
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAntirotaviral activity of dairy byproducts enriched in fractions from hyperimmune bovine colostrum: the effect of thermal and high hydrostatic pressure treatments†
Dimitra
Graikini
 ab,
Laura
García
a,
Inés
Abad
ab,
María
Lavilla
c,
Eduardo
Puértolas
c,
María Dolores
Pérez
ab and
Lourdes
Sánchez
*ab
ab,
Laura
García
a,
Inés
Abad
ab,
María
Lavilla
c,
Eduardo
Puértolas
c,
María Dolores
Pérez
ab and
Lourdes
Sánchez
*ab
aDepartmento de Producción Animal y Ciencia de los Alimentos, Universidad de Zaragoza, Zaragoza, Spain. E-mail: lousanchez@unizar.es
bInstituto Agroalimentario de Aragon (IA2), Zaragoza, Spain
cAZTI-BRTA, Food Research-Parque Tecnológico de Bizkaia, Astondo Bidea, edificio 609, 48160 Derio, Spain
First published on 6th February 2024
Abstract
Nowadays, rotaviruses remain a major health burden, especially in developing countries, and strategies complementary to vaccination are needed. In this view, dairy fractions have attracted great scientific interest, due to their high content of bioactive compounds. The objective of this study was to evaluate the antiviral activity of whey and buttermilk enriched in proteins from hyperimmune bovine colostrum (HBC) against rotavirus. The enriched fractions were spray-dried and subsequently tested for their neutralizing activity against the bovine rotavirus WC3 strain in vitro, using differentiated Caco-2/TC7 cells. The highest antirotaviral activity was observed when whey and buttermilk were enriched in purified immunoglobulin G (IgG), showing complete rotavirus neutralization at concentrations of 3 and 6 mg mL−1 for whey and buttermilk, respectively. Additionally, the use of a crude immunoglobulin fraction also gave satisfactory results. The inhibitory activities of all samples significantly decreased after the application of heat, except for the IgG-enriched buttermilk which showed a slight increase of activity following the application of short-time treatments (75 or 85 °C for 20 s). This sample also showed a significant increase of activity (13%) after the application of low-intensity high hydrostatic pressure treatment (400 MPa for 5 min). The maximum loss of bioactivity was observed at 600 MPa for 10 min (31 and 20% for whey- and buttermilk-based formulas, respectively). This study provides relevant information on the potential of whey, buttermilk, and HBC to be part of functional products as complementary strategies to combat rotavirus infections.
1. Introduction
Bovine milk represents an important source of nutrition, and it is probably the most versatile food that is consumed either as is or as processed dairy products. Although dairies are major economic drivers in rural areas, they are responsible for producing a significant volume of waste, mainly in the form of whey and buttermilk.1 Whey is generated during the cheese-making process, specifically during the coagulation stage of milk caseins. It accounts for 85–95% of the total milk volume and it can be differentiated into sweet whey (pH 6.2–6.4) and acid whey (pH 4.6–5.0) depending on the process used for casein coagulation.2 It is estimated that in the production of 1 kg of cheese, around 9 L of whey are obtained, which means that the world dairy industry generates about 145 m tons of this byproduct per year. Sweet whey is composed of lactose (74%), proteins (10%), minerals (8%), minor components (7%) and fat (1%).3 Because of the high lactose content in its dry matter, whey has a high biochemical oxygen demand (BOD of 40–60 g L−1) and a high chemical oxygen demand (COD of 50–80 g L−1).4,5 Therefore, it has been estimated that the waste load of whey is 100–175 times higher than that of a similar volume of domestic wastewater.6,7On the other hand, buttermilk is the aqueous phase released during cream churning for butter production, containing all the water-soluble components of skimmed milk, such as protein, lactose and minerals.8 It is also rich in materials derived from the milk fat globule membrane (MFGM), which ruptures during the churning step and migrates mainly to the released aqueous phase.9 Based on the weight of dry matter, the main components of sweet buttermilk are protein (31.5–33.1%), lactose (48.7–53.8%) and fat (5.7–13.1%).10 Buttermilk was considered a low-valued byproduct in the butter industry, mainly due to its low stability towards oxidation, and traditionally a part was used to be dried for animal feed.11
In the past years, various strategies have been employed for the revalorization of whey and buttermilk. Due to their composition, several successful attempts have been made for their use as emulsifiers, stabilizers, aroma and flavor contributors, and also as sources of bioactive ingredients.1,12 In recent years, the reports that indicate that proteins and peptides from whey and buttermilk have health promoting benefits beyond basic infant nutrition have notably increased. Emerging findings from in vitro, animal, and a limited number of human studies suggest that the enrichment of infant formulas with whey isolates and/or the MFGM from buttermilk gives rise to products more similar to human milk.13–15
The composition of bovine milk varies greatly during lactation, from colostrum to mature milk.16 Colostrum, the secretion of the mammary gland in the first few days after calving, is responsible for supplying the newborn with nutrients, immunological protection against pathogens, and growth and development factors.17 Among its immunological and biologically active components, the presence of several types of immunoglobulins (Igs) has been well-documented.18,19 The Igs present in bovine colostrum are IgG, IgA and IgM, with IgG being the predominant type.20 In this context, it has been found that vaccination of cows during gestation against human pathogens results in the production of specific polyclonal antibodies against them and their secretion into colostrum. This hyperimmune bovine colostrum (HBC) has been previously applied as a treatment for various gastrointestinal infections both in animal21,22 and human studies.23 In this regard, Civra et al. (2019)24 demonstrated a protective effect of HBC from cows vaccinated with bovine rotavirus (RV) against different human RV genotypes. By vaccinating the cows with a conventional veterinary vaccine against bovine RV rather than a targeted pathogen, they generated HBC on a large scale that could be used as a functional ingredient to prevent and treat RV infections.24
RV is the worldwide leading etiologic agent of severe gastroenteritis in infants and children under 5 years of age25,26 and the third leading pathogen associated with infant mortality as estimated by the World Health Organization (WHO) and the Centers for Disease Control and Prevention (CDC).27,28 So far, vaccination against RV has undoubtedly marked a significant improvement in combating the disease.29 However, logistical and economic difficulties, and their limited availability, diminish their efficacy in developing countries.30 Therefore, the search for alternative antirotaviral agents is a necessity. In this context, the use of bioactive milk fractions and proteins represents a promising avenue for intervention.
In any case, for all products to be used in the food industry, the hygienic quality must be ensured to minimize the risks associated with biological contaminants.31 Currently, thermal treatments, such as pasteurization and UHT sterilization, are the main procedures to achieve microbial inactivation. Nevertheless, there are several studies reporting that heat treatments usually applied to milk and milk derivatives may lead to an alteration of both the bioactive and organoleptic properties, affecting primarily the protein fraction.32,33 In this sense, high hydrostatic pressure (HHP) treatment has emerged as an alternative technology to inactivate pathogens, while maintaining most sensorial and nutritional properties, causing minor degradation of proteins and vitamins relative to thermal treatments.34 The process uniformity is one of the best characteristics of HHP treatment that makes it globally acceptable.35
The objective of this study was to evaluate the activity of whey and buttermilk enriched in Igs isolated from HBC to inhibit the infection by bovine RV of Caco-2/TC7 cells differentiated as enterocytes. Furthermore, the effect of industrial preservation technologies such as thermal pasteurization and HHP treatment on the antiviral activity of those preparations was investigated. This study will contribute to expanding the current knowledge about the exploitation of dairy bioactive components and fractions as complementary strategies for the treatment of rotaviral diseases.
2. Materials and methods
2.1 Obtaining milk fractions
Raw bovine milk was provided by the dairy industry Villacorona (El Burgo de Ebro, Zaragoza, Spain). The quality of milk was verified after reception by evaluating the pH, acidity, fat percentage, alkaline phosphatase and lactoperoxidase activity, and afterwards it was processed at the Food Science and Technology Pilot Plant of the University of Zaragoza. To obtain whey, milk was initially heated at 50 °C and then skimmed using an ARR-DES 125 model centrifuge (Suministros Químicos Arroyo, Santander, Spain) to separate the fat fraction. Next, caseins of the skimmed milk were precipitated by coagulation for 45 min at 35 °C, by adding bovine rennet (Laboratorios Abiasa, Pontevedra, Spain) and a 20% solution of calcium chloride (Laboratorios Arroyo, Santander, Spain) at final concentrations of 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :15000 (v/v) and 1:8000 (v/v), respectively. Finally, the whey was obtained by decantation and subsequent filtration through cheesecloth and glass wool to eliminate the remaining small curd grains. To obtain buttermilk, the cream previously isolated, with a fat content of approximately 40% (v/v), was kept overnight at 4 °C. Afterwards, it was subjected to mechanical stirring with a Philips Cucina mixer (Amsterdam, the Netherlands) until the formation of butter grains by agglomeration of the fat globules and liberation of buttermilk, which was filtered through cheesecloth and glass wool to eliminate remaining butter grains. Whey and buttermilk were stored at −20 °C either in liquid or freeze-dried form until further use. In the case of freeze-drying, the two fractions were first dialyzed against 25 mM ammonium bicarbonate, pH 7.4, and then freeze dried using a Heto PowerDry DW8 (Thermo Fisher Scientific, Rockford, IL, USA).
:15000 (v/v) and 1:8000 (v/v), respectively. Finally, the whey was obtained by decantation and subsequent filtration through cheesecloth and glass wool to eliminate the remaining small curd grains. To obtain buttermilk, the cream previously isolated, with a fat content of approximately 40% (v/v), was kept overnight at 4 °C. Afterwards, it was subjected to mechanical stirring with a Philips Cucina mixer (Amsterdam, the Netherlands) until the formation of butter grains by agglomeration of the fat globules and liberation of buttermilk, which was filtered through cheesecloth and glass wool to eliminate remaining butter grains. Whey and buttermilk were stored at −20 °C either in liquid or freeze-dried form until further use. In the case of freeze-drying, the two fractions were first dialyzed against 25 mM ammonium bicarbonate, pH 7.4, and then freeze dried using a Heto PowerDry DW8 (Thermo Fisher Scientific, Rockford, IL, USA).
2.2 Isolation and purification of Igs from HBC
The isolation of IgG was carried out starting from a frozen sample of HBC obtained from vaccinated dairy cattle and following the methodology proposed by Mainer et al. (1997),36 with slight modifications. For this, HBC was skimmed by centrifugation at 2500g for 15 min at 4 °C and then, bovine rennet was added in a 1:15000 (v/v) ratio to precipitate the caseins and obtain the whey. Afterwards, ammonium sulfate was added at a concentration of 60% (v/v) and incubated overnight at 4 °C, to induce selective Ig precipitation. Then, the mixture was centrifuged at 3000 rpm for 15 min at 4 °C, obtaining a supernatant and a precipitate containing colostral Igs. The supernatant was removed, and the precipitate was resuspended in distilled water, and dialyzed against 0.01 M potassium phosphate buffer, pH 7.7, for 2 days. The fraction obtained after dialysis was named the “immunoglobulin precipitate fraction (IPF)”. IgG was further purified by subjecting the IPF to ion exchange chromatography using the ÄKTA™ start system from GE Healthcare (Uppsala, Sweden) with a HiTrap™ DEAE FF column (5 mL) (GE Healthcare). Elution of retained proteins was carried out with a stepwise gradient of 0.01, 0.02, and 0.06 M phosphate buffer, pH 7.4, 7.2, and 7.0, respectively, eluting IgG in the third step. The fractions obtained were characterized by SDS-PAGE, and those containing IgG were pooled and dialyzed against 25 mM ammonium bicarbonate solution. Finally, they were freeze-dried and stored at −20 °C until use.
2.3 Determination of the protein content
The bicinchoninic acid assay (BCA) method was used to determine the protein content of samples. The assay was carried out using a commercial Pierce™ BCA protein assay kit (Thermo Fisher Scientific, Rockford, USA) following the manufacturer's instructions. In the case of purified IgG, the determination of the protein content was performed using a molar extinction coefficient of 1.4 M−1 cm−1.372.4 Preparation of enriched dairy fractions
Whey and buttermilk in liquid form were both enriched with Igs obtained from HBC in four different ways. Three samples were prepared using IPF, at three different concentrations: 0.5, 1, and 2 mg of total IPF protein per mL of whey or buttermilk. Moreover, an enriched solution was prepared with purified IgG, at a concentration of 0.5 mg of protein per mL of whey or buttermilk. The enriched preparations were subjected to spray drying in a Mini Spray Dryer B-290 (BUCHI, Flawil, Switzerland). The pumping speed was 35%, with an approximate flow of 12 mL min−1, an inlet temperature of 150 °C and 100% suction. The powders obtained were vacuum sealed in polyethylene bags and stored at −20 °C until use. In the following assays, the spray-dried enriched dairy preparations were reconstituted in sterile phosphate-buffered saline buffer (PBS) composed of 0.14 M NaCl, 2.6 mM KCl, 8.1 mM Na2HPO4, and KH2PO4, pH 8.5, and the different concentrations were henceforth expressed in the weight of powder per mL of PBS.2.5 Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
SDS-PAGE was carried out according to the procedure described by Laemmli (1970).38 The samples to be characterized were diluted 1:1 with a 0.12 M Tris–HCl buffer, pH 6.8, containing a final concentration of glycerol of 20% (v/v), 4% (w/v) SDS and 0.02% (w/v) bromophenol blue. For the electrophoresis performed in reducing conditions, β-mercaptoethanol was added to the buffer at a final concentration of 10% (v/v). Subsequently, all samples were heated at 100 °C for 5 min. The electrophoresis process was carried out on 4–20% polyacrylamide gels (Mini-Protean TGX, Bio-Rad Laboratories, Hercules, CA, USA). The molecular weight marker used was a PageRuler™ Prestained Protein Ladder, from 10 to 180 kDa (Thermo Fisher Scientific). The electrophoresis was run at 180 mA for 35 min and the gels were stained with Coomassie Brilliant Blue R-250 (Serva blue R, Serva Feinbiochemica GmbH & Co., Heidelberg, Germany), according to standard procedures.
2.6 Heat treatments
Aliquots (200 μL) of dairy samples were placed in glass capillary tubes (1.55 × 70 mm) that were sealed by using a small gas torch. Four replicates (n = 4) for each processing condition and sample were analyzed. Next, the tubes were placed in a preheated thermostatic bath (Unitronic 200, J. P. Selecta, Barcelona, Spain) (±0.1 °C). Heat treatment was monitored by controlling temperatures inside the water bath with a digital thermometer. The treatments applied were 75 °C for 20 s or 10 min, and 85 °C for 20 s or 10 min. After the treatment, the samples were immediately cooled in an ice bath and stored at −20 °C until use. The concentrations in which the samples were treated were 10.6, 27.2, 3.0 and 3.0 mg of powdered product per mL of PBS for whey, buttermilk, IgG-enriched whey and IgG-enriched buttermilk, respectively. For the subsequent in vitro assays, several heat-treated samples were used in a selected final concentration that showed neutralization values >90% for each non-treated control. These concentrations corresponded to 5.3, 13.6, 1.5 and 1.5 mg of powdered product per mL of PBS for whey, buttermilk, IgG-enriched whey and IgG-enriched buttermilk, respectively.2.7 HHP treatments
Spray-dried whey and buttermilk enriched in purified IgG were reconstituted in PBS at a concentration of 6 mg mL−1. HHP treatments were performed in a 55 L high-pressure unit WAVE 6000/55HT (Hiperbaric, Burgos, Spain). This system includes accurate control of the vessel wall, plugs and inlet water temperatures to ensure a constant temperature during the treatment. Samples (1.5 mL) were packed in Eppendorf tubes and isolated from pressurized water by vacuum packaging in polyethylene bags. Three replicates (n = 3) for each processing condition and sample were conducted. Untreated samples were similarly prepared as the control. The pressure values applied were 400, 500, and 600 MPa, with a pressure holding time of 5 and 10 min. The temperature of the inlet water was set for each of the three pressures at 15 °C, 13 °C and 10 °C, respectively, and therefore, the temperature of the samples under pressure remained always below 27.5 °C,39,40 avoiding heat induced side-effects on protein structures. The rate of pressure increase was about 3 MPa s−1 (∼170 MPa min−1), and the pressure release was immediate (<2 s) at the end of the treatment. After HHP treatments, the samples were stored refrigerated (4 °C ± 0.5 °C) for less than 24 h until use. The HHP treated preparations were tested for RV neutralization activity at a final concentration of 1.5 mg powder per mL.2.8 Determination of IgG by ELISA
The quantification of IgG in the enriched formulas, before and after spray drying, was performed by using the Calokit Bovino (ZEULAB, Zaragoza, Spain). This is a sandwich ELISA assay based on specific detection of bovine IgG in milk matrixes. Briefly, 50 μL of appropriately diluted samples or standards were added to each well of a microtiter 96-well plate previously coated with specific antibodies against IgG. The plate was incubated for 30 min at 25 °C and afterwards, it was washed four times with 300 μL of distilled water. Next, the plate was incubated at 25 °C for 30 min with 50 μL of conjugate solution. The plate was then washed four times with distilled water and 50 μL of substrate solution was added and incubated at 25 °C for 30 min. The enzymatic reaction was stopped by adding 50 μL of stop solution and the absorbance was immediately read at 450 nm in a Labsystem Multiskan MS Original 352 plate reader.2.9 Proteomic analysis
To assess the effect of the HHP treatment on the protein profile of the tested samples, proteomic analysis was done at the Proteomics Platform of CIBA (IACS-Universidad de Zaragoza, Spain). Protein identification was performed by analyzing the peptide fingerprint (MS) and fragmentation spectra (MS/MS) of the peptides generated after trypsin digestion of polyacrylamide gel bands in an automatic digester (Intavis, Bioanalytical Instruments, Cologne, Germany). Briefly, the bands were washed with water, ammonium bicarbonate (100 mM) and acetonitrile. Samples were then reduced by incubation with DTT (10 mM) at 60 °C for 45 min and alkylated by incubation with iodoacetamide (50 mM) at room temperature for 30 min in darkness. Finally, the proteins were digested with trypsin overnight at 37 °C (5 μg mL−1, Trypsin Gold, Promega, WI, USA). Digestion was stopped by adding 0.5% TFA (trifluoroacetic acid), and tryptic peptides were extracted sequentially with an increase in concentrations of ACN in water. Samples were evaporated in a concentrator and resuspended in 2% ACN 0.1% formic acid (FA).Proteins were identified on a hybrid triple quadrupole/linear ion trap mass spectrometer (6500QTRAP+, Sciex, Foster City, CA, USA) coupled to a nano/micro-HPLC (Eksigent LC425, Sciex). Preconcentration and desalting of the samples were performed online using a C18 precolumn (Luna® 0.3 mm id, 20 mm, 5 μm particle size, Phenomenex, CA, USA) at 10 μL min−1 for 5 min. Peptide separation was performed using a C18 column (Luna Omega® 0.3 mm id, 150 mm, 3 μm particle size, Phenomenex, CA, USA), at 5 μL min−1. The column was kept at 40 °C. The elution gradient performed was 5 to 35% ACN (0.1% FA) for 30 min. A mass spectrometer was interfaced with an ESI (Turbo V™) source using a 25 μm ID hybrid electrode, operated in the positive mode. The parameters of the source are as follows: a collision energy of 10 eV, capillary voltage of 5000 V, declustering potential of 85 V, and curtain gas and ion source (nitrogen) of 10.42 kPa. Analyses were performed using an information-dependent acquisition method with the following steps: first a scan of enhanced single mass spectra (EMS, 400–1400 m/z) from which the 5 most intense peaks were subjected to enhanced product ion scanning [EPI (MS/MS)].
The search engine used is MASCOT (MatrixScience, UK) with public databases of protein sequences (Swiss-Prot, NCBI, etc.) according to the taxonomy registered in the application. The search parameters used were missed cleavages 2, fixed carbamidomethyl modification (cysteines) and a mass tolerance of 0.5 Da peptides and fragments.
2.10 Cell culture and virus propagation
The human adenocarcinoma cell line Caco-2/TC7 (kindly donated by the Department of Physiology at the Veterinary Faculty of the University of Zaragoza) was cultured in DMEM supplemented with 10% (v/v) foetal bovine serum (FBS), 1% (v/v) L-glutamine 2 mM, 1% (v/v) non-essential amino acid solution (NEAA), 1% (v/v) antibiotic solution (100 units mL−1 penicillin, 100 μg mL−1 streptomycin) and 1 μg mL−1 amphotericin B. The rhesus monkey epithelial cell line MA104 (ATCC CRL-2378) was cultured in MEM supplemented with 10% (v/v) FBS, 1% (v/v) 2 mM L-glutamine, 1% (v/v) antibiotic solution (100 units mL−1 penicillin, 100 μg mL−1 streptomycin) and 0.25 μg mL−1 amphotericin B. Both cell types were maintained in 25 cm2 culture vessels at 37 °C in a Heraeus B5060 EK/CO2 thermostatic incubator with 5% CO2. All cell culture media and supplements were purchased from Gibco (Life Technologies Corporation, Paisley, UK).The bovine RV WC3 strain (ATCC VR-2102) was propagated on the MA104 cell line according to previously described procedures.41 Briefly, confluent 75 cm2 cell flasks were infected at a multiplicity of infection (MOI, ratio virus per cell) of 0.1 with an aliquot of a RV suspension. Once the cytopathic effect was established, the RV was harvested by subjecting the lysate to three cycles of freezing (−80 °C) and thawing (room temperature) and subsequent centrifugation at 300g for 15 min at 4 °C. The supernatant, considered as the infective viral suspension, was titrated according to the procedures described by Arnold, Patton & McDonald (2009),42 aliquoted in cryovials and stored at −80 °C until further use. The RV multiplicity of infection that was used in the following in vitro assays was 0.02.
2.11 Cell viability assay
To evaluate the effect of dairy preparations on the viability of Caco-2/TC7 cells, the CellTiter 96® AQueous One Solution Cell Proliferation Assay kit (Promega, Madison, USA) was used. The assay, based on a colorimetric reaction in which the tetrazolium compound MTS is bioreduced by metabolically active cells, was performed as previously described by Graikini et al. (2023).412.12 RV neutralization assay
The RV neutralization capacity of the samples was tested using the human cell line Caco-2/TC7, as it is considered an excellent model of the human intestinal epithelium.43 Briefly, Caco-2/TC7 cells were seeded in 96-well plates at a density of 1.4 × 104 cells per cm2 and cultured for 15 days until the differentiation stage, with a morphology similar to functional enterocytes.44 Before the assay, the cells were serum starved for 2 h using basic DMEM, supplemented with 1% (v/v) L-glutamine and 1% (v/v) NEAA, to avoid interference with FBS. Meanwhile, appropriate dilutions of samples (in basic DMEM) were mixed 1:1 (v/v) with a trypsin-activated RV suspension and incubated for 1 h at 37 °C. The basic DMEM was used as a negative control and the activated RV suspension diluted 1:1 (v/v) with basic DMEM was used as a positive control of infection. After the incubation period, the samples were transferred into the plate containing the previously serum-starved cells and incubated at 37 °C for 1 h for RV adsorption. Next, plates were added with 100 μL of DMEM per well containing 2 μg mL−1 trypsin and 6% (v/v) of FBS and incubated at 37 °C in 5% CO2 for 12 h under gentle rotation. Subsequently, virus-infected cells were detected by indirect immunofluorescence (Section 2.14).
2.13 Cell receptor blocking assay
The interaction of purified IgG with Caco-2/TC7 cells was additionally studied to evaluate the influence that it may have on RV infection. Similar to the neutralization assay, Caco-2/TC7 cells were seeded in 96-well plates and grown until differentiation. Then, the culture medium was removed, and the cells were incubated with serum-free medium for 2 h. Next, 50 μL of samples at different concentrations were added and the plate was incubated for 1 h at 4 °C, to prevent the samples from penetrating the cells by endocytosis. Afterwards, the wells were washed three times with 120 μL per well of PBS and infected with the suspension of bovine RV WC3 strain, incubating for 1 h at 37 °C. After washing the wells with 120 μL per well of PBS, 100 μL of DMEM supplemented with 6% (v/v) of FBS and 2 μg mL−1 trypsin were added to each well, and the plate was incubated at 37 °C for 12 h, subsequently proceeding to the determination of cell infection with RV by indirect immunofluorescence (Section 2.14).2.14 Indirect immunofluorescence
After 12 h of infection, the cells were washed with 200 μL per well of sterile PBS. Cell fixation was then carried out by adding 300 μL per well of a solution of acetone: methanol: formaldehyde (1:1:1, v/v/v) and incubating for 3 min at 4 °C. The plate was then washed twice with sterile PBS and incubated with 100 μL per well of antiserum anti-bovine RV obtained in lamb, kindly donated by Dr Snodgrass from Moredun Research Institute (Penicuik, UK), at 37 °C for 2 h under gentle agitation. Next, the wells were washed three times with sterile PBS and incubated with 100 μL per well of FITC-conjugated donkey anti-sheep IgG antibody (Sigma-Aldrich, St. Louis, MO, USA) for 1 h at 37 °C under gentle agitation. Finally, fluorescent cells were revealed using an Eclipse E400 fluorescence microscope with a Nikon FITC filter, and the Zen lite 2012 image processing software. The infectivity percentages were determined by enumerating fluorescent foci (infected cells) in each well in relation to the 100% infectivity obtained with the positive control, which consisted of the virus suspension without neutralizing/cell blocking agent.
2.15 Statistical analysis
Statistical analysis was conducted through the GraphPad Prism v8.0.2 software (GraphPad Software, San Diego, CA, USA) and the results are presented as the mean ± standard deviation. The normality of the data was tested through the Shapiro–Wilk test. Subsequently, for the variables characterized as normal, Student's t-test or one-way analysis of variance (ANOVA) followed by Dunnet's multiple comparisons test was performed. Variables that did not follow a normal distribution were analyzed using the Mann–Whitney U-test or Kruskal–Wallis non-parametric analysis followed by Dunn's multiple comparisons test. Differences with a p value ≤ 0.05 were considered statistically significant.3. Results
3.1 Obtention of IgG and characterization of enriched dairy preparations
 | ||
| Fig. 1 Isolation and characterization of IgG from HBC: (A) elution profile of the HiTrap™ DEAE FF chromatography of IPF for the isolation of IgG, with a stepwise gradient of potassium phosphate buffer (0.01, 0.02 and 0.06 M). (B) SDS-PAGE on 4–20% polyacrylamide gel: lane 1: molecular weight standard; lane 2: IPF; lane 3: fractions 2–6; lane 4: fractions 37–39; and lane 5: fractions 57–98. All samples were loaded at a concentration of 0.2 mg mL−1 and tested under reducing conditions. (C) SDS-PAGE on 4–20% polyacrylamide gel: lane 1: molecular weight standard, lane 2: IPF under non-reducing conditions (3 mg mL−1); lane 3: IPF under reducing conditions (3 mg mL−1); lane 4: purified IgG under non-reducing conditions (1 mg mL−1); and lane 5: purified IgG under reducing conditions (1 mg mL−1). | ||
Accordingly, Fig. 1B shows the electrophoretic profile of the different fractions obtained throughout the purification of IgG. As can be observed, at the final step of purification (lane 5) the intensity of bands with molecular weights of around 55 and 25 kDa is increased. These bands correspond to the heavy and light chains of IgG, respectively, which appear separated since electrophoresis was performed under reducing conditions. To visualize better the purification process, IPF obtained after precipitation with ammonium sulfate and dialysis against potassium phosphate buffer and purified IgG after lyophilization were both subjected to SDS electrophoresis performed under both reducing and non-reducing conditions (Fig. 1C). As can be observed in lanes 4 and 5, the bands corresponding to the molecular weights of purified IgG (∼160 kDa in its monomeric form and 55 and 25 kDa for the two chains in its reduced form) appear more intense and there is absence of additional bands, in comparison with the IPF profile (lanes 2 and 3). In these rows, apart from the typical IgG bands, additional trace bands of other colostrum proteins are displayed.
As seen in Table 1, the buttermilk samples demonstrated a higher concentration of total IgG in comparison with the whey samples. Within the same fraction, the non-enriched freeze-dried and spray-dried powders had the same IgG content. As expected, the enrichment of whey and buttermilk with 0.5 mg mL−1 of either IPF or IgG resulted in an increased total IgG content. This increase was similar independently of the type of the enriching factor used, IPF or IgG, with values of 131.90 and 130.30 μg mL−1 for whey and 138.50 and 138.24 μg mL−1 for buttermilk, respectively. Nonetheless, these values did not correspond to the theoretical pre-drying enrichment of the two fractions (500 μg mL−1). Furthermore, enrichment with increasing concentrations of IPF (0.5, 1 and 2 mg mL−1) did not result in an increase of the IgG estimated by ELISA.
| Fraction | IgG (μg mL−1) | Fraction | IgG (μg mL−1) |
|---|---|---|---|
| WH fd | 61.78 | BM fd | 64.52 |
| WH sd | 61.78 | BM sd | 64.87 |
| WH + 0.5 IPF | 131.90 | BM + 0.5 IPF | 138.50 |
| WH + 1 IPF | 126.44 | BM + 1 IPF | 135.47 |
| WH + 2 IPF | 123.65 | BM + 2 IPF | 130.42 |
| WH + 0.5 IgG | 130.30 | BM + 0.5 IgG | 138.24 |
Accordingly, Fig. 2 shows the electrophoretic profiles of enriched whey (A) and buttermilk (B) where bands of the typical proteins contained in whey and buttermilk can be observed. With regard to both matrixes, an overall increase of the intensity of bands corresponding to the light and heavy chains of the IgG, with molecular weights of 25 and 55 kDa, respectively, can be depicted (lanes 4, 5, 6 and 7). In the case of buttermilk (Fig. 2B), a greater intensity of the bands between 25 and 35 kDa is shown, which corresponds to caseins, proteose peptone component 3 and the light chain of IgG.
 | ||
| Fig. 2 SDS-PAGE on 4–20% polyacrylamide gel carried out under reducing conditions. (A) Whey fractions: freeze-dried whey (WH fd); whey enriched with 0.5 mg mL−1 IgG (WH + 0.5 IgG); whey enriched with 0.5 mg mL−1 IPF (WH + 0.5 IPF); whey enriched with 1 mg mL−1 IPF (WH + 1 IPF); and whey enriched with 2 mg mL−1 IPF (WH + 2 IPF). (B) Buttermilk fractions: freeze-dried buttermilk (BM fd); buttermilk enriched with 0.5 mg mL−1 IgG (BM + 0.5 IgG); buttermilk enriched with 0.5 mg mL−1 IPF (BM + 0.5 IPF); buttermilk enriched with 1 mg mL−1 IPF (BM + 1 IPF); and buttermilk enriched with 2 mg mL−1 IPF (BM + 2 IPF). Protein bands: α-lactalbumin (α-LA); β-lactoglobulin (β-LB); butyrophilin (BTN); immunoglobulin G (IgG); lactadherin (LDH); lactoferrin (LF); mucins (MUCs); proteose-peptone component 3 (PP3); bovine serum albumin (BSA); and xanthine oxidase (XOD). | ||
3.2 Antirotaviral activity
The antiviral activity of the samples was tested in vitro against the bovine RV WC3 strain. Prior to these assays, the potential cytotoxic effect that the samples might have on differentiated Caco-2/TC7 was investigated by using the MTS colorimetric assay. No significant loss of cell viability was found for any of the samples tested (see the ESI, Fig. S1 and S2†).As seen in Fig. 3, both whey and buttermilk were able to neutralize RV in a dose–response manner with IC50 values of 0.58 and 1.33 mg mL−1, respectively. Whey presented a potent activity already at a concentration of 0.4 mg mL−1 (45.5%) while complete RV neutralization was observed at a concentration of 14.3 mg mL−1. On the other hand, buttermilk at 0.4 mg mL−1 exerted 25.3% neutralization and its activity surpassed 99.8% neutralization at the two highest concentrations tested (27.3 and 45.5 mg mL−1).
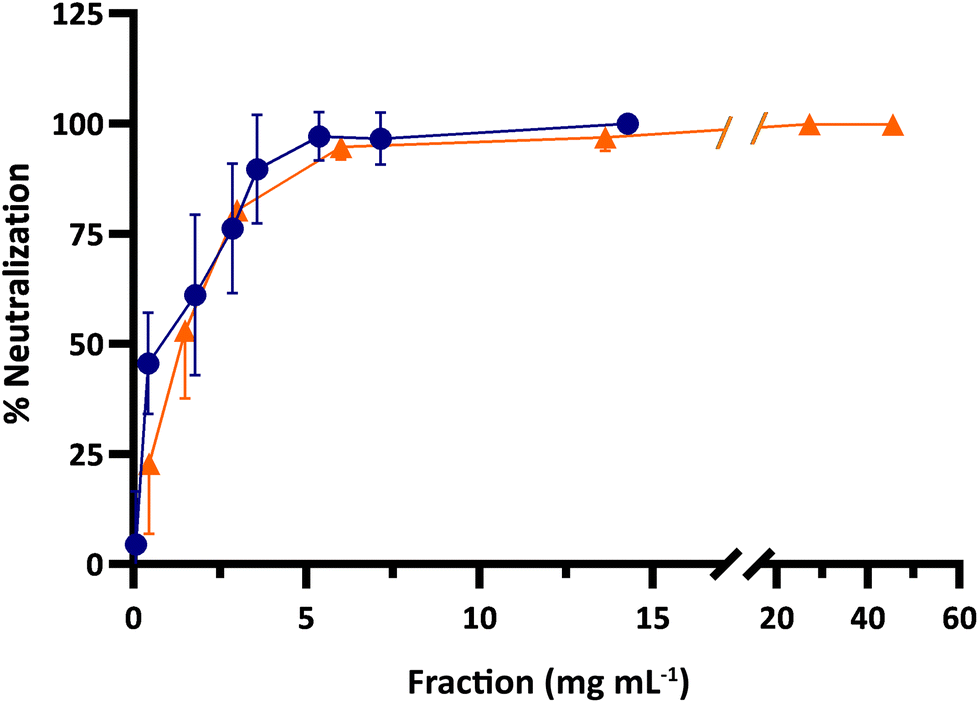 | ||
| Fig. 3 Dose–response of freeze-dried bovine whey (blue circles) and buttermilk (orange triangles) on neutralizing activity against infection of Caco-2/TC7 cells by bovine RV WC3. The results are shown as the mean ± standard deviation of duplicates from four independent experiments (n = 8). Concentration is expressed as mg of powder per mL of culture medium. | ||
The purified IgG exerted an even stronger antirotaviral activity (IC50 value of 3.55 μg mL−1). As can be seen in Fig. 4, IgG neutralized the RV infection of Caco-2/TC7 cells in a dose–response way with inhibition values over 95% at concentrations as low as 50 μg mL−1 and complete neutralization at 100 μg mL−1.
 | ||
| Fig. 4 Neutralization activity of IgG from HBC against infection of Caco-2/TC7 cells by bovine RV WC3. The results are shown as mean ± standard deviation of duplicates from three independent experiments (n = 6). Concentration is expressed as μg of protein per mL of culture medium. | ||
Since IgG presented such potent neutralization activity, we sought to investigate the involvement of additional antirotaviral mechanisms of this protein. For this, the neutralizing concentrations of IgG were evaluated in a cell receptor blocking assay and the results obtained are shown in Table 2. The inhibition of infection that occurred through cell blocking was lower at all tested concentrations compared to that demonstrated in the neutralization assay. Furthermore, we did not observe a clear dose-effect pattern. This suggests that the antirotaviral mechanism of action of IgG is mostly related to the interaction between IgG and RV (virucidal effect) and less to the interaction of these proteins with the cell receptors for RV (protective effect).
| IgG concentration (μg mL−1) | Inhibition of infection (%) | No of replicates (n) |
|---|---|---|
| 0.4 | 13.6 ± 4.40 | 7 |
| 2 | 8.81 ± 6.58 | 8 |
| 10 | 14.30 ± 9.82 | 7 |
| 50 | 13.28 ± 8.74 | 6 |
| 100 | 29.32 ± 2.34 | 6 |
| 250 | 27.13 ± 8.58 | 6 |
Fig. 5 shows the percentages of neutralization of RV infection that the different whey-based enriched fractions produced. With regard to the two treatments used for drying, in non-enriched whey, a lower neutralizing activity was observed in the case of the freeze-dried sample in comparison with the spray-dried one, although the differences between the two drying treatments were only significant at a concentration of 3 mg mL−1. At this point, it is important to stress that the initial whey used for freeze-drying was not from the same batch as that used for spray drying; therefore, such differences could be primarily attributed to the variability of protein composition rather than to the effect of the treatment itself.
 | ||
| Fig. 5 Effect of whey-based preparations on the neutralization of the infection of Caco-2/TC7 cells by bovine RV WC3. Freeze-dried whey (WH fd); spray-dried whey (WH sd); whey enriched with 0.5 mg mL−1 IPF (WH + 0.5 IPF); whey enriched with 1 mg mL−1 IPF (WH + 1 IPF); whey enriched with 2 mg mL−1 IPF (WH + 2 IPF); and whey enriched with 0.5 mg mL−1 IgG (WH + 0.5 IgG). The neutralizing activity of preparations was studied at three concentrations: 0.5, 1.5 and 3 mg of powder per mL. The results are shown as the mean ± standard deviation of duplicates from at least three independent experiments (n ≥ 6). Statistical analysis was performed using the Student's t-test in the case that data followed normal distribution and the Mann–Whitney U-test in the case that data did not follow normal distribution. #Indicates significant differences (p < 0.05) between drying treatments. *Indicates significant differences (*p < 0.05; **p < 0.01) of each preparation with respect to the non-enriched whey. | ||
With regard to the neutralization capacity of the different enriched whey samples, statistically significant differences were observed in almost all cases between the non-enriched and the enriched fractions, for all the concentrations studied. Exceptions were the enrichment with 0.5 mg of IPF, at the lowest concentration (0.5 mg powder per mL) and the enrichment with 2 mg of IPF at the highest concentration (3 mg powder per mL). The highest neutralization was demonstrated when whey was enriched with purified IgG at the highest concentrations (1.5 and 3 mg powder per mL). However, at the lowest concentration tested (0.5 mg powder per mL), it was the sample enriched with 2 mg mL−1 of IPF that showed the highest activity, with 89% neutralization, compared to the 76 and 78% percentages shown by spray-dried whey without enrichment and enriched with IgG, respectively.
The neutralization activity of the different buttermilk-based enriched preparations is shown in Fig. 6. In this case, the comparison between the two drying methods did not result in significant differences at any of the concentrations studied. Overall, the best results were obtained with the fraction enriched in purified IgG, although it should be noted that enrichment with different concentrations of IPF protein resulted in a statistically significant increase in the neutralization activity compared with non-enriched buttermilk, when tested at 1.5 mg powder per mL. However, this increase in activity was not maintained in all cases at the highest concentrations (3 and 6 mg powder per mL).
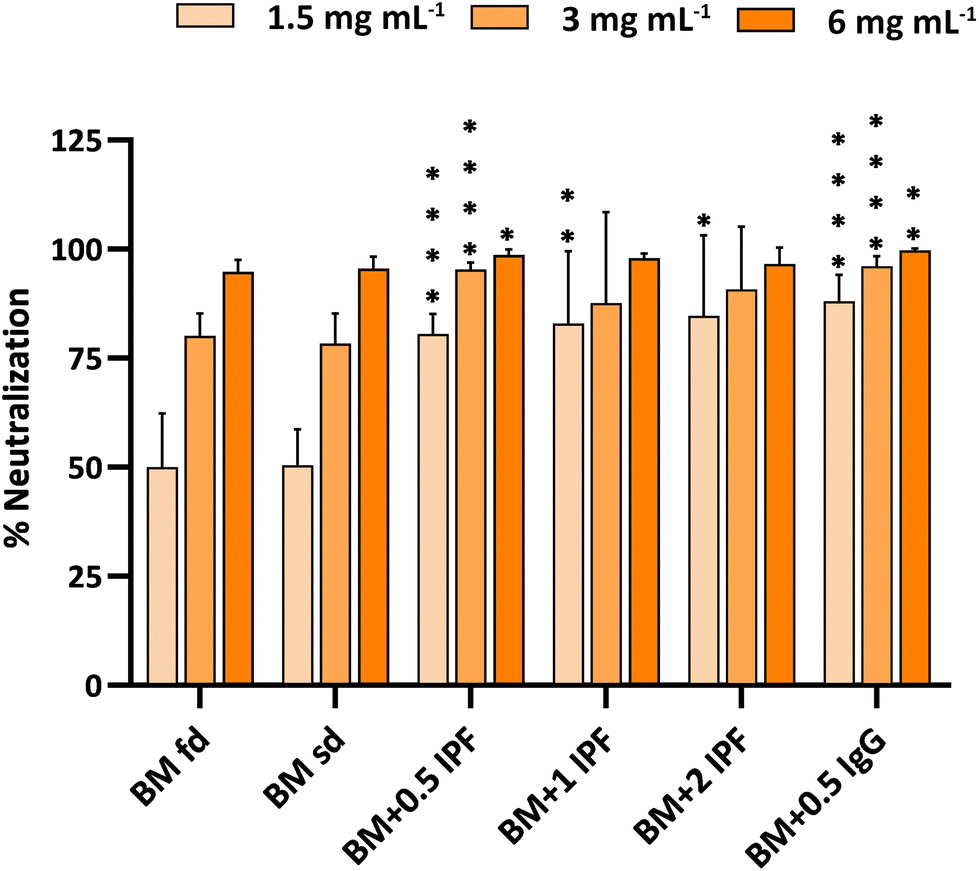 | ||
| Fig. 6 Effect of different buttermilk-based preparations on the neutralization of the infection of differentiated Caco-2/TC7 cells by WC3 RV. Freeze-dried buttermilk (BM fd); spray-dried buttermilk (BM sd); buttermilk enriched with 0.5 mg mL−1 IPF (BM + 0.5 IPF); buttermilk enriched with 1 mg mL−1 IPF (BM + 1 IPF); buttermilk enriched with 2 mg mL−1 IPF (BM + 2 IPF); and buttermilk enriched with 0.5 mg mL−1 IgG (BM + 0.5 IgG). The neutralizing activity of preparations was studied at three concentrations: 1.5, 3 and 6 mg of powder per mL. The results are shown as the mean ± standard deviation of duplicates from at least three independent experiments (n ≥ 6). Statistical analysis was performed using Student's t-test in the case that data followed normal distribution and the Mann–Whitney U-test in the case that data did not follow normal distribution. *Indicates statistically significant differences (*p < 0.05; **p < 0.01; ****p < 0.0001) of each preparation in relation to the non-enriched fraction. | ||
Overall, the enrichment with purified IgG demonstrated the best results, in both whey and buttermilk preparations, exceeding 99% neutralization at concentrations of 3 and 6 mg powder per mL, respectively. Furthermore, whey turned out to be a better enrichment medium than buttermilk, having a higher percentage of neutralization at lower concentrations.
3.3 Effect of technological treatments on the bioactivity of the enriched dairy fractions
| Treatment | Neutralization (%) | |||
|---|---|---|---|---|
| WH fd | BM fd | WH + 0.5 IgG | BM + 0.5 IgG | |
| a Indicates significant differences (p < 0.0001) of heat-treated samples in relation to the corresponding non-treated control. The concentration of IgG is expressed in mg mL−1. | ||||
| 75 °C for 20 s | 81.23 ± 5.89 | 90.69 ± 7.67 | 92.17 ± 4.21 | 105.60 ± 4.23 |
| 75 °C for 10 min | 1.67 ± 12.94a | 17.00 ± 12.26a | 3.39 ± 16.78a | 43.64 ± 19.69a |
| 85 °C for 20 s | 29.93 ± 13.95a | 30.43 ± 14.48a | 96.47 ± 2.60 | 106.50 ± 4.04a |
| 85 °C for 10 min | 13.94 ± 13.94a | 27.34 ± 21.13a | 0.00 ± 13.82a | 0.51 ± 20.39a |
The results showed that pasteurization (75 °C for 20 s) had a mild effect on the antiviral activity of the non-enriched whey and buttermilk, causing a neutralization loss of 18.77 and 9.31%, respectively. However, this decrease in activity was not significant in either of the cases. In contrast, the application of intermediate and high intensity heat treatments (75 °C for 10 min, 85 °C for 20 s and 85 °C for 10 min), resulted in a significant decrease in the neutralization effect for both fractions, with minimum neutralization values at 1.67% and 17.00% for whey and buttermilk, respectively, when the treatment applied was of 75 °C for 10 min.
Slightly different results were observed, when assessing the effect on enriched whey and buttermilk. Application of short-time (20 s) treatments, regardless of the temperature, did not significantly affect the antiviral activity of the fractions. Interestingly, in the case of the enriched buttermilk, a slight increase of the neutralization activity was shown in relation to the non-treated preparation, for both short-time treatments. Nevertheless, when the duration of the heat treatment increased (10 min), a significant loss of neutralization activity was observed. Enriched buttermilk appeared more resistant at 75 °C for 10 min as it marked 43.64% neutralization activity in contrast to the enriched whey that lost all activity (3.39% neutralization achieved). Finally, at the maximum intensity (85 °C for 10 min) the neutralization activity of both enriched fractions disappeared.
As can be observed in Fig. 7A, the loss of antirotaviral activity of the enriched whey samples was proportional to the intensity of the applied HHP treatment. Treatments at 400 and 500 MPa did not affect the antiviral activity of the preparations at any of the times tested. A decrease in neutralization was observed when samples were treated at 600 MPa for 5 min resulting in 17% loss of activity, while the same pressure for 10 min caused a significant 32% loss which was the maximum decrease observed. SDS-PAGE analysis of HHP treated preparations performed under non-reducing conditions (Fig. 7B, lanes 1–7) revealed that the intensity of the IgG (∼160 kDa in its monomeric form) decreased progressively as the treatment intensity increased. The same effect was observed for the two bands with molecular weights of about 78 and 80 kDa, corresponding to lactoperoxidase and lactoferrin, respectively. Furthermore, an additional band with a molecular weight of around 32–34 kDa appeared in the profile of all treated samples (lanes 2–7), which was absent in the control (lane 1). Proteomic analysis of this band revealed the correspondence to β-lactoglobulin. On the other hand, electrophoresis performed under reducing conditions (Fig. 7B, lanes 9–15) did not show any differences between the HHP treated and the non-treated control.
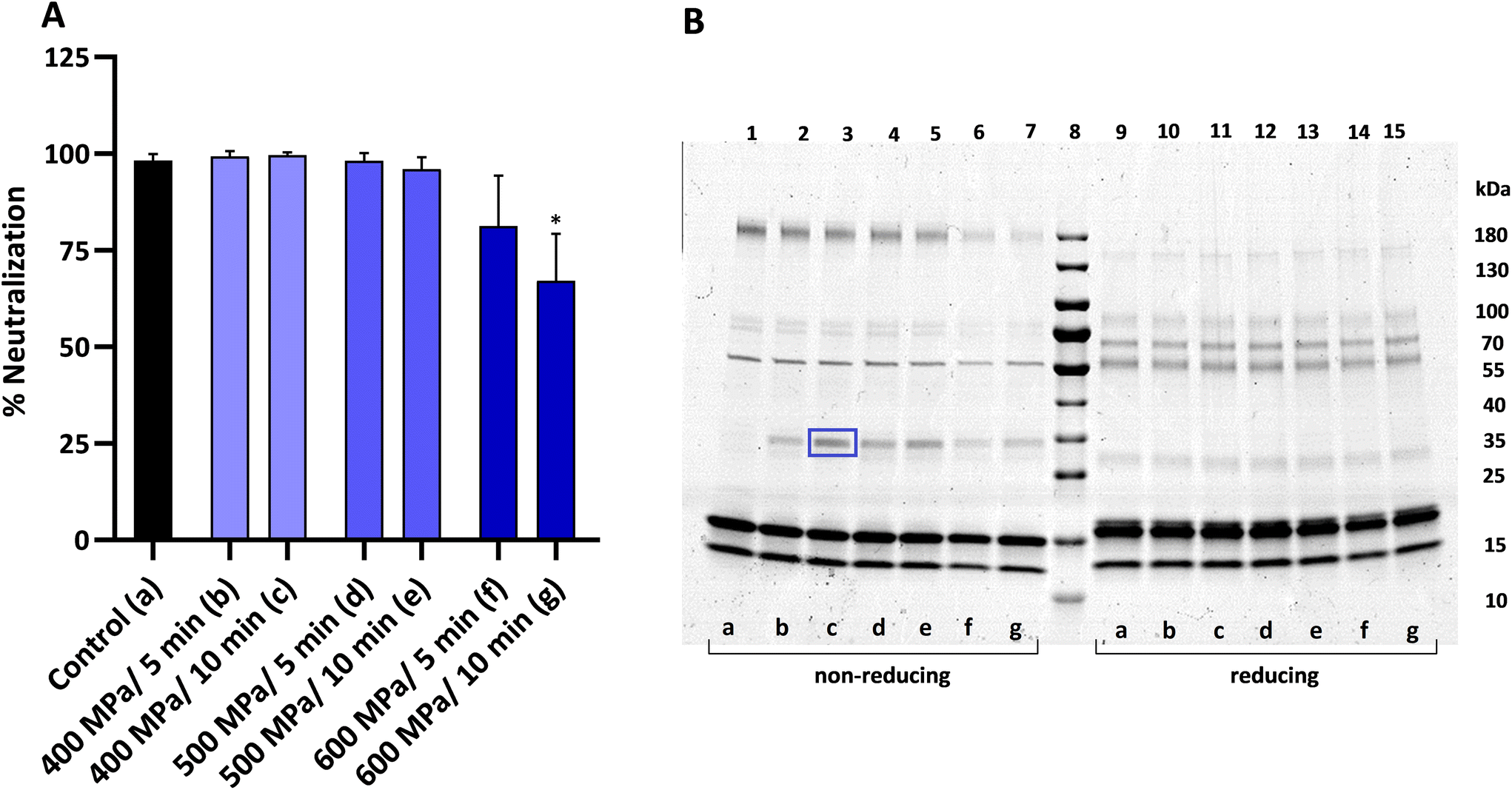 | ||
| Fig. 7 (A) Effect of HHP treatments on the bovine RV WC3 neutralization activity on Caco-2/TC7 cells of whey enriched in IgG (0.5 mg mL−1). The results are expressed as a percentage of the neutralization activity as mean ± standard deviation of duplicates from at least three independent experiments (n ≥ 6). Statistical analysis was performed using the Kruskal–Wallis test followed by Dunn's multiple comparisons test. *Indicates statistically significant differences (*p < 0.05) of the HHP treated samples in relation to the non-treated control (black bar). (B) SDS-PAGE on 4–20% polyacrylamide gel performed under non-reducing (lanes 1–7) and reducing (lanes 9–15) conditions. The code letter corresponds to the letter in brackets with regard to the HHP treatment. The framed band in lane 3 was subsequently identified by proteomic analysis. | ||
With regard to the IgG-enriched buttermilk preparation, an increase in the neutralization activity was observed when treatments of lower and intermediate intensity were applied. Specifically, treatment at 400 MPa for 5 min caused a significant increase of 13% of antirotaviral activity. Treatments at 400 MPa for 10 min and 500 MPa for 5 min also increased the activity at 8% in both cases, though not at a significant level. Treatment at 500 MPa for 10 min did not impact the neutralization activity. In contrast, application of pressure at 600 MPa to the IgG-enriched buttermilk caused a loss of activity at 7 and 18% when treated for 5 and 10 min, respectively, the latter being significant. Contrary to what was observed in the case of enriched whey preparations, SDS-PAGE analysis of the enriched buttermilk fractions did not show relevant changes in the protein profile of HHP-treated preparations with respect to the non-treated preparations (Fig. 8B). However, it should be mentioned that the bands corresponding to caseins (∼35 kDa) appeared more intense in the case of the HHP treated samples in comparison with the control. On the other hand, bands corresponding to α-lactalbumin and β-lactoglobulin (14 and 18 kDa, respectively) appeared slightly less intense when treated at the highest intensity (600 MPa for 10 min, lane 7), and it was evident only when SDS-PAGE was performed under non-reducing conditions. SDS-PAGE performed under reducing conditions did not reflect these differences in the electrophoretic pattern.
 | ||
| Fig. 8 (A) Effect of HHP treatments on the WC3 bovine RV neutralization activity on Caco-2/TC7 cells of buttermilk enriched in IgG (0.5 mg mL−1). The results are expressed as a percentage of RV neutralization, as mean ± standard deviation of duplicates from at least three independent experiments (n ≥ 6). Statistical analysis was performed using one-way ANOVA followed by Dunnett's multiple comparisons test. *Indicates statistically significant differences of the HHP treated samples in relation to the non-treated control (*p < 0.05, ***p < 0.001). (B) SDS-PAGE on 4–20% polyacrylamide gel performed under non-reducing (lanes 1–7) and reducing (lanes 9–15) conditions. The code letter corresponds to the letter in brackets with regard to the HHP treatment. | ||
4. Discussion
Scientific evidence indicates that HBC is a promising nutraceutical that could be used for the prevention or treatment of several diseases in newborns and adults, in particular to prevent gastrointestinal infections and alleviate their symptoms.45 Considering this, in the present study, a non-purified fraction of IgG (IPF) and a purified IgG fraction were obtained from HBC against bovine RV and added to bovine whey and buttermilk, to elaborate dairy preparations with enhanced antirotaviral activity.Initially, the active concentrations against RV of whey, buttermilk and IgG were determined separately. It was found that IgG from HBC exerted a direct neutralizing effect against the bovine RV strain WC3 when tested in the Caco-2/TC7 human enterocyte model. High inhibitory activity (82.5%) was observed at 10 μg mL−1 and complete inhibition at 100 μg mL−1, concentrations comparable to those previously shown as active in the MA104 cell line.46,47 On the other hand, a protective effect of IgG against RV infection by blocking Caco-2/TC7 cells was not clearly observed, although partial inhibition was shown at high concentrations, suggesting that the antirotaviral mechanism of action of IgG, at least during the early stages of infection, is more related to the interaction between IgG and RV capsid proteins (virucidal effect) and less to the interaction of these proteins with the cell receptors for RV (protective effect). In this view, previous studies were conducted in MA104 cells by our research group using reassortant UK x RRV having different combinations of VP4 and VP7 to determine which of the viral surface proteins was associated with their inhibitory effect.47 It was shown that the main mechanism of RV neutralization by milk components was exerted at the level of binding to the cell surface and, more specifically, that IgG interacts mainly with VP7. In the study conducted by den Hartog et al. (2004), a similar neutralization in vitro assay was utilized and it was found that IgG from commercial bovine colostrum was shown to be effective in neutralizing the human respiratory syncytial virus (RSV) on HEp2 cells through direct binding of the protein to renilla-GFP RSV.48 Overall, the authors supported that IgG may contribute to immune protection as it could bind to human RSV and other human respiratory pathogens and induce effector functions through binding to human FcγRII on phagocytes.
Additionally, the antirotaviral potential of whey and buttermilk has been demonstrated. Whey exerted complete neutralization of RV infection at a concentration of 14.3 mg powder per mL and buttermilk reached similar levels of activity at about a two-fold concentration (27.3 mg powder per mL). The antirotaviral activity of bovine whey has been mainly attributed to its protein fraction.49 Proteins with known antirotaviral activity in bovine whey are lactoferrin,50 IgG51 and proteose peptone component 3.52 These bioactive proteins are also found in buttermilk at some level; however, the antirotaviral potential of this fraction has been predominantly attributed to the proteins of the MFGM complex.53 More specifically, the MFGM proteins mucin 1,54 lactadherin,55 and xanthine oxidase52 have been reported as important antirotaviral agents. In the present study, all the above proteins were identified by SDS-PAGE in the buttermilk preparations (Fig. 2). Finally, apart from the protein fraction, it should be noted that there are more active compounds present in whey and buttermilk that could have contributed to the protective effect against RV, such as specific oligosaccharides56 and lipid components associated with the MFGM.57
It is important to stress that the results obtained on the antirotaviral activity of the above cited molecules or fractions are sometimes conflicting among different studies. These controversial results can be due to differences in the experimental conditions, such as the viral strain, the incubation times of the virus/sample, and the purification protocols of the proteins. Additionally, an important parameter is the cell line in which the antirotaviral activity is evaluated. For example, the response of MA104 cells after RV infection is very well characterized since this cell line has been traditionally used in this type of study due to its particularly high RV susceptibility.58,59 However, MA104 cells are considered a less physiologically relevant model since they do not express the morphological and functional characteristics of the intestinal cells. On the other hand, Caco-2/TC7 cells, although being colonic epithelial cells, when differentiated, resemble the mature enterocytes of the human small intestinal epithelium.60 Accordingly, comparative studies have shown that the antirotaviral effect of milk proteins or fractions is exerted differently on these two cell lines.54,61 Furthermore, RV shows different infectivity depending on the cell line, with the murine RV strain EHP described as neuraminidase (NA)-resistant when tested in the MA104 cell line,62 while it behaved as NA-sensitive when tested in Caco-2 cells.63 The above observations indicate the importance of considering the methodological details when choosing a suitable experimental design. Since RV particles principally infect the upper villi of the mature enterocytes in the small intestine and cause structural changes in the intestinal epithelium,64,65 the use of Caco-2/TC7 as an in vitro model could represent one step closer to physiological conditions. In any case, it should be noted that cultured Caco-2 cells are more difficult to work with as they must be treated with extreme caution to avoid detachment of the monolayer during the washing and immunostaining steps to detect RV.
After determining the range of active antiviral concentrations for whey and buttermilk in our study, the fractions were enriched in IPF and IgG from HBC and spray dried. The choice of spray drying was made as this is the most used procedure of drying at industrial level. Next, a sandwich ELISA was used for the quantification of the IgG present in the PBS-reconstituted powders. Interestingly, the IgG levels determined by the immunoassay were very low compared to the theoretical amount of total IgG added in the enrichment (approximately seven times lower). One possible explanation could be the interaction that the proteins of whey, buttermilk and HBC may have established with the added IgG during spray drying, which may have interfered with the binding of the antibody in the ELISA.66 In a previous study, low detection of IgG was reported by sandwich ELISA compared to radial immunodiffusion in samples such as liquid milk, whey and skimmed milk powders, however, the authors did not identify a specific component responsible for IgG underestimation.67 An explanation may be that the drying process may have caused a decrease in the IgG immunoreactivity which ultimately resulted in a weaker signal in the measurement. Nevertheless, as was observed in the antirotaviral activity assays, the functionality of our samples was retained after spray drying; therefore, in this case, a loss of immunoreactivity did not result in a loss of bioactivity, at least regarding the antiviral activity.
With regard to the antirotaviral activity of the enriched fractions, it was observed that, overall, the whey preparations, enriched either with IgG or IPF, were more active than the corresponding buttermilk preparations, as higher neutralization values were observed for lower whey concentrations. This is in accordance with previous observations showing that the antirotaviral activity exerted by bovine milk fractions was mainly associated with the whey fraction of milk and colostrum, rather than with the cream derived fractions.47 In this regard, the electrophoretic profile of the buttermilk samples (Fig. 2B) revealed the predominance of bands corresponding to caseins; although these are associated with some antirotaviral activity,68 it is significantly lower than that of IgG and lactoferrin. Furthermore, compared to the whey preparations, the values for the neutralization activity of the buttermilk preparations had greater deviations. In this sense, it must be taken into account that there is a high percentage of bioactive proteins present in buttermilk that are associated with the MFGM and are distributed asymmetrically, with peripheral and integral proteins and with weak protein interactions with the lipidic membrane itself.69 Since the structural organization of the MFGM affects greatly its biological activity,70 this different exposure of the proteins on the membrane could have resulted in differences in the neutralization activity of the buttermilk preparations between different assays.
The enrichment of both whey and buttermilk with the different Ig fractions from HBC resulted in an increase in the antirotaviral activity compared to the non-enriched fractions. Similarly, a recent study investigated the antibacterial effect of acid whey enriched in IgG and lactoferrin and it was found that the enriched powdered formula had lower IC50 values than a pure lactoferrin powder, indicating that there are synergistic effects between the ingredients of the enriched whey that could amplify its biofunctional outcome.71 In the present study, enrichment with purified IgG resulted in the highest RV neutralization, both for whey and buttermilk preparations, with inhibition values above 99% at concentrations of 3 and 6 mg of powder per mL, respectively. However, significant neutralization values were also observed when the enrichment of the two fractions was performed with IPF. SDS-PAGE analysis of this fraction showed that IgG was the dominant protein, although other Igs such as IgA and IgM may also be present (Fig. 1B and C). Specifically, ELISA analysis showed that 38.1% of the total protein content of IPF corresponded to IgG. The potent antirotaviral activity shown by IPF is a particularly important finding as it underscores the possibility of exploitation of bioactive HBC components without the need for complete specific antibody purification, which would require higher expenses with regard to costs, resources and energy. Accordingly, the study of Civra et al. (2019) demonstrated that the conventional bovine RV vaccine was sufficient to enhance the antihuman RV protective efficiency of bovine colostrum due to a high titer of cross-reactive anti-human RV IgG.24 Taken together, these findings indicate a conservative and feasible approach to produce HBC derived formulas that could be exploited in the fight against RV infections in both developed and developing countries. Such preparations could be a way of confronting the RV infections as a complementary strategy, or as a way of prophylaxis by their addition to infant formulas. In any case, the need for a safe product is required and for this, the effect of industrial preservation treatments on the bioactivity of the milk fractions needs to be assessed.
Four different thermal treatments of increasing intensity were applied to whey and buttermilk and their corresponding IgG-enriched preparations. It was observed that the inhibitory activity of all samples was mildly affected by pasteurization, but significantly decreased after the application of more intense treatments. The exception was the enriched buttermilk preparation, which showed an ∼6% activity increase after the application of short-time treatments (75 or 85 °C for 20 s). In this regard, the effect of heat treatments on the antirotaviral activity of whey and buttermilk had been previously studied in MA104 cells.46,52 In that study, a significant loss of neutralization activity was also observed when whey and buttermilk were tested after treatment at 85 °C for either 20 s or 10 min, and the activity loss was equivalent to the values observed in the present study. Contrary to our results, the above-cited study showed that pasteurization had no effect on the activity of whey but slightly affected that of buttermilk (23.3% decrease). Finally, the response of preparations to mild heat treatments (75 °C for 10 min) found in the present study, revealed that the loss of activity is greater when evaluated in Caco-2 cells in comparison with MA104 cells. In another study, the thermal treatment at 85 °C for 10 min of a commercial macromolecular whey protein (MMWP) at 0.5 mg mL−1 resulted in a 54% increase of RV infectivity in Caco-2 cells.51
Thermal treatments have been shown to cause a significant reduction in the levels of rotaviral antibodies in bovine milk and different products based on bovine milk and, in some cases, they become undetectable.72,73 In this regard, we have previously observed by western blot analysis that bovine lactoferrin and IgG were also susceptible to heat denaturation at 85 °C for 10 min.46 On the other hand, bovine mucin 1 and lactadherin were shown to have heat resistance,52 and thermal treatments at 80 °C for up to 120 min did not affect their secondary structure, as shown in other studies.74 This could explain the lower effect on the neutralization activity that was observed in the case of buttermilk and enriched buttermilk preparations, in comparison with those based on whey. Furthermore, concerning the increase in the neutralization activity that was observed for the enriched buttermilk, it can be hypothesized that IgG could have been protected during thermal treatment by the MFGM. In this regard, the possible application of whole buttermilk as a novel encapsulating material for the protection of bioactive compounds was previously explored,75 finding that high heat treatment in combination with an increased pH improved the performance of buttermilk as an encapsulant for the manufacture of recombined omega-3 spray-dried powders.
Finally, in the present study the effect of HHP on the neutralization activity of IgG-enriched whey and buttermilk preparations was investigated. Upon HHP treatments, the covalent bonds of the proteins do not break, which allows the maintenance of their primary structures.76 Therefore, some milk proteins, such as α-lactalbumin and bovine serum albumin, present stability under pressure treatment mainly due to the high number of disulfide bonds, unlike β-lactoglobulin which is considered pressure sensitive and tends to associate with casein micelles.77–79 Lactoferrin and IgG have also been found to be relatively more resistant to pressure than to heat treatment.77,80 However, several factors such as the pressure level and holding time, temperature and cycling conditions affect the proteins differently.34 For pressures under 400 MPa, IgG is fairly stable to HHP treatment for holding times under 10 min. However, for pressures above 500 MPa the immunoreactivity of IgG starts to decrease to eventually reaching a dramatic loss of immunoreactivity at 600 MPa.34,81 Accordingly, in the present study, treatments at 600 MPa for 10 min caused the maximum loss of antirotaviral activity, of 32% and 18% for the IgG-enriched whey and buttermilk, respectively (Fig. 7A and 8A). On the other hand, low-intensity HHP treatment (400 MPa for 5 min) of the enriched buttermilk resulted in a 13% increase of its neutralization activity compared to the untreated control (Fig. 8A). In this regard, it has been shown that the presence of casein micelles in the medium has a protective effect against pressure denaturation of lactoferrin and Igs. It is possible that lactoferrin can electrostatically bind to caseins, resulting in increased lactoferrin stability under pressure.34 Similarly, casein micelles had a protective effect against pressure-induced denaturation of Igs (450–700 MPa for 0–30 min),82 but the exact mechanism is not known. Another explanation for the increase of the antirotaviral activity, which could also apply to the results obtained under heat treatments, might be the unraveling of bioactive MFGM proteins, that otherwise are partially or totally embedded in the phospholipid membranes of milk fat globules. Hence, a low intensity treatment could induce exposure of regions with bioactive properties, higher access to proteolytic enzymes in these regions and an overall improvement of the bioavailability of bioactive compounds against RV.
Several studies have indicated that HHP treatment at 600 MPa for 3 min83 or 5 min84 can be considered as a safe alternative to the pasteurization process for raw milk. Overall, as reviewed by Silva (2023), HHP treatments at 400–600 MPa are equivalent to thermal pasteurization treatments (5–8 logs of bacterial reduction).85 However, the variability of the existing data is high and there is a strong dependence on the bacterial strain tested in each study.85 With regard to the effect of the tested treatments on the antirotaviral activity of the enriched dairy preparations, the results of this study showed that HHP conditions corresponding to those considered alternative to thermal pasteurization have not caused significant reduction in their RV neutralizing potential, in agreement with previous observations.86 More specifically, we found that the neutralization values resulting after thermal pasteurization (75 °C for 20 s) and HHP treatment (500 MPa for 10 min or 600 MPa for 5 min) were above 90% except for WH + 0.5 IgG (82.8%). Therefore, HHP treatment could be considered safe with regard to the preservation of high antirotaviral activity.
Currently, a number of food products processed by HHP exist in the market; however, its application on dairy products is still limited, mainly because the advantages of this technology in comparison with conventional thermal treatments have not yet been completely demonstrated. There are several comparative studies between HHP and thermal technologies, regarding microbial safety87 and the effect on protein denaturation and immunogenicity.78 Furthermore, some research is being conducted to investigate the possible use of indigenous milk enzymes (alkaline phosphatase, γ-glutamyltransferase and phosphohexose isomerase) as process indicators for HPP.88,89 Hence, there is a need for an official agreement for the establishment of specific indicators to ensure the safety of HHP treated dairy products.
5. Conclusions
The present study showed that whey and buttermilk together with fractions sourced from HBC can be used for developing new valuable functional products to combat viral infections. Accordingly, they could play an important role in countries where RV has a high prevalence. Valorization of dairy byproducts by incorporating them into novel products with relevant nutritional and biological properties should be one of the target points in developing future foods. Non-thermal treatments such as HHP could be an alternative key method to preserve their bioactivity; however, further in vivo trials should be considered in order to ensure the efficacy of these products in patients challenged with RV.Author contributions
D. G.: conceptualization, methodology, investigation, formal analysis, software, visualization, and writing – original draft; L. G.: investigation, software, visualization, and writing – original draft; I. A.: methodology and investigation; M. L.: methodology and writing – review and editing; E. P.: methodology and writing – review and editing; M. D. P.: methodology; L. S.: conceptualization, methodology, validation, supervision, writing – review and editing, funding acquisition, and project administration. All authors have accepted the final manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This project was funded by the European Union's H2020 research and innovation program under Marie Sklodowska-Curie grant agreement no. 801586, the AGL2017-82987R and PID2022-139104OB-I00 projects from the Ministerio de Ciencia e Innovacion (Spanish Government) and the European Social Fund, and the Aragon Regional Government (A20_23R).References
- A. F. Pires, N. G. Marnotes, O. D. Rubio, A. C. Garcia and C. D. Pereira, Dairy By-Products: A Review on the Valorization of Whey and Second Cheese Whey, Foods, 2021, 10, 1067 CrossRef CAS PubMed.
- V. Lavelli and M. P. Beccalli, Cheese whey recycling in the perspective of the circular economy: Modeling processes and the supply chain to design the involvement of the small and medium enterprises, Trends Food Sci. Technol., 2022, 126, 86–98 CrossRef CAS.
- A. Panghal, R. Patidar, S. Jaglan, N. Chhikara, S. K. Khatkar, Y. Gat and N. Sindhu, Whey valorization: current options and future scenario – A critical review, Nutr. Food Sci., 2018, 48, 520–535 CrossRef.
- S. R. Patel and Z. V. P. Murthy, Waste valorization: Recovery of lactose from partially deproteinated whey by using acetone as anti-solvent, Dairy Sci. Technol., 2011, 91, 53–63 CAS.
- A. A. Chatzipaschali and A. G. Stamatis, Biotechnological Utilization with a Focus on Anaerobic Treatment of Cheese Whey: Current Status and Prospects, Energies, 2012, 5, 3492–3525 CrossRef CAS.
- G. W. Smithers, Whey and whey proteins—From ‘gutter-to-gold’, Int. Dairy J., 2008, 18, 695–704 CrossRef CAS.
- C. Mollea, L. Marmo and F. Bosco, Valorisation of Cheese Whey, a By-Product from the Dairy Industry, Food Ind., In TechOpen, 2013 DOI:10.5772/53159.
- M. Corredig and D. G. Dalgleish, Isolates from industrial buttermilk: Emulsifying properties of materials derived from the milk fat globule membrane, J. Agric. Food Chem., 1997, 45, 4595–4600 CrossRef CAS.
- R. Roesch, A. Rincon and M. Corredig, Emulsifying properties of fractions prepared from commercial buttermilk by microfiltration, J. Dairy Sci., 2004, 87, 4080–4087 CrossRef CAS PubMed.
- I. Sodini, P. Morin, A. Olabi and R. Jiménez-Flores, Compositional and functional properties of buttermilk: A comparison between sweet, sour, and whey buttermilk, J. Dairy Sci., 2006, 89, 525–536 CrossRef CAS PubMed.
- J. O'Connell and P. F. Fox, Heat stability of buttermilk, J. Dairy Sci., 2000, 83, 1728–1732 CrossRef PubMed.
- A. H. Ali, Current knowledge of buttermilk: Composition, applications in the food industry, nutritional and beneficial health characteristics, Int. J. Dairy Technol., 2019, 72, 169–182 CrossRef CAS.
- L. Claumarchirant, A. Cilla, E. Matencio, L. M. Sanchez-Siles, P. Castro-Gomez, J. Fontecha, A. Alegría and M. J. Lagarda, Addition of milk fat globule membrane as an ingredient of infant formulas for resembling the polar lipids of human milk, Int. Dairy J., 2016, 61, 228–238 CrossRef CAS.
- B. Lönnerdal, Bioactive proteins in human milk: health, nutrition, and implications for infant formulas, J. Pediatr., 2016, 173, 4–9 CrossRef PubMed.
- J. Fontecha, L. Brink, S. Wu, Y. Pouliot, F. Visioli and R. Jiménez-Flores, Sources, production, and clinical treatments of milk fat globule membrane for infant nutrition and well-being, Nutrients, 2020, 12, 1607 CrossRef CAS PubMed.
- B. A. McGrath, P. F. Fox, P. L. McSweeney and A. L. Kelly, Composition and properties of bovine colostrum: a review, Dairy Sci. Technol., 2016, 96, 133–158 CrossRef CAS.
- R. J. Playford and M. J. Weiser, Bovine colostrum: Its constituents and uses, Nutrients, 2021, 13, 265 CrossRef CAS PubMed.
- P. Marnila and H. Korhonen, in Encyclopedia of Dairy Sciences, ed. J. W. Fuquay, Academic Press, San Diego, 2nd edn, 2011, pp. 591–597. DOI:10.1016/B978-0-12-374407-4.00322-8.
- R. D. Gomes, K. Anaya, A. B. Galdino, J. P. Oliveira, M. A. Gama, C. A. Medeiros, E. C. Gavioli, A. L. F. Porto and A. H. Rangel, Bovine colostrum: A source of bioactive compounds for prevention and treatment of gastrointestinal disorders, NFS J., 2021, 25, 1–11 CrossRef CAS.
- G. Barrington, T. Besser, W. Davis, C. Gay, J. Reeves and T. B. McFadden, Expression of immunoglobulin G1 receptors by bovine mammary epithelial cells and mammary leukocytes, J. Dairy Sci., 1997, 80, 86–93 CrossRef CAS PubMed.
- M. Inagaki, M. Yamamoto, K. Uchida, H. Yamaguchi, M. Kawasaki, K. Yamashita, T. Yabe and Y. Kanamaru, In vitro and in vivo evaluation of the efficacy of bovine colostrum against human rotavirus infection, Biosci., Biotechnol., Biochem., 2010, 74, 680–682 CrossRef CAS PubMed.
- M. Inagaki, M. Yamamoto, T. Yabe, K. Uchida, M. Kawasaki, T. Nakagomi, O. Nakagomi, N. Minamoto and Y. Kanamaru, Multiple-dose therapy with bovine colostrum confers significant protection against diarrhea in a mouse model of human rotavirus-induced gastrointestinal disease, J. Dairy Sci., 2013, 96, 806–814 CrossRef CAS PubMed.
- S. H. Barakat, M. A. Meheissen, O. M. Omar and D. A. Elbana, Bovine Colostrum in the Treatment of Acute Diarrhea in Children: A Double-Blinded Randomized Controlled Trial, J. Trop. Pediatr., 2019, 66, 46–55 Search PubMed.
- A. Civra, A. Altomare, R. Francese, M. Donalisio, G. Aldini and D. Lembo, Colostrum from cows immunized with a veterinary vaccine against bovine rotavirus displays enhanced in vitro anti-human rotavirus activity, J. Dairy Sci., 2019, 102, 4857–4869 CrossRef CAS PubMed.
- C. Troeger, I. A. Khalil, P. C. Rao, S. Cao, B. F. Blacker, T. Ahmed, G. Armah, J. E. Bines, T. G. Brewer, D. V. Colombara, G. Kang, B. D. Kirkpatrick, C. D. Kirkwood, J. M. Mwenda, U. D. Parashar, W. A. Petri Jr., M. S. Riddle, A. D. Steele, R. L. Thomson, J. L. Walson, J. W. Sanders, A. H. Mokdad, C. J. L. Murray, S. I. Hay and R. C. Reiner Jr., Rotavirus vaccination and the global burden of rotavirus diarrhea among children younger than 5 years, JAMA Pediatr., 2018, 172, 958–965 CrossRef PubMed.
- T. Oppong, H. Yang, C. Amponsem-Boateng, E. D. Kyere, T. Abdulai, G. Duan and G. Opolot, Enteric pathogens associated with gastroenteritis among children under 5 years in sub-Saharan Africa: A systematic review and meta-analysis, Epidemiol. Infect., 2020, 148, 1–26 CrossRef PubMed.
- M. Ciarlet and F. Schödel, Development of a rotavirus vaccine: clinical safety, immunogenicity, and efficacy of the pentavalent rotavirus vaccine, RotaTeq®, Vaccine, 2009, 27, G72–G81 CrossRef CAS PubMed.
- C. A. Omatola and A. O. Olaniran, Rotaviruses: from pathogenesis to disease control—a critical review, Viruses, 2022, 14, 875 CrossRef CAS PubMed.
- S. Caddy, G. Papa, A. Borodavka and U. Desselberger, Rotavirus research: 2014–2020, Virus Res., 2021, 304, 198499 CrossRef CAS PubMed.
- M. F. Carvalho and D. Gill, Rotavirus vaccine efficacy: current status and areas for improvement, Hum. Vaccines Immunother., 2019, 15, 1237–1250 CrossRef PubMed.
- A. Mediwaththe, D. Bogahawaththa, M. K. Grewal, J. Chandrapala and T. Vasiljevic, Structural changes of native milk proteins subjected to controlled shearing and heating, Food Res. Int., 2018, 114, 151–158 CrossRef CAS PubMed.
- V. Raikos, Effect of heat treatment on milk protein functionality at emulsion interfaces. A review, Food Hydrocolloids, 2010, 24, 259–265 CrossRef CAS.
- S. G. Anema, in Milk Proteins, ed. M. Boland and H. Singh, Academic Press, 3rd edn, 2020, pp. 325–384. DOI:10.1016/B978-0-12-815251-5.00009-8.
- N. Gharbi, A. Marciniak and A. Doyen, Factors affecting the modification of bovine milk proteins in high hydrostatic pressure processing: An updated review, Compr. Rev. Food Sci. Food Saf., 2022, 21, 4274–4293 CrossRef CAS PubMed.
- A. Grumezescu and A. M. Holban, Milk-Based Beverages: Volume 9: The Science of Beverages, Woodhead Publishing, 2019 Search PubMed.
- G. Mainer, L. Sanchez, J. Ena and M. Calvo, Kinetic and thermodynamic parameters for heat denaturation of bovine milk IgG, IgA and IgM, J. Food Sci., 1997, 62, 1034–1038 CrossRef CAS.
- G. Abernethy, D. Otter, K. Arnold, J. Austad, S. Christiansen, I. Ferreira, F. Irvine, C. Marsh, L. R. Massom, D. Otter, K. Pearce, J. Stevens, J. Szpylka, P. Vyas, D. Woollard and C. Wu, Determination of immunoglobulin G in bovine colostrum and milk powders, and in dietary supplements of bovine origin by protein G affinity liquid chromatography: collaborative study, J. AOAC Int., 2010, 93, 622–627 CrossRef CAS PubMed.
- U. K. Laemmli, Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature, 1970, 227, 680–685 CrossRef CAS PubMed.
- A. Ardia, D. Knorr and V. Heinz, Adiabatic heat modelling for pressure build-up during high-pressure treatment in liquid-food processing, Food Bioprod. Process., 2004, 82, 89–95 CrossRef.
- K. Knoerzer, R. Buckow, P. Sanguansri and C. Versteeg, Adiabatic compression heating coefficients for high-pressure processing of water, propylene-glycol and mixtures – A combined experimental and numerical approach, J. Food Eng., 2010, 96, 229–238 CrossRef CAS.
- D. Graikini, A. B. Soro, S. P. Sivagnanam, B. K. Tiwari and L. Sánchez, Bioactivity of Fucoidan-Rich Extracts from Fucus vesiculosus against Rotavirus and Foodborne Pathogens, Mar. Drugs, 2023, 21, 478 CrossRef CAS PubMed.
- M. Arnold, J. T. Patton and S. M. McDonald, Culturing, storage, and quantification of rotaviruses, Curr. Protoc. Microbiol., 2009, 15, 15C.3.1–15C.3.24 Search PubMed.
- S. Ferruzza, C. Rossi, M. L. Scarino and Y. Sambuy, A protocol for differentiation of human intestinal Caco-2 cells in asymmetric serum-containing medium, Toxicol. In Vitro, 2012, 26, 1252–1255 CrossRef CAS PubMed.
- J. Mesonero, L. Mahraoui, M. Matosin, A. Rodolosse, M. Rousset and E. Brot-Laroche, Expression of the hexose transporters GLUTI-GLUT5 and SGLTI in clones of Caco-2 cells, Biochem. Soc. Trans., 1994, 22, 681–684 CrossRef CAS PubMed.
- L. H. Ulfman, J. H. Leusen, H. F. Savelkoul, J. O. Warner and R. J. Van Neerven, Effects of bovine immunoglobulins on immune function, allergy, and infection, Front. Nutr., 2018, 5, 52 CrossRef PubMed.
- J. A. Parrón, D. Ripollés, M. D. Pérez, M. Calvo, J. T. Rasmussen and L. Sánchez, Effect of heat treatment on antirotaviral activity of bovine and ovine whey, Int. Dairy J., 2016, 60, 78–85 CrossRef.
- J. A. Parrón, D. Ripollés, A. C. Sánchez, M. D. Pérez, M. Calvo, S. López, C. F. Arias and L. Sánchez, Antirotaviral activity of bovine milk components: Extending the list of inhibitory proteins and seeking a better understanding of their neutralization mechanism, J. Funct. Foods, 2018, 44, 103–111 CrossRef.
- G. den Hartog, S. Jacobino, L. Bont, L. Cox, L. H. Ulfman, J. H. W. Leusen and R. J. J. van Neerven, Specificity and Effector Functions of Human RSV-Specific IgG from Bovine Milk, PLoS One, 2014, 9, e112047 CrossRef PubMed.
- T. B. Ng, R. C. F. Cheung, J. H. Wong, Y. Wang, D. T. M. Ip, D. C. C. Wan and J. Xia, Antiviral activities of whey proteins, Appl. Microbiol. Biotechnol., 2015, 99, 6997–7008 CrossRef CAS PubMed.
- D. Graikini, C. Conesa, I. Abad, M. D. Pérez and L. Sánchez, Evaluation of in vitro antirotaviral activity of lactoferrin from different species using a human intestinal model, Int. Dairy J., 2023, 149, 105818 CrossRef.
- A. Bojsen, J. Buesa, R. Montava, A. S. Kvistgaard, M. B. Kongsbak, T. E. Petersen, C. W. Heegaard and J. T. Rasmussen, Inhibitory Activities of Bovine Macromolecular Whey Proteins on Rotavirus Infections In Vitro and In Vivo, J. Dairy Sci., 2007, 90, 66–74 CrossRef CAS PubMed.
- J. A. Parrón, D. Ripollés, M. D. Pérez, M. Calvo, J. T. Rasmussen and L. Sánchez, Antirotaviral activity of bovine and ovine dairy byproducts, J. Agric. Food Chem., 2017, 65, 4280–4288 CrossRef PubMed.
- M. H. Abd El-Salam and S. El-Shibiny, Milk fat globule membrane: An overview with particular emphasis on its nutritional and health benefits, Int. J. Dairy Technol., 2020, 73, 639–655 CrossRef CAS.
- A. S. Kvistgaard, L. T. Pallesen, C. F. Arias, S. López, T. E. Petersen, C. W. Heegaard and J. T. Rasmussen, Inhibitory Effects of Human and Bovine Milk Constituents on Rotavirus Infections, J. Dairy Sci., 2004, 87, 4088–4096 CrossRef CAS PubMed.
- M. Inagaki, S. Nagai, T. Yabe, S. Nagaoka, N. Minamoto, T. Takahashi, T. Matsuda, O. Nakagomi, T. Nakagomi and T. Ebina, The bovine lactophorin C-terminal fragment and PAS6/7 were both potent in the inhibition of human rotavirus replication in cultured epithelial cells and the prevention of experimental gastroenteritis, Biosci., Biotechnol., Biochem., 2010, 74, 1386–1390 CrossRef CAS PubMed.
- D. R. Laucirica, V. Triantis, R. Schoemaker, M. K. Estes and S. Ramani, Milk Oligosaccharides Inhibit Human Rotavirus Infectivity in MA104 Cells, J. Nutr., 2017, 147, 1709–1714 CrossRef CAS PubMed.
- K. L. Fuller, T. B. Kuhlenschmidt, M. S. Kuhlenschmidt, R. Jiménez-Flores and S. M. Donovan, Milk fat globule membrane isolated from buttermilk or whey cream and their lipid components inhibit infectivity of rotavirus in vitro, J. Dairy Sci., 2013, 96, 3488–3497 CrossRef CAS PubMed.
- C. A. Guerrero, S. Zárate, G. Corkidi, S. López and C. F. Arias, Biochemical characterization of rotavirus receptors in MA104 cells, J. Virol., 2000, 74, 9362–9371 CrossRef CAS PubMed.
- S. L. Londrigan, M. J. Hewish, M. J. Thomson, G. M. Sanders, H. Mustafa and B. S. Coulson, Growth of rotaviruses in continuous human and monkey cell lines that vary in their expression of integrins, J. Gen. Virol., 2000, 81, 2203–2213 CrossRef CAS PubMed.
- Y. Sambuy, I. De Angelis, G. Ranaldi, M. Scarino, A. Stammati and F. Zucco, The Caco-2 cell line as a model of the intestinal barrier: influence of cell and culture-related factors on Caco-2 cell functional characteristics, Cell Biol. Toxicol., 2005, 21, 1–26 CrossRef CAS PubMed.
- M. H. Monaco, G. Gross and S. M. Donovan, Whey protein lipid concentrate high in milk fat globule membrane components inhibit porcine and human rotavirus in vitro, Front. Pediatr., 2021, 9, 731005 CrossRef PubMed.
- M. Ciarlet, J. E. Ludert, M. Iturriza-Gómara, F. Liprandi, J. J. Gray, U. Desselberger and M. K. Estes, Initial interaction of rotavirus strains with N-acetylneuraminic (sialic) acid residues on the cell surface correlates with VP4 genotype, not species of origin, J. Virol., 2002, 76, 4087–4095 CrossRef CAS PubMed.
- J. E. Ludert, N. Feng, J. H. Yu, R. L. Broome, Y. Hoshino and H. B. Greenberg, Genetic mapping indicates that VP4 is the rotavirus cell attachment protein in vitro and in vivo, J. Virol., 1996, 70, 487–493 CrossRef CAS PubMed.
- U. S. Geletu, M. A. Usmael and F. D. Bari, Rotavirus in calves and its zoonotic importance, Vet. Med. Int., 2021, 6639701 CAS.
- O. Lundgren and L. Svensson, Pathogenesis of rotavirus diarrhea, Microbes Infect., 2001, 3, 1145–1156 CrossRef CAS PubMed.
- J. G. Kenna, G. N. Major and R. S. Williams, Methods for reducing non-specific antibody binding in enzyme-linked immunosorbent assays, J. Immunol. Methods, 1985, 85, 409–419 CrossRef CAS PubMed.
- A. Kummer, D. Kitts, E. Li-Chan, J. Losso, B. Skura and S. Nakai, Quantification of bovine IgG in milk using enzyme-linked immunosorbent assay, Food Agric. Immunol., 1992, 4, 93–102 CrossRef CAS.
- M. Inagaki, H. Muranishi, K. Yamada, K. Kakehi, K. Uchida, T. Suzuki, T. Yabe, T. Nakagomi, O. Nakagomi and Y. Kanamaru, Bovine κ-casein inhibits human rotavirus (HRV) infection via direct binding of glycans to HRV, J. Dairy Sci., 2014, 97, 2653–2661 CrossRef CAS PubMed.
- K. Dewettinck, R. Rombaut, N. Thienpont, T. T. Le, K. Messens and J. Van Camp, Nutritional and technological aspects of milk fat globule membrane material, Int. Dairy J., 2008, 18, 436–457 CrossRef CAS.
- R. Zanabria, M. W. Griffiths and M. Corredig, Does structure affect biological function? Modifications to the protein and phospholipids fraction of the milk fat globule membrane after extraction affect the antiproliferative activity of colon cancer cells, J. Food Biochem., 2020, 44, e13104 CrossRef PubMed.
- F. Ostertag and J. Hinrichs, Enrichment of Lactoferrin and Immunoglobulin G from Acid Whey by Cross-Flow Filtration, Foods, 2023, 12, 2163 CrossRef CAS PubMed.
- J. Lecce and M. W. King, Rotaviral antibodies in cow's milk, Can. J. Comp. Med., 1982, 46, 434 CAS.
- R. H. Yolken, G. A. Losonsky, S. Vonderfecht, F. Leister and S. B. Wee, Antibody to human rotavirus in cow's milk, N. Engl. J. Med., 1985, 312, 605–610 CrossRef CAS PubMed.
- J. B. Madsen, K. I. Pakkanen and S. Lee, Thermostability of bovine submaxillary mucin (BSM) in bulk solution and at a sliding interface, J. Colloid Interface Sci., 2014, 424, 113–119 CrossRef CAS PubMed.
- M. A. Augustin, S. Bhail, L. J. Cheng, Z. Shen, S. Øiseth and L. Sanguansri, Use of whole buttermilk for microencapsulation of omega-3 oils, J. Funct. Foods, 2015, 19, 859–867 CrossRef CAS.
- W. Wang, P. Yang, L. Rao, L. Zhao, X. Wu, Y. Wang and X. Liao, Effect of high hydrostatic pressure processing on the structure, functionality, and nutritional properties of food proteins: A review, Compr. Rev. Food Sci. Food Saf., 2022, 21, 4640–4682 CrossRef CAS PubMed.
- T. Huppertz, T. Vasiljevic, B. Zisu and H. Deeth, Whey Proteins, Elsevier, 2019, pp. 281–334 Search PubMed.
- D. Bogahawaththa, R. Buckow, J. Chandrapala and T. Vasiljevic, Comparison between thermal pasteurization and high pressure processing of bovine skim milk in relation to denaturation and immunogenicity of native milk proteins, Innovative Food Sci. Emerging Technol., 2018, 47, 301–308 CrossRef CAS.
- M. Munir, M. Nadeem, T. M. Qureshi, T. S. Leong, C. J. Gamlath, G. J. Martin and M. Ashokkumar, Effects of high pressure, microwave and ultrasound processing on proteins and enzyme activity in dairy systems—A review, Innovative Food Sci. Emerging Technol., 2019, 57, 102192 CrossRef CAS.
- A. Soni, L. M. Samuelsson, S. M. Loveday and T. B. Gupta, Applications of novel processing technologies to enhance the safety and bioactivity of milk, Compr. Rev. Food Sci. Food Saf., 2021, 20, 4652–4677 CrossRef CAS PubMed.
- H. E. Indyk, J. W. Williams and H. A. Patel, Analysis of denaturation of bovine IgG by heat and high pressure using an optical biosensor, Int. Dairy J., 2008, 18, 359–366 CrossRef CAS.
- C. Mazri, L. Sánchez, S. Ramos, M. Calvo and M. D. Pérez, Effect of high-pressure treatment on denaturation of bovine lactoferrin and lactoperoxidase, J. Dairy Sci., 2012, 95, 549–557 CrossRef CAS PubMed.
- G. Liu, C. Carøe, Z. Qin, D. M. Munk, M. Crafack, M. A. Petersen and L. Ahrné, Comparative study on quality of whole milk processed by high hydrostatic pressure or thermal pasteurization treatment, LWT – Food Sci. Technol., 2020, 127, 109370 CrossRef CAS.
- M. Buffa, B. Guamis, C. Royo and A. J. Trujillo, Microbiological changes throughout ripening of goat cheese made from raw, pasteurized and high-pressure-treated milk, Food Microbiol., 2001, 18, 45–51 CrossRef.
- F. V. M. Silva, Pasteurization of Food and Beverages by High Pressure Processing (HPP) at Room Temperature: Inactivation of Staphylococcus aureus, Escherichia coli, Listeria monocytogenes, Salmonella, and Other Microbial Pathogens, Appl. Sci., 2023, 13, 1193 CrossRef CAS.
- J. A. Parrón, D. Ripollés, S. J. Ramos, M. D. Pérez, M. Calvo and L. Sánchez, Effect of thermal and high-pressure treatments on the antirotaviral activity of human milk fractions, Innovative Food Sci. Emerging Technol., 2018, 47, 262–270 CrossRef.
- A. C. Stratakos, E. S. Inguglia, M. Linton, J. Tollerton, L. Murphy, N. Corcionivoschi, N. A. Koidis and B. K. Tiwari, Effect of high-pressure processing on the safety, shelf life and quality of raw milk, Innovative Food Sci. Emerging Technol., 2019, 52, 325–333 CrossRef CAS.
- B. Rademacher and J. Hinrichs, Effects of high-pressure treatment on indigenous enzymes in bovine milk: Reaction kinetics, inactivation and potential application, Int. Dairy J., 2006, 16, 655–661 CrossRef CAS.
- A. Koncz, L. Meszaros, J. Farkas, K. Pasztor-Huszar, R. Helt and N. Lechner, Pasteurisation of raw milk by high hydrostatic pressure, Acta Aliment., 2007, 36, 471–481 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3fo05250h |
| This journal is © The Royal Society of Chemistry 2024 |