
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Effect of high pressure homogenization on in vitro digestibility and colon fermentability of pea protein-rich bread designed for elderly consumers†
Martina
Moretton
 ab,
Melania
Casertano
c,
Nicoletta
Pellegrini
cd,
Monica
Anese
d,
Vincenzo
Fogliano
c and
Edoardo
Capuano
*c
ab,
Melania
Casertano
c,
Nicoletta
Pellegrini
cd,
Monica
Anese
d,
Vincenzo
Fogliano
c and
Edoardo
Capuano
*c
aResearch and Innovation Centre, Fondazione Edmund Mach, San Michele all'Adige, Italy
bONFoods - Research and innovation network on food and nutrition Sustainability, Safety and Security - Working ON Foods, Parma, Italy
cFood Quality and Design Group, Wageningen University, Wageningen, The Netherlands
dDepartment of Agricultural, Food, Environmental and Animal Sciences, University of Udine, Italy
First published on 30th September 2024
Abstract
Enrichment of staple foods with proteins can be a solution to tackle protein-energy malnutrition in the elderly. For instance, bread can be enriched with pea proteins that are cheap, sustainable and easily digestible. Non-conventional technologies, such as high pressure homogenization (HPH), can improve the digestibility of plant proteins. To characterize the health functionality of pea-enriched bread, a functional bread tailored to elderly consumers was developed by substituting 5% wheat flour with untreated or HPH-treated pea protein concentrate. Protein digestibility and colon fermentability were assessed by mimicking elderly in vitro gastrointestinal and gut microbiota conditions and compared with adult conditions. Bread reformulation with pea proteins affected physical and chemical properties and produced an increase in hardness, which is one of the key features for the acceptability of bread by the elderly. The highest hardness value was observed for pea protein bread, followed by HPH-treated pea protein bread and wheat bread. In vitro protein digestibility and fermentability were affected by reformulation and by physiological digestive conditions, with lower digestibility under elderly conditions compared to adult ones. The obtained results may contribute to a better understanding of food digestibility under different gastrointestinal conditions and its dependence on physiological and formulation factors, and ultimately would help to design age-tailored foods.
1 Introduction
The world's population is aging at an unprecedented rate. In many countries, the number of the elderly is growing faster than the overall population1 (WHO, 2002a). The increase in the elderly population has created a need for additional knowledge of age-related changes and nutrition needs, which has importance in the prevention and treatment of malnutrition and disease.2,3 In particular, protein-energy malnutrition, resulting from reduced and/or unbalanced food protein intake and impaired metabolism is quite common in the elderly, who do not introduce the recommended protein amount (i.e., 1.1 g per kg body weight per day).4 It is a matter of fact that a state of malnutrition in the elderly increases the risk of chronic diseases, such as sarcopenia, and frailty, worsens geriatric syndromes and important functional abilities, such as immunity, bone health, and cognitive functions, and is associated with longer hospital stays and high demands of medical services.5,6Increasing protein intake in the elderly is challenging. Increasing portion sizes or meal frequency is usually not a successful strategy in the elderly, because of eating fatigue or low appetite.7 Therefore, there is a need for new food product development with functional health benefits specifically designed to address the food-related needs of elderly consumers.8 In line with this, protein enrichment of staple foods can represent a strategy to increase protein intake in the elderly.9,10 Bread is an important food in the daily diet of the elderly, especially in the Mediterranean area.11 It is generally produced with soft wheat flour, but proteins can be added to increase its nutritional value.12 Pea proteins have been gaining interest in the development of functional foods because they are cheap, sustainable, quite well-balanced in terms of amino acid profile, despite the admitted deficiency in sulphur amino acids in legumes, and easily digestible. Based on high digestibility, and balanced amino acid composition, pea protein may be considered as a high-quality plant protein.13,14 It is worthy to note that pea proteins aggregate in the stomach producing a kinetics of amino acid absorption in the small intestine that is intermediate between that of whey, considered fast-digestible proteins, and caseins, considered slow-digestible proteins.15 To prevent or treat malnutrition in the elderly both aspects should be tackled: food enrichment with a significant protein amount per portion and quality of proteins that should be rich in essential amino acids and fast to digest. To this aim, the application of non-conventional technologies has been suggested to improve vegetable protein digestibility. High pressure homogenization (HPH) has been recently proposed as an effective tool to induce structural changes in proteins, enhancing their functional and nutritional properties. Melchior et al.16 demonstrated that HPH induced protein structural modifications. Because of HPH application, unfolded protein resulted in more easily accessible by the digestive enzymes during the gastric phase, with increased gastric digestion kinetics.
Currently, one of the main hurdles in the development of foods for the elderly is represented by the limited methodologies available to assess their effective functionality. The nutritional assessment should be conducted considering the influence of gastrointestinal (GI) modifications in the elderly, which could decrease the ability to digest and absorb nutrients due to changes in salivary flow, chewing ability, digestive enzyme production, gastric acid secretion.3 The composition and function of the gut microbiota also change with age, potentially contributing to unhealthy aging and the onset of various age-related diseases. Preserving a healthy gut microbiota offers a promising avenue for promoting healthy aging.17 In this sense, in vitro digestion and fermentation models could represent a valuable tool to pre-set the most promising technological interventions enabling the development of age-tailored foods.18 The relationship between technological intervention and nutrient digestibility under elderly GI conditions has been investigated recently,12,19–21 while during colonic fermentation is still largely unexplored and mostly focused on adult GI conditions.22
In this context, a functional bread tailored to elderly consumers was developed by substituting 5% wheat flour with untreated or HPH-treated pea protein concentrate. This percentage was chosen to allow, for the functional bread, the claim “source of protein” (Reg. EU No 1924/2006). This claim is allowed when at least 12% of the total energy of the food is provided by proteins. Nutritional quality of the developed bread was assessed by mimicking elderly in vitro GI and gut microbiota conditions and results were compared with those obtained by applying protocols simulating adult conditions.
2 Material and methods
2.1 Materials
Wheat flour, sunflower oil, dry yeast, sugar, and salt were purchased from a local supermarket (Wageningen, the Netherlands). Pea protein concentrate (80% protein, 8% fat, 4.9% carbohydrates, 4.2% fiber, 1.1% salt, 1.8% moisture, from Raab Vitalfood GmbH, Rohrbach, Germany) was acquired from a local supermarket (Udine, Italy). All ingredients were used for breadmaking. Pepsin (P6887) from porcine gastric mucosa (≥3200 units per mg protein), pancreatin (P7545) from porcine pancreas (8 × USP), porcine bile extract (B8631), ammonium carbonate ((NH4)2CO3), sodium bicarbonate (NaHCO3), potassium chloride (KCl), sodium chloride (NaCl), calcium chloride dihydrate (CaCl2(H2O)2), magnesium chloride hexahydrate (MgCl2(H2O)6), potassium dihydrogen phosphate (KH2PO4), potassium hydrogen phosphate (K2HPO4), sodium hydroxide (NaOH), hydrochloric acid (HCl), o-phthaldialdehyde (OPA), dithiothreithol, sodium dodecyl sulfate (SDS), L-serine, trichloroacetic acid (TCA), sodium tetraborate, sodium thioglucolate, 2-ethylbutyric acid, oxalic acid, acetic acid, propionic acid, butyric acid, valeric acid, iso-butyric acid, isovaleric acid, tryptophan, tryptamine, serotonin, indole-3-lactic acid, oxindole, indole-3-aldehyde, kynurenine, D-methionine, rhodamine B and fluorescein-5-isothiocyanate (FITC) were provided by Sigma-Aldrich (St Louis MO, USA). Pierce™ BCA Protein Assay Kit, NuPAGE®LDS, NuPAGE® MES SDS Running Buffer, NuPAGE® Novex® 4–12% Bis-Tris Gel 1.0 mm were acquired from Thermo Scientific Inc. (Massachusetts, USA). Coomassie Brilliant Blue R-250 Staining Solution was purchased from Bio-Rad (Hercules, CA, USA). BlueRay Prestained Protein Marker was purchased from Jena Bioscience GmbH (Jena, Germany). Ammonia Assay Kit (AMIAR) was provided by Megazyme (Bray, Co. Wicklow, Ireland). Ultra-pure water (Purelab® Flex Pure Water Systems, Veolia Water Technologies Netherlands B.V., Ede, The Netherlands) was used for all analyses.2.2 High pressure homogenization (HPH) treatment
Pea protein concentrate was suspended in water at 5% (w/v) concentration and stirred overnight. The protein aqueous dispersion was then treated at a pressure of 70 MPa for 3 passes by a continuous lab-scale high-pressure homogenizer (Panda Plus 2000; GEA Niro Soavi, Parma, Italy) as reported by Melchior et al.16 Samples were freeze-dried (Epsilon 2–4 LSCplus, Martin Christ GmbH, Osterode am Harz, Germany) and stored into a desiccator at room temperature before breadmaking.2.3 Breadmaking
The reference wheat bread was prepared according to the following dough formulation: 60.5% wheat flour, 30.0% water, 5.9% sunflower oil, 2.0% sugar, 1.0% yeast and 0.6% salt. Two pea protein-rich breads were obtained by substituting wheat flour with 5% w/w of either pea protein or HPH-treated pea protein concentrate while maintaining a constant ratio among the other ingredients.12Before mixing with other ingredients, pea protein and HPH-treated pea protein concentrates were preliminarily suspended in water and stirred overnight at room temperature to allow protein hydration. All ingredients were mixed in a kneading machine (Hobart, Spronk Bakkerijmachines, Andelst, The Netherlands) at speed 3 for 15 min, until the dough was completely stuck around the mixing hook, leaving the mixing bowl completely clean. Subsequently, dough portions (100 g) were manually rounded, and leavened at 35 °C and 80 RH% for 60 min (Weiss, Tiel, The Netherlands). The fermented dough was baked in a professional oven (Leventi Bakermat Mastermind, Spronk Bakkerijmachines, Andelst, The Netherlands) at 160 °C for 35 min, with steam injections every 3 min from 0 to 21 min. Finally, bread samples were cooled to room temperature for 1 h. Wheat bread was used as the reference sample. Wheat bread, pea protein bread and HPH-treated pea protein bread were named W, PP and HPH-PP, respectively.
2.3 Chemical and physical characterization of bread
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g at 4 °C (centrifuge 5430 R, Eppendorf, Nijmegen, The Netherlands). The supernatant was drained, and the precipitate was weighed. WHC was calculated as gwater gdry weight−1.
000g at 4 °C (centrifuge 5430 R, Eppendorf, Nijmegen, The Netherlands). The supernatant was drained, and the precipitate was weighed. WHC was calculated as gwater gdry weight−1.
| Hue angle = arctan (b*/a*) | (1) |
:2.2 mm lens (Basler AG, Germany). ImageJ software was used to analyze the porosity of the bread. The highest b-value was changed from 255 to a value between 130 and 178, to isolate the correct area of the pores. The porosity was calculated as the mean of three different pictures for each bread type.
2.4 Bread digestibility
:1 (v/v) with SGF, CaCl2(H2O)2 and pepsin solution, the pH was adjusted with HCl 1 M to 3 under adult conditions and to 4.5 under elderly ones, water was added to maintain the ratio bolo to fluids 1:1 (v/v), and the sample was stirred at 37 °C during 2 or 3 h under adult or elderly conditions. The intestinal phase was carried out by mixing the endpoint of the gastric phase with SIF, CaCl2(H2O)2, pancreatin and bile salts solutions. Then, the pH was adjusted with NaOH 1 M to 7 or 6.6 under adult or elderly conditions, respectively, water was added to maintain the ratio chyme to fluids 1:1 (v/v), and the sample was stirred at 37 °C during 2 or 3 h under adult or elderly settings. In this study, distinct in vitro experiments were conducted, both in triplicate, respectively for the digesta characterization and colonic fermentation. In the case of the digestion procedure, two parallel in vitro procedures were performed one until the end of the gastric phase and the other until the end of the intestinal phase. The blank in vitro digestion was performed substituting bread with water.
000g for 30 min at 4 °C (Eppendorf Centrifuge 5430 R, Eppendorf, Nijmegen, The Netherlands). The in vitro protein digestibility was determined using the OPA spectrophotometric assay.32 OPA (0.04 g mL−1) was dissolved in ethanol and added to an aqueous solution of sodium tetraborate (0.038 g mL−1), dithiothreitol (0.88 mg mL−1), and SDS (1 mg mL−1). Aliquots of 5 μL of the supernatant were mixed with 200 μL of OPA reagent into a 96-well microplate and incubated for 15 min at room temperature in the dark. The absorbance was measured at 340 nm using a multi-mode microplate reader (Spectramax M3, Molecular Devices, San Jose, USA). A calibration curve was obtained from standard L-serine solutions ranging from 0 to 10 mM. In vitro protein digestibility was measured through the degree of hydrolysis, calculated as follows: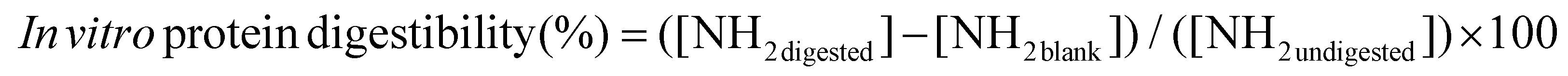 | (2) |
:20 and 0.5 mL of antioxidant. An aliquot of 10 μL of sample was loaded onto NuPAGE® Novex® 4–12% Bis-Tris Gel to guarantee the same protein content in each well. The run was performed on a XCell SureLock™ Mini-Cell apparatus (Thermofisher Scientific, Waltham, Massachusetts, USA) at 150 V for 1 h (VWR 250 V Power Supply, VWR, Radnor, Pennsylvania, USA). Subsequently, the gel was washed with deionized water 3 times, stained with 100 mL Coomassie Brilliant Blue R-250 Staining Solution for 1 h, while shaking slightly, rinsed with deionized water 3 times and washed with washing buffer, containing 10% (v/v) ethanol and 7.5% (v/v) acetic acid in deionized water, for 2 h. The gel was scanned using GS-900 Calibrated Densitometer (Bio-Rad, Hercules, CA, USA) and the image was acquired with Image Lab (Bio-Rad, Hercules, CA, USA). A mixture of proteins with known molecular weights (MW), ranging from 10 to 180 kDa, was used as a reference.
2.5 Bread fermentability
Fresh fecal samples were collected from three European adults, 28–40 years old, and three Dutch elderly, 74–76 years old, with a body mass index of 21.2 ± 1.4 and 20.9 ± 2.2, respectively. All donors were in good health and with no history of gastrointestinal disorders or antibiotic treatment for at least 3 months before this study. Fecal slurries were processed within 2 h after defecation following the method described by Pérez-Burillo et al.34 Healthy volunteers gave written consent for a single fecal donation and their anonymity was maintained. According to the guidelines of the Medical Ethical Advisory Committee of Wageningen University (METC-WU), this research did not need an ethics approval. The fecal slurry was prepared at 32% feces (w/v) in phosphate buffer, consisting of (per L) 8.8 g K2HPO4, 6.8 g KH2PO4 and 0.1 g of sodium thioglycolate in water, using a Stomacher 400 circulator (VWR, Radnor, Pennsylvania, USA). Basal nutrient medium contained (per L): 5.22 g K2HPO4, 16.32 g KH2PO4, 2 g NaHCO3, 2 g yeast extract, 2 g peptone, 1 g mucin, 0.5 g L-cysteine HCl and 2 mL Tween-80. Phosphate buffer, basal nutrient medium and all the materials were autoclaved to sterilize at 121 °C and 1 bar for 15 min. Each fermentation tube carried 5.25 mL of basal nutrient medium, 1.40 mL of fecal slurry, 0.35 g of undigested pellet and 0.45 g of supernatant from in vitro digestion. Digesta pellet and supernatant were thawed immediately before analysis. Earlier addition of fecal slurries, tubes were flushed with N2/CO2 (80/20, v/v) gases to create an anaerobic condition. A control (blank) was included containing only fecal slurry. Samples were kept at 37 °C with oscillating shaking at 200 rpm (MaxQ 2000, Thermofisher Scientific, Waltham, Massachusetts, USA) and taken at 0, 2, 8 and 24 h. Microbial activity was stopped by placing the tubes on ice and after centrifugation at 4500g for 10 min at 4 °C, the supernatants and pellets were separated and stored at −20 °C for further analysis.
000g for 5 min at 4 °C and 2 mL were filtered (15 mm ∅, 0.2 μm regenerated cellulose filter, Phenomenex, Torrance, USA). An internal standard of 2-ethylbutyric acid in 0.3 M HCl and 0.9 M oxalic acid was added to the samples. Nitrogen was used as a carrier gas with a flow rate of 10 mL min−1. The temperature of GC-FID started at 100 °C, then increased to 180 °C for 2 min at a rate of 10.8 °C min−1. Then, it increased at 50 °C min−1 to 240 °C and was maintained at 240 °C for 2 min. Standard calibration curves of acetic, propionic, butyric, valeric, isobutyric and isovaleric acids were prepared in the range from 0 to 2 mg mL−1. The data was processed using Chromeleon 7.2 sr5® software (Thermo Scientific, Waltham, Massachusetts, USA).
000g for 5 min at 4 °C and filtered with 15 mm ∅, 0.2 μm regenerated cellulose filter. Tryptophan-derived catabolites in supernatants were quantified via a Shimadzu Nexera XR LC-20ADxr UPLC system coupled with a Shimadzu LCMS-8050 mass spectrometer (Kyoto, Japan) according to the methodology reported by Huang et al.36 Compounds were identified by comparing the transitions (m/z) and retention time (RT) with reference standards including tryptophan (Trp, m/z 204.9 → 188.1; RT 2.6 min) and catabolite: tryptamine (TA; m/z 161.1 → 144.0; RT 2.0 min), serotonin (5HT; m/z 177.0 → 160.1; RT 1.1 min), indole-3-lactic acid (ILA; m/z 205.9 → 118.1; RT 8.0 min), oxindole (Oxi; m/z 134.0 → 77.1; RT 7.5 min), indole-3-aldehyde (I3A; m/z 146.0 → 118.1; RT 8.4 min) and kynurenine (Kyn; m/z 209.0 → 192.1; RT 1.92 min).
2.6 Statistical analysis
Results are expressed as mean ± standard deviation (SD) of at least three measurements on three replicated samples on a single batch of each bread sample. Statistical analysis was performed by using R v. 4.2.2 for Windows (The R foundation for statistical computing, 2022). Bartlett's test was used to check the homogeneity of variance, one-way ANOVA was carried out and the Tukey test was used to determine statistically significant differences among means (p < 0.05).3 Results and discussion
3.1 Chemical and physical properties of bread
The effect of the protein addition on the chemical and physical properties of bread samples is summarized in Table 1.| W | PP | HPH-PP | |||
|---|---|---|---|---|---|
| a Color parametres: L*, lightness; HA, Hue angle. Values are expressed as mean ± standard deviation (number of repetitions = 3 for protein content, moisture content and aw, 10 for color and texture profiles). Means in the same row indicated by different letters are significantly different (p < 0.05). | |||||
| Protein content (g per 100 gdw) | Crumb | 10.6 ± 0.1b | 13.2 ± 0.3a | 13.6 ± 0.1a | |
| Moisture content (%) | Crust | 20.4 ± 3.4a | 19.0 ± 0.0a | 19.3 ± 0.2a | |
| Crumb | 33.3 ± 0.9b | 35.4 ± 0.7a | 36.1 ± 0.2a | ||
| a w | Crust | 0.90 ± 0.01a | 0.95 ± 0.03a | 0.89 ± 0.00a | |
| Crumb | 0.95 ± 0.01a | 0.96 ± 0.01a | 0.97 ± 0.00a | ||
| WHC (gwater gdry weight−1) | Crumb | 1.1 ± 0.1a | 0.8 ± 0.1a | 1.0 ± 0.1a | |
| Colora | L* | Crust | 50.8 ± 1.0a | 43.7 ± 1.2b | 43.6 ± 0.7b |
| Crumb | 63.2 ± 0.7a | 62.4 ± 1.2ab | 61.4 ± 1.4b | ||
| HA | Crust | 66.9 ± 1.2a | 61.4 ± 1.2b | 61.7 ± 0.7b | |
| Crumb | 86.6 ± 0.2a | 83.8 ± 0.1b | 83.2 ± 0.2c | ||
| Hardness (N) | Crumb | 3.1 ± 0.5c | 7.7 ± 1.9a | 6.2 ± 0.8b | |
| Chewiness (N mm) | Crumb | 1.9 ± 0.2b | 4.2 ± 1.3a | 3.7 ± 0.5a | |
| Porosity (%) | Crumb | 19.1 ± 2.0a | 15.0 ± 3.9a | 14.0 ± 2.9a | |
| Image |

|

|

|
||
The replacement of 5% of wheat flour with PP and HPH-PP concentrates caused an increase in protein content from 10 to 13%, which corresponded to an increase in the energy provided by proteins from 100 kcal g−1 to 130 kcal g−1.12 This allowed the reformulated bread to be claimed as a “source of protein” according to Regulation (EU) No 1924/2006. The presence of pea protein caused an increase in the moisture content of crumb (Table 1), in agreement with the literature.37 Wheat flour substitution with protein concentrate did not modify either aw or WHC values (Table 1).
Color is an important attribute because it influences the elderly's food choices and preferences. As expected, the addition of PP and HPH-PP concentrates significantly decreased L* and HA values of both crumb and crust of bread, as compared to wheat bread (Table 1). This result can be attributed to an increase in Maillard browning reaction with the increase in pea protein content.38,39 No differences in color parameter values (L* and HA) were observed between the protein-enriched samples (p > 0.05).
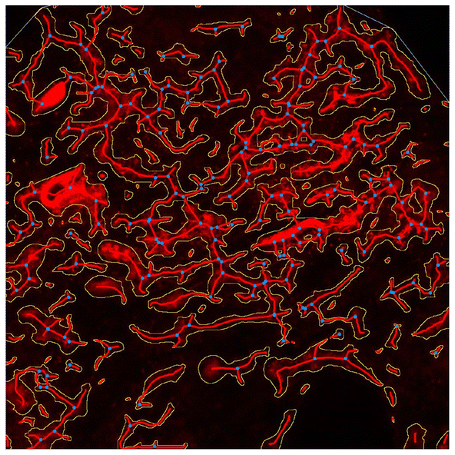
Pea protein addition significantly increased crumb hardness and chewiness as compared to wheat bread (Table 1), in agreement with literature data.37–39 Interestingly, the addition of HPH-PP concentrate led to a slight but significantly reduced crumb hardness in comparison to PP concentrate. Hardness and chewiness represent key features for the acceptability of bread by the elderly, who described their ideal bread as a soft crumb, easy to chew and swallow.40 The increase in hardness and chewiness reveals that bread requires higher teeth force during the first mastication and a longer time and more energy for mastication before swallowing, respectively.41 These results are supported by crumb images, showing a more expanded structure of the control sample as compared with those of the pea-enriched ones without differences in porosity (Table 1). To visualize the changes in the protein network, as affected by the addition of PP and HPH-PP concentrates, a CLSM study was performed (Table 2).
| W | PP | HPH-PP | ||
|---|---|---|---|---|
| Values are expressed as mean ± standard deviation (number of repetitions = 10). Means in the same row indicated by different letters are significantly different (p < 0.05). | ||||
| Protein microstructure | a |

|

|

|
| b |

|

|
|
|
| Average protein length (μm) | 158.1 ± 29.2a | 101.7 ± 24.0b | 162.9 ± 39.9a | |
The images of W bread showed a compact, continuous protein network enclosing starch granules. However, incorporating pea protein into the bread resulted in discernible changes in both the protein network and starch granule structure (Table 2). PP and HPH-PP bread samples exhibited a less interconnected gluten network by increasing sponginess due to the presence of numerous large cavities.37,42 The addition of PP concentrate resulted in a significant reduction in protein length (Table 2). Conversely, there was no change in protein length between the W and HPH-PP bread samples. Compared to the W protein network, the HPH-PP one showed some larger protein aggregates, likely due to protein unfolding caused by the HPH treatment,16 with compromission of the gluten network.
3.2 Bread in vitro protein digestibility
Table 3 shows protein in vitro protein digestibility of W, PP and HPH-PP bread samples at the end of gastric and intestinal phases, under adult and elderly conditions.| Phase | Bread type | In vitro protein digestibility (%) | |
|---|---|---|---|
| Adult | Elderly | ||
| Values are expressed as mean ± standard deviation (number of repetitions = 3). Lower letters indicate statistically significant (p < 0.05) differences between bread types at either the end of gastric or the end of intestinal phase under adult GI conditions. Capital letters indicate statistically significant (p < 0.05) differences between bread types at either the end of the gastric or intestinal phase under the elderly GI conditions. *: indicate statistically significant (p < 0.05) difference between physiological conditions (adult and the elderly) within each bread type. | |||
| Gastric | W | 6.4 ± 0.4b,* | 2.8 ± 1.2A |
| PP | 7.3 ± 0.5a,* | 3.1 ± 0.7A | |
| HPH-PP | 7.5 ± 0.3a,* | 3.0 ± 0.9A | |
| Intestinal | W | 40.8 ± 2.8b,* | 36.9 ± 6.3A |
| PP | 46.3 ± 1.5a,* | 38.4 ± 3.5A | |
| HPH-PP | 48.7 ± 3.9a,* | 38.7 ± 3.3A | |
At the end of the gastric phase, very few peptide bonds were cleaved under both GI conditions (Table 3), in agreement with the literature43 showing that gastric pepsin being an endopeptidase performs few cuts of the polypeptides to achieve protein denaturation. In vitro protein digestibility increased considerably after the intestinal phase: all bread types showed lower in vitro protein digestibility under the elderly GI mimicking conditions than under the adult ones. These results are in agreement with the literature,12,19,21,44 and further confirm that physiological changes during ageing may significantly compromise in vitro protein digestibility.45 Under adult GI conditions, the in vitro protein digestibility of pea protein-enriched bread samples was higher (p < 0.05) than that of wheat bread, and no differences were found between PP and HPH-PP breads. According to the literature, these higher in vitro protein digestibility values can be associated with lower interaction between proteins and starch matrix, making them more accessible to proteases.46 On the contrary, under the elderly conditions, the in vitro protein digestibility values of PP and HPH-PP bread were comparable (p > 0.05) with that of W bread at the end of the intestinal phase (Table 3). It is indeed important to underline that the in vitro digestibility of proteins in the intestinal phase is lower compared to our previous study.12 These results can be attributed to slight methodological differences regarding the oral phase (in vitro vs. in vivo mastication)47 and the treatment the samples underwent (in the previous study both bread and bolus were frozen and thawed) before and after digestion.
Unexpectedly, HPH treatment did not increase (p > 0.05) in vitro protein digestibility of pea protein concentrate-containing bread, both under the adult and elderly GI conditions (Table 3). This finding seems to be in contrast with the results described by Melchior et al.16 for pea protein model systems, showing that shear forces and cavitation effects upon HPH treatment favoured the enzymatic proteolysis, due to the protein unfolding with consequent exposure of SH-groups. In a complex matrix, such as bread, the lack increasing of in vitro protein digestibility of HPH-PP bread compared to that of PP bread (Table 3) can be attributable to the formation of a gluten-pea protein network between the SH-groups of unfolded pea protein and gluten during bread making.42 Therefore, pea proteins would result less available for digestion.
Overall, the slight differences in in vitro protein digestibility observed among samples and under different physiological conditions are confirmed by a few differences found from the particle size distribution and SDS-PAGE profiles of W, PP and HPH-PP bread undigested pellets collected at the end of the gastric and intestinal phases (Fig. 1 and 2).
 | ||
| Fig. 1 Particle size distribution of undigested pellet of wheat (W), pea protein (PP) and HPH-treated pea protein (HPH-PP) bread collected at the end of the gastric under adult (a) and elderly (b) conditions and intestinal phases under the adult (c) and elderly (d) conditions. Average of three consecutive measurements. | ||
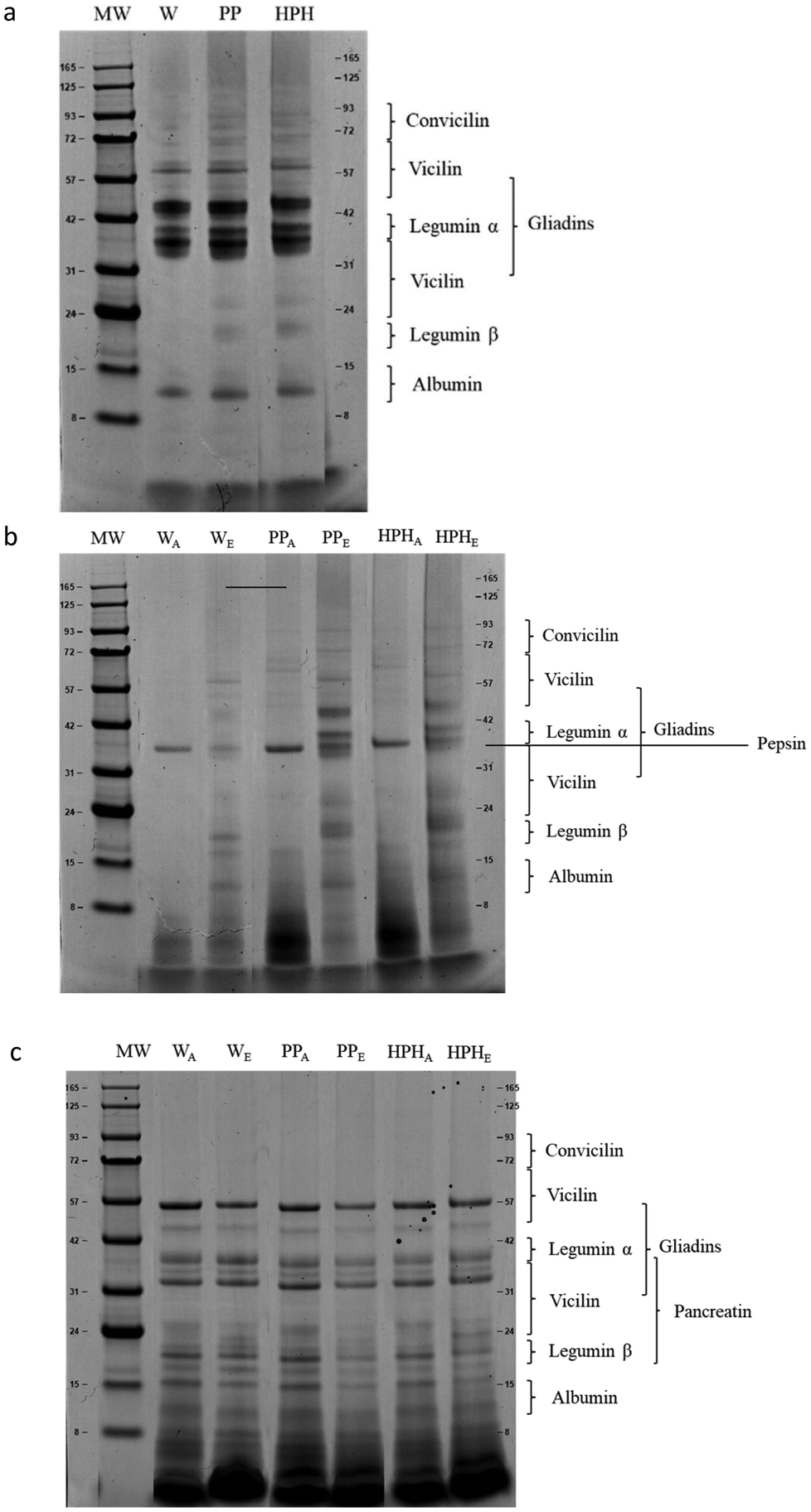 | ||
| Fig. 2 SDS-PAGE of wheat (W), pea protein (PP) and HPH-treated pea protein (HPH-PP) bread before digestion (panel a), at the end of the gastric (panel b) and intestinal (panel c) phases. Letter A indicates adult GI conditions, letter E indicates elderly GI conditions. | ||
Particle size profile (Fig. 1) explained the lack of differences observed in in vitro protein digestibility. Specifically, the pellet of HPH-PP bread exhibited a higher volume of particles in the larger size range (100–1000 μm) compared to other samples at the end of the gastric phase (Fig. 1a and b), confirming that the stronger protein network previously observed (Table 2) is not dissolved during the gastric phase.
SDS-PAGE profile identified the main proteins in bread types (Fig. 2a) in agreement with Laguna et al.48 The application of the HPH treatment did not modify the protein pattern of HPH-PP bread compared to the PP bread. The band distribution changes in intensity with the digestion time of each bread type. At the end of the gastric phase (Fig. 2b) under adult GI conditions, the high molecular weight proteins, including key components such as vicilin, legumin α and gliadins, completely disappeared, which is indicative of proteolysis, and several bands of molecular weight <15 kDa progressively appeared. Among the remaining protein fragments, the low molecular weight forms of vicilin and legumin β exhibited resistance to pepsin during the gastric phase but they were hydrolysed during the intestinal phase. Interestingly, although no differences were observed in in vitro protein digestibility, under adult GI conditions, the intensity of bands <8 kDa of pea protein enriched bread was higher than that of wheat bread. This suggests that, despite similar digestibility, the digestion of pea proteins resulted in larger amounts of low molecular weight peptides. On the other hand, under the elderly condition, the protein degradation pattern differed noticeably. Although all of the protein bands remained present by the end of the gastric phase, their intensity was reduced, indicating that proteolysis was less extensive compared to adult conditions. This could be attributed to the age-related differences in digestive efficiency. A similar trend was previously observed and reported by Melchior et al.31 At the end of the intestinal phase (Fig. 2c), all high molecular weight proteins were digested into smaller fragments, regardless of the bread composition. The resulting digesta profiles were remarkably consistent and showed minimal variation between the adult and elderly GI conditions. The electrophoretic bands between 20 and 50 kDa corresponded to digestive enzymes that were visible across all samples. Additionally, the bands corresponding to the smaller degradation products, specifically those <8 kDa, became significantly more intense, indicating a high degree of protein breakdown into low molecular weight peptides. The results obtained could contribute to better understanding of protein fate under elderly GI conditions and their relation to physiological factors. Preliminary in vitro screening must be validated by in vivo studies that are compulsorily required to understand the actual physiological response.
3.3 In vitro colonic fermentation of bread
After the intestinal phase that mimicked what happens in the small intestine, the pellets contained around 17–20% of dry matter (Table S1†). This value did not significantly differ (p > 0.05) among bread types under adult GI conditions. However, under elderly GI conditions, the dry matter of wheat bread was slightly but significantly lower (p < 0.05) than that of enriched breads. Although more than 97% of proteins initially present in samples were digested, the protein content in the elderly pellets was up to 3 times that of adult ones and enriched bread samples led to a higher content of protein in the adult and elderly pellets, in agreement with the results of the in vitro digestion (Table 3).The undigested pellet of bread types was then subjected to in vitro colonic fermentation to explore the effects of the bread enrichment with pea protein on microbial metabolism. The fermentative activity of the microbiota was monitored by measuring SCFAs and BCFAs (Table 4), which are produced by the gut microbiota by the fermentation of carbohydrates and the reductive deamination of amino acids.49
| Time (h) | Bread type | Butyric acid (mg mL−1) | Acetic acid (mg mL−1) | Propionic acid (mg mL−1) | |||
|---|---|---|---|---|---|---|---|
| Adult | Elderly | Adult | Elderly | Adult | Elderly | ||
| Lower letters indicate a statistically significant (p < 0.05) difference among bread types at different time points using adult batch fermentations. Capital letters indicate a statistically significant (p < 0.05) difference among bread types at different time points using the elderly batch fermentations. | |||||||
| 0 | W | 0.02 ± 0.01a | 0.04 ± 0.01A | 0.37 ± 0.07c | 0.31 ± 0.07D | 0.05 ± 0.02c | 0.05 ± 0.01C |
| PP | 0.02 ± 0.01a | 0.04 ± 0.01A | 0.30 ± 0.05c | 0.31 ± 0.06D | 0.05 ± 0.01c | 0.05 ± 0.01C | |
| HPH-PP | 0.02 ± 0.01a | 0.04 ± 0.01A | 0.40 ± 0.13c | 0.31 ± 0.06D | 0.05 ± 0.01c | 0.05 ± 0.01C | |
| 2 | W | 0.02 ± 0.01a | 0.05 ± 0.03A | 0.44 ± 0.03c | 0.59 ± 0.24C | 0.07 ± 0.00c | 0.25 ± 0.24C |
| PP | 0.02 ± 0.01a | 0.05 ± 0.03A | 0.41 ± 0.02c | 0.61 ± 0.23C | 0.07 ± 0.01c | 0.25 ± 0.23C | |
| HPH-PP | 0.02 ± 0.01a | 0.05 ± 0.03A | 0.53 ± 0.16c | 0.62 ± 0.22C | 0.08 ± 0.02c | 0.26 ± 0.24C | |
| 8 | W | 0.02 ± 0.01a | 0.04 ± 0.01A | 1.91 ± 0.46b | 1.96 ± 0.46B | 0.68 ± 0.04b | 1.55 ± 1.78B |
| PP | 0.02 ± 0.01a | 0.04 ± 0.01A | 2.06 ± 0.74b | 2.06 ± 0.53B | 0.69 ± 0.07b | 1.54 ± 1.79B | |
| HPH-PP | 0.02 ± 0.01a | 0.04 ± 0.01A | 2.39 ± 1.28b | 2.27 ± 0.49B | 0.73 ± 0.12b | 1.76 ± 1.77B | |
| 24 | W | 0.02 ± 0.01a | 0.04 ± 0.01A | 4.14 ± 0.57a | 3.50 ± 1.00A | 3.84 ± 2.02a | 3.97 ± 1.50A |
| PP | 0.02 ± 0.01a | 0.04 ± 0.01A | 4.04 ± 0.78a | 3.35 ± 0.95A | 4.03 ± 2.17a | 3.57 ± 1.93A | |
| HPH-PP | 0.02 ± 0.01a | 0.03 ± 0.01A | 3.95 ± 0.63a | 3.61 ± 1.06A | 3.93 ± 2.05a | 4.14 ± 1.29A | |
Acetic and propionic acids were the two major SCFAs produced during batch fermentations in the 24 h (Table 4). Their concentration increased (p < 0.05) over time, even though no differences (p > 0.05) were detected among bread types as well as between the adult and elderly microbiota conditions, indicating that the type of microbiota (from adult or elderly donors) did not affect SCFAs production.
BCFAs, i.e. isovalerate, isobutyrate, and valerate, were also determined. At the beginning of fermentation (time 0) their concentrations were 0.05 ± 0.02 mg mL−1, 0.01 ± 0.01 mg mL−1, and 0.01 ± 0.01 mg mL−1, respectively, and did not increase over time. Additionally, ammonia production was detected after 24 h of batch fermentation of pellets collected after in vitro digestion of W, PP and HPH-PP breads under the adult and elderly microbiota conditions. In all cases, ammonia concentration was 0.10 ± 0.02 mg mL−1, regardless of bread type and gut condition. Generally, deamination of amino acids increases luminal concentrations of H2, CO2, ammonia and amines.50 However, the ammonia concentration of the faecal blank was higher for both gut conditions (0.16 ± 0.05 mg mL−1 under the adult microbiota conditions and 0.34 ± 0.09 mg mL−1 under the elderly ones). This is probably because faecal blank contained only proteins from basal nutrient medium and not fermentable carbohydrates. Therefore, these results confirm that protein fermentation is reduced in the presence of fermentable carbohydrates.51 Moreover, the higher level of ammonia detected under elderly conditions may be due to the higher level of proteins of the pellets collected after in vitro digestion of breads under elderly conditions.
In light of the modest differences in the extent of colonic fermentation among the bread types, the concentration of tryptophan (Trp) and the production of its catabolites were assessed after 0 and 24 h of fermentation (Table 5) in PP bread, being peas, like other plant-based derivatives, sources of Trp. Consequently, pea could serve as a carrier for delivering Trp to intestinal microorganisms in various forms, to prevent hyperinflammation and induce long-term immune tolerance in the elderly.52
| Time (h) | PP | ||
|---|---|---|---|
| Adult | Elderly | ||
| Lower letters indicate a statistically significant difference (p < 0.05) between sampling times using the adult batch fermentations. Capital letters indicate a statistically significant difference (p < 0.05) between sampling times using the elderly batch fermentations. *: indicates statistically significant (p < 0.05) difference between batch fermentations (adult and the elderly). | |||
| Tryptophan (Trp) (μM g−1 protein) | 0 | 6111 ± 574a,* | 2544 ± 236A |
| 24 | 6029 ± 291a,* | 2692 ± 354A | |
| Tryptamine (TA) (μM g−1 protein) | 0 | 47.4 ± 9.4a,* | 20.6 ± 4.4A |
| 24 | 41.8 ± 5.1a,* | 27.1 ± 6.7A | |
| Serotonin (5HT) (μM g−1 protein) | 0 | 1.5 ± 0.3 | 1.2 ± 0.4A |
| 24 | — | 1.4 ± 0.2A | |
| Indole-3-acetic acid (ILA) (μM g−1 protein) | 0 | — | — |
| 24 | 1734.9 ± 122.5* | 293.6 ± 3.7 | |
| Oxindole (Oxi) (μM g−1 protein) | 0 | 775.1 ± 234.5b,* | 112.1 ± 16.7B |
| 24 | 1087.1 ± 2.6a,* | 265.9 ± 32.1A | |
| Indole-3-aldehyde (I3A) (μM g−1 protein) | 0 | 348.0 ± 77.2a,* | 156.3 ± 4.5A |
| 24 | 399.9 ± 54.9a,* | 140.3 ± 21.3A | |
| Kynurenine (Kyn) (μM g−1 protein) | 0 | 1.3 ± 0.2b,* | 0.5 ± 0.7B |
| 24 | 4.5 ± 0.7a,* | 1.5 ± 0.1A | |
Therefore, investigating the changes in Tpr and its metabolites can provide information on the physiological conditions in the GI tract. As can be observed in Table 5, at the beginning of fermentation, the Trp concentration was significantly lower in the elderly gut microbiota than in adult ones. A lower Trp concentration in the elderly people is relevant in age-related changes, and is connected with increased chronic inflammatory conditions.53 During colonic fermentation, Trp concentration remained constant, suggesting a low ability of gut bacteria to utilize Trp in both physiological conditions. No significant differences (p > 0.05) in the concentration of Trp were found between time samplings within each GI condition.
Trp is catabolized by gut microbiota in a range of catabolites, which play a positive role in the communication between the host and microbiota in both health and diseases,54 as they are capable of modulating the host immune response and thereby affecting the host's defence mechanisms against pathogens or diseases. When looking at specific catabolites (Table 5), ILA, Oxi and Kyn were produced during the colonic fermentation, while the concentration of TA, 5HT and I3A did not change over time. Adult and elderly gut microbiota bacteria presented different capacities to produce Trp catabolites, with higher concentrations of all catabolites produced by the adult gut microbiota. The differences observed between different GI conditions are probably attributable to the higher amount of Trp released on amount of fermented protein under the adult conditions, which serves as a substrate for Trp conversion in catabolites. Such a higher release of Trp may be ascribed to a higher proteolytic activity of adult microbiota compared to the elderly microbiota. It is also worth noticing that the data reported in Tables 4 and 5 refer to the fermentation of the same amount of pellet residuing after in vitro digestion of bread and therefore do not account for the differences in vitro protein digestibility among adult and elderly conditions.
These differences emphasize the variability in microbial metabolic responses depending on the type of dietary substrate and highlight the importance of considering the inherent microbial communities present in each donor. The ability to monitor these compounds provides valuable insights into individual metabolic responses and could serve to aid in developing tailored dietary interventions.
4. Conclusions
In the present study, bread was reformulated by partially replacing wheat flour with untreated and HPH-treated pea protein concentrate. The addition of pea protein concentrate was responsible for an increase in bread color and hardness, regardless the HPH was applied or not. Pea protein enrichment caused an increase in in vitro protein digestibility under adult and elderly conditions compared to that of the wheat bread, although no differences were found between PP and HPH-PP bread types. Moreover, in vitro protein digestibility was affected by ageing-related changes resulting in an overall reduced digestion efficiency irrespective of the bread tested. Furthermore, results on in vitro colonic fermentation of bread, showed increased levels of tryptophan catabolites produced from undigested proteins under adult conditions compared to elderly ones, suggesting a modification of microbiota functionality due to aging. In this context, these results provide new knowledge on microbial catabolism of protein enrich-bread under different gut conditions, which could serve as a foundation for future research endeavours in the development of age-specific foods aimed at enhancing gut health.Author contributions
Martina Moretton: methodology, investigation, data curation, formal analysis, visualization, writing – original draft, writing – review & editing. Melania Casertano: methodology, writing – review. Nicoletta Pellegrini: conceptualization, supervision, writing – review & editing. Monica Anese: conceptualization, methodology, resources, supervision, writing – review & editing. Vincenzo Fogliano: supervision, writing – review & editing. Edoardo Capuano: supervision, methodology, writing – review & editing.Data availability
Data will be made available on request.Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.Acknowledgements
The authors are grateful to Ms Rianne Oudejans for contributing to the acquisition of experimental data, to Mr Erik Meulenbroeks and Dr Christos Fryganas from the Food Quality and Design research group for their help in the analysis of the samples by CLSM and LCMS, and to Dr Stefano Renzetti and coworkers for the use of the bakery lab at Wageningen Food Biobased Research.This work was supported by the University of Udine (Italy) in the framework of the Strategic Plan 2022-25 – Interdepartmental Research Project CibiAmo.
References
- World Health Organization, Active Ageing: A policy framework, 2002 Search PubMed.
- World Health Organization, Keep fit for life: meeting the nutritional needs of older persons, 2002 Search PubMed.
- Y. Qin, C. Pillidge, B. Harrison and B. Adhikari, Pathways in formulating foods for the elderly, Food Res. Int., 2024, 186, 114324 CrossRef CAS PubMed.
- N. E. P. Deutz, J. M. Bauer, R. Barazzoni, G. Biolo, Y. Boirie, A. Bosy-Westphal, T. Cederholm, A. Cruz-Jentoft, Z. Krznariç, K. S. Nair, P. Singer, D. Teta, K. Tipton and P. C. Calder, Protein intake and exercise for optimal muscle function with aging: Recommendations from the ESPEN Expert Group, Clin. Nutr., 2014, 33, 929–936 CrossRef CAS PubMed.
- S. Brownie, Why are elderly individuals at risk of nutritional deficiency?, Int. J. Nurs. Pract., 2006, 12, 110–118 CrossRef PubMed.
- D. Rémond, D. R. Shahar, D. Gille, P. Pinto, J. Kachal, M.-A. Peyron, C. Nunes, D. Santos, B. Walther, A. Bordoni, D. Dupont, L. Tomás-Cobos and G. Vergères, Understanding the gastrointestinal tract of the elderly to develop dietary solutions that prevent malnutrition, Oncotarget, 2015, 6, 13858–13898 CrossRef PubMed.
- L. Laguna, A. Sarkar, G. Artigas and J. Chen, A quantitative assessment of the eating capability in the elderly individuals, Physiol. Behav., 2015, 147, 274–281 CrossRef CAS PubMed.
- S. Calligaris, M. Moretton, S. Melchior, A. C. Mosca, N. Pellegrini and M. Anese, Designing food for the elderly: the critical impact of food structure, Food Funct., 2022, 13, 6467–6483 RSC.
- P. Satusap, V. Chavasit, W. Kriengsinyos and K. Judprasong, Development of cereal andlegume based food products for the elderly, SpringerPlus, 2014, 3, 451 CrossRef PubMed.
- M. Assad-Bustillos, J. Palier, H. Rabesona, Y. Choiset, G. Della Valle and G. Feron, Role of the bolus degree of structure on the protein digestibility during in vitro digestion of a pea protein-fortified sponge cake chewed by elderly, J. Texture Stud., 2020, 51, 134–143 CrossRef PubMed.
- D. Angelino, A. Rosi, E. Ruggiero, D. Nucci, G. Paolella, V. Pignone, N. Pellegrini and D. Martini, Analysis of food labels to evaluate the nutritional quality of bread products and substitutes sold in Italy: Results from the food labelling of italian products (flip) study, Foods, 2020, 9, 1905 CrossRef CAS PubMed.
- M. Moretton, M. Alongi, N. Renoldi and M. Anese, Steering protein and carbohydrate digestibility by food design to address elderly needs: The case of pea protein enriched bread, LWT – Food Sci. Technol., 2023, 190, 115530 CrossRef CAS.
- M. Barac, S. Cabrilo, S. Stanojevic, M. Pesic, M. Pavlicevic, B. Zlatkovic and M. Jankovic, Functional properties of protein hydrolysates from pea (Pisum sativum, L) seeds, Int. J. Food Sci. Technol., 2012, 47, 1457–1467 CrossRef CAS.
- F. M. Guillin, C. Gaudichon, L. Guérin-Deremaux, C. Lefranc-Millot, G. Airinei, N. Khodorova, R. Benamouzig, P. H. Pomport, J. Martin and J. Calvez, Real ileal amino acid digestibility of pea protein compared to casein in healthy humans: a randomized trial, Am. J. Clin. Nutr., 2022, 115, 353–363 CrossRef PubMed.
- J. Overduin, L. Guérin-Deremaux, D. Wils and T. T. Lambers, NUTRALYS® pea protein: Characterization of in vitro gastric digestion and in vivo gastrointestinal peptide responses relevant to satiety, Food Nutr. Res., 2015, 59, 25622 CrossRef PubMed.
- S. Melchior, M. Moretton, S. Calligaris, L. Manzocco and M. C. Nicoli, High pressure homogenization shapes the techno-functionalities and digestibility of pea proteins, Food Bioprod. Process., 2022, 131, 77–85 CrossRef CAS.
- Z. Ling, X. Liu, Y. Cheng, X. Yan and S. Wu, Gut microbiota and aging, Crit. Rev. Food Sci. Nutr., 2022, 62, 3509–3534 CrossRef CAS PubMed.
- A. Mackie, A. I. Mulet-Cabero and A. Torcello-Gomez, Simulating human digestion: Developing our knowledge to create healthier and more sustainable foods, Food Funct., 2020, 11, 9397–9431 RSC.
- M. Moretton, M. Alongi, S. Melchior and M. Anese, Adult and elderly in vitro digestibility patterns of proteins and carbohydrates as affected by different commercial bread types, Food Res. Int., 2023, 167, 112732 CrossRef CAS PubMed.
- E. Hernández-Olivas, S. Muñoz-Pina, A. Andrés and A. Heredia, Impact of elderly gastrointestinal alterations on in vitro, digestion of salmon, sardine, sea bass and hake: Proteolysis, lipolysisand bioaccesibilityt of calcium and vitamins, Food Chem., 2020, 326, 127024 CrossRef PubMed.
- E. Hernández-Olivas, S. Muñoz-Pina, J. Sánchez-García, A. Andrés and A. Heredia, Understanding the role of food matrix on the digestibility of dairy products under elderly gastrointestinal conditions, Food Res. Int., 2020, 137, 109454 CrossRef PubMed.
- L. Nissen, F. Casciano, E. Chiarello, M. Di Nunzio, A. Bordoni and A. Gianotti, Colonic in vitro model assessment of the prebiotic potential of bread fortified with polyphenols rich olive fiber, Nutrients, 2021, 13, 1–17 CrossRef PubMed.
- A. H. Simonne, E. H. Simonne, R. R. Eitenmiller, H. A. Millsc and C. Iiia, Could the dumas method replace the Kjeldahl digestion for nitrogen and crude protein determinations in foods?, J. Sci. Food Agric., 1997, 73, 39–45 CrossRef CAS.
- AOAC, Official Methods of Analysis, Association of Official Analytical Chemists, Washington, DC, 2019 Search PubMed.
- G. M. Bornhorst and R. P. Singh, Kinetics of in vitro bread bolus digestion with varying oral and gastric digestion parameters, Food Biophys., 2013, 8, 50–59 CrossRef.
- F. M. Clydesdale and E. M. Ahmed, Colorimetry - methodology and applications, Crit. Rev. Food Sci. Nutr., 1978, 10, 243–301 CrossRef CAS PubMed.
- F. Boukid, E. Carini, E. Curti, G. Bardini, E. Pizzigalli and E. Vittadini, Effectiveness of vital gluten and transglutaminase in the improvement of physico- chemical properties of fresh bread, LWT – Food Sci. Technol., 2018, 92, 465–470 CrossRef CAS.
- S. Renzetti, M. Theunissen and K. Horrevorts, A systematic comparison of the intrinsic properties of wheat and oat bran fractions and their effects on dough and bread properties: Elucidation of chemical mechanisms, water binding, and steric hindrance, Foods, 2021, 10, 2311 CrossRef CAS PubMed.
- M. Tagliasco, M. Tecuanhuey, R. Reynard, R. Zuliani, N. Pellegrini and E. Capuano, Monitoring the effect of cell wall integrity in modulating the starch digestibility of durum wheat during different steps of bread making, Food Chem., 2022, 396, 133678 CrossRef CAS PubMed.
- A. Brodkorb, L. Egger, M. Alminger, P. Alvito, R. Assunção, S. Ballance, T. Bohn, C. Bourlieu-Lacanal, R. Boutrou, F. Carrière, A. Clemente, M. Corredig, D. Dupont, C. Dufour, C. Edwards, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. R. Mackie, C. Martins, S. Marze, D. J. McClements, O. Ménard, M. Minekus, R. Portmann, C. N. Santos, I. Souchon, R. P. Singh, G. E. Vegarud, M. S. J. Wickham, W. Weitschies and I. Recio, INFOGEST static in vitro simulation of gastrointestinal food digestion, Nat. Protoc., 2019, 14, 991–1014 CrossRef CAS PubMed.
- S. Melchior, M. Moretton, M. Alongi, S. Calligaris, M. C. Nicoli and M. Anese, Comparison of protein in vitro digestibility under adult and elderly conditions: the case study of wheat, pea, rice, and whey proteins, Food Res. Int., 2023, 163, 112147 CrossRef CAS PubMed.
- S. L. Bavaro, G. Mamone, G. Picariello, M. J. Callanan, Y. Chen, A. Brodkorb and L. Giblin, Thermal or membrane processing for Infant Milk Formula: Effects on protein digestion and integrity of the intestinal barrier, Food Chem., 2021, 347, 129019 CrossRef CAS PubMed.
- U. K. Laemmli, Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature, 1970, 227, 680–685 CrossRef CAS PubMed.
- S. Pèrez-Burillo, S. Molino, B. Navajas-Porras, A. J. Valverde-Moya, D. Hinojosa-Nogueira, A. Lopez-Moldonado, S. Pastoriza and J. A. Rufian-Henares, An in vitro batch fermentation protocol for studying the contribution of food to gut microbiota composition and functionality, Nat. Protoc., 2021, 16, 3186–3209 CrossRef PubMed.
- Z. Huyan, N. Pellegrini, W. Steegenga and E. Capuano, Insights into gut microbiota metabolism of dietary lipids: the case of linoleic acid, Food Funct., 2022, 13, 4513 RSC.
- Z. Huang, T. Schoones, J. M. Wells, V. Fogliano and E. Capuano, Substrate-driven differences in tryptophan catabolism by gut microbiota and aryl hydrocarbon receptor activation, Mol. Nutr. Food Res., 2021, 65, 2100092 CrossRef CAS PubMed.
- A. Hoehnel, C. Axel, J. Bez, E. K. Arendt and E. Zannini, Combining high-protein ingredients from pseudocereals and legumes for the development of fresh high-protein hybrid pasta: maintained technological quality and adequate sensory attributes, J. Cereal Sci., 2019, 89, 102816 CrossRef CAS.
- P. Garciá-Segovia, M. Igual and J. Martínez-Monzó, Physicochemical properties and consumer acceptance of bread enriched with alternative proteins, Foods, 2020, 9, 1–22 CrossRef PubMed.
- G. D. Stamatie, I. E. Susman, S. A. Bobea, E. Matei, D. E. Duta and F. Israel-Roming, The influence of the technological process on improving the acceptability of bread enriched with pea protein, hemp and sea buckthorn press cake, Foods, 2022, 11, 3667 CrossRef CAS PubMed.
- M. Moretton, C. Cattaneo, A. C. Mosca, C. Proserpio, M. Anese, E. Pagliarini and N. Pellegrini, Identification of desirable mechanical and sensory properties of bread for the elderly, Food Qual. Prefer., 2023, 104, 104716 CrossRef.
- R. Di Monaco, S. Cavella and P. Masi, Predicting sensory cohesiveness, hardness and springiness of solid foods from instrumental measurements, J. Texture Stud., 2008, 39, 129–149 CrossRef.
- H. Yano and W. Fu, Effective use of plant proteins for the development of “new” foods, Foods, 2022, 11, 1185 CrossRef CAS PubMed.
- M. Hiolle, V. Lechevalier, J. Floury, N. Boulier-Monthéan, C. Prioul, D. Dupont and F. Nau, In vitro digestion of complex foods: How microstructure influences food disintegration and micronutrient bioaccessibility, Food Res. Int., 2020, 128, 108817 CrossRef CAS PubMed.
- C. Wang, F. Zhao, Y. Bai, C. Li, X. Xu, K. Kristiansen and G. Zhou, Effect of gastrointestinal alterations mimicking elderly conditions on in vitro digestion of meat and soy proteins, Food Chem., 2022, 383, 132465 CrossRef CAS PubMed.
- S. Lee, K. Jo, H. G. Jeong, Y.-S. Choi, H. I. Yong and S. Jung, Understanding protein digestion in infants and the elderly: Current in vitro digestion models, Crit. Rev. Food Sci. Nutr., 2021, 63, 975–992 CrossRef PubMed.
- F. Smith, X. Pan, V. Bellido, G. A. Toole, F. K. Gates, M. S. J. Wickham, P. R. Shewry, S. Bakalis, P. Padfield and E. N. C. Mills, Digestibility of gluten proteins is reduced by baking and enhanced by starch digestion, Mol. Nutr. Food Res., 2015, 59, 2034–2043 CrossRef CAS PubMed.
- J. W. Woolnough, A. R. Bird, J. A. Monro and C. S. Brennan, The effect of a brief salivary α-amylase exposure during chewing on subsequent in vitro starch digestion curve profiles, Int. J. Mol. Sci., 2010, 11, 2780–2790 CrossRef CAS PubMed.
- L. Laguna, P. Picouet, M. D. Guàrdia, C. M. G. C. Renard and A. Sarkar, In vitro gastrointestinal digestion of pea protein isolate as a function of pH, food matrices, autoclaving, high-pressure and re-heat treatments, LWT – Food Sci. Technol., 2017, 84, 511–519 CrossRef CAS.
- V. Vinelli, P. Biscotti, D. Martini, C. Del Bo’, M. Marino, T. Meroño, O. Nikoloudaki, F. M. Calabrese, S. Turroni, V. Taverniti, A. U. Caballero, C. Andrés-Lacueva, M. Porrini, M. Gobbetti, M. De Angelis, P. Brigidi, M. Pinart, K. Nimptsch, S. Guglielmetti and P. Riso, Effects of dietary fibers on short-chain fatty acids and gut microbiota composition in healthy adults: a systematic review, Nutrients, 2022, 14, 2559 CrossRef CAS PubMed.
- C. K. Yao, J. G. Muir and P. R. Gibson, Review article: insights into colonic protein fermentation, its modulation and potential health implications, Aliment. Pharmacol. Ther., 2016, 43, 181–196 CrossRef CAS PubMed.
- K. Windey, V. de Preter and K. Verbeke, Relevance of protein fermentation to gut health, Mol. Nutr. Food Res., 2012, 56, 184–196 CrossRef CAS PubMed.
- S. Comai, A. Bertazzo, L. Bailoni, M. Zancato, C. V. L. Costa and G. Allegri, Protein and non-protein (free and protein-bound) tryptophan in legume seeds, Food Chem., 2007, 103, 657–661 CrossRef CAS.
- S. Collino, I. Montoliu, F. P. J. Martin, M. Scherer, D. Mari, S. Salvioli, L. Bucci, R. Ostan, D. Monti, E. Biagi, P. Brigidi, C. Franceschi and S. Rezzi, Metabolic signatures of extreme longevity in northern italian centenarians reveal a complex remodeling of lipids, amino acids, and gut microbiota metabolism, PLoS One, 2013, 8, e56564 CrossRef CAS PubMed.
- J. Gao, K. Xu, H. Liu, G. Liu, M. Bai, C. Peng, T. Li and Y. Yin, Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism, Front. Cell. Infect. Microbiol., 2018, 8, 13 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4fo02767a |
| This journal is © The Royal Society of Chemistry 2024 |