized and detoxified derivatives of lipopolysaccharide (LPS), demonstrate high biosafety and have been established as effective adjuvants for activating immunity in cancer immunotherapy.25,26 Additionally, owing to their favorable biocompatibility and similarity to biofilms, lipid NPs serve as efficient carriers, demonstrating competence as a delivery system in immunotherapy.27 However, the hydrophilic nature of lipid NPs limits their ability to stably release drugs in the body, resulting in a short half-life of drugs.28 Polymer-based NPs, such as poly(lactic acid) (PLA) and poly(lactic-co-glycolic acid) (PLGA), are approved by the U.S. Food and Drug Administration (FDA) and hold promise for nanovaccine formulations in cancer treatment, antiviral therapy, and antibacterial drugs.18,29 Nevertheless, challenges, including low drug-loading rates, issues with drug release, and special processing requirements, must be addressed for extensive applications.30,31
Proteins, intricate in their sophisticated structures and versatile functions, play integral roles in all fundamental life processes.32 Compared with the basic monomeric form of protein, protein nanoparticles (PNPs) exhibit superior performance in functional control, stability, allosteric regulation, and higher-order complexity.33–35 Specifically, these qualities are evident in several aspects: (1) functional control: PNPs are homomerized of a single kind of protein or heteromerized of several kinds of different proteins. Thus, PNPs can serve as the integrated platform for multiple protein functions.23,24,36 By chemical modification or genetic fusion on different protein subunits, PNPs can realize the carrying of drugs and antigens while targeting tumor cells or APCs.24,37 Besides, by intervening in the interaction between subunits of PNPs, controlled release of cargo can be achieved.34 (2) Stability: stability encompasses two facets. Firstly, the inherent stability of PNPs themselves, as many proteins are naturally stable or derived from the body; secondly, the stability of functionalized PNPs in carrying drugs and antigens. Functionalized PNPs can deliver antigens in a reactive array, addressing issues like poor antigen stability and insufficient immune stimulation commonly seen in traditional vaccines.23,37–39 (3) Allosteric regulation: allosteric regulation occurs through the specific binding of small molecular substrates or ligands to proteins, thereby regulating their biological activity and functions. Hemoglobin, an α2β2 tetrameric assembly which contains four haem prosthetic groups, efficiently transports oxygen from the lungs into tissues via allosteric regulation.35 Besides, by assembly of allosteric protein into PNPs, drugs can be released controllably under allosteric signals.40 (4) Higher-order complexity: biomolecules exhibit remarkable complexity and diversity, rendering them unique building blocks for generating precisely assembled components. Leveraging complex biomolecules to construct more intricate structures is essential for achieving multifunctionality. Among biomolecules, proteins stand out due to their inherent structural complexity, which is both precise and controllable. Various structures of PNPs have demonstrated high-order complexity, including tubes, rings, knots, and cages, laying the foundation for expanding the applications of PNPs.35,41 Besides, the intrinsic high biocompatibility and biodegradability of PNPs make them extensively applicable in immunotherapy.24,36 Happily, the FDA has already sanctioned two types of PNPs. The first, Abraxane, is an albumin-bound paclitaxel NP utilized for treating breast and pancreatic cancer. The second, Ontak, is a Denileukin diftitox NP, an engineered protein combining IL-2 and diphtheria toxin, designed for treating cutaneous T-cell lymphoma.42
In cancer immunotherapy, PNPs mainly play two roles, as platforms for carrying antigens, immune adjuvant, or drugs/photosensitizers/photothermal agents, or themselves directly as adjuvants for immune enhancement.24,36,43,44 As for the platforms, here we discuss them point-by-point according to the different cargoes they carry. (1) antigens: PNPs can carry cancer-related antigens through chemical conjugation or gene fusion and act as cancer vaccines. PNPs with specific spatial structures are easily captured by APCs to present cancer-related antigens, thereby inducing specific T-cell immune responses.45–47 (2) Immune adjuvants: PNPs can carry adjuvants like CpG through covalent interaction and other methods, further activating APCs by means of promoting the maturation of dendritic cells (DCs) and the polarization of tumor-associated macrophage (TAM) into the tumor-suppressing M1 phenotype, also releasing inflammatory cytokines to reverse the immunosuppressive tumor microenvironment.48–50 (3) Drugs/photosensitizers/photothermal agents: due to the hydrophobic interactions of proteins like albumin and the pH-responsive characteristic of ferritin, various cargoes can be loaded into PNPs. For example, loading with chemotherapeutic drugs such as doxorubicin (DOX) and paclitaxel can induce ICD and enhance anti-tumor immune responses, or loading with photosensitizers/photothermal agents for photodynamic therapy (PDT)/photothermal therapy (PTT) also can induce ICD or generate reactive oxygen species (ROS) capable of directly killing cancer cells.51–57 Achieving these processes often requires PNPs to accumulate in cancer tissues via the EPR effect or to acquire targeting abilities through molecular modifications to target cancer cells. In addition to their role as carriers, some PNPs, especially virus-like particles (VLPs), can directly act as immune-activating adjuvants.44 Even without carrying other antigens or adjuvants, some VLPs such as cowpea mosaic virus (CPMV) exhibit excellent anti-tumor efficacy.58
As for other diseases such as influenza, pneumonia, and hepatitis, PNPs are also usually equipped with related antigens and designed as vaccines. Similar to the design of cancer vaccines, PNPs present disease antigens through strategies such as gene fusion or chemical conjugation, forming self-assembled platforms that efficiently deliver antigens to APCs, activating helper T cells, and further maturing B cells into IgG-producing plasma cells.59 Additionally, the repetitive array of antigens presented by PNPs can effectively activate multiple B-cell receptors, thereby activating B cells.60 Therefore, as a safe, functionalized, and multivalent antigen-presenting vaccine platform, PNPs induce stronger and more sustained neutralizing antibody titers, providing robust protection.61–63
Naturally occurring protein nanoparticles (NPNPs), such as the ferritin nanocage and polyhedral virus capsid, represent promising candidates for immunotherapy.36,64,65 Drawing inspiration from nature, researchers have adeptly assembled natural protein building blocks into sophisticated NPs or modified the surfaces of certain NPNPs while retaining their three-dimensional structures. These lines of research aim to bolster the effectiveness of immunotherapy.17,23,43
In addition to naturally evolved protein building blocks, computational methodologies have significantly contributed to the progress in developing de novo designed protein building blocks. These novel designs enable exploration of the complete sequence space of amino acids.66,67 In comparison with their naturally evolved counterparts, de novo designed proteins generally exhibit enhanced stability due to their adherence to the physical principle of the lowest energy state.64,68 Furthermore, scientists have employed a bottom-up strategy to assemble these de novo designed protein building blocks into highly ordered NPs, namely de novo designed protein nanoparticles (DPNPs), exploring their potential applications in immunotherapy.32,37
Thus, in this review, we first outline the recent advance of NPNPs in immunotherapy for cancer, and followed by a discussion of the applications of NPNPs and DPNPs in immunotherapy for other diseases (Fig. 1). Finally, we close with a discussion of the challenges, and future directions of PNPs in immunotherapy.
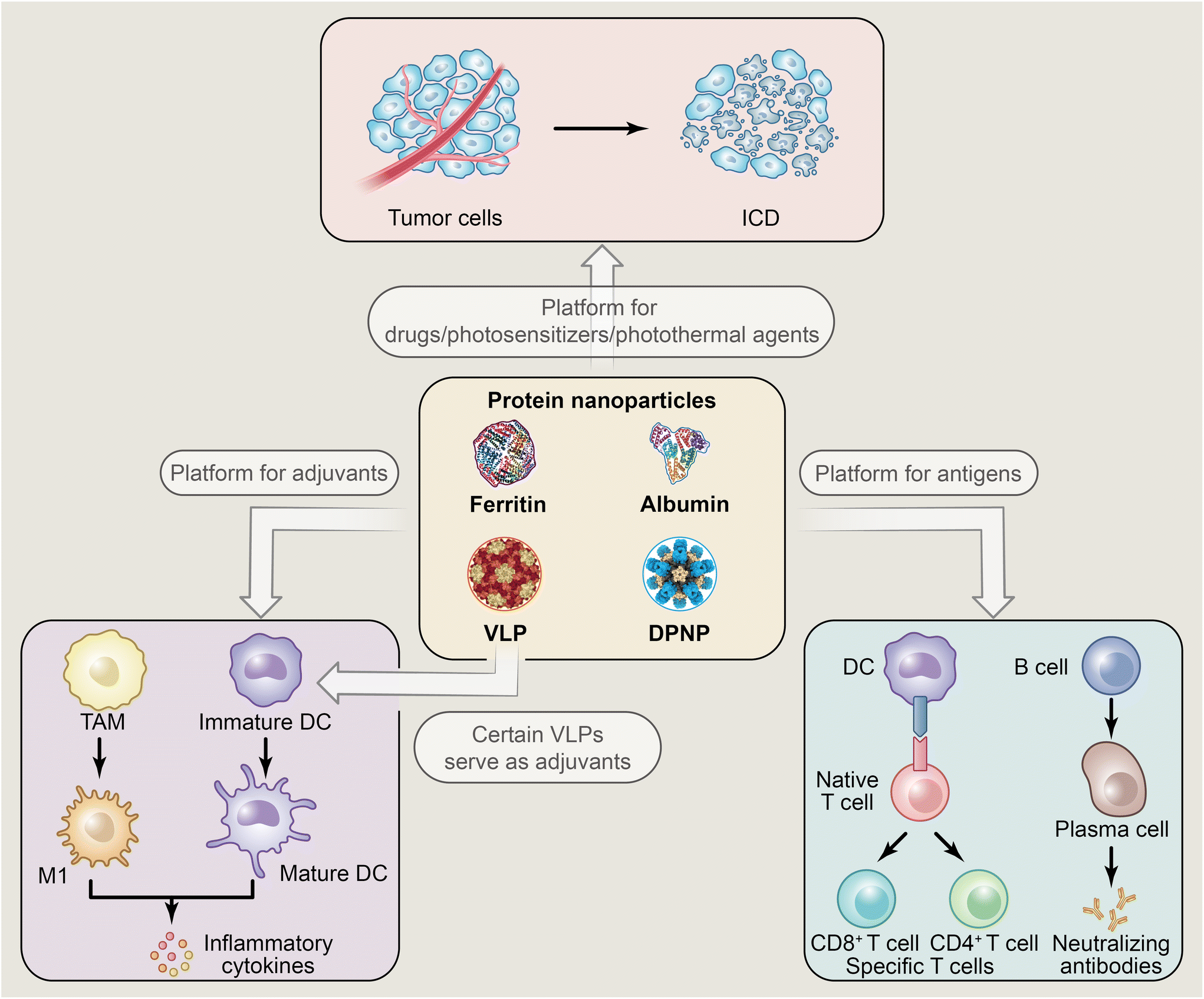 | ||
| Fig. 1 The applications of PNPs in immunotherapy. | ||
2. Immunotherapy for cancer
Cancer poses a formidable public health challenge, resulting in nearly 96 million deaths annually and emerging as the second leading cause of global mortality.69 The American Cancer Society projects approximately 2 million new cancer cases and 609![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 820 cancer-related deaths in the United States by 2023.70 In cancer immunotherapy, the typical approach to achieving an anti-tumor effect involves activating APCs to present antigens and stimulating anti-tumor-specific T lymphocytes to directly target and eliminate tumors.71 However, cancer immunotherapy faces challenges in immune tolerance and other adverse reactions during clinical practice.72 For instance, checkpoint blockade immunotherapy has demonstrated remarkable anti-tumor effects on solid tumors, but there is a need for refinement in its targeting strategy.73,74 Adoptive cell transfer immunotherapy can significantly boost the induction of tumor-specific T cells, albeit with the potential for severe cytokine storms.75 Besides, while immune adjuvants are commonly employed to enhance adaptive immune responses, they encounter challenges such as susceptibility to clearance by the body and limited immunogenicity.14,76 PNPs offer a promising alternative in immunotherapy, capable of augmenting anti-tumor efficacy by incorporating immune adjuvants and integrating multiple therapeutic modalities. Notably, the current understanding of DPNP applications in cancer immunotherapy remains unclear based on the retrieved research articles. Hence, the following discussion primarily highlights recent advancements in NPNPs within the context of cancer immunotherapy.
820 cancer-related deaths in the United States by 2023.70 In cancer immunotherapy, the typical approach to achieving an anti-tumor effect involves activating APCs to present antigens and stimulating anti-tumor-specific T lymphocytes to directly target and eliminate tumors.71 However, cancer immunotherapy faces challenges in immune tolerance and other adverse reactions during clinical practice.72 For instance, checkpoint blockade immunotherapy has demonstrated remarkable anti-tumor effects on solid tumors, but there is a need for refinement in its targeting strategy.73,74 Adoptive cell transfer immunotherapy can significantly boost the induction of tumor-specific T cells, albeit with the potential for severe cytokine storms.75 Besides, while immune adjuvants are commonly employed to enhance adaptive immune responses, they encounter challenges such as susceptibility to clearance by the body and limited immunogenicity.14,76 PNPs offer a promising alternative in immunotherapy, capable of augmenting anti-tumor efficacy by incorporating immune adjuvants and integrating multiple therapeutic modalities. Notably, the current understanding of DPNP applications in cancer immunotherapy remains unclear based on the retrieved research articles. Hence, the following discussion primarily highlights recent advancements in NPNPs within the context of cancer immunotherapy.
2.1 VLPs
VLPs are NPs ranging in size from tens to hundreds of nanometers, spontaneously assembled by viral capsid shell proteins.17 NPs containing antigens sourced from either viral or non-viral origins, exhibiting a size and shape similar to the virus, are also categorized as VLPs.77In cancer immunotherapy, like other PNPs, VLPs can serve as the platform to carry cargos. The surface of VLPs can be equipped with tumor antigens, peptides, and small molecules via surface modification. The inner cavity of VLPs can also carry chemotherapy drugs, proteins, peptides and nucleic acids.
It should be noted that more than half of global gene therapy clinical trials utilize VLPs to transport genetic material.78 This is evident in several ways: (1) many VLPs have the innate ability to encapsulate nucleic acids from the host cells they infect. For instance, RNA phage VLPs produced by E. coli recombination can naturally encapsulate bacterial-derived single-stranded RNAs (ssRNAs), which are recognized by TLRs.79 (2) Some plant viruses carry nucleic acids that can also be recognized by TLRs, providing them with immune stimulation potential. For instance, the nucleic acid carried by CPMV itself can be recognized by cytoplasmic TLR7, thereby inducing cytokine secretion and enhancing its anti-tumor immune function.80 (3) VLPs lacking their own genome can readily encapsulate therapeutic nucleic acids like CpG oligodeoxynucleotide for cancer immunotherapy.78,81
Apart from the delivery capability, the noteworthy reason behind the high regard for VLPs as a promising platform for cancer immunotherapy lies in their structural similarity to real viruses, and VLPs of diverse shapes and sizes create conditions conducive to efficient drainage to lymph nodes and the capture of APCs like DCs, thereby achieving immune activation.60,77,82
Moreover, some VLPs themselves can also serve as adjuvants. Plant-derived VLPs are non-infectious, have low toxicity in animals whether carrying genetic information or not, and can stimulate the immune system. Animal-derived VLPs without genetic information possess advantages such as natural biocompatibility, high immunogenicity, low allergenicity, and high clinical efficacy while alleviating concerns regarding contagion.
These unique advantages of VLPs enable their participation in a wide array of combined cancer therapies, encompassing chemotherapy, radiation therapy and PDT.51,83,84 The VLPs introduced here encompass some plant/animal-derived viruses and oncolytic viruses (OVs) capable of specifically infecting cancer cells (Fig. 2). We also summarize the applications of VLPs in cancer immunotherapy, as shown in Table 1.
 | ||
| Fig. 2 The utilization of certain VLPs in the context of cancer immunotherapy. (a) Anti-tumor effects of CPMV, CPSMV, and TRSV as cancer immunotherapy vaccines [reprinted with permission from ref. 85; Copyright (2022) American Chemical Society]. (b) Synthesis of PVX–DOX and tumor growth curves of different groups [reprinted with permission from ref. 51; Copyright (2017) American Chemical Society]. (c) Synthesis of CPMV-1V209, CCMV-1V209, and Qβ-1V209 particles [reprinted with permission from ref. 48; Copyright (2023) American Chemical Society]. (d) Schematic diagram of the self-assembly mechanism of p22 VLP vaccines and tumor growth curves from the prophylactic tumor inhibition model [reprinted with permission from ref. 91; Copyright (2021) Elsevier]. (e) Schematic representation of the self-assembly process of hybrid fluorescent VLPs and images depicting B16-OVA tumors and lung metastases isolated from immunized mice in various groups at the end of the treatment period [reprinted with permission from ref. 45; Copyright (2020) American Chemical Society]. (f) FH VLP design strategy and tumor growth curves from two mouse tumor models [reprinted with permission from ref. 94; Copyright (2022) American Chemical Society]. (g) Schematic of biometric PNPs stimulating immature dendritic cell (iDC) to mature dendritic cell (mDC) through loading antigen peptides and CpG, and the bar chart of enhanced activation of BMDCs by CpG encapsulated in E2 PNPs (termed as CpG-E2) [reprinted with permission from ref. 97; Copyright (2013) American Chemical Society]. | ||
| Sources | Types | Carrying virus genome or not | Immune strategies | Therapeutic effects (in the mouse model) | Ref. |
|---|---|---|---|---|---|
| Plant viruses | CPMV | Yes | As an adjuvant in activating TLR2, TLR4, TRL7, TLR8 | Suppressing B16F10 dermal melanoma/ID8-Defb29/VEGF-a ovarian cancer/CT-26 colon cancer | 58 and 85 |
| CPMV | Yes | Using RNA-laden, but non-infectious CPMV as an adjuvant in activating TLRs | Suppressing B16F10 melanoma | 86 | |
| CPMV | Yes | As a platform in delivering TLR7/TLR3 agonist | Suppressing B16F10 melanoma/CT26 colon cancer | 48 | |
| CCMV | Yes | As a platform in delivering TLR7 agonist | Suppressing B16F10 melanoma/CT26 colon cancer | 48 | |
| CCMV | No | As a platform in delivering TLR9 agonist | Suppressing B16F10 melanoma/CT26 colon cancer | 186 | |
| CPSMV | Yes | As an adjuvant in activating TLR2, TLR4 | Suppressing B16F10 melanoma | 85 | |
| TRSV | Yes | As an adjuvant in activating TLR2, TLR4 | Suppressing B16F10 melanoma | 85 | |
| PVX | Yes | As a platform for delivering DOX | Suppressing B16F10 melanoma | 51 | |
| TMV | Yes | As an adjuvant in polarizing macrophage to M1 phenotype via TLR4 | Suppressing 4T1 tumors | 87 | |
| TMV | Yes | As a platform for delivering tumor-associated carbohydrate antigens | Inducing strong humoral immune responses | 187 | |
| Bacteriophages | Qβ | No | As a platform for delivering TLR7 agonist | Suppressing B16F10 melanoma/CT26 colon cancer | 48 |
| Qβ | No | As a platform for delivering tumor antigens | Suppressing the metastasis of melanoma | 188 | |
| Qβ | No | As a platform for delivering TLR9 agonist | Inducing durable tumor regression at injected and distant tumors and significantly prolonged survival | 189 | |
| P22 | No | As a platform for delivering OVA peptides | Suppressing E.G7-OVA lymphoma | 91 | |
| MS2 | No | As a platform for displaying the cystine-glutamate antiporter protein xCT to generate high titer antibodies that inhibited the function of xCT expressing breast cancer stem cells | Suppressing 4T1 tumors | 190 | |
| Animal viruses | HBV | No | As a platform for delivering two antigens | Suppressing B16-OVA melanoma and lung metastatic | 45 |
| HBV | No | Flagellin/HBVc hybrid VLP acting as a platform for antigen delivery | Suppressing B16-OVA melanoma E.G7-OVA lymphoma | 94 | |
| Human papillomavirus (HPV) | No | As a platform for delivering siRNA oligonucleotides (for Cd274 knockdown) cooperating with immune checkpoint blockade therapy | Suppressing 4T1 tumor and improving immune checkpoint blockade therapy immunotherapy | 191 | |
| Norovirus | No | As a platform for delivering antigen | Suppressing TC-1 tumors and B16F10 melanoma | 46 | |
| Rabbit haemorrhagic disease virus (RHDV) | No | Chimaeric RHDV VLP containing epitopes derived from colorectal cancer tumour-associated antigens topoisomerase IIα and surviving as an immunotherapeutic vaccine | Suppressing MC38-OVA tumors and improving overall survival | 192 | |
| RHDV | No | As a platform modified to contain the universal helper T-cell epitope PADRE and decorated with an MHC I-restricted peptide from the HPV16 E6 | Suppressing TC-1 tumors and prolonging survival for HPV tumor-bearing mice | 193 |
Numerous studies have explored the application of plant virus-derived VLPs in cancer immunotherapy. CPMV, an icosahedral VLP non-infectious to mammals, has demonstrated superior anti-tumor effects as an immune stimulator compared with various other VLPs. In a previous investigation, Beiss et al. established CPMV's efficacy as an in situ vaccine for cancer immunotherapy through in vivo experiments, surpassing cowpea chlorotic mottle virus (CCMV), physalis mosaic virus (PhMV), sesbania mosaic virus (SeMV), bacteriophage Qβ VLPs, and hepatitis B virus capsids (HBVc).58 Subsequently, Beiss et al. extended the comparison to other plant-generated viruses, such as cowpea severe mosaic virus (CPSMV) and tobacco ring spot virus (TRSV), in the context of cancer immunotherapy. Results from a mouse dermal melanoma model revealed that CPMV, as an in situ vaccine, exhibited significantly superior anti-tumor efficacy compared with CPSMV and TRSV (Fig. 2a).85 CPMV could significantly stimulate the production of pro-inflammatory cytokines in lymphocytes within the draining lymph nodes (DLN) of tumor-bearing mice. Besides, it has been confirmed that the action of three plant viruses was reliant on the adapter molecule MyD88 from the experimental results of the mouse knockout (KO) model. Interestingly, in comparison with the other two VLPs, the absence of TLR2 or TLR4 signaling did not diminish the ability of the CPMV in situ vaccine to induce interferon-β (IFN-β). Additionally, the mouse KO model revealed that TLR7 signaling played a pivotal role in mediating the effectiveness of CPMV-induced IFN-β production, highlighting its significance in generating differential responses. Moreover, CPMV induced more robust and persistent pro-inflammatory cytokines, suggesting an immune memory response. Another study by Beiss et al. compared the immunogenicity and cancer immunotherapy efficacy of chemically inactivated CPMV containing nucleic acids with non-infectious VLPs of CPMV devoid of nucleic acids. In vivo experiment results indicated that chemically inactivated CPMV more effectively reduced tumor volume and increased the survival rate compared with empty CPMV.86 These findings underscore CPMV VLP's role as a potent immune stimulator in cancer immunotherapy.
Other NPs derived from plant viruses, such as tobacco mosaic virus (TMV) and potato virus X (PVX), have demonstrated potential as vaccines for cancer immunotherapy. TMV has been observed to activate MAPK and NF-κB pro-inflammatory signaling pathways via TLR4, leading to the polarization of macrophages into M1 phenotype and the production of inflammatory cytokines.87 Nkanga et al. utilized TMV to deliver a small-molecule TLR7 antigen for cancer photothermal immunotherapy.88 In an example of combined immunotherapy and chemotherapy, Lee et al. loaded the chemotherapy drug doxorubicin (DOX) into PVX (Fig. 2b).51 In a mouse melanoma model, PVX alone resulted in slower tumor growth. However, when PVX was combined with DOX, the ‘PVX + DOX’ treatment group exhibited significantly greater inhibition of tumor progression compared with the DOX-loaded ‘PVX-DOX’ group. Furthermore, the combination therapy group induced significantly higher levels of inflammatory cytokines IFN-γ, IL-1α, and IL-1β. These indicators clarify that, when combined with chemotherapy drugs, PVX enhances its individual therapeutic effect as an immune enhancer.
Plant-derived viruses can not only be directly used for immune activation, but also serve as carriers for drug delivery. To address the pharmacokinetic challenges associated with small-molecule TLR agonists in clinical settings, Jung et al. employed plant virus NPs, including CPMV, CCMV, and bacteriophage Qβ VLPs, as carriers for the TLR7 agonist 1 V209 using the method of chemical conjugation (Fig. 2c).48In vivo studies revealed that compared with free agonists or simple mixtures of agonists and VLPs, agonist chemically conjugated to VLPs elicited higher production of inflammatory cytokines, restrained mouse tumor growth, and prolonged mouse survival.
Apart from viruses derived from plant sources, the protein capsid of bacteriophages infecting bacteria has been established as a self-assembly VLP antigen-loading platform. In addition to the bacteriophage Qβ VLPs mentioned above, the P22 VLP derived from the Salmonella typhimurium bacteriophage P22 is considered a versatile VLP platform owing to its outstanding stability, modifiability, and facile heterologous expression.89 Featuring a 58 nm icosahedral structure, the P22 VLP self-assembles from 420 47 kDa shell proteins and 100–300 internalized scaffold proteins.90 Patterson et al. accomplished the external covalent linking of protein domains in P22 VLPs using a sortase-mediated ligation strategy. The stability and structural characteristics of these VLPs were confirmed through Dynamic Light Scattering (DLS) and Small-Angle X-ray Scattering (SAXS) analyses.89 The results demonstrated the plasticity of bacteriophage P22 VLPs, showcasing their ability to achieve external bioconjugation by covalently attaching proteins to the C-terminus of the P22 coat protein. This underscores the robust nature of P22 VLPs as scaffolds for constructing diverse therapeutic NPs.
In a study on cancer immunotherapy using the P22 VLP platform, Li et al. devised two self-assembling P22 VLP vaccines, termed VLP-OVAB and VLP-OVAT, each presenting either the OVAB peptide (ISQAVHAAHAEINEAGR) or the OVAT peptide (SIINFEKL) at the C-terminus of the P22 CP protein. These peptides are well-known CD4+ and CD8+ T cell epitopes originating from OVA protein, which can be presented by DCs via macropinocytosis to execute different tumor-killing mechanisms (Fig. 2d).91In vivo, VLP-OVAB elicited higher levels of total IgG, IgG1, and IgG2a in mouse serum compared with free OVAB peptide. Additionally, an increase in immunization frequency correlated with elevated antibody titers. Notably, both VLP-OVAT and the unmodified VLP (termed VLP-WT) induced similar levels of maturation and inflammatory cytokines from bone marrow-derived dendritic cells (BMDCs) in vitro. This finding indicated that P22 VLPs themselves could enhance BMDCs’ immune activation, while the incorporation of OVAT peptide created a more conducive environment for BMDCs to activate CD8+ T cells. Furthermore, in vivo, both VLP-OVAT and VLP-WT migrated to lymph nodes through lymphatic vessels and accumulated there. However, in a tumor inhibition model using the immune adjuvant polyinosinic:polycytidylic acid, VLP-WT failed to impede tumor progression, whereas VLP-OVAB exhibited partial tumor growth inhibition. Although free OVAT peptide demonstrated preventive and therapeutic efficacy, its ability to induce CD8+ T cells was inferior to that of VLP-OVAT. These experimental findings underscore the significance of a NP platform with specific spatial structure and antigen selection in designing cancer immunotherapy vaccines.
Besides plant-derived viruses and bacteriophage VLPs, certain animal-infecting viruses are also employed as VLP-based platforms in cancer immunotherapy. The HBVc protein is a frequently utilized platform for VLP construction, easily obtained and purified via the E. coli expression system. It exhibits robust immunogenicity for B cells and CD8+ cytotoxic T-lymphocytes (CD8+ T).92,93
Typically, some VLP platforms either solely leverage the immune adjuvant properties of the VLP itself or carry only one specific antigen alone, often resulting in inadequate anti-tumor efficacy for inhibiting tumor metastasis. Cheng et al. developed a dual-antigen delivery system utilizing HBVc VLPs. Hybrid VLPs, referred to as dual-antigen-loaded VLPs, were created by incorporating the antigens OVA257–264 (SIINFEKL) or gp100 (KVPRNQDWL) peptides through conjugation with a glycine-rich linker.45 By developing a fluorescent probe that chemically conjugated the Cyanine5.5 (Cy5.5) fluorescence molecule and the corresponding quercetin BHQ-3 onto VLP monomers carrying distinct antigens, the reduction in fluorescence intensity could indicate the formation of Hybrid VLPs. This validated the assembly of two VLP monomers with different antigens into a single VLP (Fig. 2e). In vitro and in vivo experiments have demonstrated that hybrid VLPs could enhance the maturation of DCs (CD11C+CD80+CD86+) in BMDCs and lymph nodes. Following hybrid VLP stimulation, the maturation of dendritic cells in both in vitro and in vivo settings was approximately 30% higher than that observed in the control group, highlighting their superior ability to induce innate immune responses. Fig. 2e also illustrates the antitumor effects of hybrid VLPs in the mouse melanoma model and the lung metastatic tumor model. The immunotherapeutic effects and tumor metastasis inhibition abilities of the hybrid VLPs group were superior to those of VLPs loaded with a single antigen. This superiority arose from the ability of hybrid VLPs to enable DCs to present multiple antigens, thereby enhancing the immune response of CD8+ T cells. The promising therapeutic effects of dual-antigen-loaded VLPs underscore the benefits of presenting multi-antigen cancer immunotherapy vaccines.
In another study based on HBVc VLP, Zhao et al. developed a self-assembled hybrid platform termed FH VLP, which combined Flagellin with VLP based on HBVc, achieved by integrating the modified flagellin gene into the c/e1 loop of HBVc (Fig. 2f).94 To mitigate the risk of Flagellin-induced adverse reactions, they replaced the exposed D3 domain on the surface of flagellin with heterologous antigens. In vivo, analysis of inflammatory cytokines in serum revealed FH-VLP's superior biological safety compared with FljB (phase 2 flagellin of Salmonella typhimurium strain LT2). By incorporating the OVA peptide antigen, the FH VLP-based OVA peptide vaccine (termed FH-OVA) significantly impeded the progression of B16F10-OVA melanoma and E.G7-OVA lymphoma, as illustrated in Fig. 2f, while also stimulating the expansion of CD8+ T cells. Furthermore, the addition of clinical CpG 1018 adjuvant enhanced FH-OVA VLP immunization. Overall, FH VLP demonstrated robust immunogenicity and biosafety, suggesting its potential as a versatile platform for cancer immunotherapy.
Another animal-derived virus, norovirus, has been repurposed as an antigen presentation platform. The capsid protein VP1 of norovirus could self-assemble into VLPs when expressed in Pichia pastoris or baculovirus expression systems. Zheng et al. adopted a gene fusion approach, integrating SpyCatcher3 into the N-terminal shell domain of Norovirus VP1.46 This facilitated the binding of SpyTag-modified tumor antigens to the VLPs formed by SpyCatcher. Notably, the resulting vaccine exhibited stability across multiple freeze–thaw cycles. In vitro studies revealed a significant promotion of BMDC maturation by the vaccine. In vivo, the vaccine effectively targeted DCs in lymph nodes, triggered a systemic anti-tumor immune response, and reversed the immunosuppressive tumor microenvironment. Encouragingly, tumors established in TC-1 tumor-bearing mice even experienced complete regression post-vaccine treatment. Moreover, VLP modified with SpyCatcher could be extensively prepared from E. coli. Hence, the high yield, stability, and potent anti-tumor effects of this norovirus VLP position it as a widely applicable tumor vaccine platform.
Although various vaccine platforms based on viruses or VLPs inherently possess immune-enhancing properties akin to those of immune adjuvants, not all disease scenarios necessitate such nonspecific immune enhancement effects.44,77,95 Certain vaccine platforms rooted in viruses or VLPs may not be suitable for specific situations, making biomimetic VLPs a valuable alternative. In an earlier study, Ren et al. reported that protein nanocages assembled using the E2 protein demonstrated efficacy in delivering the anti-tumor chemotherapy drug doxorubicin.96 Molino et al. engineered the structural core of the non-viral E2 subunit of pyruvate dehydrogenase, creating a protein nanocage platform for therapeutic applications. This platform was covalently linked with DC-activating CpG molecules using an acid-releasable strategy, along with the inclusion of MHC I-restricted SIINFEKL peptide as an antigen (Fig. 2g).97In vitro, as shown in the bar chart in Fig. 2g, the concentration of activated BMDCs encapsulated with E2 NPs (termed CpG-E2) was 25 times lower than that of unbound CpG alone. Moreover, E2-bound SIINFEKL exhibited a threefold higher antigen presentation rate by BMDCs compared with the unbound antigen peptide, leading to increased activation of CD8+ T cells. To some extent, biomimetic PNPs can serve as a platform to emulate the characteristics of viruses or VLPs, thereby enhancing immunotherapy.
In general, viruses or VLPs can induce a certain degree of immune response as they are recognized as viral invasions by APCs. However, most of this response is nonspecific. Recently, OVs have gained significant attention due to their ability to directly target and kill tumors.98 Furthermore, OVs can act as immune adjuvants for peptide-based cancer vaccines.99 Capitalizing on these advantages, VLPs based on OVs enhance anti-tumor immune responses by providing tumor-derived antigens to APCs.100 Fusciello et al. designed VLPs based on OVs that encapsulated tumor cell membranes artificially, containing tumor-associated antigens. These VLPs exhibited increased infectivity and oncolytic effects both in vivo and in vitro.101 However, the therapeutic efficacy of OVs is limited by low permeability of tumor tissues and insufficient tumor-targeting effects. Addressing this, Yoon et al. utilized the tumor-homing characteristics of mesenchymal stromal cells to deliver oncolytic viruses specifically targeting tumors.102,103
2.2 Ferritin
In addition to VLPs, ferritin is also widely used as a platform for cancer immunotherapy. Generally, ferritin is a protein with a molecular weight of 450 kDa, consisting of 24 subunits self-assembled into a cage-like structure, with inner and outer sizes of 8 nm and 12 nm, respectively.104–106 Ferritin derived from eukaryotes generally has two genes encoding the heavy chain and the light chain. The heavy chain and light chain subunits have different biological functions individually, and the proportion of heavy chain and light chain subunits in ferritin varies among different organisms and tissues, displaying distinct characteristics.107 Besides, ferritin without an inorganic core is generally called apoferritin.108Its remarkable properties make ferritin widely used in cancer immunotherapy. Similar to other PNPs, ferritins can be modified through strategies such as chemical conjugation, mutation, gene fusion, and hybridization to optimize the functions. Besides, ferritin exhibits good stability can maintain a stable structure under 85 °C or some denaturants such as urea, which make it easier to be modified.109,110 Importantly, ferritin NPs possess distinct advantages over other NPNPs, such as pH responsiveness, tumor targeting and its unique structural symmetry. Specifically, ferritin exhibits controllable pH responsiveness, depolymerizing into subunits when the solution pH is <3.4 or >10 and reassembling into nanocages when the environment returns to a neutral solution. This unique pH-responsive characteristic enables ferritin to load and release drugs or other molecules.43,111 The heavy chain ferritin (HFn) exhibits a remarkable affinity for various tumor cells that express high levels of transferrin receptor 1 (TfR1), including HT-29 human colon cancer cells, SMMC-7721 human liver cancer cells, A375 melanoma cells, MDA-MB-231 breast cancer cells and many other cancer cell lines, which underscores HFn's exceptional capability in recognizing and targeting cancer cells.112 At the same time, the unique structural symmetry of ferritin can help to elicit stronger immune responses. For example, since ferritin has a 3-fold axis, it can help trimeric antigens to present with specific spatial structures on the surface of ferritin.39,113
Due to these properties, ferritin NPs loaded with photosensitizers and DOX were developed by Zhen et al. for cancer PDT and chemotherapy. Additionally, targeting cancer cells with ferritin NPs was accomplished by modifying the RGD4C sequence, facilitating specific binding to αvβ3 on the membrane of cancer cells.52,114 Numerous studies have employed ferritin NPs as a platform to contribute to cancer chemotherapy, radiotherapy, and PDT.65,115–119 Here, we will specifically focus on introducing the application of ferritin NPs in immunotherapy (Fig. 3).
 | ||
| Fig. 3 The utilization of certain ferritin NPs in the context of cancer immunotherapy. (a) OT peptide-loaded FPCNs induce proliferation of CD4+ T cells and CD8+ T cells through DC-mediated antigen presentation process [reprinted with permission from ref. 47; Copyright (2014) Elsevier]. (b) Schematic diagram of the TLR-activating nucleic acid loading process of HFn(+) through pH-mediated disassembly/reassembly [reprinted with permission from ref. 50; Copyright (2022) Elsevier]. (c) Schematic illustration of self-assembly SR717@RGE-HFn NPs [reprinted with permission from ref. 129; Copyright (2022) Elsevier]. (d) Left part illustrates human ferritin monomers modified with SIRPγ self-assembling into 24-mer ferritin NPs; right part shows FSγ and CpG potentiating antitumor immunity by promoting phagocytic activity of phagocytes [reprinted with permission from ref. 130; Copyright (2021) Springer Nature]. (e) Schematic illustration of the preparation of Nb-Ftn@ICG [reprinted with permission from ref. 133; Copyright (2024) Advanced Science]. | ||
Han's team utilized the ferritin nanoplatform as an antigen delivery NP for the development of DC-based vaccines.47 Using a gene fusion strategy, they modified OT peptides (ovalbumin antigen peptides) on the outer surface or inner cavity of ferritin to prepare ferritin protein cage NPs loaded with antigens, termed OT-FPCNs (Fig. 3a). In both in vitro and in vivo experiments, various OT-FPCNs induced the proliferation of antigen-specific CD4+ T cells and CD8+ T cells after being phagocytosed by DCs. This was accompanied by the production of cytokines such as IFN-γ and IL-2.
To enhance the efficacy and utilization of TLR agonists for activating APCs in vivo, Zhang et al. utilized biotechnology to design a human HFn NP with a positively charged cavity, denoted as HFn(+), for loading TLR agonists.50 They specifically replaced negatively charged glutamate and aspartate within the ferritin cavity with positively charged lysine or arginine, screening positive amino acids by constructing a series of HFn(+) with different mutant subunits. Loading TLR-activating nucleic acids into HFn(+) was achieved through a pH-mediated disassembly/reassembly strategy (Fig. 3b). Fluorescence tracking demonstrated that HFn carrying CpG (termed CpG@HFn(+)) was more effectively captured by DCs. In vitro, CpG@HFn(+) proved more advantageous than free CpG in promoting DC maturation, accompanied by a significant release of inflammatory cytokines such as TNF-α and IL-6. Moreover, in vivo, it exhibited a similar trend in promoting maturation of DCs in tumors. Subsequently, the carboxyl groups of chlorin e6 (Ce6), a photosensitizer, were covalently bound to the amino groups on the surface of CPG@HFn(+) through EDC/NHS-mediated carboxyl activation. The prepared Ce6-CPG@HFn(+) was used in combination with PDT, demonstrating excellent immune response and tumor-killing effects. Modifying amino acids to alter the intracavity charge of ferritin provides a novel approach for drug delivery based on ferritin NPs in immunotherapy.
In addition to its role as a modifiable NP platform, ferritin possesses unique properties. The presence of the blood–brain barrier (BBB) poses a significant challenge in treating brain diseases like gliomas, hindering the passage of drug molecules such as proteins and peptides due to its high selective permeability.120–124 The breakthrough comes in the delivery of NPs through the BBB and targeting tumor tissue. Studies have excitingly shown that ferritin has the ability to bind to the highly expressed TfR1 on endothelial cells of the BBB, facilitating delivery to the tumor through receptor-mediated endocytosis.125–128 Building upon this, Wang et al. modified the glioma-targeting REG motif on HFn through gene fusion and employed a pH-mediated disassembly/reassembly strategy to load the non-nucleotide stimulator of interferon genes, namely STING agonist SR717 (Fig. 3c).129 In the glioma model, the analysis of fluorescence signals in mice injected intravenously with Cy5.5-labeled HFn or RGE-HFn confirmed the accumulation of ferritin NPs in tumor tissue. Additionally, RGE-HFn carrying STING agonists (termed SR717@RGE-HFn) exhibited stronger CD8+ T-cell induction ability and more release of inflammatory cytokines than free agonists. The design of a dual-targeting NP platform, which can target both BBB and tumors, provides a new approach for the treatment of glioma.
Adjuvant-loaded NPs generally elicit a robust immune response by enhancing the activation of inflammatory signaling pathways, phagocytic function, and antigen presentation in APCs. In an alternative approach, Choi et al. engineered ferritin NPs with signal-regulatory protein γ (SIRPγ) to obstruct cluster of differentiation 47 (CD47) on cancer cells, inhibiting the phagocytic ability of phagocytes (Fig. 3d).130 Termed FSγ, these SIRPγ-modified ferritin NPs demonstrated the ability to enhance the phagocytosis of macrophages and DCs in vitro. In vivo, their high binding affinity with CD47 potentiated the phagocytic activity of macrophages and DCs in a mouse melanoma model, successfully achieving tumor suppression. Furthermore, the synergistic effect of CpG and NPs, depicted in the right part of Fig. 3d, further enhanced the phagocytosis of antigen-presenting cells, leading to a more successful anti-tumor effect through combination therapy.
Immunotherapy for liver cancer, exemplified by immune checkpoint inhibitors, encounters the challenge of limited efficacy.131 To enhance the effectiveness of liver cancer immunotherapy utilizing immune checkpoint inhibitors, Chen et al. devised an HFn platform loaded with DOX, termed DOX@HFn, for chemoimmunotherapy.132 Leveraging the tumor-targeting and pH-responsive properties of ferritin, NPs prompted the ICD effect of cancer cells in vitro and facilitated maturation of DCs. In vivo studies using the H22 subcutaneous liver cancer model and Hepa1–6 in situ liver tumor model demonstrated that DOX@HFn induced potent anti-tumor immune responses and exhibited efficacy in tumor eradication.
In addition to their application in combination with chemotherapy, ferritin NPs have been investigated for their potential in combining immunotherapy with PTT. Liu et al. developed an anti-PD-L1 nanobody (Nb)-ferritin nanoplatform for cancer immunotherapy.133 The association between anti-PD-L1 Nb and ferritin is facilitated by maleimido and amino bifunctionalized PEG (MAL-PEG-NH2). By employing a click reaction between the maleimide of PEG and the cysteine residue on the surface of ferritin, site-specific polyethylene glycol functionalization of ferritin nanocages was achieved. Indigo green (ICG), a photothermal agent approved by the FDA, could be encapsulated into ferritin NPs through a disassembly/reassembly process. Moreover, the antibody carrying the LLQS tag through gene fusion bound specifically to the amino base of PEG via mTGase-mediated connections, thereby obtaining NPs termed Nb-Ftn@ICG (Fig. 3e). The fusion of Nb and ferritin demonstrated prolonged in vivo circulation time and improved physical stability, addressing the issue of NP clearance by the kidneys due to the small size of the NPs. The Nb-Ftn@ICG NPs exhibited tumor-targeting capabilities, accumulating at tumor sites and inducing ICD of cancer cells upon laser irradiation. They also promoted the maturation of DCs and elicited a robust anti-tumor T-cell immune response. Moreover, the ability to block immune checkpoints and degrade PD-L1 has been confirmed. In vivo studies demonstrated this NP's effectiveness in inhibiting primary tumor progression and metastasis, highlighting the potential of this platform for cancer immunotherapy.
2.3 Albumin
Albumin abundantly exists in human blood, with a concentration of about 40 mg mL−1.134 Albumin possesses not only the advantages of non-toxicity, non-immunogenicity, excellent biodegradability and biocompatibility, but also some unique characteristics.135–137 Firstly, albumin's extended half-life makes it a great platform for carrying drugs. It mainly stems from its ability to evade renal clearance through reabsorption mediated by receptors in the proximal tubules of the kidneys. A key receptor involved in this process is the neonatal Fc receptor, widely distributed throughout the body, which is recognized for the role it plays in extending the half-life of serum albumin.138,139 Secondly, albumin's hydrophobic pockets serve as natural carriers for hydrophobic substances, facilitating the delivery of hydrophobic drugs like paclitaxel via an albumin NP platform.138,139 Moreover, enhancing the hydrophobicity of albumin can be achieved by incorporating lipophilic drugs and reducing the primary amino groups on its surface, thereby promoting the self-assembly of albumin and the formation of polymer micelles for delivering DOX.140 Hoogenboezem et al. summarized albumin NP fabrication methods, including desolvation, emulsion, thermal gelation, nanospraying, and self-assembly.139 Lin et al. induced albumin self-assembly into NPs loaded with paclitaxel by increasing hydrophobicity, thereby enhancing BBB penetration for glioma treatment.141 There are also numerous studies focusing on the use of albumin NPs in cancer chemotherapy, similar to this approach.142–144 In this review, we mainly seek examples of applications in cancer immunotherapy (Fig. 4). | ||
| Fig. 4 The utilization of certain albumin NPs in the context of cancer immunotherapy. (a) Albumin NPs enhance the efficacy of PD-1/PD-L1 blockade immunotherapy, leading to the reversal of the tumor's immunosuppressive microenvironment, which includes PGE2, Treg, and M2 macrophages [reprinted with permission from ref. (146); Copyright (2024) Springer Nature]. (b) Schematic diagram of the synthesis and therapeutic process of IR780-ZnS@HSA NPs [reprinted with permission from ref. 147; Copyright (2023) Frontiers]. (c) Preparation and application schematic of albumin NP in anti-tumor immunotherapy [reprinted with permission from ref. 149; Copyright (2024) Elsevier]. (d) Schematic diagram of the synthesis route of BSA-MHI148@SRF NPs and the mechanism of enhanced tumor photodynamic immunotherapy mediated by BSA-MHI148@SRF NPs, facilitating cascade two-stage reoxygenation and immune resensitization strategy [reprinted with permission from ref. 151; Copyright (2022) Elsevier]. | ||
Chu et al. integrated cancer immunotherapy with nanotechnology to establish a strategy of neutrophil-mediated therapeutic albumin NP deposition in tumor tissues.145 Upon injection of the monoclonal antibody TA99 specific for the gp75 antigen of melanoma in vivo, there was an enhanced transport of albumin NPs in the tumor, which was mediated by neutrophils and did not affect the function of neutrophils. This approach provides a new avenue for delivering therapeutic drugs to the tumor site, thereby enhancing cancer immunotherapy.
To enhance checkpoint blockade cancer immunotherapy, Feng et al. designed controlled-release albumin NPs called Cele-BMS-NPs, composed of pH-responsive human serum albumin derivative, BMS-202 compound as a PD-1/PD-L1 inhibitor, and a GSH-responsive celecoxib prodrug targeting COX-2 to improve anti-PD-1/PD-L1 immunotherapy (Fig. 4a).146 Under acidic tumor microenvironment conditions, NPs released BMS-202 and GSH-responsive COX-2 inhibitor. Moreover, in the mouse breast cancer model, Cele-BMS-NPs induced stronger tumor infiltration of CD8+ T cells, while inhibiting the differentiation of tumor-related macrophages and regulatory T cells (Treg) and the impact of immune suppression of prostaglandin E2 (PGE2), providing a robust strategy for improving cancer immunotherapy.
Upregulating innate immunity via the cyclic guanosine monophosphate-adenosine monophosphate synthase/interferon gene stimulator (cGAS/STING) signaling pathway has emerged as a promising strategy for enhancing cancer immunotherapy. In another study boosting the effectiveness of αPD-L1 immunotherapy, Yang et al. employed synergistic PTT, PDT, and immunotherapy to induce pyroptosis in triple-negative breast cancer.147 They developed a self-assembled human serum albumin (HSA) NP with combined PTT/PDT/immunotherapy functionalities, termed IR780-ZnS@HSA, loaded with the mitochondria-targeting PTT/PDT drug IR780 and the cGAS/STING agonist zinc sulphide (ZnS) (Fig. 4b). Additionally, intracellular zinc ions could generate ROS, further enhanced by inhibiting catalase in tumor cells and producing hydrogen sulphide (H2S) gas from ZnS.148In vitro, the NPs successfully induced ICD and pyroptosis in cancer cells by triggering the cGAS/STING signaling pathway. Furthermore, combining the NPs with laser treatment significantly impeded breast cancer progression in tumor-bearing mice, activating immunity and enhancing the efficacy of aPD-L1. This study harnessed target-specific albumin as a NP carrier to develop a novel inducer of cancer cell pyroptosis.
Utilizing the mechanism of activating the STING pathway similarly, Zheng et al. developed albumin NPs carrying STING-activating peptide, termed HSA-MnO2-pep.149 They employed an oxidation–reduction reaction between HSA and potassium permanganate (KMnO4) to synthesize biodegradable manganese dioxide for vaccine design. Manganese dioxide (MnO2) degraded into manganese ions within cells, triggering the activation of the STING pathway, which promotes DC maturation and antigen presentation (Fig. 4c). Consequently, this enhanced the immune response of anti-tumor T cells. In vitro, experiments demonstrated that the binding of antigen-stimulating peptides and albumin enhances uptake and maturation of DCs compared with free peptides. Protein imprinting and enzyme-linked immunosorbent assay (ELISA) were utilized to investigate the activation of the STING pathway by detecting downstream markers such as phosphorylated TANK-binding kinase 1 and IFN-β expression levels, affirming the efficacy of MnO2-induced pathway activation. In vivo, HSA-MnO2-pep effectively accumulated in mouse lymph nodes, promoting maturation of DCs and inducing increased IFN-γ secretion, thus contributing to tumor prevention. Furthermore, in tumor treatment models, HSA-MnO2-pep induced robust anti-tumor T-cell levels, thereby suppressing tumor progression. This albumin NP vaccine provides a solution to address issues of poor antigen stability and weak immunogenicity without apparent toxic side effects.
PDT, aside from generating ROS with photosensitizers in the presence of molecular oxygen to eliminate cancer cells, can also bolster the immune response by inducing T-lymphocyte infiltration, thereby converting “cold” tumors into “hot” ones for immunotherapy.150
To address the challenge of PDT's limited efficacy in generating reactive oxygen species due to the hypoxic tumor microenvironment, Zhou et al. employed a hydrophilic–hydrophobic self-assembly approach for fabricating bovine serum albumin (BSA)-based BSA-MHI148@SRF NPs.151 These NPs consist of the near-infrared photodynamic dye MHI148 covalently linked and sorafenib (SRF), a typical vascular endothelial growth factor (VEGF) inhibitor with effective tumor vascular normalization ability and proved to reduce tumor oxygen consumption rate, attached via hydrophilic–hydrophobic interactions, facilitating a two-stage cascade for tumor reoxygenation and immune sensitization (Fig. 4d).152In vitro, NPs demonstrated the ability to impede mitochondrial respiration, enhance reactive oxygen species (ROS) generation, and exert potent cytotoxic effects on tumor cells. In vivo, NPs exhibited tumor-targeting properties and successful accumulation in the tumor region. Moreover, they effectively induced normalization of tumor blood vessels and alleviated the hypoxic tumor microenvironment, resulting in significant suppression of solid tumor growth, which offered promising prospects for better cancer PDT.
3. Immunotherapy for other diseases
In addition, some respiratory diseases such as influenza and pneumonia also pose significant public health problems. An estimated 300000 to 500000 people die annually due to seasonal influenza epidemics, with a mortality rate of 0.2%.153 Universal inactivated, live attenuated vaccines are commonly used in the face of these diseases; however, their toxicity and low immunogenicity still have not been overcome. For example, in recent years, the world has been plagued by COVID-19. Despite the eventual success of vaccination against COVID-19, it still appears to be inadequate in the face of the new pandemic. Live attenuated vaccines currently fail to overcome worrisome toxicity and low immunogenicity.154 Other diseases, including hepatitis, face challenges similar to influenza and pneumonia. Like cancer immunotherapy, successful immunotherapy for these diseases also needs to be achieved by fully activating the body's immune response, leading to the induction of broader and more persistent antibody levels.
Using SARS-CoV-2 as an example, with the constant emergence of new variants, the efficacy of existing vaccines has gradually become a concern. Therefore, there is a need to continue developing cost-effective, heat-stable vaccines to combat widely spreading variants and potential future strains. In addition to cancer immunotherapy, NP-based platforms are also involved in the development of vaccines against other human diseases.11 NP-based vaccines, leveraging the advantages of easy self-assembly and multivalent antigen presentation, have been widely utilized in the treatment of various diseases.155 This section will introduce the applications of NPNPs and DPNPs in immunotherapy for other diseases.
3.1 NPNPs
NP-based platforms, including viruses or VLPs and ferritin, can also serve as platforms for immunotherapy of some infectious diseases (Fig. 5). | ||
| Fig. 5 The utilization of certain VLPs and ferritin NPs in immunotherapy for other diseases. (a) The modification of the icosahedral PLP shell with lambda decorative protein gpD in vitro [reprinted with permission from ref. 159; Copyright (2022) Springer Nature]. (b) Structures of SARS-CoV-2 spike-functionalized ferritin NPs and spike-pseudotyped lentivirus neutralizing titers from different groups after single antigen stimulation [reprinted with permission from ref. 113; Copyright (2021) American Chemical Society]. (c) 3D model of a spike-ferritin NP. The NP is depicted along the four-fold and three-fold symmetry axes of the spike-ferritin. Distinguish different parts with different colors: (1) ferritin molecule: alternating gray and white, (2) C-terminal coiled coil N-terminal to hinge 1: purple, (3) RBD: green, (4) NTD: blue, (5) S1 and S2 polypeptides: red and cyan [reprinted with permission from ref. 164; Copyright (2021) Elsevier]. (d) Schematic illustration of RBD and RBD-HR NPs based on ferritin [reprinted with permission from ref. 163; Copyright (2020) Elsevier]. (e) Schematic diagram of assembled HMNF NPs [reprinted with permission from ref. 162; Copyright (2023) American Chemical Society]. | ||
Plant-derived VLP vaccines containing influenza hemagglutinin (HA) have demonstrated the ability to elicit humoral immune responses and CD4+ T-cell responses. Makarkov et al. explored the immune activation mechanism of VLPs carrying plant-derived H1 protein (termed H1-VLP) by examining the intracellular fate of human monocyte-derived macrophages (MDMs).158 They discovered that MDMs underwent endocytosis mediated by clathrin, as well as macropinocytosis/phagocytosis, resulting in a threefold higher accumulation of HA from VLPs in cells compared with soluble HA. Additionally, MDMs treated with H1-VLP were found to express 115 HA-derived peptides associated with MHC I, indicating that influenza HA carried by VLPs could enhance the functions of APCs, such as antigen capture, processing, and presentation. This study clarifies the intracellular mechanism through which VLPs activate immune responses.
In a unique approach, Davenport et al. developed a vaccine utilizing the phage lambda platform.159 Their method involved engineering the lambda phage-like particle (PLP) platform to present the RBD of the coronavirus spike protein. The RBD proteins of both SARS-CoV-2 and Middle East respiratory syndrome coronavirus (MERS-CoV) were labeled RBDSARS and RBDMERS, respectively. These RBD proteins were chemically cross-linked to the decorative protein mutant gpD, S42C, on the lambda phage for display on the PLP surface (Fig. 5a). Subsequently, monovalent vaccines were developed based on RBDSARS-PLP and RBDMERS-PLP, respectively. Additionally, they designed a bivalent vaccine, termed hCoV-RBD-PLP, capable of simultaneously presenting the spike RBD proteins of both viruses. Remarkably, mice immunized with 60% RBDSARS-PLPs maintained high levels of RBDSARS-specific IgG at day 174 post-immunization, indicating the vaccine's ability to induce durable humoral immune responses, including the production of neutralizing antibodies. This immunity conferred protection against pulmonary infection, inflammation, and pathology when challenged with SARS-CoV-2 and MERS-CoV. These findings underscore the potential of this vaccine platform for flexible and controlled vaccine design and development.
In addition to the biomimetic VLPs employed in diseases treatment, they have also been designed for diagnostic purposes. For example, Chan et al. have pioneered the design of a biomimetic VLP platform for detecting SARS-CoV-2, which was proved to possess enhanced stability, rendering it suitable for clinical trials.160
Ferritin is emerging as an ideal carrier for the development of COVID-19 vaccines. Powell and collaborators devised subunit vaccines utilizing self-assembling ferritin NPs, showcasing either of the two polymeric SARS-CoV-2 spike proteins: the full-length extracellular domain (termed S-Fer) or a C-terminal 70 amino acid deletion (termed SΔC-Fer).113 Initial NPs were generated through DNA plasmid construction and protein purification, with western blot confirmation of spike antigen expression levels. Additionally, two spike outer domain proteins fused to a trimeric coiled-coil, GCN4-pIQI (termed S-GCN4 and SΔC-GCN4), were produced for comparison (Fig. 5b). Mice immunized with the two spike ferritin particles exhibited significantly higher induced neutralizing antibody intensity compared with RBD or spike domain trimer alone. Notably, the average neutralizing antibody titers in the spike-like ferritin particle-treated group were twice that of plasma from 20 convalescent COVID-19 patients (termed CCP), as illustrated in Fig. 5b. Upon the second antigen stimulation, animals immunized with SΔC-Fer demonstrated higher neutralizing antibody titers than other groups. Consequently, developing a multivalent SARS-CoV-2 spike antigen presentation platform vaccine based on ferritin NPs proves to be a viable strategy against COVID-19.
There exist many other structures of ferritin NPs like this. Joyce et al. designed a spike-ferritin NP that could display eight spikes.164 In this design, the Helicobacter pylori ferritin molecule underwent genetic modification at the C-terminus of the SARS-CoV-2 spike (abbreviated as S) antigen, encompassing the S1 attachment subunit, S2 fusion subunit from the class I fusion glycoprotein S, RBD, and N-terminal domain (termed NTD) (Fig. 5c). The stable binding between the ferritin scaffold and trimer relied on the C-terminal coiled N-terminal to hinge 1, depicted in purple in the protein structure of Fig. 5c. In vivo experiments verified that these immunogens could elicit a robust and protective neutralizing antibody response against SARS-CoV-2.
In addition, researchers have also developed some multivalent ferritin NP vaccines that can present two or more antigens. Ma et al. strategically linked SpyTag (abbreviated as ST) to the N-terminus of RBD or heptad repeat (termed HR), and SpyCatcher (abbreviated as SC) to the N-terminus of ferritin through gene fusion. Subsequently, purified SC-ferritin was combined with ST-RBD and/or ST-HR in buffer, facilitating the conjugation of antigen subunits to ferritin via a stable intermolecular isopeptide bond formed between SC and ST (Fig. 5d).163 Following SARS-CoV-2 challenge, mice immunized with RBD and RBD-HR NPs exhibited significantly lower viral loads in their lungs. Additionally, RBD-HR NPs effectively elicited neutralizing antibody responses and immune activation against other coronaviruses. These studies offer viable vaccination strategies for combatting SARS-CoV-2 and other coronaviruses.
With another multivalent ferritin NP vaccine, Pan et al. devised a universal influenza candidate vaccine, aiming to confer comprehensive protection against both influenza A and B viruses to address the limited efficacy of influenza vaccines.162 The engineered self-assembling ferritin NP, termed HMNF, efficiently displayed HA A Helix antigens (abbreviated as H), M2e antigens (abbreviated as M), and NA HCA-2 (abbreviated as N) antigens simultaneously after purification (Fig. 5e). This achievement involved fusing three types of antigen information in tandem to the human heavy chain ferritin. Mice subjected to intranasal vaccination with HMNF exhibited an augmented production of viral peptide-specific IgG antibodies, along with elevated secretion of IFN-γ and IL-4 in the spleens. Upon challenging mice with diverse influenza strains from IAV and IBV, such as H1N1, H3N2, H6N6, H9N2, etc., the outcomes in terms of body weight and survival rate underscored the prophylactic potential and cross-protective capability of HMNF NPs. Most notably, HMNF NPs effectively induced sustained antibody protection that endured for six months. This universal influenza vaccine, demonstrating competence in providing enduring protection, furnishes a strategic response to both existing and potential future influenza viruses.
3.2 DPNPs
The extensive coding information embedded in protein amino acid sequences endows them with the capacity to engage in various intricate life processes. The number of distinct sequences for a protein of typical length is 20200 sequences, but the number of these proteins that are discovered in nature is on the order of 1012. In other words, naturally occurring proteins occupy a tiny fraction of protein sequence space, leaving a huge space for de novo designed proteins that might have more coding information and functions. Besides, de novo designed proteins generally exhibit enhanced stability due to their adherence to the physical principle of the lowest energy state, which makes DPNPs more tolerant of surface modification through genetic fusion or chemical modification.66 Additionally, in contrast to the top-down modification of NPNPs discussed in the preceding section, which constrained them to a design space near their initial state, for DPNPs the bottom-up design strategy is adopted, boasting a broader design spectrum.37 Moreover, they possess controllable physicochemical properties, environmental responsiveness, and substantial loading capacity, thereby opening avenues for DPNPs to actively contribute to immunotherapy.66,165 Herein, we showcase some promising structures poised for inclusion in experimental immunotherapy model research, along with an introduction to experimentally validated applications of DPNPs.King et al. have devoted themselves to the computational design of protein assemblies, generating numerous DPNPs with specific spatial structures through precise computational design. These structures hold promise as platforms for future immunotherapy (Fig. 6). The computational method allows for the predictive design of self-assembled protein nanomaterials with atomic-level accuracy, and they can be synthesized through in vitro assembly.166 In 2014, King et al. designed five 24-subunit protein nanocages using symmetric docking and protein–protein interface design methods, anticipating their widespread application in designing symmetric structures (Fig. 6a).67 The experimental findings have validated the alignment between protein structure and computational design, showcasing the potential of DPNPs in drug delivery and vaccine development. In 2016, King's team engineered an icosahedral protein nanocage with a meticulously ordered structure, employing computational design and self-assembly through ternary protein building blocks (Fig. 6b).167 Verification of the congruence between the calculated design and the actual protein structure was accomplished through electron microscopy observation, and reversible protein decomposition could be executed under specific conditions. Subsequently, they synthesized icosahedral protein assemblies using computational design. The inner surface of these positively charged protein assemblies encapsulated their own full-length mRNA genomes through electrostatic interactions, mirroring the way viruses carry their genetic information (Fig. 6c).168 The attainment of high stability in the bloodstream and the enhancement of genome encapsulation efficiency were realized through the ongoing evolution of nucleocapsids. These non-viral DPNPs can function as a non-toxic platform for drug loading and mRNA influenza vaccines, opening avenues to novel strategies in immunotherapy.
 | ||
| Fig. 6 Several structures of DPNPs are anticipated for utilization in immunotherapy. (a) Symmetrical displayed crystal structure of one designed protein [reprinted with permission from ref. 67; Copyright (2014) Springer Nature]. (b) Computational design of icosahedral protein nanocage structures in various orientations [reprinted with permission from ref. 167; Copyright (2016) Springer Nature]. (c) Strategy for self-assembly icosahedral capsids encapsulating own mRNA genomes [reprinted with permission from ref. (168); Copyright (2017) Springer Nature]. | ||
In the following years, King's team continued to develop DPNP-based vaccines for various diseases (Fig. 7). Previously, they synthesized a protein structure known as I53-50, a computationally designed two-component protein complex composed of 20 trimer “A” and 12 pentamer “B”, totaling 120 subunits.169 Based on this structure, to address the challenge of effectively eliciting broadly neutralizing antibodies against human immunodeficiency virus 1 (HIV-1) through vaccination, they introduced a novel PNP platform termed SOSIP-I53 50A NPs. This platform employed a strategy of presenting antigens in a multivalent manner on PNPs to enhance immunogenicity and stimulate extensive antibody production in immunotherapy (Fig. 7a).170 The two-component PNPs could present 20 stable SOSIP trimers from different HIV-1 strains. Leveraging the properties of bicomponent proteins, native-like HIV-1 envelope trimers could be efficiently loaded into DPNPs. Experimental results demonstrated that the designed two-component protein induced more potent neutralizing antibodies compared with traditional one-component NPs.
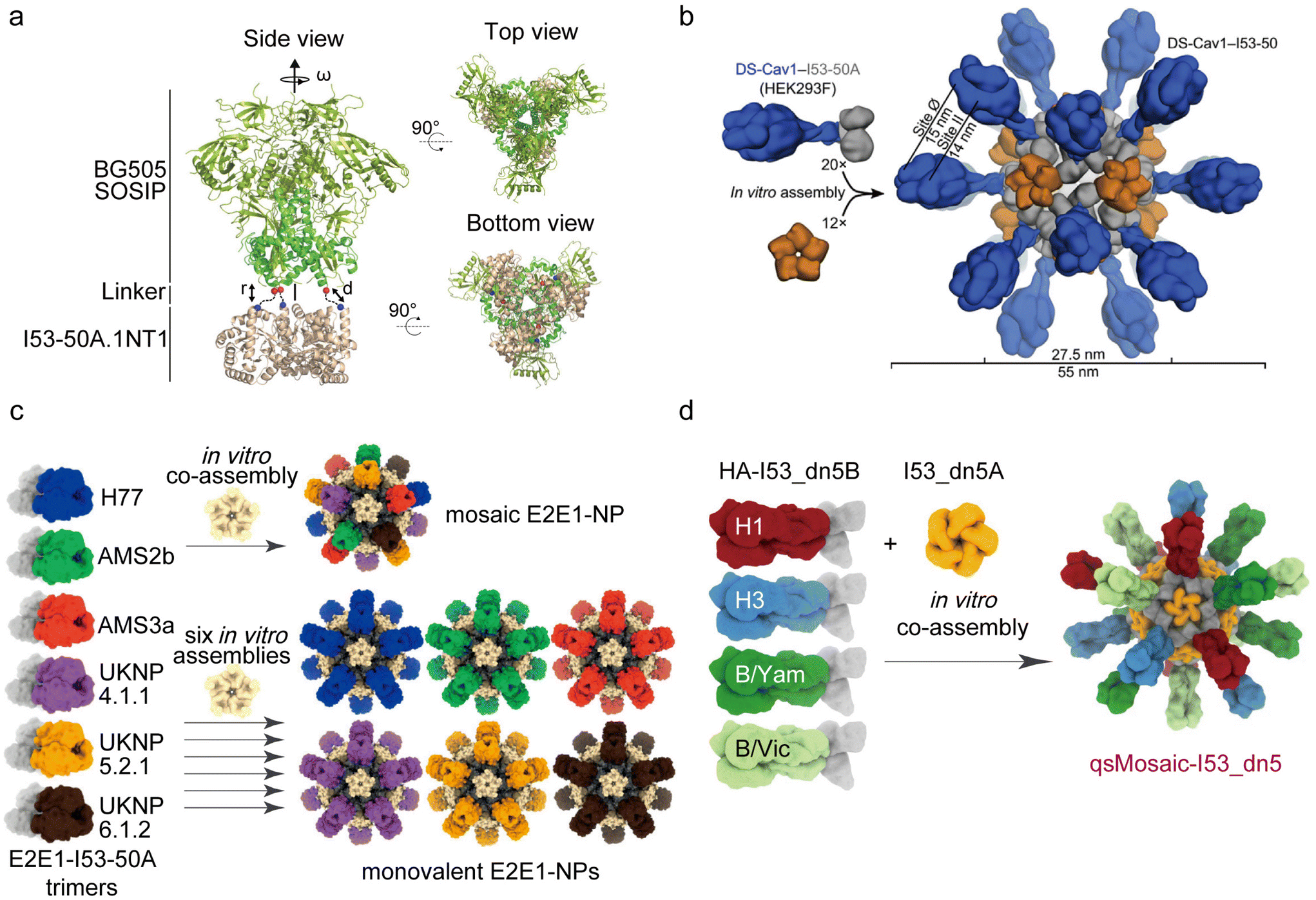 | ||
| Fig. 7 Some DPNPs used in immunotherapy for certain diseases. (a) Schematic representation of the computational docking protocol for SOSIP-I53 50A trimeric fusion protein [reprinted with permission from ref. 170; Copyright (2019) Springer Nature]. (b) Schematic representation of the computational docking protocol used to identify NP components suitable for fusion to DS-Cav1 [reprinted with permission from ref. 171; Copyright (2019) Elsevier]. (c) Six E2E1-I53-50A trimers are combined and mixed with I53-50B pentamer to co-assemble as mosaic E2E1-NPs (top) or are individually mixed with I53-50B to assemble as monovalent E2E1-NP (bottom) [reprinted with permission from ref. 172; Copyright (2022) Springer Nature]. (d) Strategy of designing a mosaic NP immunogen that co-displayed the four HAs [reprinted with permission from ref. 174; Copyright (2021) Springer Nature]. | ||
In another DPNP they designed, termed DS-Cav1-I53-50, the respiratory syncytial virus (RSV) F glycoprotein trimer (termed DS-Cav1) was presented in a repeated array outside the DPNP (Fig. 7b).171 Animal experiments revealed that the neutralizing antibody response induced by the full-valent NP immunogen of 20 DS-Cav1 trimers was 10 times higher than that of the trimeric DS-Cav1. This demonstrates that the two-component DPNP platform holds significant promise in developing multivalent antigen-presenting respiratory syncytial virus vaccines.
Similarly, a vaccine platform for hepatitis C virus (HCV), named E2E1-NP, has been developed to present the trimeric E1E2 glycoprotein. The E1E2 glycoprotein plays a crucial role in inducing potent neutralizing antibodies, distinguishing it from commonly used recombinant glycoprotein vaccines that only carry E2. King et al. designed a two-component DPNP with the ability to simultaneously display the E2E1 immunogens, resulting in a broader range of neutralizing antibody reactions compared with E2 alone. Additionally, they designed a mosaic PNP that displays six different E2E1 immunogens (H77, AMS2b, AMS3a, UKNP4.1.1, UKNP5.2.1, UKNP6.1.2) (Fig. 7c).172 This mosaic E2E1-NP vaccine could induce a more effective neutralization effect compared with DPNPs with a monovalent display of E2E1 immunogens. This suggests that the mosaic E2E1-NP vaccine may be more competitive in HCV vaccines when compared with vaccines targeting severe E2-specific broadly neutralizing antibodies.
In the realm of influenza vaccines, the HA glycoprotein serves as the primary immunogen for inducing antibody responses.173 King's team designed another mosaic influenza DPNP vaccine, termed qsMosaic-I53_dn5, capable of co-displaying four trimeric HAs (Fig. 7d).174 This quadrivalent influenza vaccine demonstrated the ability to induce stronger antibody responses than commercial quadrivalent influenza vaccines in several animal models. Importantly, the vaccine-induced serum antibodies effectively addressed the challenge of heterosubtypic influenza viruses, promoting potent receptor-blocking and cross-reactive stem-directed responses. Collectively, these findings suggest the potential of replacing conventional seasonal vaccines.
Hence, DPNPs exhibit extensive potential in the realm of immunotherapy, particularly for various infectious diseases. Leveraging computational design methodologies, DPNPs acquire distinct nanocage structures conducive to drug loading, concurrently facilitating the presentation of multiple immunogens to amplify neutralizing antibody responses.171,175 The I53-50NP platform, serving as the foundation for an RSV vaccine, is presently undergoing assessment in Phase I clinical trials. Notably, following the successful culmination of Phase 1 and Phase 3 investigations, South Korea has recently granted approval for a COVID-19 vaccine rooted in the I53-50NP platform.176
4. Conclusions and future perspectives
This review consolidates recent applications of PNPs, categorized as NPNPs and DPNPs, within the realm of immunotherapy for cancer and various other diseases. We highlighted the advantages of VLPs, ferritin, and albumin as immunotherapy platforms among NPNPs, all boasting excellent biological safety, chemical and genetic modifiability. The difference is that while ferritin and albumin exhibit low immunogenicity, many VLPs possess a certain degree of immune stimulation capability. We also introduced several structures of DPNPs and discussed their applications as vaccines for other diseases, like influenza, pneumonia, and hepatitis.With ongoing exploration, an increasing number of scholars are considering intervention in the body's immune response, aiming to stimulate immune system activation, particularly enhancing APC function to induce robust proliferation of specific T cells as part of immunotherapy.17,177–179 A common impediment encountered in immunotherapy pertains to the low immunogenicity of therapeutic drugs and vaccines in combatting cancer and viral infections.11,43,180,181 PNPs, characterized by high biosafety and biocompatibility, offer novel solutions for antigen or immune adjuvant delivery. Unlike other materials, the cell-targeting capability of PNPs can be augmented through genetic modification of antigens, such as peptides or proteins. This genetically modified structure can be efficiently expressed and purified in large quantities using E. coli and can be more effectively delivered to APCs than chemically bonded antigens.111 Consequently, PNPs possess the potential for multifunctionality, with further emphasis on functionalization evident in DPNPs based on computational design. Furthermore, PNP-based cancer immunotherapy can be synergistically combined with chemotherapy, PDT, and other treatment modalities, thereby bolstering the anti-tumor effect. In the context of influenza, pneumonia, etc., PNP-based vaccines exhibit advantages over traditional vaccines by eliciting stronger and more enduring neutralizing antibodies.
However, the widespread applications of PNPs still face certain challenges, which need attention to find solutions.
Firstly, in cancer immunotherapy, when VLPs serve as immune adjuvants, antibodies generated against VLPs do not interfere with their immune-stimulating ability, as seen with some plant-derived viruses like TMV and CPMV. However, when VLPs act as antigen delivery platforms, frequent stimulation may lead to the production of antibodies against VLPs, reducing their circulating half-life and leading to clearance.182 Similarly, in immunotherapy for diseases like influenza and pneumonia, neutralizing antibodies against VLP platforms pose a significant limitation, hindering cargo delivery to target tissues or cells.37 Modifying “don't-eat-me” signal such as CD47 can prevent clearance by phagocytosis, while lipid encapsulation of PNPs is another effective method to mitigate antibody responses.183,184 Nevertheless, these modifications may affect the efficacy of immune stimulation, necessitating the search for new strategies to address neutralizing antibody responses when VLPs serve as carriers. Beyond VLPs, nonspecific cell uptake, protein corona formation, and the mononuclear phagocytic system present barriers to antigen/drug delivery systems based on most PNPs.23 These obstacles also apply to other types of nanomaterials such as inorganic and polymeric NPs, emphasizing the need for comprehensive consideration and cross-referencing when addressing these challenges.
Secondly, when PNPs function as a delivery platform, careful consideration must be given to the intensity of their induced immunogenicity. In certain cases, the immunogenicity of these carrier platforms surpasses that of the adjuvants or antigens they carry, as APCs perceive them as authentic viral invasions. For example, CPMV VLPs have demonstrated the activation of TLR7/8 and the induction of interferon secretion.86 Additionally, previously introduced P22 VLPs exhibit robust immune activation effects.91 However, in certain circumstances, such induced immune responses have not been thoroughly investigated for their impact on the immune mechanism, potentially leading to an excessive immune response. Therefore, the immunogenicity of PNPs should be thoroughly considered when addressing diverse diseases.
Thirdly, designing PNPs with smart responsiveness is crucial in immunotherapy. Some NPNPs exhibit responsiveness, such as ferritin, which disassembles when the pH is lower than 3, a condition not precisely matched in the tumor microenvironment (pH ≈ 6). Therefore, creating protein materials with genuine responsiveness is important. Computational development has facilitated the design of DPNPs with specific structures and dynamic functional controls. By precisely designing the number and location of histidines on the surface of DPNP subunits, responsiveness to disease microenvironments can be achieved. In addition to pH responsiveness, the design of enzyme-responsive or light-responsive NPNPs is one of the future directions for PNPs in immunotherapy.
In addition, other characteristics of PNPs, such as particle size, charge, and hydrophobicity, warrant comprehensive exploration regarding their impact on drug delivery and immune stimulation before clinical application. For instance, ferritin can achieve positively charged lumens through modifications in individual amino acids, thereby improving drug loading efficiency. In the realm of cancer treatment, alterations in particle size can further enhance the EPR effect of PNPs, rendering them more advantageous for accumulation in the lesion area.
Amid mounting concerns regarding the safety of chemical and biomaterials over recent decades, the concept of green biomaterials has emerged, aligning with environmental and physiological principles.185 In cancer treatment, the unavoidable toxic side effects of chemotherapy associated with chemical materials persist, while vaccine development for different diseases also demands stringent safety considerations. Hence, the advancement of green biomaterials is imperative. In accordance with the core tenets of green biomaterials, PNPs offer promising attributes for environmentally friendly materials for several reasons: (1) protein materials can be synthesized using biological systems like E. coli, reducing reliance on fossil fuels and minimizing the carbon footprint; (2) most protein materials are compatible with aqueous environments, eliminating the need for hazardous solvents; (3) many protein materials, such as ferritin and albumin, are sourced from nature, with their biological safety extensively validated; (4) protein materials exhibit exceptional biodegradability, undergoing enzymatic degradation within the human body.24 Furthermore, gene modification and the DPNPs inspired by nature contribute to the expanded functionalities of protein materials. Consequently, protein materials hold vast potential as green biomaterials.
Over the decades, PNPs have undergone continuous development, evolving from the initial exploration of protein properties to their ultimate applications. Their contribution to disease treatment extends far beyond the scope outlined in this article. From an immunotherapy standpoint, the utilization of PNPs must parallel the ongoing progress in developing superior immune adjuvants and more effective immunotherapy strategies. The anticipation lies in the discovery of enhanced NPNP-based platforms. Moreover, the anticipation extends to the envisioning of diverse designs for DPNPs and their applications in cancer immunotherapy.
Abbreviations
| NPs | Nanoparticles |
| PNPs | Protein nanoparticles |
| NPNPs | Naturally occurring protein nanoparticles |
| DPNPs | de novo designed protein nanoparticles |
| LNPs | Lipid nanoparticles |
| EPR | Enhanced permeability and retention |
| APCs | Antigen-presenting cells |
| ICD | Immunogenic cell death |
| LPS | Lipopolysaccharide |
| PLA | Poly(lactic acid) |
| PLGA | Poly(lactic-co-glycolic acid) |
| FDA | The U.S. Food and Drug Administration |
| E. coli | Escherichia coli |
| TLRs | Toll-like receptors |
| TAM | Tumor-associated macrophage |
| DOX | Doxorubicin |
| PDT | Photodynamic therapy |
| PTT | Photothermal therapy |
| ROS | Reactive oxygen species |
| VLPs | Virus-like particles |
| DCs | Dendritic cells |
| iDC | Immature dendritic cell |
| mDC | Mature dendritic cell |
| BMDCs | Bone marrow-derived dendritic cells |
| OVs | Oncolytic virus |
| CPMV | Cowpea mosaic virus |
| CCMV | Chlorotic mottle virus |
| PhMV | Physalis mosaic virus |
| SeMV | Sesbania mosaic virus |
| HBVc | Hepatitis B virus capsids |
| CPSMV | Cowpea severe mosaic virus |
| TRSV | Tobacco ring spot virus |
| HPV | Human papillomavirus |
| RHDV | Rabbit haemorrhagic disease virus |
| ssRNAs | Single-stranded RNAs |
| DLN | Draining lymph nodes |
| KO | Knockout |
| TMV | Tobacco mosaic virus |
| PVX | Potato virus X |
| Cy5.5 | Cyanine5.5 |
| FljB | Phase 2 flagellin of Salmonella typhimurium strain LT2 |
| DLS | Dynamic light scattering |
| SAXS | Small-angle X-ray scattering |
| HFn | Heavy chain ferritin |
| Ce6 | Chlorin e6 |
| BBB | Blood–brain barrier |
| TfR1 | Transfer receptor 1 |
| SIRPγ | Signal-regulatory protein γ |
| CD47 | Cluster of differentiation 47 |
| Nb | Nanobody |
| MAL-PEG-NH2 | Maleimido and amino bifunctionalized PEG |
| ICG | Indigo green |
| Treg | Regulatory T cells |
| PGE2 | Prostaglandin E2 |
| HSA | Human serum albumin |
| ZnS | Zinc sulphide |
| H2S | Hydrogen sulphide |
| KMnO4 | Potassium permanganate |
| MnO2 | Manganese dioxide |
| ELISA | Enzyme-linked immunosorbent assay |
| BSA | Bovine serum albumin |
| SRF | Sorafenib |
| VEGF | Vascular endothelial growth factor |
| SARS | Severe acute respiratory syndrome |
| HIV-1 | Human immunodeficiency virus 1 |
| NDV | Newcastle disease virus |
| LAIV | Live attenuated influenza virus |
| RBD | Receptor binding domain |
| HA | Hemagglutinin |
| MDMs | Monocyte-derived macrophages |
| PLP | Phage-like particle |
| MERS-CoV | Middle East respiratory syndrome coronavirus |
| RSV | Respiratory syncytial virus |
| HCV | Hepatitis C virus |
Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was financially supported by the National Natural Science Foundation of China (62227803, 62288102, 62235008, 22307056), the Natural Science Foundation of Jiangsu Province-Major Project (BK20212012), Natural Science Foundation of Jiangsu Province (BK20220387), the Belt and Road Innovation Cooperation Project of Jiangsu (BZ2022011).References
- K. Yang, A. Halima and T. A. Chan, Nat. Rev. Clin. Oncol., 2023, 20, 604–623 CrossRef CAS PubMed.
- Y. Chao and Z. Liu, Nat. Rev. Bioeng., 2023, 1, 125–138 CrossRef.
- F. L. van de Veerdonk, E. Giamarellos-Bourboulis, P. Pickkers, L. Derde, H. Leavis, R. van Crevel, J. J. Engel, W. J. Wiersinga, A. P. J. Vlaar, M. Shankar-Hari, T. van der Poll, M. Bonten, D. C. Angus, J. W. M. van der Meer and M. G. Netea, Nat. Med., 2022, 28, 39–50 CrossRef CAS PubMed.
- B. A. Arslan and A. C. Timucin, Acta Virol., 2020, 64, 389–395 CrossRef CAS PubMed.
- R. S. Wallis, A. O'Garra, A. Sher and A. Wack, Nat. Rev. Immunol., 2023, 23, 121–133 CrossRef CAS PubMed.
- S.-Y. Qin, Y.-J. Cheng, Q. Lei, A.-Q. Zhang and X.-Z. Zhang, Biomaterials, 2018, 171, 178–197 CrossRef CAS PubMed.
- N. Shaverdian, A. E. Lisberg, K. Bornazyan, D. Veruttipong, J. W. Goldman, S. C. Formenti, E. B. Garon and P. Lee, Lancet Oncol., 2017, 18, 895–903 CrossRef CAS PubMed.
- A. Brufsky, S. B. Kim, Ž. Zvirbule, A. Eniu, J. Mebis, J. H. Sohn, M. Wongchenko, S. Chohan, R. Amin, Y. Yan, V. McNally, D. Miles and S. Loi, Ann. Oncol., 2021, 32, 652–660 CrossRef CAS PubMed.
- G. I. Ellis, N. C. Sheppard and J. L. Riley, Nat. Rev. Genet., 2021, 22, 427–447 CrossRef CAS PubMed.
- M. N. Wykes and S. R. Lewin, Nat. Rev. Immunol., 2018, 18, 91–104 CrossRef CAS PubMed.
- F. Zarreen Simnani, D. Singh, P. Patel, A. Choudhury, A. Sinha, A. Nandi, S. Kumar Samal, S. K. Verma and P. Kumar Panda, Mater. Today, 2023, 66, 371–408 CrossRef CAS.
- V.-K. Lakshmanan, S. Jindal, G. Packirisamy, S. Ojha, S. Lian, A. Kaushik, A. I. M. A. Alzarooni, Y. A. F. Metwally, S. P. Thyagarajan, Y. Do Jung and S. Chouaib, Cancer Gene Ther., 2021, 28, 911–923 CrossRef CAS PubMed.
- V. Gowd, A. Ahmad, M. Tarique, M. Suhail, T. A. Zughaibi, S. Tabrez and R. Khan, Semin. Cancer Biol., 2022, 86, 624–644 CrossRef CAS PubMed.
- B. Zhou, J. Liu, M. Lin, J. Zhu and W. R. Chen, Coord. Chem. Rev., 2021, 442, 214009 CrossRef CAS.
- E. J. Comparetti, N. N. Ferreira, L. M. B. Ferreira, R. Kaneno and V. Zucolotto, J. Biomed. Mater. Res., Part A, 2022, 110, 1166–1181 CrossRef CAS PubMed.
- G. Yang, H. Gong, T. Liu, X. Sun, L. Cheng and Z. Liu, Biomaterials, 2015, 60, 62–71 CrossRef CAS PubMed.
- J. Chen and X. Cong, Biomed. Pharmacother., 2023, 157, 113998 CrossRef CAS PubMed.
- R. S. Riley, C. H. June, R. Langer and M. J. Mitchell, Nat. Rev. Drug Discovery, 2019, 18, 175–196 CrossRef CAS PubMed.
- M. Yang, J. Li, P. Gu and X. Fan, Bioact. Mater., 2021, 6, 1973–1987 CAS.
- M. E. Davis, Z. Chen and D. M. Shin, Nat. Rev. Drug Discovery, 2008, 7, 771–782 CrossRef CAS PubMed.
- A. Bhardwaj, J. Kaur, M. Wuest and F. Wuest, Nat. Commun., 2017, 8, 1 CrossRef PubMed.
- M. Kunitski, N. Eicke, P. Huber, J. Köhler, S. Zeller, J. Voigtsberger, N. Schlott, K. Henrichs, H. Sann, F. Trinter, L. P. H. Schmidt, A. Kalinin, M. S. Schöffler, T. Jahnke, M. Lein and R. Dörner, Nat. Commun., 2019, 10, 1 CrossRef CAS PubMed.
- B. Nguyen and N. H. Tolia, npj Vaccines, 2021, 6, 70 CrossRef CAS PubMed.
- A. Jain, S. K. Singh, S. K. Arya, S. C. Kundu and S. Kapoor, ACS Biomater. Sci. Eng., 2018, 4, 3939–3961 CrossRef CAS PubMed.
- X. Zhuang, T. Wu, Y. Zhao, X. Hu, Y. Bao, Y. Guo, Q. Song, G. Li, S. Tan and Z. Zhang, J. Controlled Release, 2016, 228, 26–37 CrossRef CAS PubMed.
- A. Pantel, C. Cheong, D. Dandamudi, E. Shrestha, S. Mehandru, L. Brane, D. Ruane, A. Teixeira, L. Bozzacco, R. M. Steinman and M. P. Longhi, Eur. J. Immunol., 2012, 42, 101–109 CrossRef CAS PubMed.
- P. L. Felgner, T. R. Gadek, M. Holm, R. Roman, H. W. Chan, M. Wenz, J. P. Northrop, G. M. Ringold and M. Danielsen, Proc. Natl. Acad. Sci. U. S. A., 1987, 84, 7413–7417 CrossRef CAS PubMed.
- X. Yang, S. Yang, H. Chai, Z. Yang, R. J. Lee, W. Liao and L. Teng, PLoS One, 2015, 10, e0136649 CrossRef PubMed.
- Y. Wang, Z. Zhang, J. Luo, X. Han, Y. Wei and X. Wei, Mol. Cancer, 2021, 20, 33 CrossRef CAS PubMed.
- C. F. van der Waalse, G. Sharma and M. N. V. Ravi Kumar, Expert Opin. Drug Delivery, 2009, 6, 177–186 CrossRef PubMed.
- L. Mao, P. Ma, X. Luo, H. Cheng, Z. Wang, E. Ye, X. J. Loh, Y.-L. Wu and Z. Li, ACS Nano, 2023, 17, 9826–9849 CrossRef CAS PubMed.
- I. W. Hamley, Biomacromolecules, 2019, 20, 1829–1848 CrossRef CAS PubMed.
- J. López-Sagaseta, E. Malito, R. Rappuoli and M. J. Bottomley, Comput. Struct. Biotechnol. J., 2016, 14, 58–68 CrossRef PubMed.
- F. Sandra, N. U. Khaliq, A. Sunna and A. Care, Nanomaterials, 2019, 9, 1329 CrossRef CAS PubMed.
- B. J. G. E. Pieters, M. B. van Eldijk, R. J. M. Nolte and J. Mecinović, Chem. Soc. Rev., 2016, 45, 24–39 RSC.
- N. Zhang, K. Mei, P. Guan, X. Hu and Y. Zhao, Small, 2020, 16, 1907256 CrossRef CAS PubMed.
- A. Olshefsky, C. Richardson, S. H. Pun and N. P. King, Bioconjugate Chem., 2022, 33, 2018–2034 CrossRef CAS PubMed.
- C. N. Fries, E. J. Curvino, J.-L. Chen, S. R. Permar, G. G. Fouda and J. H. Collier, Nat. Nanotechnol., 2021, 16, 1–14 CrossRef CAS PubMed.
- M. Kanekiyo, C.-J. Wei, H. M. Yassine, P. M. McTamney, J. C. Boyington, J. R. R. Whittle, S. S. Rao, W.-P. Kong, L. Wang and G. J. Nabel, Nature, 2013, 499, 102–106 CrossRef CAS PubMed.
- M. Xu, R. Zeng, J. Xiang and Q. Yan, CCS Chem., 2020, 3, 2223–2232 CrossRef.
- L. Li and G. Chen, J. Am. Chem. Soc., 2022, 144, 16232–16251 CrossRef CAS PubMed.
- Y. H. Choi and H.-K. Han, J. Pharm. Investig., 2017, 48, 43–60 CrossRef PubMed.
- S. Hong, D. W. Choi, H. N. Kim, C. G. Park, W. Lee and H. H. Park, Pharmaceutics, 2020, 12, 604 CrossRef CAS PubMed.
- J. C. Caldeira, M. Perrine, F. Pericle and F. Cavallo, Viruses, 2020, 12, 488 CrossRef CAS PubMed.
- K. Cheng, T. Du, Y. Li, Y. Qi, H. Min, Y. Wang, Q. Zhang, C. Wang, Y. Zhou, L. Li, S. Ye, X. Zhou, S. Bi, J. Yang and L. Ren, ACS Appl. Mater. Interfaces, 2020, 12, 53682–53690 CrossRef CAS PubMed.
- P. Zheng, Y. Yang, Y. Fu, J. He, Y. Hu, X. Zheng, B. Duan, M. Wang, Q. Liu, W. Li, D. Li, Y. Yang, Z. Yang, X. Yang, W. Huang and Y. Ma, ACS Nano, 2023, 17, 3412–3429 CrossRef CAS PubMed.
- J.-A. Han, Y. J. Kang, C. Shin, J.-S. Ra, H.-H. Shin, S. Y. Hong, Y. Do and S. Kang, Nanomedicine, 2014, 10, 561–569 CrossRef CAS PubMed.
- E. Jung, Y. H. Chung and N. F. Steinmetz, Bioconjugate Chem., 2023, 34, 1596–1605 CrossRef CAS PubMed.
- C. I. Nkanga, O. A. Ortega-Rivera and N. F. Steinmetz, Nanomedicine, 2022, 44, 102573 CrossRef CAS PubMed.
- B. Zhang, X. Chen, G. Tang, R. Zhang, J. Li, G. Sun, X. Yan and K. Fan, Nano Today, 2022, 46, 101564 CrossRef CAS.
- K. L. Lee, A. A. Murray, D. H. T. Le, M. R. Sheen, S. Shukla, U. Commandeur, S. Fiering and N. F. Steinmetz, Nano Lett., 2017, 17, 4019–4028 CrossRef CAS PubMed.
- Z. Zhen, W. Tang, H. Chen, X. Lin, T. Todd, G. Wang, T. Cowger, X. Chen and J. Xie, ACS Nano, 2013, 7, 4830–4837 CrossRef CAS PubMed.
- S. Geninatti Crich, M. Cadenazzi, S. Lanzardo, L. Conti, R. Ruiu, D. Alberti, F. Cavallo, J. C. Cutrin and S. Aime, Nanoscale, 2015, 7, 6527–6533 RSC.
- M. Liu, Y. Zhu, T. Wu, J. Cheng and Y. Liu, Chem. – Eur. J., 2020, 26, 7442–7450 CrossRef CAS PubMed.
- P. P. Karmali, V. R. Kotamraju, M. Kastantin, M. Black, D. Missirlis, M. Tirrell and E. Ruoslahti, Nanomedicine, 2009, 5, 73–82 CrossRef CAS PubMed.
- R. Prajapati, E. Garcia-Garrido and Á. Somoza, Cancers, 2021, 13, 3011 CrossRef CAS PubMed.
- H. Ren, J. Liu, F. Su, S. Ge, A. Yuan, W. Dai, J. Wu and Y. Hu, ACS Appl. Mater. Interfaces, 2017, 9, 3463–3473 CrossRef CAS PubMed.
- S. Shukla, C. Wang, V. Beiss, H. Cai, T. Washington, A. A. Murray, X. Gong, Z. Zhao, H. Masarapu, A. Zlotnick, S. Fiering and N. F. Steinmetz, Biomater. Sci., 2020, 8, 5489–5503 RSC.
- D. C. Parker, Annu. Rev. Immunol., 1993, 11, 331–360 CrossRef CAS PubMed.
- M. F. Bachmann and G. T. Jennings, Nat. Rev. Immunol., 2010, 10, 787–796 CrossRef CAS PubMed.
- L. He, N. de Val, C. D. Morris, N. Vora, T. C. Thinnes, L. Kong, P. Azadnia, D. Sok, B. Zhou, D. R. Burton, I. A. Wilson, D. Nemazee, A. B. Ward and J. Zhu, Nat. Commun., 2016, 7, 12041 CrossRef CAS PubMed.
- K. D. Brune, D. B. Leneghan, I. J. Brian, A. S. Ishizuka, M. F. Bachmann, S. J. Draper, S. Biswas and M. Howarth, Sci. Rep., 2016, 6, 19234 CrossRef CAS PubMed.
- P. J. M. Brouwer, A. Antanasijevic, Z. Berndsen, A. Yasmeen, B. Fiala, T. P. L. Bijl, I. Bontjer, J. B. Bale, W. Sheffler, J. D. Allen, A. Schorcht, J. A. Burger, M. Camacho, D. Ellis, C. A. Cottrell, A.-J. Behrens, M. Catalano, I. del Moral-Sánchez, T. J. Ketas, C. LaBranche, M. J. van Gils, K. Sliepen, L. J. Stewart, M. Crispin, D. C. Montefiori, D. Baker, J. P. Moore, P. J. Klasse, A. B. Ward, N. P. King and R. W. Sanders, Nat. Commun., 2019, 10, 4272 CrossRef PubMed.
- Q. Luo, C. Hou, Y. Bai, R. Wang and J. Liu, Chem. Rev., 2016, 116, 13571–13632 CrossRef CAS PubMed.
- A. Varanko, S. Saha and A. Chilkoti, Adv. Drug Delivery Rev., 2020, 156, 133–187 CrossRef CAS PubMed.
- P.-S. Huang, S. E. Boyken and D. Baker, Nature, 2016, 537, 320–327 CrossRef CAS PubMed.
- N. P. King, J. B. Bale, W. Sheffler, D. E. McNamara, S. Gonen, T. Gonen, T. O. Yeates and D. Baker, Nature, 2014, 510, 103–108 CrossRef CAS PubMed.
- D. Baker, Protein Sci., 2019, 28, 678–683 CrossRef CAS PubMed.
- H. Sung, J. Ferlay, R. L. Siegel, M. Laversanne, I. Soerjomataram, A. Jemal and F. Bray, CA Cancer J. Clin., 2021, 71, 209–249 CrossRef PubMed.
- R. L. Siegel, K. D. Miller, N. S. Wagle and A. Jemal, CA Cancer J. Clin., 2023, 73, 17–48 CrossRef PubMed.
- T. N. Schumacher and R. D. Schreiber, Science, 2015, 348, 69–74 CrossRef CAS PubMed.
- M.-Z. Jin and W.-L. Jin, Signal Transduction Targeted Ther., 2020, 5, 166 CrossRef PubMed.
- P. Sharma and J. P. Allison, Science, 2015, 348, 56–61 CrossRef CAS PubMed.
- Y. Ye, J. Wang, Q. Hu, G. M. Hochu, H. Xin, C. Wang and Z. Gu, ACS Nano, 2016, 10, 8956–8963 CrossRef CAS PubMed.
- K. A. Hay, Br. J. Haematol., 2018, 183, 364–374 CrossRef CAS PubMed.
- E. Montomoli, S. Piccirella, B. Khadang, E. Mennitto, R. Camerini and A. De Rosa, Expert Rev. Vaccines, 2011, 10, 1053–1061 CrossRef CAS PubMed.
- G. Guillén, J. C. Aguilar, S. Dueñas, L. Hermida, M. G. Guzmán, E. Penton, E. Iglesias, J. Junco, I. Torrens, Y. Lobaina, V. Muzio and L. Herrera, Procedia Vaccinol., 2010, 2, 128–133 CrossRef.
- Y. H. Chung, H. Cai and N. F. Steinmetz, Adv. Drug Delivery Rev., 2020, 156, 214–235 CrossRef CAS PubMed.
- G. G. Pickett and D. S. Peabody, Nucleic Acids Res., 1993, 21, 4621–4626 CrossRef CAS PubMed.
- C. Mao, V. Beiss, J. Fields, N. F. Steinmetz and S. Fiering, Biomaterials, 2021, 275, 120914 CrossRef CAS PubMed.
- T. Storni, C. Ruedl, K. Schwarz, R. A. Schwendener, W. A. Renner and M. F. Bachmann, J. Immunol., 2004, 172, 1777–1785 CrossRef CAS PubMed.
- A. Shahrivarkevishahi, L. M. Hagge, O. R. Brohlin, S. Kumari, R. Ehrman, C. Benjamin and J. J. Gassensmith, Mater. Today Chem., 2022, 24, 100808 CrossRef CAS.
- A. Shahrivarkevishahi, M. A. Luzuriaga, F. C. Herbert, A. C. Tumac, O. R. Brohlin, Y. H. Wijesundara, A. V. Adlooru, C. Benjamin, H. Lee, P. Parsamian, J. Gadhvi, N. J. De Nisco and J. J. Gassensmith, J. Am. Chem. Soc., 2021, 143, 16428–16438 CrossRef CAS PubMed.
- Y. H. Chung, J. Park, H. Cai and N. F. Steinmetz, Adv. Sci., 2021, 8, 2101796 CrossRef CAS PubMed.
- V. Beiss, C. Mao, S. N. Fiering and N. F. Steinmetz, Mol. Pharm., 2022, 19, 1573–1585 CrossRef CAS PubMed.
- P. L. Chariou, V. Beiss, Y. Ma and N. F. Steinmetz, Mater. Adv., 2021, 2, 1644–1656 RSC.
- J. Ou, M. Zhu, X. Ju, D. Xu, G. Lu, K. Li, W. Jiang, C. Wan, Y. Zhao, Y. Han, Y. Tian and Z. Niu, Nano Lett., 2023, 23, 2056–2064 CrossRef CAS PubMed.
- C. I. Nkanga, O. A. Ortega-Rivera and N. F. Steinmetz, Nanomedicine, 2022, 44, 102573 CrossRef CAS PubMed.
- D. Patterson, B. Schwarz, J. Avera, B. Western, M. Hicks, P. Krugler, M. Terra, M. Uchida, K. McCoy and T. Douglas, Bioconjugate Chem., 2017, 28, 2114–2124 CrossRef CAS PubMed.
- D. Botstein, C. H. Waddell and J. King, J. Mol. Biol., 1973, 80, 669–695 CrossRef CAS PubMed.
- W. Li, Z. Jing, S. Wang, Q. Li, Y. Xing, H. Shi, S. Li and Z. Hong, Biomaterials, 2021, 271, 120726 CrossRef CAS PubMed.
- P. Pumpens and E. Grens, Intervirology, 2001, 44, 98–114 CrossRef CAS PubMed.
- M. Pasek, T. Goto, W. Gilbert, B. Zink, H. Schaller, P. MacKay, G. Leadbetter and K. Murray, Nature, 1979, 282, 575–579 CrossRef CAS PubMed.
- Y. Zhao, Z. Li, J. Voyer, Y. Li and X. Chen, ACS Appl. Mater. Interfaces, 2022, 14, 21872–21885 CrossRef CAS PubMed.
- E. M. Plummer and M. Manchester, Wiley Interdiscip. Rev.: Nanomed. Nanobiotechnol., 2011, 3, 174–196 CAS.
- D. Ren, F. Kratz and S.-W. Wang, Small, 2011, 7, 1051–1060 CrossRef CAS PubMed.
- N. M. Molino, A. K. L. Anderson, E. L. Nelson and S.-W. Wang, ACS Nano, 2013, 7, 9743–9752 CrossRef CAS PubMed.
- D. Lin, Y. Shen and T. Liang, Signal Transduction Targeted Ther., 2023, 8, 156 CrossRef PubMed.
- D. G. Roy, K. Geoffroy, M. Marguerie, S. T. Khan, N. T. Martin, J. Kmiecik, D. Bobbala, A. S. Aitken, C. T. de Souza, K. B. Stephenson, B. D. Lichty, R. C. Auer, D. F. Stojdl, J. C. Bell and M. C. Bourgeois-Daigneault, Nat. Commun., 2021, 12, 2626 CrossRef CAS PubMed.
- V. Cerullo, M. Vähä-Koskela and A. Hemminki, OncoImmunology, 2012, 1, 979–981 CrossRef PubMed.
- M. Fusciello, F. Fontana, S. Tähtinen, C. Capasso, S. Feola, B. Martins, J. Chiaro, K. Peltonen, L. Ylösmäki, E. Ylösmäki, F. Hamdan, O. K. Kari, J. Ndika, H. Alenius, A. Urtti, J. T. Hirvonen, H. A. Santos and V. Cerullo, Nat. Commun., 2019, 10, 5747 CrossRef CAS PubMed.
- R. Ma, Z. Li, E. A. Chiocca, M. A. Caligiuri and J. Yu, Trends Cancer, 2023, 9, 122–139 CrossRef CAS PubMed.
- A. R. Yoon, C. Rivera-Cruz, J. M. Gimble, C.-O. Yun and M. L. Figueiredo, Mol. Ther.–Oncolytics, 2022, 25, 78–97 CrossRef CAS PubMed.
- M. Truffi, L. Fiandra, L. Sorrentino, M. Monieri, F. Corsi and S. Mazzucchelli, Pharmacol. Res., 2016, 107, 57–65 CrossRef CAS PubMed.
- Y. Hu, D. Chen, S. Park, T. Emrick and T. P. Russell, Adv. Mater., 2010, 22, 2583–2587 CrossRef CAS PubMed.
- G. Deng, Y. Li, N. Liang, P. Hu, Y. Zhang, L. Qiao, Y. Zhang, J. Xie, H. Luo, F. Wang, F. Chen, F. Liu, D. Xu and J. Zhang, Cancer Med., 2023, 12, 11570–11588 CrossRef CAS PubMed.
- P. Arosio, R. Ingrassia and P. Cavadini, Biochim. Biophys. Acta, Gen. Subj., 2009, 1790, 589–599 CrossRef CAS PubMed.
- G. Jutz, P. van Rijn, B. Santos Miranda and A. Böker, Chem. Rev., 2015, 115, 1653–1701 CrossRef CAS PubMed.
- M. C. Linder, H. R. Kakavandi, P. Miller, P. L. Wirth and G. M. Nagel, Arch. Biochem. Biophys., 1989, 269, 485–496 CrossRef CAS PubMed.
- X. Liu, W. Jin and E. C. Theil, Proc. Natl. Acad. Sci. U. S. A., 2003, 100, 3653–3658 CrossRef CAS PubMed.
- S. Yin, K. Davey, S. Dai, Y. Liu and J. Bi, Particuology, 2022, 64, 65–84 CrossRef CAS.
- K. Fan, C. Cao, Y. Pan, D. Lu, D. Yang, J. Feng, L. Song, M. Liang and X. Yan, Nat. Nanotechnol., 2012, 7, 459–464 CrossRef CAS PubMed.
- A. E. Powell, K. Zhang, M. Sanyal, S. Tang, P. A. Weidenbacher, S. Li, T. D. Pham, J. E. Pak, W. Chiu and P. S. Kim, ACS Cent. Sci., 2021, 7, 183–199 CrossRef CAS PubMed.
- Z. Zhen, W. Tang, C. Guo, H. Chen, X. Lin, G. Liu, B. Fei, X. Chen, B. Xu and J. Xie, ACS Nano, 2013, 7, 6988–6996 CrossRef CAS PubMed.
- G. Fracasso, E. Falvo, G. Colotti, F. Fazi, T. Ingegnere, A. Amalfitano, G. B. Doglietto, S. Alfieri, A. Boffi, V. Morea, G. Conti, E. Tremante, P. Giacomini, A. Arcovito and P. Ceci, J. Controlled Release, 2016, 239, 10–18 CrossRef CAS PubMed.
- E. Falvo, E. Tremante, R. Fraioli, C. Leonetti, C. Zamparelli, A. Boffi, V. Morea, P. Ceci and P. Giacomini, Nanoscale, 2013, 5, 12278 RSC.
- M. Liu, Y. Zhu, T. Wu, J. Cheng and Y. Liu, Chem. – Eur. J., 2020, 26, 7442–7450 CrossRef CAS PubMed.
- L. N. Turino, M. R. Ruggiero, R. Stefanìa, J. C. Cutrin, S. Aime and S. Geninatti Crich, Bioconjugate Chem., 2017, 28, 1283–1290 CrossRef CAS PubMed.
- L. Conti, S. Ciambellotti, G. E. Giacomazzo, V. Ghini, L. Cosottini, E. Puliti, M. Severi, E. Fratini, F. Cencetti, P. Bruni, B. Valtancoli, C. Giorgi and P. Turano, Inorg. Chem. Front., 2022, 9, 1070–1081 RSC.
- M. S. Lesniak and H. Brem, Nat. Rev. Drug Discovery, 2004, 3, 499–508 CrossRef CAS PubMed.
- C. D. Arvanitis, G. B. Ferraro and R. K. Jain, Nat. Rev. Cancer, 2020, 20, 26–41 CrossRef CAS PubMed.
- L. You, J. Wang, T. Liu, Y. Zhang, X. Han, T. Wang, S. Guo, T. Dong, J. Xu, G. J. Anderson, Q. Liu, Y.-Z. Chang, X. Lou and G. Nie, ACS Nano, 2018, 12, 4123–4139 CrossRef CAS PubMed.
- O. Khorkova and C. Wahlestedt, Nat. Biotechnol., 2017, 35, 249–263 CrossRef CAS PubMed.
- P. Y. Collins, V. Patel, S. S. Joestl, D. March, T. R. Insel, A. S. Daar, I. A. Bordin, E. J. Costello, M. Durkin, C. Fairburn, R. I. Glass, W. Hall, Y. Huang, S. E. Hyman, K. Jamison, S. Kaaya, S. Kapur, A. Kleinman, A. Ogunniyi, A. Otero-Ojeda, M.-M. Poo, V. Ravindranath, B. J. Sahakian, S. Saxena, P. A. Singer, D. J. Stein, W. Anderson, M. A. Dhansay, W. Ewart, A. Phillips, S. Shurin and M. Walport, Nature, 2011, 475, 27–30 CrossRef CAS PubMed.
- K. Fan, X. Jia, M. Zhou, K. Wang, J. Conde, J. He, J. Tian and X. Yan, ACS Nano, 2018, 12, 4105–4115 CrossRef CAS PubMed.
- M. Liang, K. Fan, M. Zhou, D. Duan, J. Zheng, D. Yang, J. Feng and X. Yan, Proc. Natl. Acad. Sci. U. S. A., 2014, 111, 14900–14905 CrossRef CAS PubMed.
- F. Palombarini, E. Di Fabio, A. Boffi, A. Macone and A. Bonamore, Molecules, 2020, 25, 825 CrossRef CAS PubMed.
- Z. Yuan, B. Wang, Y. Teng, W. Ho, B. Hu, K. O. Boakye-Yiadom, X. Xu and X.-Q. Zhang, Nanoscale, 2022, 14, 6449–6464 RSC.
- B. Wang, M. Tang, Z. Yuan, Z. Li, B. Hu, X. Bai, J. Chu, X. Xu and X.-Q. Zhang, Bioact. Mater., 2022, 16, 232–248 CAS.
- Y. Choi, G.-H. Nam, G. B. Kim, S. Kim, Y. K. Kim, S. A. Kim, H.-J. Kim, E. J. Lee and I.-S. Kim, Cancer Gene Ther., 2021, 28, 960–970 CrossRef CAS PubMed.
- B. Sangro, P. Sarobe, S. Hervás-Stubbs and I. Melero, Nat. Rev. Gastroenterol. Hepatol., 2021, 18, 525–543 CrossRef PubMed.
- Y. Chen, L. Zeng, H. Zhu, Q. Wu, R. Liu, Q. Liang, B. Chen, H. Dai, K. Tang, C. Liao, Y. Huang, X. Yan, K. Fan, J.-Z. Du, R. Lin and J. Wang, Small Methods, 2023, 7, 2201086 CrossRef CAS PubMed.
- M. Liu, D. Jin, W. Yu, J. Yu, K. Cao, J. Cheng, X. Zheng, A. Wang and Y. Liu, Adv. Sci., 2024, 2308248 CrossRef CAS PubMed.
- M. T. Larsen, M. Kuhlmann, M. L. Hvam and K. A. Howard, Mol. Cell. Ther., 2016, 4, 3 CrossRef PubMed.
- F. Kratz, J. Controlled Release, 2008, 132, 171–183 CrossRef CAS PubMed.
- D. Sleep, J. Cameron and L. R. Evans, Biochim. Biophys. Acta, Gen. Subj., 2013, 1830, 5526–5534 CrossRef CAS PubMed.
- A. O. Elzoghby, W. M. Samy and N. A. Elgindy, J. Controlled Release, 2012, 157, 168–182 CrossRef CAS PubMed.
- E. S. Lee and Y. S. Youn, J. Pharm. Invest., 2016, 46, 305–315 CrossRef CAS.
- E. N. Hoogenboezem and C. L. Duvall, Adv. Drug Delivery Rev., 2018, 130, 73–89 CrossRef CAS PubMed.
- R. Xu, M. Fisher and R. L. Juliano, Bioconjugate Chem., 2011, 22, 870–878 CrossRef CAS PubMed.
- T. Lin, P. Zhao, Y. Jiang, Y. Tang, H. Jin, Z. Pan, H. He, V. C. Yang and Y. Huang, ACS Nano, 2016, 10, 9999–10012 CrossRef CAS PubMed.
- W.-W. Qi, H.-Y. Yu, H. Guo, J. Lou, Z.-M. Wang, P. Liu, A. Sapin-Minet, P. Maincent, X.-C. Hong, X.-M. Hu and Y.-L. Xiao, Mol. Pharm., 2015, 12, 675–683 CrossRef CAS PubMed.
- J. H. Lee, H. Moon, H. Han, I. J. Lee, D. Kim, H. J. Lee, S. W. Ha, H. Kim and J. W. Chung, Cancers, 2019, 11, 581 CrossRef CAS PubMed.
- W. K. Nevala, S. A. Buhrow, D. J. Knauer, J. M. Reid, E. A. Atanasova and S. N. Markovic, Cancer Res., 2016, 76, 3954–3964 CrossRef CAS PubMed.
- D. Chu, Q. Zhao, J. Yu, F. Zhang, H. Zhang and Z. Wang, Adv. Healthcare Mater., 2016, 5, 1088–1093 CrossRef CAS PubMed.
- L. Feng, L. Yang, L. Li, J. Xiao, N. Bie, C. Xu, J. Zhou, H. Liu, L. Gan and Y. Wu, Nano Res., 2021, 15, 593–602 CrossRef.
- J. Yang, W. Guo, R. Huang, J. Bian, S. Zhang, T. Wei, C. He, Z. Hu, J. Li, C. Zhou and M. Lu, Front. Immunol., 2023, 14, 1173487 CrossRef CAS PubMed.
- D. Cen, Q. Ge, C. Xie, Q. Zheng, J. Guo, Y. Zhang, Y. Wang, X. Li, Z. Gu and X. Cai, Adv. Mater., 2021, 33, 2104037 CrossRef CAS PubMed.
- A. Zheng, Z. Ning, X. Wang, Z. Li, Y. Sun, M. Wu, D. Zhang, X. Liu, J. Chen and Y. Zeng, Mater. Today Bio, 2024, 25, 100955 CrossRef CAS PubMed.
- C. Zhang, W. Chen, T. Zhang, X. Jiang and Y. Hu, J. Mater. Chem. B, 2020, 8, 4726–4737 RSC.
- Z. Zhou, J. Chen, Y. Liu, C. Zheng, W. Luo, L. Chen, S. Zhou, Z. Li and J. Shen, Acta Pharm. Sin. B, 2022, 12, 4204–4223 CrossRef CAS PubMed.
- O. Karroum, J. Kengen, P. Danhier, J. Magat, L. Mignion, C. Bouzin, J. Verrax, N. Charette, P. Starkel, P. B. Calderon, P. Sonveaux, O. Feron, V. Grégoire, B. Gallez and B. F. Jordan, Radiother. Oncol., 2012, 105, 64–71 CrossRef CAS PubMed.
- W.-S. Ryu, in Molecular Virology of Human Pathogenic Viruses, 2017, pp. 195–211, DOI:10.1016/b978-0-12-800838-6.00015-1.
- M. Sharma and B. Sood, World J. Microbiol. Biotechnol., 2010, 27, 471–477 CrossRef.
- L. Deng and B.-Z. Wang, ACS Infect. Dis., 2018, 4, 1656–1665 CrossRef CAS PubMed.
- W. Sun, Y. Liu, F. Amanat, I. González-Domínguez, S. McCroskery, S. Slamanig, L. Coughlan, V. Rosado, N. Lemus, S. Jangra, R. Rathnasinghe, M. Schotsaert, J. L. Martinez, K. Sano, I. Mena, B. L. Innis, P. Wirachwong, D. H. Thai, R. D. N. Oliveira, R. Scharf, R. Hjorth, R. Raghunandan, F. Krammer, A. García-Sastre and P. Palese, Nat. Commun., 2021, 12, 6197 CrossRef PubMed.
- S. Deng, Y. Liu, R. C.-Y. Tam, P. Chen, A. J. Zhang, B. W.-Y. Mok, T. Long, A. Kukic, R. Zhou, H. Xu, W. Song, J. F.-W. Chan, K. K.-W. To, Z. Chen, K.-Y. Yuen, P. Wang and H. Chen, Nat. Commun., 2023, 14, 2081 CrossRef CAS PubMed.
- A. I. Makarkov, M. Golizeh, E. Ruiz-Lancheros, A. A. Gopal, I. N. Costas-Cancelas, S. Chierzi, S. Pillet, N. Charland, N. Landry, I. Rouiller, P. W. Wiseman, M. Ndao and B. J. Ward, npj Vaccines, 2019, 4, 17 CrossRef PubMed.
- B. J. Davenport, A. Catala, S. M. Weston, R. M. Johnson, J. Ardanuy, H. L. Hammond, C. Dillen, M. B. Frieman, C. E. Catalano and T. E. Morrison, npj Vaccines, 2022, 7, 57 CrossRef CAS PubMed.
- S. K. Chan, P. Du, C. Ignacio, S. Mehta, I. G. Newton and N. F. Steinmetz, ACS Nano, 2020, 15, 1259–1272 CrossRef PubMed.
- P. A. B. Weidenbacher, M. Sanyal, N. Friedland, S. Tang, P. S. Arunachalam, M. Hu, O. S. Kumru, M. K. Morris, J. Fontenot, L. Shirreff, J. Do, Y.-C. Cheng, G. Vasudevan, M. B. Feinberg, F. J. Villinger, C. Hanson, S. B. Joshi, D. B. Volkin, B. Pulendran and P. S. Kim, Nat. Commun., 2023, 14, 2149 CrossRef CAS PubMed.
- J. Pan, Q. Wang, M. Qi, J. Chen, X. Wu, X. Zhang, W. Li, X.-E. Zhang and Z. Cui, ACS Nano, 2023, 17, 13474–13487 CrossRef CAS PubMed.
- X. Ma, F. Zou, F. Yu, R. Li, Y. Yuan, Y. Zhang, X. Zhang, J. Deng, T. Chen, Z. Song, Y. Qiao, Y. Zhan, J. Liu, J. Zhang, X. Zhang, Z. Peng, Y. Li, Y. Lin, L. Liang, G. Wang, Y. Chen, Q. Chen, T. Pan, X. He and H. Zhang, Immunity, 2020, 53, 1315–1330 CrossRef CAS PubMed.
- M. G. Joyce, W.-H. Chen, R. S. Sankhala, A. Hajduczki, P. V. Thomas, M. Choe, E. J. Martinez, W. C. Chang, C. E. Peterson, E. B. Morrison, C. Smith, R. E. Chen, A. Ahmed, L. Wieczorek, A. Anderson, J. B. Case, Y. Li, T. Oertel, L. Rosado, A. Ganesh, C. Whalen, J. M. Carmen, L. Mendez-Rivera, C. P. Karch, N. Gohain, Z. Villar, D. McCurdy, Z. Beck, J. Kim, S. Shrivastava, O. Jobe, V. Dussupt, S. Molnar, U. Tran, C. B. Kannadka, S. Soman, C. Kuklis, M. Zemil, H. Khanh, W. Wu, M. A. Cole, D. K. Duso, L. W. Kummer, T. J. Lang, S. E. Muncil, J. R. Currier, S. J. Krebs, V. R. Polonis, S. Rajan, P. M. McTamney, M. T. Esser, W. W. Reiley, M. Rolland, N. de Val, M. S. Diamond, G. D. Gromowski, G. R. Matyas, M. Rao, N. L. Michael and K. Modjarrad, Cell Rep., 2021, 37, 110143 CrossRef CAS PubMed.
- H. Shen, E. M. Lynch, S. Akkineni, J. L. Watson, J. Decarreau, N. P. Bethel, I. Benna, W. Sheffler, D. Farrell, F. DiMaio, E. Derivery, J. J. De Yoreo, J. Kollman and D. Baker, Nat. Nanotechnol., 2024 DOI:10.1038/s41565-024-01641-1.
- A. J. Wargacki, T. P. Wörner, M. van de Waterbeemd, D. Ellis, A. J. R. Heck and N. P. King, Nat. Commun., 2021, 12, 883 CrossRef CAS PubMed.
- Y. Hsia, J. B. Bale, S. Gonen, D. Shi, W. Sheffler, K. K. Fong, U. Nattermann, C. Xu, P.-S. Huang, R. Ravichandran, S. Yi, T. N. Davis, T. Gonen, N. P. King and D. Baker, Nature, 2016, 535, 136–139 CrossRef CAS PubMed.
- G. L. Butterfield, M. J. Lajoie, H. H. Gustafson, D. L. Sellers, U. Nattermann, D. Ellis, J. B. Bale, S. Ke, G. H. Lenz, A. Yehdego, R. Ravichandran, S. H. Pun, N. P. King and D. Baker, Nature, 2017, 552, 415–420 CrossRef CAS PubMed.
- J. B. Bale, S. Gonen, Y. Liu, W. Sheffler, D. Ellis, C. Thomas, D. Cascio, T. O. Yeates, T. Gonen, N. P. King and D. Baker, Science, 2016, 353, 389–394 CrossRef CAS PubMed.
- P. J. M. Brouwer, A. Antanasijevic, Z. Berndsen, A. Yasmeen, B. Fiala, T. P. L. Bijl, I. Bontjer, J. B. Bale, W. Sheffler, J. D. Allen, A. Schorcht, J. A. Burger, M. Camacho, D. Ellis, C. A. Cottrell, A.-J. Behrens, M. Catalano, I. del Moral-Sánchez, T. J. Ketas, C. LaBranche, M. J. van Gils, K. Sliepen, L. J. Stewart, M. Crispin, D. C. Montefiori, D. Baker, J. P. Moore, P. J. Klasse, A. B. Ward, N. P. King and R. W. Sanders, Nat. Commun., 2019, 10, 4272 CrossRef PubMed.
- J. Marcandalli, B. Fiala, S. Ols, M. Perotti, W. de van der Schueren, J. Snijder, E. Hodge, M. Benhaim, R. Ravichandran, L. Carter, W. Sheffler, L. Brunner, M. Lawrenz, P. Dubois, A. Lanzavecchia, F. Sallusto, K. K. Lee, D. Veesler, C. E. Correnti, L. J. Stewart, D. Baker, K. Loré, L. Perez and N. P. King, Cell, 2019, 176, 1420–1431 CrossRef CAS PubMed.
- K. Sliepen, L. Radić, J. Capella-Pujol, Y. Watanabe, I. Zon, A. Chumbe, W.-H. Lee, M. de Gast, J. Koopsen, S. Koekkoek, I. del Moral-Sánchez, P. J. M. Brouwer, R. Ravichandran, G. Ozorowski, N. P. King, A. B. Ward, M. J. van Gils, M. Crispin, J. Schinkel and R. W. Sanders, Nat. Commun., 2022, 13, 7271 CrossRef CAS PubMed.
- A. H. Ellebedy, F. Krammer, G.-M. Li, M. S. Miller, C. Chiu, J. Wrammert, C. Y. Chang, C. W. Davis, M. McCausland, R. Elbein, S. Edupuganti, P. Spearman, S. F. Andrews, P. C. Wilson, A. García-Sastre, M. J. Mulligan, A. K. Mehta, P. Palese and R. Ahmed, Proc. Natl. Acad. Sci. U. S. A., 2014, 111, 13133–13138 CrossRef CAS PubMed.
- S. Boyoglu-Barnum, D. Ellis, R. A. Gillespie, G. B. Hutchinson, Y.-J. Park, S. M. Moin, O. J. Acton, R. Ravichandran, M. Murphy, D. Pettie, N. Matheson, L. Carter, A. Creanga, M. J. Watson, S. Kephart, S. Ataca, J. R. Vaile, G. Ueda, M. C. Crank, L. Stewart, K. K. Lee, M. Guttman, D. Baker, J. R. Mascola, D. Veesler, B. S. Graham, N. P. King and M. Kanekiyo, Nature, 2021, 592, 623–628 CrossRef CAS PubMed.
- A. C. Walls, B. Fiala, A. Schäfer, S. Wrenn, M. N. Pham, M. Murphy, L. V. Tse, L. Shehata, M. A. O'Connor, C. Chen, M. J. Navarro, M. C. Miranda, D. Pettie, R. Ravichandran, J. C. Kraft, C. Ogohara, A. Palser, S. Chalk, E. C. Lee, K. Guerriero, E. Kepl, C. M. Chow, C. Sydeman, E. A. Hodge, B. Brown, J. T. Fuller, K. H. Dinnon, L. E. Gralinski, S. R. Leist, K. L. Gully, T. B. Lewis, M. Guttman, H. Y. Chu, K. K. Lee, D. H. Fuller, R. S. Baric, P. Kellam, L. Carter, M. Pepper, T. P. Sheahan, D. Veesler and N. P. King, Cell, 2020, 183, 1367–1382 CrossRef CAS PubMed.
- J. Y. Song, W. S. Choi, J. Y. Heo, J. S. Lee, D. S. Jung, S.-W. Kim, K.-H. Park, J. S. Eom, S. J. Jeong, J. Lee, K. T. Kwon, H. J. Choi, J. W. Sohn, Y. K. Kim, J. Y. Noh, W. J. Kim, F. Roman, M. A. Ceregido, F. Solmi, A. Philippot, A. C. Walls, L. Carter, D. Veesler, N. P. King, H. Kim, J. H. Ryu, S. J. Lee, Y. W. Park, H. K. Park and H. J. Cheong, eClinicalMedicine, 2022, 51, 101569 CrossRef PubMed.
- G. Cappellano, H. Abreu, C. Casale, U. Dianzani and A. Chiocchetti, Vaccines, 2021, 9, 606 CrossRef CAS PubMed.
- Q.-V. Le, J. Choi and Y.-K. Oh, J. Pharm. Invest., 2018, 48, 527–539 CrossRef CAS.
- S. A. Rosenberg, N. P. Restifo, J. C. Yang, R. A. Morgan and M. E. Dudley, Nat. Rev. Cancer, 2008, 8, 299–308 CrossRef CAS PubMed.
- S. Yang, I.-C. Sun, H. S. Hwang, M. K. Shim, H. Y. Yoon and K. Kim, J. Mater. Chem. B, 2021, 9, 3983–4001 RSC.
- M. Sharma and B. Sood, World J. Microbiol. Biotechnol., 2011, 27, 471–477 CrossRef CAS.
- M. Shahgolzari, H. Dianat-Moghadam and S. Fiering, Semin. Cancer Biol., 2022, 86, 1076–1085 CrossRef CAS PubMed.
- G. K. Mutwiri, A. K. Nichani, S. Babiuk and L. A. Babiuk, J. Controlled Release, 2004, 97, 1–17 CrossRef CAS PubMed.
- G. Deng, Z. Sun, S. Li, X. Peng, W. Li, L. Zhou, Y. Ma, P. Gong and L. Cai, ACS Nano, 2018, 12, 12096–12108 CrossRef CAS PubMed.
- N. Rabiee, M. R. Dokmeci, A. Zarrabi, P. Makvandi, M. R. Saeb, H. Karimi-Maleh, S. Jafarzadeh, C. Karaman, Y. Yamauchi, M. E. Warkiani, S. A. Bencherif, G. Mehta, M. Eguchi, A. Kaushik, M.-A. Shahbazi, A. C. Paiva-Santos, J. Ryl, E. C. Lima, M. R. Hamblin, R. S. Varma, Y. Huh, A. T. E. Vilian, P. K. Gupta, S. K. Lakhera, K. K. Kesari, Y.-T. Liu, M. Tahriri, G. S. Rama Raju, M. Adeli, A. Mohammadi, J. Wang, M. Z. Ansari, T. Aminabhavi, H. Savoji, G. Sethi, T. Bączek, A. Kot-Wasik, M. E. Penoff, A. M. Nafchi, J. Kucinska-Lipka, M. Zargar, M. Asadnia, A. R. Aref, M. Safarkhani, M. Ashrafizadeh, R. Umapathi, A. Ghasemi and M. Radisic, Green Biomater., 2023, 1, 1–4 CrossRef.
- H. Cai, S. Shukla and N. F. Steinmetz, Adv. Funct. Mater., 2020, 30, 1908743 CrossRef CAS PubMed.
- Z. Yin, H. G. Nguyen, S. Chowdhury, P. Bentley, M. A. Bruckman, A. Miermont, J. C. Gildersleeve, Q. Wang and X. Huang, Bioconjugate Chem., 2012, 23, 1694–1703 CrossRef CAS PubMed.
- X. Wu, C. McKay, C. Pett, J. Yu, M. Schorlemer, S. Ramadan, S. Lang, S. Behren, U. Westerlind, M. G. Finn and X. Huang, ACS Chem. Biol., 2019, 14, 2176–2184 CrossRef CAS PubMed.
- C. Yinwen, D. L.-M. Caitlin, W. Wattawan, W. Zhaoming, H. F. C. Carlos, K. S. Aliasger, J. W. George and L. S. Andrean, J. ImmunoTher. Cancer, 2020, 8, e000940 CrossRef PubMed.
- E. Bolli, J. P. O'Rourke, L. Conti, S. Lanzardo, V. Rolih, J. M. Christen, G. Barutello, M. Forni, F. Pericle and F. Cavallo, OncoImmunology, 2018, 7, e1408746 CrossRef PubMed.
- D.-W. Zheng, F. Gao, Q. Cheng, P. Bao, X. Dong, J.-X. Fan, W. Song, X. Zeng, S.-X. Cheng and X.-Z. Zhang, Nat. Commun., 2020, 11, 1985 CrossRef CAS PubMed.
- D. Braeden, A.-B. Farah, J. P. Simon, Y. Katie, K. W. Vernon and L. Y. Sarah, J. ImmunoTher. Cancer, 2017, 5, 69 CrossRef PubMed.
- K. Jemon, V. Young, M. Wilson, S. McKee, V. Ward, M. Baird, S. Young and M. Hibma, PLoS One, 2013, 8, e66866 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2024 |