
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceSelenium supplementation via modulation of selenoproteins ameliorates binge drinking-induced oxidative, energetic, metabolic, and endocrine imbalance in adolescent rats’ skeletal muscle
Inés
Romero-Herrera
 a,
Fátima
Nogales
*a,
María del Carmen
Gallego-López
a,
Javier
Díaz-Castro
bc,
Olimpia
Carreras
a and
María Luisa
Ojeda
a
a,
Fátima
Nogales
*a,
María del Carmen
Gallego-López
a,
Javier
Díaz-Castro
bc,
Olimpia
Carreras
a and
María Luisa
Ojeda
a
aDepartment of Physiology, Faculty of Pharmacy, University of Seville, C/Professor García González 2, 41012-Seville, Spain. E-mail: iromero3@us.es; fnogales@us.es; mgallego3@us.es; olimpia@us.es; ojedamuri11@us.es
bInstitute of Nutrition and Food Technology “José Mataix Verdú”, University of Granada, Avenida del Conocimiento s/n, 18071-Armilla, Granada, Spain. E-mail: javierdc@ugr.es
cDepartment of Physiology, University of Granada, Granada, Spain
First published on 10th July 2024
Abstract
Adolescence is characterized by increased vulnerability to addiction and ethanol (EtOH) toxicity, particularly through binge drinking (BD), a favored acute EtOH-ingestion pattern among teenagers. BD, highly pro-oxidant, induces oxidative stress (OS), affecting skeletal muscle (SKM), where selenium (Se), an antioxidant element and catalytic center of selenoproteins, is stored, among other tissues. Investigating the effects of Se supplementation on SKM after BD exposure holds therapeutic promise. For this, we randomised 32 adolescent Wistar rats into 4 groups, exposed or not to intermittent i.p. BD [BD and control (C)] (3 g EtOH per kg per day), and supplemented with selenite [BDSe and CSe] (0.4 ppm). In SKM, we examined the oxidative balance, energy status (AMPK, SIRT-1), protein turnover (IRS-1, Akt1, mTOR, IGF-1, NF-κB p65, MAFbx, ULK1, pelF2α), serum myokines (myostatin, IL-6, FGF21, irisin, BDNF, IL-15, fractalkine, FSTL-1, FABP-3), and selenoproteins (GPx1, GPx4, SelM, SelP). In the pancreas, we studied the oxidative balance and SIRT-1 expression. Selenite supplementation mitigated BD-induced OS by enhancing the expression of selenoproteins, which restored oxidative balance, notably stimulating protein synthesis and normalizing the myokine profile, leading to improved SKM mass growth and metabolism, and reduced inflammation and apoptosis (caspase-3). Selenite restoration of SelP's receptor LRP1 expression, reduced by BD, outlines the crucial role of SKM in the SelP cycle, linking Se levels to SKM development. Furthermore, Se attenuated pancreatic OS, preserving insulin secretion. Se supplementation shows potential for alleviating SKM damage from BD, with additional beneficial endocrine effects on the pancreas, adipose tissue, liver, heart and brain that position it as a broad-spectrum treatment for adolescent alcohol consumption, preventing metabolic diseases in adulthood.
1. Introduction
Adolescence is a period marked by intense developmental, endocrine, and metabolic changes, characterized by impulsive behavior attributed to the immaturity of the prefrontal cortex.1 Therefore, addictive disorders related to substances and/or behaviors typically begin during adolescence or early adulthood.2 Besides, this age group is especially susceptible to the toxicity of ethanol (EtOH);3–8 in this context, binge drinking (BD) is a pattern of acute EtOH consumption that has lately been prominently favored by teenagers,9,10 consisting of reaching a blood alcohol concentration of 0.08% or higher within 2 hours.11 The harmfulness of BD resides in its pronounced pro-oxidant effect on DNA, proteins and lipids, as it greatly induces the hepatic activity of cytochrome P450 2E1 (CYP2E1), much more than chronic alcoholism.12 CYP2E1 generates a large amount of reactive oxygen species (ROS) to metabolize the excess of EtOH ingested, leading to oxidative stress (OS) when the antioxidant defense capacity of the cell is surpassed.13,14 NADPH oxidase 4 (NOX4) is another well-known EtOH-induced source of ROS and OS, whose activity increases serum hydrogen peroxide levels (H2O2).6,15The adverse impact of BD on the central nervous system (CNS) has been pervasively studied;16,17 however, its detrimental effects on other systems, such as the hepatic, renal or cardiovascular systems, have also been demonstrated, promoting the emergence of cardiometabolic diseases over time.3,5,18,19 Moreover, in the three principal target organs for insulin action, the liver, skeletal muscle (SKM) and adipose tissue, adolescent rats after BD exposure exhibited dysregulation of two crucial cellular energy sensors, AMP activated protein kinase (AMPK) and NAD+-dependent deacetylase sirtuin 1 (SIRT-1); this can be related to depletion in the expression of the insulin receptor substrate 1 (IRS-1), which was also found.5,7,8 This situation predisposes adolescents to the development of insulin resistance (IR),20,21 which is a factor in metabolic syndrome, that has lately become a highly prevalent disease in adolescents.22,23
SKM is a tissue that stands out in this context for being the primary site for glucose disposal in response to insulin or exercise;24 it can also store glucose and serves as the reservoir of major metabolic amino acid, to be used in case of energy requirements. Regrettably, BD during adolescence has recently been linked to impaired SKM protein turnover.7,25 Furthermore, OS in SKM leads to AMPKα stimulation,7,26 which induces a catabolic state that inhibits protein synthesis routes, such as the IRS-1/PI3K/Akt pathway of the mammalian target of rapamycin complex 1 (mTORC1); and activates the degradation mechanisms, the ubiquitin-proteasome (UPS) and autophagic-lysosomal systems, via stimulation of atrogin-1 (or muscle atrophy F-box, MAFbx) and Unc-51-like autophagy activating kinase 1 (ULK1), respectively.7,25 Additionally, although predominantly secreted by the liver, insulin-like growth factor 1 (IGF-1) can also be produced locally in SKM, where it governs protein synthesis via stimulation of the mTOR pathway.27
In the last decade, SKM has been identified as a secretory organ. Thus, cytokines and other peptides produced, expressed, or released by muscle fibers and exerting paracrine, autocrine, or endocrine effects are classified as myokines. It is crucial that SKM has been established with this role, as it provides us with a conceptual basis for understanding how it communicates with other organs, such as adipose tissue, the liver, pancreas, or the brain, to name but a few.28,29 Furthermore, disbalance of myokines has been related not only to SKM pathology, but also to general metabolic diseases and to OS.7,30,31
Given that it has been reported that OS partially mediates the impairment caused by BD,7 an antioxidant treatment appears to be a suitable therapeutic approach. Selenium (Se) is an essential trace element with renowned antioxidant properties that is the catalytic center of selenoproteins, such as the glutathione peroxidase family (GPx) or selenoprotein P (SelP).32 Our research team has discovered through several studies that during adolescence, BD alters Se homeostasis and Se tissue distribution,19 and that dietary Se supplementation is an effective strategy to mitigate BD-induced liver,5,33,34 kidney3 and cardiovascular damage;18 attributed to the ability of Se to reduce oxidative imbalance by upregulating the expressions of GPx1 and GPx4, and by modulating nuclear factor kappa B (NF-κB) activation and apoptosis-related caspase-3 protein in these organs. Selenoproteins differ with regard to their location and function within cells. GPx1 and SelP are cytosolic; GPx1 is involved in redox regulation, reducing H2O2 to water, and is also related to IR;33 SelP also has antioxidant activity, being mainly responsible for the plasmatic transport of Se from the liver to target organs, thus indirectly controlling the tissue expression of all selenoproteins. This transport is mediated through receptors in each organ: e.g., LRP1 (low-density lipoprotein receptor-related protein 1) mediates the uptake of SelP in SKM.35 GPx4 has an essential role in mitochondria, preventing their oxidation, NF-κB inflammatory response and apoptosis; this selenoprotein eliminates intracellular lipid hydroperoxides, also being implicated in ferroptosis, a regulated cell death characterized by iron-dependent lipid peroxidation. Finally, we find endoplasmic reticulum (ER)-resident selenoproteins, such as selenoprotein M (SelM), which controls ER homeostasis, protecting it through scavenging excessive ROS and suppressing apoptosis.36 In the ER, excessive ROS produce ER stress, leading to the accumulation of unfolded proteins and thus activating the unfolded protein response (UPR).37
SKM harbors approximately half of the body's total Se content,38 serving as a reservoir for the body when there are deficiencies, such as those produced by BD-induced OS. Consequently, it could be crucial to explore Se dietary supplementation in SKM after BD exposure. Therefore, the aim of this study is to analyze the oxidative, metabolic, energetic, and endocrine balance after BD exposure in the SKM of adolescent rats receiving Se supplementation, exploring a possible mechanism of action through selenoprotein modulation and synergic activity.
2. Materials and methods
2.1. Animals
Animal care procedures and experimental protocols were performed in accordance with European Union regulations (Directive 2010/63/UE) and with a Spanish Royal Decree (BOE 34/11 370, 2013), which establishes the basic rules applicable for the protection of animals used in experimentation and other scientific purposes. The experiments were also reviewed and approved by the Ethics Committee of the University of Seville (CEEA-US2019-4) and the Andalusian Regional Government (05-04-2019-065). They were meticulously planned and executed with the goal of minimizing unnecessary pain and suffering, adhering to the principles of replacement, reduction, and refinement wherever possible to decrease both the total number of animals and their use.In this experiment, 32 adolescent male Wistar rats (Centre of Production and Animal experimentation, Vice-rector's Office for Scientific Research, University of Seville) were used. Rats were received at 21 days of age and housed in groups of 2 rats per cage for 1 week, to acclimatize them to the housing conditions and handling. The experimental treatment took place during a 3-week period, starting at postnatal day (PND) 28 and ending at PND 46 (sacrifice). This period corresponds to adolescence in Wistar rats.39 The animals were kept at an automatically controlled temperature (22–23 °C) and in a 12![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :12 h light/dark photocycle (light: 07:00 a.m. to 19:00 p.m.).
:12 h light/dark photocycle (light: 07:00 a.m. to 19:00 p.m.).
On PND 28, when the adolescent period began, rats were randomly assigned into 4 groups (n = 8 per group) according to their treatment: control group (C), which received a standard pellet diet and drinking water ad libitum, and on the corresponding days, an isotonic physiological saline solution (PSS) intraperitoneally (i.p.); BD alcohol group (BD), which received a standard pellet diet and drinking water ad libitum, and on the corresponding days, an ethanol solution in PSS i.p.; control Se group (CSe), which received a standard pellet diet and Se supplementation in drinking water ad libitum, and on the corresponding days an injection of PSS i.p.; and BD alcohol Se group (BDSe), which received a standard pellet diet and drinking water supplemented with Se ad libitum, and on the corresponding days, an alcohol solution in PSS i.p.
The standard pellet diet (2014 Teklad Global 14% Protein Rodent Maintenance Diet, Harlan Laboratories, Barcelona, Spain) contained 0.23 ppm of Se. The Se-supplemented groups (CSe and BDSe) received 0.14 ppm of extra Se as anhydrous sodium selenite (PanReac, Barcelona, Spain) in drinking water over the whole experimental period.
2.2. Nutritional control
Body weight and the amount of food consumed by the rats were monitored daily until the end of the experimental period. The amount of food ingested every day was calculated by measuring this parameter every morning and the next day; the difference between them was the amount consumed. Total Se intake was calculated by multiplying the known Se concentration in the diet (0.23 ppm) and in the drinking water (0.14 ppm for CSe and BDSe groups) by the amounts of food and water ingested every day. All measurements were taken at 9:00 to avoid changes due to circadian rhythms.2.3. Ethanol treatment
Alcohol BD-exposed groups (BD and BDSe) received an i.p. injection of alcohol in PSS (20% v/v) with a dose of 3 g/ kg−1 day−1. Alcohol injections were given starting at 19:00, when the dark cycle began, for 3 consecutive days each week for 3 weeks (a total of 9 injections for each rat). No i.p. injections were given during the remaining 4 days of each week. This BD model is one of the most commonly used since it easily ensures a blood alcohol concentration of 80 mg dL−1, the value established by the NIAAA for BD.11 In addition, this method of alcoholization in adolescent rats has previously been used by this research group, registering a blood alcohol concentration of almost 125.0 mg dL−1 1 h after the last injection.12 The control groups (C and CSe) received an i.p. injection of an equal volume of PSS at the same time as the injections to the alcohol BD-exposed group.2.4. Samples
At the end of the experimental period (PND 46), the rats were fasted for 12 h (from 07:00 to 19:00). Then, 24 h after their last EtOH exposure or treatment with saline solution (last injection: PND 45, 19:00), the adolescent rats were anesthetized with an i.p. injection of 28% w/v urethane (0.5 ml per 100 g of body weight) and sacrificed (PND 46, 19:00). The blood was obtained by heart puncture and collected in tubes. The serum was prepared using low-speed centrifugation for 15 min at 1300g. The abdomen was opened by a midline incision to extract the pancreas; SKM gastrocnemius was obtained from the right leg. Once removed, both organs were immediately frozen in liquid nitrogen and stored at −80 °C.2.5. Biochemical measurements in serum
In serum, the creatinine, creatine phosphokinase (CPK) and insulin were measured with an automated analyzer (Technicon RA-1000, Bayer Diagnostics, Leverkusen, Germany).2.6. Antioxidant enzyme activity and oxidative stress markers
In order to measure the activity of the antioxidant enzymes superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx), as well as the lipid and protein oxidation, pancreas and SKM tissue samples were homogenized (25000 rpm for 1 min, 1:4 w/v) using the homogenizer Fisherbrand™ 850 (Thermo Fisher Scientific Inc., Waltham, MA, USA) in a sucrose buffer (15 mM Tris/HCl, pH 7.4, 250 mM sucrose, 1 mM EDTA, and 1 mM dithiothreitol) in an ice bath. The homogenates were centrifuged at 900g for 20 min at 4 °C, and the final supernatant was aliquoted and frozen at −80 °C until analysis. The activity of SOD (U mg−1 protein) was determined by the Fridovich method (1985),40 which is based on the ability of SOD to inhibit the reduction of cytochrome c induced by the xanthine–xanthine oxidase system; it is measured by the absorbance increase at 550 nm for 3 min, due to the reduction of cytochrome c by adding xanthine oxidase. CAT activity (U mg−1 protein) was determined using H2O2 as substrate by the assay of Beers and Sizer (1952),41 where the disappearance of H2O2 was followed spectrophotometrically at 240 nm for 3 min. The GPx activity (mU mg−1) was determined using the method described by Lawrence and Burk (1976).42 In this assay, the oxidized glutathione (GSSG) formed by the action of GPx is coupled to the reaction that catalyzes the glutathione reductase (GR) enzyme, measuring the absorbance decrease at 340 nm for 3 min due to the oxidation of nicotinamide adenine dinucleotide phosphate (NADPH). In order to measure lipid peroxidation in the pancreas and SKM, we quantified the malondialdehyde (MDA) levels (mol mg−1 protein), the end-product of the oxidative degradation of lipids, using a colorimetric reaction with thiobarbituric acid (TBA) at 535 nm, as described by Draper and Hadley (1990).43 The protein oxidation in SKM was measured by the detection of carbonyl groups (CG) (nmol mg−1 protein) at 366 nm by the method described by Reznick and Packer (1994).44 where the reaction of 2,4-dinitrophenylhydrazine (DNPH) with CG takes place.
Hydrogen peroxide (H2O2) concentration was also determined (μM), this molecule being a reactive oxygen metabolic by-product. For this, we used an H2O2 colorimetric detection kit (catalog number KA1017, Abnova, Taipei, Taiwan). The kit has a color reagent that contains a dye, xylenol orange, in an acidic solution with sorbitol and ammonium iron sulfate that reacts to produce a purple color proportional to the concentration of H2O2 in the sample. The optical density was measured at 550 nm, using a plate reader.
2.7. Protein immunoblotting assays
To this end, tissue samples were homogenized (25000 rpm for 1 min, 1:10 w/v) in 50 mm phosphate buffer [K2HPO4 50 mm, KH2PO4 50 mm, EDTA 0.01 mm, protease inhibitor 1:10 (Complete Protease Inhibitor Cocktail Tablets, Roche, Mannheim, Germany), phosphatase inhibitor 1:10 (PhosSTOP Phosphatase Inhibitor Cocktail Tablets, Roche, Mannheim, Germany)] using the homogenizer Fisherbrand™ 850 (Thermo Fisher Scientific Inc., Waltham, MA, USA). Then, the homogenates were centrifuged at 500g at 4 °C for 10 min, and the final supernatant was aliquoted and frozen at −80 °C until analysis.
The protein expression measurements in our adolescent rats were conducted using western blot, a protein immunodetection technique. In the pancreas, we evaluated the expression of GPx1 and SIRT-1; in SKM, we quantified the expression of Akt1, total-AMPKα (tAMPKα), phospho-AMPKα (Thr172) (pAMPKα), atrogin-1/MAFbx, cleaved caspase-3, phospho-eukaryotic initiation factor 2α (Ser51) (pelF2α), GPx1, GPx4, IGF-1, IRS-1, LRP1, mTOR, phospho-mTOR (Ser2448) (pmTOR), NF-κB p65, NOX4, SelM, SelP, SIRT-1 and UKL1. β-Actin was used as load control for the pancreas and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for SKM. The protein content of the samples was analyzed by the method of Lowry et al. (1951)45 and the samples utilized contained 100 or 60 μg of protein for the pancreas or SKM, respectively. The calculated appropriate volumes for those protein quantities were mixed with an equal volume of a sample buffer (Laemmli 2× Concentrate, Sigma-Aldrich, Burlington, MA, USA), plus 5% 2-mercaptoethanol (BioRad, CA, USA). After that, the samples were heated at 95 °C for 5 min, to denature the proteins. The proteins were then separated with SDS-polyacrylamide gel electrophoresis (PAGE) using acrylamide gels (6%, 7%, 9%, 10% or 12%, depending on the protein) over 1 h at 150 V; then, they were transferred onto a nitrocellulose membrane (BioRad, CA, USA) using a blot system (Transblot; BioRad, CA, USA) for 2 h at 100 V. Nonspecific membrane sites were blocked for 1 h with a blocking buffer, TBS-M: TBS (50 mM TrisHCl, 150 mM NaCl, 0.1% (v/v) Tween 20, pH 7.5) and milk powder 5% (BioRad, CA, USA); thereafter, they were probed overnight at 4 °C with the specific primary antibodies diluted in TBS-M.
The antibodies were all mouse monoclonal IgG from Santa Cruz Biotechnology (Santa Cruz, CA, USA), except pAMPKα and pelF2α, rabbit monoclonal and polyclonal antibodies, respectively from Cell Signaling Technology (Beverly, MA, USA). The primary antibodies diluted in TBS-M were: Akt1 (dilution 1:750; catalogue number sc-5298); AMPKα1/2 (1:2000; sc-74461); pAMPKα (Thr172) (1:4000; #2535); atrogin-1/MAFbx (1:750; sc-166806); cleaved caspase-3 (1:1000; sc-7272); pelF2α (Ser51) (1:750; #9721); GPx1/2 (1:1000; sc-133160); GPx4 (1:1000; sc-166570); IGF-1 (1:750; sc-518040); IRS-1 (1:500; sc-8038); LRP1 (1:750; sc-57353); mTOR (1:1000; sc-517464); pmTOR (Ser2448) (1:500; sc-293133); NF-κB p65 (1:1000; sc-8008); NOX4 (1:1000; sc-518092); SelM (1:1000; sc-514952); SelP (1:1000; sc-376858); SIRT-1 (1:500; sc-74465); and ULK1 (1:500; sc-390904). As loading controls, monoclonal mouse anti β-actin (1:10000 in TBS-M; IgG1A5441, Sigma-Aldrich, Burlington, MA, USA) and monoclonal mouse anti GAPDH (1:1000 in TBS-M; sc-32233, Santa Cruz, CA, USA) were used to detect β-actin or GAPDH for the pancreas or SKM, respectively.
After 48 h incubation of the primary antibodies, the membranes were washed 5 times (5 minutes for each wash) with TBST (TBS + 0.1% Tween 20, PanReac, Barcelona, Spain), and then incubated again, this time with the corresponding secondary antibody (Goat Anti-Mouse IgG (H + L) Horseradish Peroxidase Conjugate, catalogue number 170-6516, BioRad, CA, USA; or Goat Anti-Rabbit IgG (H + L) Horseradish Peroxidase Conjugate, 170-6515, BioRad, CA, USA) diluted in TBS-M. After a 1 h probe, the membranes were washed again 5 times with TBST (5 minutes for each wash). Finally, to reveal the results, we added the commercial developer solution Luminol ECL reagent (GE Healthcare and Lumigen, Buckinghamshire, UK) and introduced the membranes into the image analyzer Amersham Imager 600 (GE Healthcare, Buckinghamshire, UK). Then, quantification of the blots was performed by densitometry with the ImageJ program (National Institutes of Health, Bethesda, Maryland, USA). The results were expressed as percent arbitrary relative units, referring to values in control animals which were defined as 100%.
2.8. Determination of myokines
Serum levels of the following myokines were measured: interleukin 6 (IL-6), myostatin, interleukin 15 (IL-15), fractalkine (CX3CL1), fibroblast growth factor 21 (FGF21), irisin, brain-derived neurotrophic factor (BDNF), follistatin-like protein 1 (FSTL-1) and fatty acid binding protein 3 (FABP-3). For this, we used a MILLIPLEX® Rat Myokine Panel (Millipore Corp., St Charles, MO, USA), based on immunoassays on the surface of fluorescent-coded beads (microspheres), following the manufacturer's specifications (50 events per bead, 50 μL sample, gate settings of 8000–15000, time out of 60 seconds, melatonin bead set of 34). The plate was read on a LABScan 100 analyzer (Luminex Corp., Austin, TX, USA) with xPONENT software for data acquisition. Average values for each set of duplicate samples or standards were within 15% of the mean. Myokine concentrations in plasma samples were determined by comparing the mean of duplicate samples with the standard curve for each assay. Serum samples were not diluted for this analysis.2.9. Statistical analysis
The results were expressed as the mean ± SD (standard deviation) and analyzed using Prism version 8.0.2 (GraphPad Software Inc., San Diego, CA, USA) statistical analysis software. Statistical differences were assessed using the ordinary two-way analysis of variance (two-way ANOVA). When ANOVA resulted in differences, Tukey's multiple comparisons post hoc test was conducted to analyze the differences between the means of groups using all possible combinations. The sample size per group was 8 (n = 8 per group). Statistical differences were expressed as p-values, with p < 0.05 considered statistically significant. Throughout the text, p-values are illustrated as follows: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001.3. Results
Table 1 shows that total solid intake was similar among groups, while liquid intake was significantly increased in BD rats with respect to C (p < 0.01) and BDSe groups (p < 0.05). In this context, the supplemented groups consumed double the amount of Se compared to the non-supplemented groups (p < 0.001). However, BD and BDSe groups had a lower increased body weight at the end of the experiment than the control groups (p < 0.01). Accordingly, the alcohol groups also presented a lower final body weight (p < 0.05 vs. C; p < 0.01 BDSe vs. CSe). BD rats also presented a lower SKM weight and protein content than C (p < 0.001, p < 0.01) or BDSe rats (p < 0.01, p < 0.001); however, BDSe rats had increased protein content vs. CSe (p < 0.01) and slightly decreased SKM weight against the C group (p < 0.05). Serum CPK and creatinine levels were similar between groups. Even though the pancreas relative mass was similar in the 4 groups studied, in this tissue, BD exposure increased the protein content in both the BD and BDSe groups (p < 0.001).| Parameters | C | BD | CSe | BDSe |
|---|---|---|---|---|
| CPK: creatine phosphokinase. The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistic difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. | ||||
| Final body weight (g) | 159.2 ± 6.0 | 143.3 ± 17.2* | 159.5 ± 11.0●● | 142.1 ± 10.7* |
| Increased body weight (g day−1) | 5.83 ± 0.33 | 4.85 ± 1.12** | 5.85 ± 0.86●● | 4.73 ± 0.57** |
| Solid intake (mg day−1) | 15.9 ± 2.0 | 14.3 ± 1.6 | 16.3 ± 1.9 | 14.5 ± 2.5 |
| Liquid intake (mL day−1) | 21.4 ± 2.4 | 24.9 ± 1.6**,● | 22.8 ± 2.8 | 22.4 ± 3.3 |
| Total Se intake (μg day−1) | 3.49 ± 0.29 | 3.35 ± 0.08●●● | 6.38 ± 0.73*** | 6.48 ± 0.79*** |
| SKM weight (g) | 0.99 ± 0.04 | 0.77 ± 0.06***,●● | 0.95 ± 0.12 | 0.89 ± 0.07* |
| Proteins in SKM (mg g−1 wet tissue) | 43.3 ± 3.4 | 37.6 ± 4.3**,●●● | 41.6 ± 2.8●● | 47.2 ± 4.7 |
| CPK (UI L−1) | 2215 ± 122 | 2292 ± 272 | 2095 ± 285 | 2481 ± 393 |
| Creatinine (mg dL−1) | 0.45 ± 0.03 | 0.47 ± 0.03 | 0.44 ± 0.06 | 0.51 ± 0.09 |
| Pancreas weight (g) | 1.01 ± 0.09 | 1.06 ± 0.07 | 1.11 ± 0.09 | 1.04 ± 0.13 |
| Proteins in pancreas (mg g−1 wet tissue) | 159 ± 35 | 284 ± 57*** | 161 ± 31●●● | 285 ± 28*** |
Fig. 1 shows that BD rats had significantly higher SOD (p < 0.001) and CAT (p < 0.001) activities, lower GPx activity (p < 0.001) and higher levels of MDA (p < 0.001), CG (p < 0.001), NOX4 expression (p < 0.001) and H2O2 (p < 0.001). Se supplementation to BD rats significantly decreased SOD activity (p < 0.001), and increased CAT (p < 0.01) and GPx (p < 0.001) activities; it also decreased MDA levels (p < 0.001), H2O2 concentration (p < 0.01) and NOX4 expression (p < 0.001) in SKM. BDSe rats had the highest CAT activity among groups, being higher than in C or CSe (p < 0.001) animals; CG levels and NOX4 expression in BDSe rats were also higher than in C or CSe ones (CAT p < 0.001; NOX4 p < 0.001 vs. C and p < 0.01 vs. CSe). In addition, MDA levels were slightly reduced in the BDSe animals (p < 0.05). The effects of selenium supplementation to C rats were more discrete, it only increased GPx activity (p < 0.001) and NOX4 expression (p < 0.01) vs. C animals.
 | ||
| Fig. 1 Skeletal muscle oxidative balance in adolescent rats after binge drinking exposure and selenium supplementation. Superoxide dismutase (SOD) activity (A); catalase (CAT) activity (B); glutathione peroxidase (GPx) activity (C); malondialdehyde levels (MDA), showing lipid peroxidation (D); carbonyl group (CG), showing protein oxidation (E); NADPH oxidase 4 (NOX4) western blot expression and image with GAPDH as load control (F); and hydrogen peroxide (H2O2) concentration, a reactive oxygen species (ROS) (G). The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistical difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. | ||
Rats exposed to BD had in SKM significantly lower levels of IRS-1 (p < 0.001), Akt1 (p < 0.01), mTOR (p < 0.001), pmTOR (p < 0.001) and IGF-1 (p < 0.001) expressions than control animals (Fig. 2). Se supplementation to BD rats significantly enhanced all these parameters: IRS-1 (p < 0. 0.001), Akt1 (p < 0.001), mTOR (p < 0.001), pmTOR (p < 0.001) and IGF-1 (p < 0.001). BDSe rats had significant higher levels of Akt1 (p < 0.001) and IGF-1 (p < 0.001) than C or CSe rats, being the most elevated group in these proteins. CSe rats presented increased expressions of IRS-1 (p < 0.001), mTOR (p < 0.001) and pmTOR (p < 0.001) vs. C or BDSe groups, having the highest expression amongst the groups.
 | ||
| Fig. 2 Expression of proteins related to protein synthesis in the skeletal muscle of adolescent rats after binge drinking exposure and selenium supplementation. Insulin receptor substrate 1 (IRS-1) (A); Akt1 (B); mammalian target of rapamycin 1 (mTOR) (C); phospho-mTOR (Ser2448) (pmTOR) (D); insulin-like growth factor 1 (IGF-1) (E); and western blot expression images with GAPDH as load control (F). The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistical difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. | ||
Fig. 3hows that BD-exposed animals presented in SKM significantly lower expression of tAMPKα (p < 0.001), and higher expression of pAMPKα (p < 0.001), pAMPK/tAMPK ratio (p < 0.001), SIRT-1 (p < 0.001), NF-κB p65 (p < 0.001), atrogin-1/MAFbx (p < 0.01), ULK1 (p < 0.01) and myostatin (p < 0.01) than control animals. Se supplementation to BD rats significantly increased tAMPKα levels (p < 0.001), and decreased pAMPKα (p < 0.001), pAMPK/tAMPK ratio (p < 0.001) and NF-κB p65 (p < 0.01); however, it increased SIRT-1 expression levels even more (p < 0.001). BDSe rats also had higher levels of tAMPKα (p < 0.01), pAMPKα (p < 0.001), pAMPK/tAMPK ratio (p < 0.01), SIRT-1 (p < 0.001), NF-κB p65 (p < 0.001), atrogin-1/MAFbx (p < 0.01) and ULK1 (p < 0.001) than control animals. In addition, CSe rats had lower pAMPKα (p < 0.05), SIRT-1 (p < 0.001), NF-κB p65 (p < 0.001), atrogin-1/MAFbx (p < 0.01) and ULK1 (p < 0.01) expressions than BDSe rats; and slightly increased expressions of tAMPKα, pAMPKα and SIRT-1 (p < 0.05) against C animals.
 | ||
| Fig. 3 Expression of proteins related to proteolysis in the skeletal muscle of adolescent rats after binge drinking exposure and selenium supplementation. Total AMP-activated protein kinase α (AMPKα) (A); phospho-AMPKα (Thr172) (pAMPKα) (B); NAD+-dependent deacetylase sirtuin 1 (SIRT-1) (C); ratio tAMPK/pAMPK (D); nuclear factor kappa B (NF-κB p65) (E); muscle atrophy F-box (atrogin-1/MAFbx) (F); Unc-51-like autophagy activating kinase 1 (ULK1) (G); western blot expression images with GAPDH as load control (H) and serum myokine myostatin concentration (I). The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistical difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. | ||
In Fig. 4, BD depleted the expressions of all selenoproteins in SKM as compared to C rats: GPx1 (p < 0.001), GPx4 (p < 0.001), SelM (p < 0.001), and SelP (p < 0.001), and the expression of SelP's receptor LRP1 (p < 0.001). Conversely, the expressions of caspase-3 (p < 0.001) and pelF2α (p < 0.001) were elevated in the BD rats. After Se supplementation to BD rats, the expression of selenoproteins significantly increased (GPx p < 0.001; GPx4 p < 0.001; SelM p < 0.01; and SelP p < 0.01), including LRP1 (p < 0.001); while caspase-3 and pelF2α were reduced (p < 0.01 and p < 0.001, respectively). BDSe rats vs. C animals also presented higher levels of GPx1 (p < 0.05), GPx4 (p < 0.01) and caspase-3 (p < 0.01), and lower of SelP (p < 0.01). As regards the CSe group, GPx1 (p < 0.01) and GPx4 (p < 0.001) expressions were higher with respect to C. Of note, in the CSe rats, GPx4 and SelP expressions were elevated (p < 0.05) vs. BDSe, while caspase-3 was reduced (p < 0.001).
 | ||
| Fig. 4 Selenoprotein expression in the skeletal muscle of adolescent rats after binge drinking exposure and selenium supplementation. Western blot expression images with GAPDH as load control (A); glutathione peroxidase 1 (GPx1) (B); glutathione peroxidase 4 (GPx4) (C); cleaved caspase-3 (D); selenoprotein M (SelM) (E); phospho-eukaryotic initiation factor 2α (Ser51) (pelF2α) (F); selenoprotein P (SelP) (G); and low-density lipoprotein receptor-related protein 1 (LRP1) (H). The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistical difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. | ||
When the BD group is compared to the C one, it increased serum IL-6 levels (p < 0.01); however, it decreased the secretion of several myokines: FGF21 (p < 0.01), irisin (p < 0.001), BDNF (p < 0.05), CX3CL1 (p < 0.001) and FSTL-1 (p < 0.01) (Fig. 5). When BD rats were supplemented with Se, compared to BD rats they presented higher serum levels of FGF21 (p < 0.05), irisin (p < 0.001), BDNF (p < 0.01), IL-15 (p < 0.01), CX3CL1 (p < 0.01) and FSTL-1 (p < 0.01), and lower levels of FABP-3 (p < 0.001). When BDSe rats were compared to C animals, they presented higher levels of IL-6 and IL-15 (p < 0.01, p < 0.001), and lower levels of CX3CL1 (p < 0.001) and FABP-3 (p < 0.01). CSe rats had higher levels of IL-6 and IL-15 (p < 0.01, p < 0.001), and lower levels of CX3CL1 (p < 0.001) vs. C rats; and significantly the highest levels of FGF21, FSTL-1 and FABP-3 of all groups (p < 0.001).
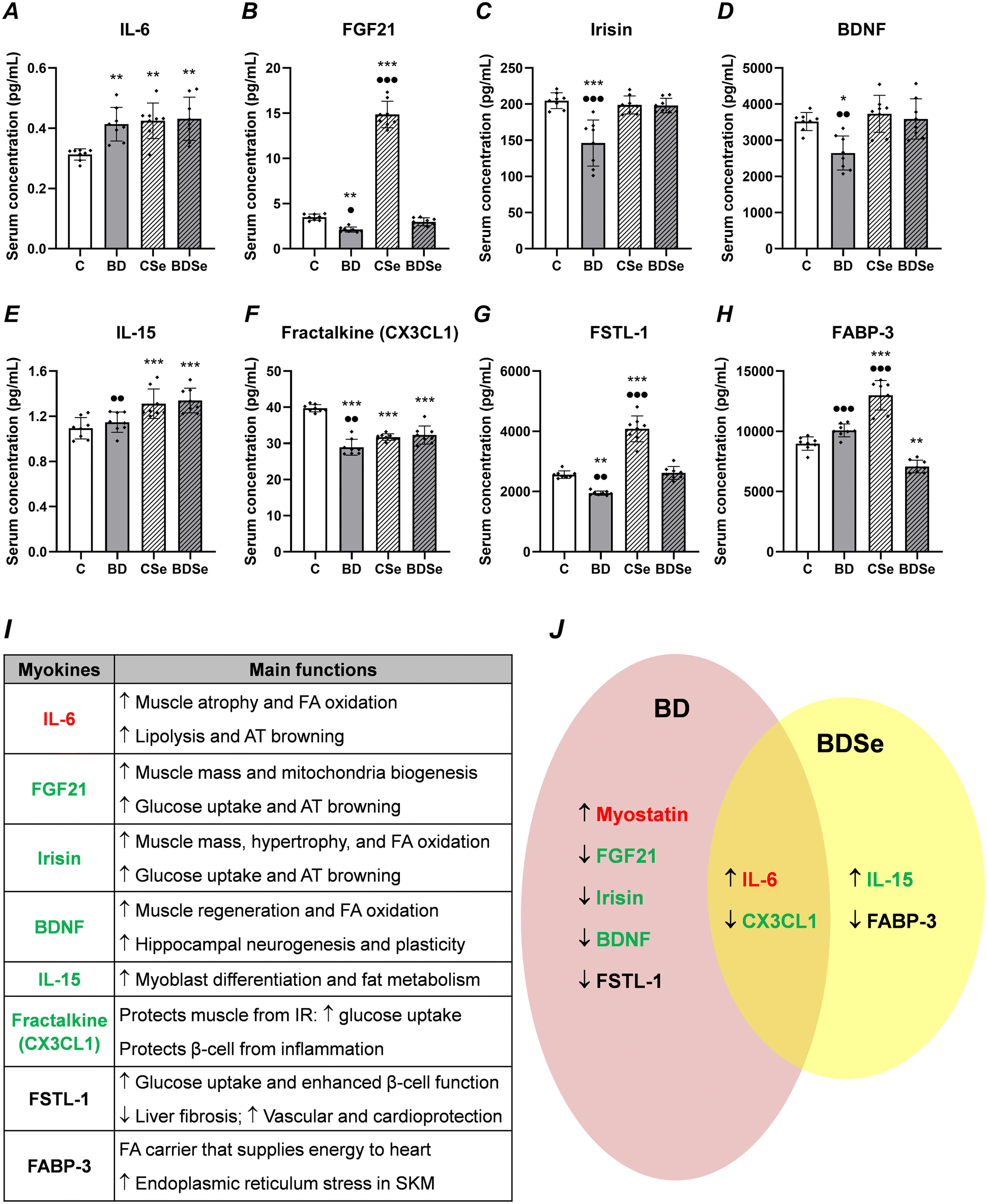 | ||
| Fig. 5 The concentration of serum myokine in adolescent rats after binge drinking exposure and selenium supplementation (A–H); table of its main known functions (I); graphic summary of the significant effects found in the BD and BDSe groups vs. the C group (J). The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistic difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. The main functions (I) were extracted from the following revisions:28,30,31,46–48 FA: fatty acid; AT: adipose tissue; SKM: skeletal muscle; IR: insulin resistance. In(I) and (J), the symbol “↑” indicates increase and “↓” indicates decrease; additionally, myokines written in red are clearly implicated in muscle atrophy, and green ones are related to hypertrophy. For (J), on the left, in a pale red color, we represent the BD group effects; on the right, in yellow, the BDSe results; in the middle, the effects in which both groups coincide. | ||
BD exposure in the pancreas increased SOD activity (p < 0.05) and decreased CAT (p < 0.05) and GPx (p < 0.01) activities, so the MDA levels increased (p < 0.01) (Fig. 6A–D). Se supplementation to BD rats, drastically increased CAT activity (p < 0.001) and GPx activity (p < 0.001), decreasing serum MDA levels (p < 0.01). The increase in CAT activity found in BDSe rats was significantly higher than in C or CSe rats (p < 0.001; p < 0.01). Se supplementation to C rats significantly increased the three antioxidant enzymes activities vs. C rats (p < 0.001), and SOD and GPx activities vs. BDSe (p < 0.001). Pancreas GPx1 expression was decreased in BD-exposed rats (p < 0.01) vs. C rats (Fig. 6E), and significantly increased in supplemented groups vs. their counterparts (p < 0.001); CSe rats also had higher GPx1 expression levels than BDSe rats (p < 0.01). In addition, rats’ exposure to BD had an elevated level of serum insulin concentration vs. C rats (p < 0.01) (Fig. 6G) and a particularly boosted pancreatic SIRT1 expression (p < 0.001) (Fig. 6H). Se supplementation to these BD rats did not affect serum insulin levels and decreased SIRT-1 expression (p < 0.01); besides, in these BDSe rats, serum insulin and SIRT-1 expression were higher than in C rats (p < 0.05 and p < 0.001, respectively). CSe rats vs. C ones presented significantly higher levels of serum insulin values (p < 0.001); additionally, CSe rats exhibited augmented serum insulin levels vs. BDSe animals (p < 0.001) and decreased pancreatic SIRT-1 expression (p < 0.001).
 | ||
| Fig. 6 Pancreas oxidative balance, serum insulin concentration and SIRT-1 pancreas expression in adolescent rats after binge drinking exposure and selenium supplementation. Superoxide dismutase (SOD) activity (A); catalase (CAT) activity (B); glutathione peroxidase (GPx) activity (C); malondialdehyde levels (MDA), showing lipid peroxidation (D); GPx1 western blot expression (E) and image (F) with β-actin as load control; serum insulin concentration (G); and SIRT-1 western blot expression (H) and image (I) with β-actin as load control. The results are expressed as mean ± SD and analyzed through two-way ANOVA, followed by Tukey's test. The number of animals in each group is 8. Groups: C: control group, BD: binge drinking group, CSe: control selenium group, and BDSe: binge drinking selenium group. Statistical difference between groups is expressed as p-value: vs. C: *p < 0.05, **p < 0.01, ***p < 0.001; vs. BDSe: ●p < 0.05, ●●p < 0.01, ●●●p < 0.001. | ||
4. Discussion
4.1. Selenite supplementation improved the compromised SKM mass and protein content induced by BD
In Table 1, we observe that the repetitive BD model used in these adolescent rats did not affect their solid intake; however, it led to a lower final body weight, which also appeared after Se supplementation. This fact indicates that there is an imbalance in growth and metabolic processes towards catabolism. Regarding liquid intake, it was significantly enhanced in BD rats, probably due to the well-known dehydrating properties of alcohol.49 Despite BD-exposed rats having underdeveloped SKM and a lower protein content, selenite supplementation prevented this situation in part, enhancing SKM weight and protein content. These effects on protein turnover balance should not only be due to the pro-oxidant effects of BD in SKM,7 but also to a depletion of Se in this tissue, as demonstrated by the lower GPx1 expression values found (Fig. 4B) in the SKM of BD rats. GPx1 is one of the lowest selenoproteins in the “hierarchy of selenoproteins”; it is highly responsive to Se levels and its expression decreases dramatically in the case of Se deficiency.50 Moreover, Se-deficiency status specifically affects SKM function51 since, as mentioned in the introduction, this tissue is one of the main Se reservoirs which releases Se into the blood when required, such as during periods of generalized OS, triggered by exposure to certain factors like BD.7,36 It has been well described that Se deficiency in SKM contributes not only to generating OS, but also to reducing the expression of cell differentiation factors such as myogenin, MyoD, and myosin heavy chain (MyHC).52 The selenite supplementation used in this study was efficient since it enhanced GPx1 expression in SKM, improving oxidative balance and SKM growth in BD rats.4.2. Selenite supplementation by improving oxidative balance contributed to modulate protein turnover in the SKM of adolescents BD exposed rats, mainly by increasing the protein synthesis process
As we have revealed, the effects on SKM mass and protein turnover from the treatments used in this study should be due in part to the pro-oxidant effects of BD exposure, also recently seen in Romero-Herrera et al.,7 and to the antioxidant activity of Se. Therefore, BD clearly disrupted oxidative balance towards an oxidative process in SKM (Fig. 1); even though BD increased the activities of SOD, which generates H2O2, and CAT, which decomposes H2O2, GPX activity was decreased. GPx did not properly eliminate the H2O2 generated by SOD, leading to oxidative damage in proteins (Fig. 1E) and lipids (Fig. 1D). Moreover, in these rats, NOX4 expression, one of the primary NOXs expressed in SKM53 with the ability to directly produce both O2˙− and mainly H2O2,54 was enhanced, and consequently the measured concentration of H2O2 was elevated. Se supplementation to BD rats partially decreased NOX4 expression, decreased SOD activity, and increased CAT and GPx expression, collaborating to partially decrease H2O2 levels and avoid lipid peroxidation; however, protein oxidation remained active. The effect of Se supplementation in GPx activity was expected since this enzyme is Se dependent, consequently affecting the activity of the other antioxidant endogenous enzymes, SOD and CAT.Interestingly, a physiological relationship has been described among NOX4, GPx1 and the insulin signaling process in SKM.55 To trigger the insulin signaling cascade, a basal amount of H2O2 is required, so over-activity of the H2O2-detoxifying selenoprotein GPx1 could lead to IR. In this context, BDSe vs. BD rats presented enhanced levels of GPx1 activity and expression (Fig. 1C and 4B) and slightly lower levels of NOX4, but these NOX4 levels were higher than in C rats (Fig. 1F) collaborating to maintain a basal amount of H2O2. Besides, the expressions of IRS-1 (Fig. 2A) and pmTOR (Fig. 2D) were enhanced, indicating that insulin sensitivity is increased, and that equilibrium in this redox axis (NOX4–GPx1) is necessary for a correct insulin process.55 Besides, Se supplementation to C adolescent rats also affected GPx1 activity and expression, NOX4 regulation, and IRS-1 expression, revealing that this axis function is dependent on Se levels.
SKM is a highly plastic organ, able to modify its mass and functional properties in response to several stimuli, such as energetic imbalance and OS.56 SKM mass is influenced by the balance between protein synthesis and breakdown: the protein turnover. Protein synthesis relies on Akt/mTOR signaling, which is usually activated by the hormones insulin and IGF-1; whereas protein breakdown hinges on the UPS, autophagy and mitophagy processes. The appearance of ROS and OS in SKM has been linked to disruption of insulin signaling,57,58 and it has recently been described that BD-induced OS specifically decreased IRS-1 and pmTOR expressions, altering protein synthesis.7 The present study found that, most probably, OS caused by BD reduced the expressions of IRS-1, IGF-1, Akt1, mTOR, and pmTOR (Fig. 2), showing a significant alteration in protein synthesis. These results are consistent with the lower SKM protein content and mass found (Table 1). Se treatment to BD adolescent rats increased the expressions of all the protein synthesis related factors analyzed, outlining the enhanced expression of IGF-1. SKM-derived IGF-1 is recognized as an important factor in muscle hypertrophy.59 The upregulation of these proteins is in consonance with the higher SKM mass and protein content found in BDSe rats. Finally, Se supplementation to control rats greatly increased IRS-1, mTOR and pmTOR expressions; nevertheless, SKM mass and protein content were similar to those of control rats. Therefore, more mechanisms should be analyzed. In any case, once again, this evidence demonstrates that Se is deeply connected to insulin function.
To fully understand protein turnover, it is necessary to also investigate the mechanisms involved in protein degradation; these are typically triggered by energy sensors like AMPK or SIRT-1 when cells require energy, as well as by the OS process.7,60 Consequently, exposure to BD would likely enhance these so-called mechanisms. In Fig. 3, our results show how BD mainly enhanced the ratio pAMPK/tAMPK, which is especially sensible to OS in SKM,26 which is also an inductor of the overexpression of protein ULK1, implicated in autophagy and mitophagy processes.27,61 Moreover, pAMPK indirectly increased atrogin-1/MAFbx expression, which activates the UPS, contributing to the breakdown of proteins.62 BD also increased SIRT-1 expression; this energy sensor specifically stimulates, via forkhead box protein O (FoxO) transcription factors, the atrogin-1/MAFbx route and autophagy–lysosome pathways.63 Finally, BD exposure enhanced NF-κB p65 expression and serum myostatin levels, both being protein UPS inductors. NF-κB is a protein complex that controls DNA transcription, involved in the cellular response to harmful stimuli such as OS, playing a key role in regulating the immune response. Relative to protein breakdown, it promotes the expression of UPS proteins, such as MuRF1.63 As regards myostatin, it is a well-known myokine that is produced and released by myocytes and acts on muscle cells, playing a key role in SKM regulation, and leading to atrophy through the Smad signaling pathway, which also stimulates the UPS.64 For all this, it is clear that BD exposure contributed to an alteration of protein turnover, increasing proteolysis and decreasing protein synthesis.
After Se administration to BD rats, the pAMPK/tAMPK ratio and NF-κB p65 expression were decreased; however, myostatin, atrogin-1/MAFbx and ULK1 levels were almost unaffected by Se. Remarkably, the expression of SIRT-1 was even more enhanced. This effect could seem deleterious, but it is important to bear in mind that SIRT-1, apart from being an energy sensor, has important antioxidant activity and is related to autophagy in a survival context.65 In fact, the crosstalk between cellular energy metabolism (AMPK/SIRT-1) and survival versus growth and repair of SKM (mTOR) has been well studied,65 since longevity is also associated with reduced mTOR signaling when it occurs alongside oxidative stress (OS) and mitochondrial catabolism.66–68 Furthermore, setting aside its negative regulation of mTOR, SIRT-1 appeared fundamental in negatively modulating acute69 or prolonged70 IGF-I increments, which could induce cell death, since SIRT-1 is crucial in maintaining survival. This could be the circumstance in the SKM of BDSe rats, where IGF-1 expression was greatly increased, leading to an overexpression of SIRT-1 to counteract the effects of IGF-1 in cell survival. In summary, the beneficial effects of Se supplementation to BD rats on protein turnover relied primarily on enhancement in protein synthesis, inducing an endocrine response via insulin and IGF-1, which was counteracted by SIRT-1 overexpression.
4.3. BD exposure during adolescence compromised the expression and functions of SKM selenoproteins. Selenite supplementation counteracted this situation, avoiding mitochondrial apoptosis and ER stress
Recently Jing et al.36 reported that OS in SKM affects selenogenes related to growth retardation, mitochondrial dysfunction, ER stress, and protein and lipid metabolism disorders. Supplementation with Se increased muscular Se deposition, exhibited protective effects against ROS levels, enhanced antioxidant capacity in SKM, and mitigated mitochondrial dysfunction and ER stress; moreover, selenoproteins improved protein biosynthesis by regulating the Akt/mTOR/S6K1 signaling pathway. Among them, GPx4 in mitochondria and SelM in ER play a unique role during this protection. For these reasons, knowing that BD exposure during adolescence led to general OS,7 we have determined for the first time some of these selenoproteins in the SKM of BD adolescent rats. After BD exposure, adolescent rats presented a depletion of the 4 selenoproteins measured (Fig. 4): GPx1, GPx4, SelM and SelP. These results are in agreement with the previously mentioned fact that SKM is one of the main Se reservoirs which release Se into the blood when needed; such is the case after BD exposure, when deep OS occurs.Therefore, as previously discussed, the expression and activity of the cytosolic GPx1, a selenoprotein very sensitive to Se status, is decreased; contributing to generating the OS detected in the SKM of BD rats. Moreover, GPx4, which ranks high in the hierarchical system of selenoprotein transcripts,71 is also decreased, indicating that Se depletion in SKM after BD exposure has an important magnitude (around 15–20% of depletion). GPx4 is specifically implicated in the protection of cell membranes and organelles, especially in mitochondria lipidic antioxidant defense. GPx4 is also an important modulator of ferroptosis and mitochondria-derived apoptosis, cell death that is ultimately activated by caspase-3. In addition, the inflammatory NF-κB pathway is negatively modulated by GPx4; therefore, it reduces the production of proinflammatory mediators.72,73 Consequently, our results are in agreement since BD rats presented lower GPx4 (Fig. 4C) and higher NF-κB p65 (Fig. 3E) and cleaved caspase-3 (Fig. 4D) expression levels in SKM, indicating that mitochondrial function was compromised.
SelM is one of the most important selenoproteins resident in the ER; apart from its mentioned role in ER protection, it also participates in other fundamental SKM functions. ER functions as a quality-control organelle for proteins, allowing only normal proteins to persist in vesicles. Excessive ROS impairs protein homeostasis (proteostasis), causing stress in the ER, which induces the accumulation of unfolded proteins in the ER, thus triggering the unfolded protein response (UPR). The consequences of UPR include inhibition of protein synthesis, modulation of gene expression, and decisions about the fate of a cell like apoptosis. Three ER-localized transmembrane sensors mediate the UPR, protein kinase RNA-like ER kinase (PERK), inositol-requiring enzyme 1 (IRE1), and activating transcription factor 6 (ATF6); these are used as UPR markers. When an unfolded protein binds to PERK, conformational changes that phosphorylate the eukaryotic translation initiation factor 2α (peIF2α) are produced; peIF2α reduces global protein synthesis and protein load. In this context, BD exposure reduces SelM expression, decreasing ER antioxidant defense, and activating UPR, as evidenced by the higher expression of the UPR marker pelF2α (Fig. 4F). This clearly contributes to the disbalance in protein turnover found in BD-exposed rats. Besides, SelM has also been directly related to the protein synthesis indicators Akt and mTOR, both decreased in BD rats (Fig. 2).
Finally, SelP expression was also inhibited by BD in the SKM of adolescent rats. SelP has 10 selenocysteine residues instead of one, as in the rest of the selenoproteins, being a perfect plasmatic transporter of Se from the liver to tissues with antioxidant functions.35 It plays a pivotal role in recycling and storing Se in essential organs, especially under conditions of limited Se supply via the SelP cycle.74 The correct SelP cycle depends on tissue SelP receptors from the lipoprotein receptor family (LRPs). Three kinds of LRPs have been identified as SelP receptors, namely, apolipoprotein E receptor 2 (ApoER2/LRP8), associated with SelP uptake in the brain, testes, and bone; megalin (LRP2), expressed in the kidney and brain; and LRP1, located only in SKM.75 The SelP–LRP1 axis is considered the main pathway implicated in the different effects that SelP provokes in SKM.35 In BD-exposed rats, SelP levels were decreased in SKM, together with the expression of LRP1 (Fig. 4H). This implies that the SelP cycle may be deriving Se to protect the brain and endocrine glands, at the expense of SKM. The idea that SKM acts as Se reservoir tissue which delivers Se to the blood when bodily circumstances need it is reinforced in the EtOH-exposure case. Similar effects of SKM as an Se reservoir were found in dams chronically exposed to alcohol.76
Selenite supplementation to BD adolescent rats significantly increased the 4 selenoproteins studied, contributing to a better cytosolic, mitochondrial and ER oxidative balance. These effects not only have repercussions in OS, but also in SKM structure and function since the increase in GPx4 with selenite reduces endogenous apoptosis and inflammation, as demonstrated by the lower expression levels of cleaved caspase-3 and NF-κB p65, respectively, found in BDSe rats. Moreover, the increase in SelM avoids ER stress and UPR, as indicated by the decreased levels of pelF2α found in these rats, having counteracted the BD-induced alterations in proteostasis. Additionally, SelM has been directly correlated to an increase in Akt and mTOR protein synthesis signaling, both of which increased in BDSe rats compared to BD ones. Finally, thanks to the additional Se supplied in this study, the SelP cycle seems to be well balanced, allowing Se to be deposited in SKM via SelP-LRP1, expressions of which were both increased in SKM.
4.4. Selenite supplementation improved the autocrine/paracrine regulation of metabolism in SKM as well as in the endocrine regulation of other tissues, by modulating serum myokine levels after BD exposure
This study reconfirmed that BD exposure alters the myokine secretion profile (Fig. 5), not only impacting SKM function and structure, but also collaborating to promote whole-body metabolic and energetic imbalance. Most importantly, this experimental design demonstrated for the first time that selenite supplementation to BD adolescent rats is an effective therapy for restoring the serum myokine profile toward a more physiological state.The selenite treatment used restored several myokines to serum control levels, such as FSTL-1, the atrophic myostatin, and the hypertrophic FGF21, irisin, and BDNF, deeply altered by BD exposure. Moreover, selenite increased serum levels of IL-5, which is implicated in myoblast differentiation and muscle mass development and facilitates glucose uptake and fat metabolism in SKM.47 Therefore, this myokine is clearly contributing to the increase in protein synthesis found in BDSe rats compared to BD rats. Se-supplementation also recovered serum FSTL-1 levels, and decreased FABP-3 ones. However, it was not able to prevent the increase in atrophic IL-6 or the decrease in hypertrophic CX3CL1 serum levels caused by BD exposure.
Apart from their autocrine/paracrine effects on SKM, the myokines FGF21, irisin and FSTL-1 play an important endocrine role in promoting insulin sensitivity by different mechanisms.28,46,77 Their reduction after BD exposure indicates that a general IR process was probably taking place; and that selenite supplementation by increasing FGF21, irisin and FSTL-1 contributed to improving insulin sensitivity. Moreover, FSTL-1 promotes endothelial cell function and revascularization, maintains cardiac growth and development, and protects the liver from fibrosis.30 In fact, previous studies obtained in similar BD experimental models, demonstrated that BD led to OS, inflammation and apoptosis in hepatocytes5,33 and cardiac myocytes,18 increasing endothelial markers and blood pressure. Therefore, selenite supplementation improved not only SKM mass, but also endocrine, liver and cardiac function, positioning itself as a broad-spectrum therapy against alcohol consumption during adolescence.
Additionally, the myokine BDNF also has important endocrine effects on neurogenesis, protecting neurons from OS and apoptosis, and promoting synaptic plasticity and memory and learning processes.47 The normalization of serum BDNF levels produced after selenite supplementation to BD adolescent rats will probably counteract the well-known neurotoxic effects of EtOH.78 Interestingly, a direct relationship between SIRT-1 expression and BNDF secretion has been described;79 this correlation was also present in BDSe rats. With regard to serum FABP-3 levels, Se supplementation to BD rats decreased them. FABP-3 is a cytoplasmic fatty acid carrier that facilitates the transport of lipophilic substances from sarcolemma to cellular compartments,80 supplying energy to SKM cells; importantly, it also contributes to increasing ER stress in SKM.48 Se supplementation to BD rats by increasing SelM decreased ER stress, which is also confirmed by the lower serum FABP-3 levels found in BDSe adolescent rats; this is consistent with an appropriate proteostasis.
Finally, relative to selenite supplementation in control rats, it should be noted that it resulted in a very significant enhancement in FGF21 and FSTL-1 serum levels, which have beneficial actions on hepatic metabolism.28,30 This beneficial role of selenite supplementation during adolescence had been previously reported, pointing to selenite as an important source of Se supplementation to act specifically on the liver.33,81
4.5. BD led to OS in the pancreas of adolescent rats, affecting insulin secretion
In order to understand whether the lower SKM protein synthesis found after BD exposure is related to the amount of insulin secreted by the pancreas, we also needed to study this endocrine organ. In our results, it seems quite clear that BD affected the pancreas exocrine/endocrine physiology, because it increased protein concentration (Table 1) and lipid peroxidation (Fig. 6D), which are indicative of protein plug formation and the progression of exocrine pancreatic damage;82 and it also affected insulin secretion (Fig. 6G). In a previous recent paper,8 we confirmed that BD exposure during adolescence led to hyperglycemia and hyperinsulinemia, which were compatible with a general IR process.BD altered all antioxidant enzyme activities (Fig. 6), which resulted in lipid peroxidation in the pancreas. However, pancreatic β-cells, being especially sensitive to OS,83 were apparently functional since serum insulin levels were increased. This could be due to the protective action of the energy sensor SIRT-1, overexpressed in our BD rats. SIRT-1 is also involved in the prevention of inflammatory and oxidative processes, being essential for the formation and maintenance of pancreatic β-cells.84–87 Therefore, the impaired SKM insulin function found could be due to a disruption of the insulin signaling cascade in this tissue, and not to reduced pancreatic insulin secretion.
Se supplementation to BD rats avoided lipid peroxidation by increasing the activity of the selenoprotein GPx and the antioxidant CAT. This enhanced CAT activity is important since this enzyme is crucial to eliminating H2O2 in the pancreas.88,89 BDSe rats also presented high SIRT-1 levels in the pancreas, collaborating with an appropriate pancreatic function; therefore, serum insulin levels were not affected in this group of rats. This fact and the proper SKM IRS-1 expression shown (Fig. 2A), indicate that the insulin signaling pathway is functioning properly and that protein biosynthesis is being actively maintained by this route, counteracting BD-exposure effects.
Eventually, CS rats presented higher activities of the three antioxidant enzymes; these increases were proportional, avoiding lipid peroxidation. It has been shown that an increase in the three enzymes and their combined action is very effective in stimulating the function of pancreatic β-cells.90 Therefore, serum insulin levels were greatly increased; however, since IRS-1 expression in SKM and in other insulin target tissues of rats exposed to the same amount of selenite such as the liver were increased, and serum glucose levels were unaffected,5 a general IR process is not taking place in CSe rats. Nevertheless, it is quite clear that Se is a potent element to induce insulin secretion and enhance insulin response to consider when required.
5. Conclusions
Based on the results obtained, it has been confirmed that BD exposure during adolescence leads to oxidation and catabolism in SKM and the pancreas. Despite the increased serum insulin levels, its anabolic effects on SKM were decreased. Interestingly, it has been established for the first time that BD exposure during adolescence specifically decreases the expression of selenoproteins in SKM (GPx1, GPx4, SelP and SelM). This points to SKM as an important tissue in the SelP cycle after EtOH insults, since LRP1 expression was clearly decreased in BD rats, and links Se status to SKM development. Therefore, the selenite supplementation used in BD-exposed adolescent rats enhanced LRP1 and expression of selenoproteins, thereby improving SKM functions by modulating oxidative balance, energy-status, proteostasis, inflammation, apoptosis and ER homeostasis (Fig. 7). Besides, selenite not only adjusted the serum myokine profile toward a more anabolic one in SKM, but Se also had beneficial endocrine repercussions in the pancreas, WAT, liver, heart and brain, which positions it as a broad-spectrum therapy against alcohol consumption during adolescence. Furthermore, Se also decreased lipid peroxidation in the pancreas, contributing to normal serum insulin secretion. | ||
| Fig. 7 Summary of results in the skeletal muscle of adolescent rats after binge drinking exposure and selenium supplementation. After BD exposure, firstly, cytoplasmatic reactive oxygen species (ROS) are generated, leading to oxidative stress (OS) in this space. Besides, ROS appeared in the mitochondria, affecting their function, and collaborating to generate mitochondrial OS, apoptosis (via caspase-3 stimulation) and inflammation (via NF-κB activation), but also probably to ferroptosis. ROS also emerged in the endoplasmic reticulum (ER), generating OS there and accumulation of unfolded proteins, which activates the unfolded protein response (UPR) via pelF2α stimulation. All these situations generated by BD lead to a stimulation of proteolytic proteins (AMPK, SIRT-1, MAFbx, ULK1, myostatin, IL-6, NF-κB p65, caspase-3 and pelF2α), and to an inhibition of protein synthesis factors (IRS-1, Akt1, pmTOR, IGF-1, FGF21, irisin, BDNF, IL-15 and CX3CL1). The imbalance produced promotes retardation of SKM growth. Supplementation with the antioxidant Se increases the expression of selenoproteins in the cytoplasm (GPx1 and Selenoprotein P), mitochondria (GPx4) and ER (Selenoprotein M), reducing ROS, OS, and their related damage. In the BDSe rats, there is inhibition of the so-called atrophic proteins, and a more profound and significant stimulation of the pro-anabolism proteins mentioned. Consequently, selenite facilitates normal SKM growth. Dashed lines indicate inhibition, while continuous lines indicate stimulation. The wider arrow above the anabolic proteins after Se supplementation indicates a stronger stimulatory effect. | ||
Data availability
Data are available as ESI for review purposes (original western blot images). The rest of the data are all presented in this article.Conflicts of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.Acknowledgements
This research was funded by grants from the Andalusian Regional Government, which support the CTS-193 research group (2021/CTS-193; 2019/CTS-193), as well as by VII Plan Propio de Investigación y Transferencia – University of Seville 2022. The first author has a predoctoral researcher and teaching personnel contract, number USE-22212-V, also funded by the Andalusian Regional Government. Additionally, we very much thank the Research, Technology and Innovation Centre (CITIUS, Centro de Investigación, Tecnología e Innovación) of the University of Seville for its continuous support with respect to the use of its facilities.References
- A. Caballero, R. Granberg and K. Y. Tseng, Mechanisms contributing to prefrontal cortex maturation during adolescence, Neurosci. Biobehav. Rev., 2016, 70, 4–12 CrossRef.
- R. A. Chambers, J. R. Taylor and M. N. Potenza, Developmental neurocircuitry of motivation in adolescence: a critical period of addiction Vulnerability, Am. J. Psychiatry, 2003, 160, 1041–1052 CrossRef PubMed.
- P. Sobrino, M. L. Ojeda, F. Nogales, M. L. Murillo and O. Carreras, Binge drinking affects kidney function, osmotic balance, aldosterone levels, and arterial pressure in adolescent rats: the potential hypotensive effect of selenium mediated by improvements in oxidative balance, Hypertens. Res., 2019, 42, 1495–1506 CrossRef CAS.
- L. Ai, E. Perez, A. Asimes, T. Kampaengsri, M. Heroux, A. Zlobin, M. A. Hiske, C. S. Chung, T. R. Pak and J. A. Kirk, Binge alcohol exposure in adolescence impairs normal heart growth, J. Am. Heart Assoc., 2020, 9, 15611 Search PubMed.
- F. Nogales, O. Cebadero, I. Romero-Herrera, R. M. Rua, O. Carreras and M. L. Ojeda, Selenite supplementation modulates the hepatic metabolic sensors AMPK and SIRT1 in binge drinking exposed adolescent rats by avoiding oxidative stress, Food Funct., 2021, 12, 3022–3032 RSC.
- M. C. Gallego-Lopez, M. L. Ojeda, I. Romero-Herrera, F. Nogales and O. Carreras, Folic acid homeostasis and its pathways related to hepatic oxidation in adolescent rats exposed to binge drinking, Antioxidants, 2022, 11, 362 CrossRef.
- I. Romero-Herrera, F. Nogales, J. Diaz-Castro, J. Moreno-Fernandez, M. D. C. Gallego-Lopez, J. J. Ochoa, O. Carreras and M. L. Ojeda, Binge drinking leads to an oxidative and metabolic imbalance in skeletal muscle during adolescence in rats: endocrine repercussion, J. Physiol. Biochem., 2023, 79, 799–810 CrossRef PubMed.
- I. Romero-Herrera, F. Nogales, M. D. C. Gallego-López, J. Díaz-Castro, J. Moreno-Fernandez, J. J. Ochoa, O. Carreras and M. L. Ojeda, Adipose tissue homeostasis orchestrates the oxidative, energetic, metabolic and endocrine disruption induced by binge drinking in adolescent rats, J. Physiol., 2023, 601, 5617–5633 CrossRef CAS PubMed.
- E. E. Bonar, M. J. Parks, M. Gunlicks-Stoessel, G. R. Lyden, C. J. Mehus, N. Morrell and M. E. Patrick, Binge drinking before and after a COVID-19 campus closure among first-year college students, Addict. Behav., 2021, 118, 106879 CrossRef PubMed.
- S. Muzi, A. Sansò and C. S. Pace, What's happened to Italian adolescents during the COVID-19 pandemic? A preliminary study on symptoms, problematic social media usage, and attachment: relationships and differences with pre-pandemic peers, Front. Psychiatry, 2021, 12, 590543 CrossRef.
- NIAAA , What is binge drinking? How common is binge drinking? Understanding binge drinking, https://www.niaaa.nih.gov/publications/brochures-and-fact-sheets/binge-drinking, (accessed 1 January 2024).
- F. Nogales, R. M. Rua, M. L. Ojeda, M. L. Murillo and O. Carreras, Oral or intraperitoneal binge drinking and oxidative balance in adolescent rats, Chem. Res. Toxicol., 2014, 27, 1926–1933 Search PubMed.
- M. A. Abdelmegeed, A. Banerjee, S. Jang, S. H. Yoo, J. W. Yun, F. J. Gonzalez, A. Keshavarzian and B. J. Song, CYP2E1 potentiates binge alcohol-induced gut leakiness, steatohepatitis and apoptosis, Free Radicals Biol. Med., 2013, 65, 1238–1245 CrossRef CAS.
- Y. Lu and A. I. Cederbaum, Cytochrome P450S and alcoholic liver disease, Curr. Pharm. Des., 2018, 24, 1502 CrossRef CAS.
- Q. Sun, W. Zhang, W. Zhong, X. Sun and Z. Zhou, Pharmacological inhibition of NOX4 ameliorates alcohol-induced liver injury in mice through improving oxidative stress and mitochondrial function, Biochim. Biophys. Acta, 2017, 1861, 2912–2921 CrossRef CAS.
- B. Lees, L. R. Meredith, A. E. Kirkland, B. E. Bryant and L. M. Squeglia, Effect of alcohol use on the adolescent brain and behavior, Pharmacol., Biochem. Behav., 2020, 192, 172906 CrossRef CAS PubMed.
- S. Lannoy, J. Billieux, V. Dormal and P. Maurage, Behavioral and cerebral impairments associated with binge drinking in youth: a critical review, Psychol. Belg., 2019, 59, 116 CrossRef PubMed.
- M. L. Ojeda, P. Sobrino, R. M. Rua, M. D. C. Gallego-Lopez, F. Nogales and O. Carreras, Selenium, a dietary-antioxidant with cardioprotective effects, prevents the impairments in heart rate and systolic blood pressure in adolescent rats exposed to binge drinking treatment, Am. J. Drug Alcohol Abuse, 2021, 47, 680–693 CrossRef CAS PubMed.
- M. L. Ojeda, F. Nogales, M. D. C. Gallego-López and O. Carreras, Binge drinking during the adolescence period causes oxidative damage-induced cardiometabolic disorders: A possible ameliorative approach with selenium supplementation, Life Sci., 2022, 301, 120618 CrossRef CAS.
- L. Giovannini and S. Bianchi, Role of nutraceutical SIRT1 modulators in AMPK and mTOR pathway: Evidence of a synergistic effect, Nutrition, 2017, 34, 82–96 CrossRef CAS.
- G. M. Reaven, Pathophysiology of insulin resistance in human disease, Physiol. Rev., 1995, 75, 473–486 CrossRef PubMed.
- H. Sharif, S. S. Sheikh, T. Seemi, H. Naeem, U. Khan and S. S. Jan, Metabolic syndrome and obesity among marginalised school-going adolescents in Karachi, Pakistan: a cross-sectional study, Lancet Reg. Health Southeast Asia, 2024, 21, 100354 CrossRef PubMed.
- V. Higgins and K. Adeli, Pediatric metabolic syndrome: pathophysiology and laboratory assessment, eJIFCC, 2017, 28, 25 Search PubMed.
- L. Peifer-Weiß, H. Al-Hasani and A. Chadt, AMPK and beyond: the signaling network controlling RabGAPs and contraction-mediated glucose uptake in skeletal muscle, Int. J. Mol. Sci., 2024, 25, 1910 CrossRef.
- C. Cáceres-Ayala, R. G. Mira, M. J. Acuña, E. Brandan, W. Cerpa and D. L. Rebolledo, Episodic binge-like ethanol reduces skeletal muscle strength associated with atrophy, fibrosis, and inflammation in young rats, Int. J. Mol. Sci., 2023, 24, 1655 CrossRef.
- T. Toyoda, T. Hayashi, L. Miyamoto, S. Yonemitsu, M. Nakano, S. Tanaka, K. Ebihara, H. Masuzaki, K. Hosoda, G. Inoue, A. Otaka, K. Sato, T. Fushiki and K. Nakao, Possible involvement of the α1 isoform of 5′AMP-activated protein kinase in oxidative stress-stimulated glucose transport in skeletal muscle, Am. J. Physiol. Endocrinol. Metab., 2004, 287, E166–E173 CrossRef CAS PubMed.
- T. Yoshida and P. Delafontaine, Mechanisms of IGF-1-mediated regulation of skeletal muscle hypertrophy and atrophy, Cells, 2020, 9, 1970 CrossRef CAS.
- M. C. K. Severinsen and B. K. Pedersen, Muscle–organ crosstalk: The emerging roles of myokines, Endocr. Rev., 2020, 41, 594 CrossRef.
- B. K. Pedersen and M. A. Febbraio, Muscle as an endocrine organ: focus on muscle-derived interleukin-6, Physiol. Rev., 2008, 88, 1379–1406 CrossRef CAS.
- W. Chen, L. Wang, W. You and T. Shan, Myokines mediate the cross talk between skeletal muscle and other organs, J. Cell Physiol., 2021, 236, 2393–2412 CrossRef CAS PubMed.
- R. Mancinelli, F. Checcaglini, F. Coscia, P. Gigliotti, S. Fulle and G. Fanò-Illic, Biological aspects of selected myokines in skeletal muscle: focus on aging, Int. J. Mol. Sci., 2021, 22, 8520 CrossRef PubMed.
- M. P. Rayman, Selenium and human health, Lancet, 2012, 379, 1256–1268 CrossRef PubMed.
- M. L. Ojeda, O. Carreras, P. Sobrino, M. L. Murillo and F. Nogales, Biological implications of selenium in adolescent rats exposed to binge drinking: Oxidative, immunologic and apoptotic balance, Toxicol. Appl. Pharmacol., 2017, 329, 165–172 CrossRef PubMed.
- M. L. Ojeda, R. M. Rua, M. L. Murillo, O. Carreras and F. Nogales, Binge drinking during adolescence disrupts se homeostasis and its main hepatic selenoprotein expression, Alcohol: Clin. Exp. Res., 2015, 39, 818–826 CrossRef PubMed.
- Y. Saito, Selenium transport mechanism via selenoprotein P—its physiological role and related diseases, Front. Nutr., 2021, 8, 685517 CrossRef PubMed.
- J. Jing, Y. He, Y. Liu, J. Tang, L. Wang, G. Jia, G. Liu, X. Chen, G. Tian, J. Cai, L. Che, B. Kang and H. Zhao, Selenoproteins synergistically protect porcine skeletal muscle from oxidative damage via relieving mitochondrial dysfunction and endoplasmic reticulum stress, J. Anim. Sci. Biotechnol., 2023, 14, 79 CrossRef PubMed.
- X. Chen, C. Shi, M. He, S. Xiong and X. Xia, Endoplasmic reticulum stress: molecular mechanism and therapeutic targets, Signal Transduction Targeted Ther., 2023, 8, 352 CrossRef PubMed.
- L. T. Wesolowski, P. L. Semanchik and S. H. White-Springer, Beyond antioxidants: Selenium and skeletal muscle mitochondria, Front. Vet. Sci., 2022, 9, 1011159 CrossRef PubMed.
- L. Spear, Modeling adolescent development and alcohol use in animals, Alcohol Res. Health, 2000, 24, 115 CAS.
- I. Fridovich, in CRC Handb. Methods Oxyg. Radic. Res, ed. R. A. Greenwald, CRC Press, 1st edn, 1985, pp. 213–216 Search PubMed.
- R. F. Beers and I. W. Sizer, A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase, J. Biol. Chem., 1952, 195, 133–140 CrossRef CAS.
- R. A. Lawrence and R. F. Burk, Glutathione peroxidase activity in selenium-deficient rat liver, Biochem. Biophys. Res. Commun., 1976, 71, 952–958 CrossRef CAS PubMed.
- H. H. Draper and M. Hadley, Malondialdehyde determination as index of lipid Peroxidation, Methods Enzymol., 1990, 186, 421–431 CAS.
- A. Z. Reznick and L. Packer, Oxidative damage to proteins: Spectrophotometric method for carbonyl assay, Methods Enzymol., 1994, 233, 357–363 CAS.
- O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J. Randall, Protein measurement with the folin phenol reagent, J. Biol. Chem., 1951, 193, 265–275 CrossRef CAS PubMed.
- A. R. D. O. Dos Santos, B. D. O. Zanuso, V. F. B. Miola, S. M. Barbalho, P. C. Santos-Bueno, U. A. P. Flato, C. R. P. Detregiachi, D. V. Buchaim, R. L. Buchaim, R. J. Tofano, C. G. Mendes, V. A. C. Tofano and J. F. D. S. Haber, Adipokines, myokines, and hepatokines: crosstalk and metabolic repercussions, Int. J. Mol. Sci., 2021, 22, 2639 CrossRef.
- J. H. Lee and H. S. Jun, Role of myokines in regulating skeletal muscle mass and function, Front. Physiol., 2019, 10, 42 CrossRef PubMed.
- S. M. Lee, S. H. Lee, Y. Jung, Y. Lee, J. H. Yoon, J. Y. Choi, C. Y. Hwang, Y. H. Son, S. S. Park, G. S. Hwang, K. P. Lee and K. S. Kwon, FABP3-mediated membrane lipid saturation alters fluidity and induces ER stress in skeletal muscle with aging, Nat. Commun., 2020, 11, 5661 CrossRef CAS PubMed.
- M. Epstein, Alcohol's impact on kidney function, Alcohol Health Res. World, 1997, 21, 84–92 CAS.
- M. S. Saedi, C. G. Smith, J. Frampton, I. Chambers, P. R. Harrison and R. A. Sunde, Effect of selenium status on mRNA levels for glutathione peroxidase in rat liver, Biochem. Biophys. Res. Commun., 1988, 153, 855–861 CrossRef CAS PubMed.
- A. Bartholomew, D. Latshaw and D. E. Swayne, Changes in blood chemistry, hematology, and histology caused by a selenium/vitamin E deficiency and recovery in chicks, Biol. Trace Elem. Res., 1998, 62, 7–16 CrossRef PubMed.
- B. W. Bao, Z. Kang, Y. Zhang, K. Li, R. Xu and M. Y. Guo, Selenium deficiency leads to reduced skeletal muscle cell differentiation by oxidative stress in mice, Biol. Trace Elem. Res., 2023, 201, 1878–1887 CrossRef.
- G. K. Sakellariou, A. Vasilaki, J. Palomero, A. Kayani, L. Zibrik, A. Mcardle and M. J. Jackson, Studies of mitochondrial and nonmitochondrial sources implicate nicotinamide adenine dinucleotide phosphate oxidase(s) in the increased skeletal muscle superoxide generation that occurs during contractile activity, Antioxid. Redox Signal, 2013, 18, 603–621 CrossRef PubMed.
- K. D. Martyn, L. M. Frederick, K. Von Loehneysen, M. C. Dinauer and U. G. Knaus, Functional analysis of NOX4 reveals unique characteristics compared to other NADPH oxidases, Cell Signal, 2006, 18, 69–82 CrossRef PubMed.
- C. E. Xirouchaki, Y. Jia, M. J. Mcgrath, S. Greatorex, M. Tran, T. L. Merry, D. Hong, M. J. Eramo, S. C. Broome, J. S. T. Woodhead, R. F. D′souza, J. Gallagher, E. Salimova, C. Huang, R. B. Schittenhelm, J. Sadoshima, M. J. Watt, C. A. Mitchell and T. Tiganis, Skeletal muscle NOX4 is required for adaptive responses that prevent insulin resistance, Sci. Adv., 2021, 7, eabl4988 CrossRef.
- R. P. Ferreira and J. A. Duarte, Protein turnover in skeletal muscle: looking at molecular regulation towards active lifestyle, Int. J. Sports Med., 2023, 44, 763–777 Search PubMed.
- P. Llanos and J. Palomero, Cells, 2022, 11, 4008 CrossRef PubMed.
- B. B. Dokken, V. Saengsirisuwan, J. S. Kim, M. K. Teachey and E. J. Henriksen, Oxidative stress-induced insulin resistance in rat skeletal muscle: role of glycogen synthase kinase-3, Am. J. Physiol. Endocrinol. Metab., 2008, 294, 615–621 CrossRef.
- W. Matheny, E. Merritt, S. V. Zannikos, R. P. Farrar and M. L. Adamo, Serum IGF-I-deficiency does not prevent compensatory skeletal muscle hypertrophy in resistance exercise, Exp. Biol. Med., 2009, 234, 164–170 CrossRef PubMed.
- B. B. Kahn, T. Alquier, D. Carling and D. G. Hardie, Review AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism, Cell Metab., 2005, 1, 15–25 CrossRef.
- R. Iorio, G. Celenza and S. Petricca, Mitophagy: molecular mechanisms, new concepts on parkin activation and the emerging role of AMPK/ULK1 axis, Cells, 2021, 11, 30 CrossRef.
- T. S. Bowen, G. Schuler and V. Adams, Skeletal muscle wasting in cachexia and sarcopenia: Molecular pathophysiology and impact of exercise training, J. Cachexia Sarcopenia Muscle, 2015, 6, 197–207 CrossRef PubMed.
- M. Sandri, Signaling in muscle atrophy and hypertrophy, Physiology, 2008, 23, 160–170 CrossRef.
- N. El Shafey, M. Guesnon, F. Simon, E. Deprez, J. Cosette, D. Stockholm, D. Scherman, P. Bigey and A. Kichler, Inhibition of the myostatin/Smad signaling pathway by short decorin-derived peptides, Exp. Cell Res., 2016, 341, 187–195 CrossRef.
- A. P. Sharples, D. C. Hughes, C. S. Deane, A. Saini, C. Selman and C. E. Stewart, Longevity and skeletal muscle mass: the role of IGF signalling, the sirtuins, dietary restriction and protein intake, Aging Cell, 2015, 14, 511–523 CrossRef PubMed.
- W. M. Passtoors, M. Beekman, J. Deelen, R. van der Breggen, A. B. Maier, B. Guigas, E. Derhovanessian, D. van Heemst, A. J. M. de Craen, D. A. Gunn, G. Pawelec and P. E. Slagboom, Gene expression analysis of mTOR pathway: association with human longevity, Aging Cell, 2013, 12, 24–31 CrossRef PubMed.
- P. E. Slagboom, M. Beekman, W. M. Passtoors, J. Deelen, A. A. M. Vaarhorst, J. M. Boer, E. B. Van Den Akker, D. Van Heemst, A. J. M. De Craen, A. B. Maier, M. Rozing, S. P. Mooijaart, B. T. Heijmans and R. G. J. Westendorp, Genomics of human longevity, Philos. Trans. R. Soc., B, 2011, 366, 35–42 CrossRef PubMed.
- D. Papadopoli, K. Boulay, L. Kazak, M. Pollak, F. A. Mallette, I. Topisirovic and L. Hulea, mTOR as a central regulator of lifespan and aging, F1000Research, 2019, 8, F1000 Faculty Rev–998 Search PubMed.
- A. Saini, N. Al-Shanti, A. P. Sharples and C. E. Stewart, Sirtuin 1 regulates skeletal myoblast survival and enhances differentiation in the presence of resveratrol, Exp. Physiol., 2012, 97, 400–418 CrossRef.
- D. Tran, J. Bergholz, H. Zhang, H. He, Y. Wang, Y. Zhang, Q. Li, J. L. Kirkland and Z. X. Xiao, Insulin-like growth factor-1 regulates the SIRT1-p53 pathway in cellular senescence, Aging Cell, 2014, 13, 669–678 CrossRef PubMed.
- L. Schomburg and U. Schweizer, Hierarchical regulation of selenoprotein expression and sex-specific effects of selenium, Biochim. Biophys. Acta, 2009, 1790, 1453–1462 CrossRef.
- K. Weaver and R. Skouta, The selenoprotein glutathione peroxidase 4: from molecular mechanisms to novel therapeutic opportunities, Biomedicines, 2022, 10, 891 CrossRef.
- Y. Xie, R. Kang, D. J. Klionsky and D. Tang, GPX4 in cell death, autophagy, and disease, Autophagy, 2023, 19, 2621–2638 CrossRef PubMed.
- L. Schomburg, Selenoprotein P - Selenium transport protein, enzyme and biomarker of selenium status, Free Radicals Biol. Med., 2022, 191, 150–163 CrossRef.
- A. Mizuno, T. Toyama, A. Ichikawa, N. Sakai, Y. Yoshioka, Y. Nishito, R. Toga, H. Amesaka, T. Kaneko, K. Arisawa, R. Tsutsumi, Y. Mita, S. I. Tanaka, N. Noguchi and Y. Saito, An efficient selenium transport pathway of selenoprotein P utilizing a high-affinity ApoER2 receptor variant and being independent of selenocysteine lyase, J. Biol. Chem., 2023, 299, 105009 CrossRef.
- K. Jotty, M. L. Ojeda, F. Nogales, J. M. Rubio, M. L. Murillo and O. Carreras, Selenium tissue distribution changes after ethanol exposure during gestation and lactation: selenite as a therapy, Food Chem. Toxicol., 2009, 47, 2484–2489 CrossRef PubMed.
- M. C. K. Severinsen, C. Schéele and B. K. Pedersen, Exercise and browning of white adipose tissue - a translational perspective, Curr. Opin. Pharmacol., 2020, 52, 18–24 CrossRef.
- F. T. Crews, R. P. Vetreno, M. A. Broadwater and D. L. Robinson, Adolescent alcohol exposure persistently impacts adult neurobiology and behavior, Pharmacol. Rev., 2016, 68, 1074–1109 CrossRef PubMed.
- G. I. Caruso, S. F. Spampinato, G. Costantino, S. Merlo and M. A. Sortino, SIRT1-dependent upregulation of BDNF in human microglia challenged with Aβ: an early but transient response rescued by melatonin, Biomedicines, 2021, 9, 466 CrossRef.
- G. S. de Castro, J. Correia-Lima, E. Simoes, C. E. Orsso, J. Xiao, L. R. Gama, S. P. Gomes, D. C. Gonçalves, R. G. F. Costa, K. Radloff, U. Lenz, A. E. Taranko, F. C. Bin, F. B. Formiga, L. G. L. de Godoy, R. P. de Souza, L. H. A. Nucci, M. Feitoza, C. C. de Castro, F. Tokeshi, P. S. M. Alcantara, J. P. Otoch, A. F. Ramos, A. Laviano, D. Coletti, V. C. Mazurak, C. M. Prado and M. Seelaender, Myokines in treatment-naïve patients with cancer-associated cachexia, Clin. Nutr., 2021, 40, 2443–2455 CrossRef PubMed.
- M. L. Ojeda, F. Nogales, J. A. Carrasco-López, M. D. C. Gallego-López, O. Carreras, A. Alcudia and E. Pajuelo, Microbiota-liver-bile salts axis, a novel mechanism involved in the contrasting effects of sodium selenite and selenium-nanoparticle supplementation on adipose tissue development in adolescent rats, Antioxidants, 2023, 12, 1123 CrossRef.
- A. Vonlaufen, J. S. Wilson, R. C. Pirola and M. V. Apte, Role of alcohol metabolism in chronic pancreatitis, Alcohol Res. Health, 2007, 30, 48–54 Search PubMed.
- N. Eguchi, N. D. Vaziri, D. C. Dafoe and H. Ichii, The role of oxidative stress in pancreatic β cell dysfunction in diabetes, Int. J. Mol. Sci., 2021, 22, 1–18 Search PubMed.
- R. H. Wang, X. Xu, H. S. Kim, Z. Xiao and C. X. Deng, SIRT1 deacetylates FOXA2 and is critical for Pdx1 transcription and β-cell formation, Int. J. Biol. Sci., 2013, 9, 934 CrossRef PubMed.
- V. Singh and S. Ubaid, Role of silent information regulator 1 (SIRT1) in regulating oxidative stress and inflammation, Inflammation, 2020, 43, 1589–1598 CrossRef CAS.
- G. J. Prud'Homme, Y. Glinka, O. Udovyk, C. Hasilo, S. Paraskevas and Q. Wang, GABA protects pancreatic beta cells against apoptosis by increasing SIRT1 expression and activity, Biochem. Biophys. Res. Commun., 2014, 452, 649–654 CrossRef.
- L. Bordone, M. C. Motta, F. Picard, A. Robinson, U. S. Jhala, J. Apfeld, T. McDonagh, M. Lemieux, M. McBurney, A. Szilvasi, E. J. Easlon, S. J. Lin and L. Guarente, Sirt1 regulates insulin secretion by repressing UCP2 in pancreatic β cells, PLos Biol., 2006, 4, 210–220 CrossRef CAS.
- K. Bloch, E. Shichman, M. Vorobeychik, D. Bloch and P. Vardi, Catalase expression in pancreatic alpha cells of diabetic and non-diabetic mice, Histochem. Cell Biol., 2007, 127, 227–232 CrossRef CAS PubMed.
- R. Tatsuki, K. Satoh, A. Yamamoto, K. Hoshi and K. Ichihara, Lipid peroxidation in the pancreas and other organs in streptozotocin diabetic rats, Jpn. J. Pharmacol., 1997, 75, 267–273 CrossRef CAS PubMed.
- M. Tiedge, S. Lortz, R. Munday and S. Lenzen, Complementary action of antioxidant enzymes in the protection of bioengineered insulin-producing RINm5F cells against the toxicity of reactive oxygen species, Diabetes, 1998, 47, 1578–1585 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2024 |