
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
The quercetin metabolite 4-methylcatechol causes vasodilation via voltage-gated potassium (KV) channels†
Patrícia
Dias‡
ab,
Rudy
Salam‡
 cd,
Jana
Pourová
a,
Marie
Vopršalová
a,
Lukáš
Konečný
a,
Eduard
Jirkovský
a,
Jurjen
Duintjer Tebbens
c and
Přemysl
Mladěnka
*a
cd,
Jana
Pourová
a,
Marie
Vopršalová
a,
Lukáš
Konečný
a,
Eduard
Jirkovský
a,
Jurjen
Duintjer Tebbens
c and
Přemysl
Mladěnka
*a
aDepartment of Pharmacology and Toxicology, Faculty of Pharmacy, Charles University, Hradec Kralove, 500 05, Czech Republic. E-mail: mladenkap@faf.cuni.cz; Fax: +420 495 067 170; Tel: +420 495 067 295
bDivision of Outcomes & Translational Sciences, Pelotonia Research Center, The Ohio State University, 2255 Kenny Rd, Columbus, OH, USA
cDepartment of Biophysics and Physical Chemistry, Faculty of Pharmacy, Charles University, Hradec Kralove, 500 05, Czech Republic
dDepartment of Pharmacy, Faculty of Medicine, Brawijaya University, Malang, Indonesia
First published on 3rd October 2024
Abstract
Dietary polyphenols have been associated with many beneficial cardiovascular effects. However, these effects are rather attributed to small phenolic metabolites formed by the gut microbiota, which reach sufficient concentrations in systemic circulation. 4-Methylcatechol (4-MC) is one such metabolite. As it is shown to possess considerable vasorelaxant effects, this study aimed to unravel its mechanism of action. To this end, experimental in vitro and in silico approaches were employed. In the first step, isometric tension recordings were performed on rat aortic rings. 4-MC potentiated the effect of cyclic nucleotides, but the effect was not mediated by either soluble guanylyl cyclase (sGC), modification of cyclic adenosine monophosphate levels, or protein kinase G. Hence, downstream targets such as calcium or potassium channels were considered. Inhibition of voltage-gated K+ channels (KV) markedly decreased the effect of 4-MC, and vasodilation was partly decreased by inhibition of the KV7 isoform. Contrarily, other types of K+ channels or L-type Ca2+ channels were not involved. In silico reverse docking confirmed that 4-MC binds to KV7.4 through hydrogen bonding and hydrophobic interactions. In particular, it interacts with two crucial residues for KV7.4 activation: Trp242 and Phe246. In summary, our findings suggested that 4-MC exerts vasorelaxation by opening KV channels with the involvement of KV7.4.
Introduction
Hypertension is a multifactorial and, in its early stages, a silent disease that affects more than 30% of the adult population worldwide. It can cause or facilitate a cardiovascular event and even premature death.1 Arterial blood pressure is basically determined by two parameters: cardiac output and total peripheral resistance. In many cases, hypertensive patients have an unaltered cardiac output, whereas the total peripheral resistance is elevated.2 This fact highlights the major role played by blood vessels in the pathogenesis of hypertension. Indeed, small arteries and arterioles often appear more constricted and thicker in hypertension owing to abnormalities in vascular smooth muscle cells (VSMCs).3 Physiologically, the dynamic changes in vessel contractility are monitored by a tight interplay between the potassium (K+) and calcium (Ca2+) channels located in VSMCs. However, in chronic hypertension, vascular remodeling frequently occurs and contributes to an abnormal depolarized state of VSMCs.2,4 Flavonoids are polyphenolic substances that are abundant in foods such as vegetables, fruits, tea, red wine, and dark chocolate and have been positively linked to protective effects on the cardiovascular system.5,6 Indeed, epidemiological studies have suggested that a high intake of flavonoids is inversely related to arterial blood pressure.7 The probable explanation for this phenomenon is based on vasorelaxation as a consequence of flavonoid intake. However, flavonoids are not active in vivo in their parent forms as they are poorly absorbed.8 Hence, rather their metabolites, most commonly small phenolic compounds formed by human microflora, are responsible for such an effect. To date, a large repertoire of small phenolic metabolites has been identified and detected in human biological fluids9–11 and many have been shown to relax in different magnitude vessels in vitro or even in vivo.12–15 Yet, in most cases, the mechanisms of action of these metabolites remain enigmatic as there has not been much research in this field. 4-Methylcatechol (4-MC) is one such metabolite. It was suggested to be formed upon microbial catabolism of different classes of (poly)phenols (e.g., flavonol, quercetin, and its glycosides, flavan-3-ols, chlorogenic acids, Fig. 1).9,16 Although its detection in the urine of rats fed with vanillin was described as early as in the mid-1970s,17 its significant vasorelaxant effects were discovered by our team recently, and such data were confirmed in animals where 4-MC was able to decrease arterial blood pressure.14,15 More concretely, 4-MC induced relaxation of the rat aorta and mesenteric arteries in vitro, and caused a drop in the mean arterial pressure by 10% in spontaneously hypertensive rats after a single i.v. dose of 2.5 mg kg−1. Importantly, the observed effects were not mediated through the heart, and most likely relied on a decrease in the resistance of the vasculature.14 Therefore, the aim of the current study was to unravel the specific mechanisms that mediate the vasodilatory action of 4-MC. To achieve this, we carried out in vitro experiments on the isolated rat aorta, and in silico reverse docking studies. | ||
| Fig. 1 Simplified scheme of the suggested metabolic pathways for distinct classes of polyphenols leading to the formation of 4-methylcatechol: (a) procyanidin (PC), (b) chlorogenic acids, (c) flavonol rutin, and (d) vanillin. Data were obtained from the following ref. 9 and 17–22. | ||
Materials and methods
Animals
For the in vitro experiments, male Wistar Han rats (n = 32) obtained from Charles River, Germany (av. weight 300 g) were used. The animals were bred in the animal facility of the Faculty of Pharmacy, Charles University, and maintained at a constant temperature of 23–25 °C with the 12 h dark/light cycle. Rats were provided a standard diet and tap water ad libitum. The study was conformed to The Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (8th edition, revised 2011, ISBN-13:978-0-309-15400-0), and was performed in agreement with approval of the Czech Ministry of Education, Youth and Sports (No. 4937_2019-9).Chemicals
Urethane, noradrenaline, acetylcholine, sodium nitroprusside (SNP), dimethyl sulfoxide (DMSO), nifedipine, 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ), membrane-permeable protein kinase G Iα inhibitory peptide: RQIKIWFQNRRMKWKKLRKKKKKH (DT-3), forskolin, bradykinin, iberiotoxin, glibenclamide, BaCl2, 4-aminopyridine, linopirdine, and thapsigargin, as well as the tested compound 4-MC were purchased from Sigma-Aldrich (Germany). Bay K8644 was obtained from Axon Medchem BV (Netherlands). 4-MC was dissolved in 100% DMSO (10 mM), and the subsequent solutions were prepared with ultrapure water. KCl and CaCl2 were purchased from Sigma-Aldrich. NaCl, NaHCO3, and D-glucose were provided by PENTA s.r.o. (Czech Republic), MgSO4·7H2O was purchased from Erba Lachema (Czech Republic) s.r.o., whereas KH2PO4 was obtained from Dr Kulich Pharma s.r.o. (Czech Republic). The Krebs solution contained the following composition in mM: NaCl, 119; KCl, 4.7; CaCl2, 1.25; KH2PO4, 1.18; MgSO4·7H2O, 1.17; NaHCO3, 25; and D-glucose, 11.In vitro experiments
| Targets | Activator (A)/inhibitor (I) |
|---|---|
| Ion channels | |
| L-type Ca2+ channels (CaV1.2) | Bay K8644 (A) |
| Nifedipine (I) | |
| BKCa (large conductance calcium- activated K+ channels) | Iberiotoxin (I) |
| KATP (ATP-sensitive K+ channels) | Glibenclamide (I) |
| KIR (inwardly-rectifier K+ channels) | Ba2+ (I) |
| KV (voltage-gated K+ channels) | 4-Aminopyridine (I) |
| KV7 (voltage-gated K+ channels KV7 subfamily) | Linopirdine (I) |
| Enzymes | |
| sGC (soluble guanylate cyclase) | Sodium nitroprusside (A) |
| ODQ (I) | |
| PKG-Iα (protein kinase G isoform Iα) | DT3 (I) |
| PKG-Iα,β (protein kinase G isoforms Iα,β) | Rp-8-pCPT-cGMPS (I) |
| AC (adenylate cyclase) | Forskolin (A) |
| ATPases | |
| SERCA (sarco/endoplasmic reticulum calcium ATPase) | Thapsigargin (I) |
Lastly, the aortic rings were treated with the sarco/endoplasmic reticulum calcium ATPase (SERCA) inhibitor thapsigargin (1 μM), followed by the addition of noradrenaline (10 μM) in the presence of 7 mM KCl. Once the contraction was stable, 4-MC was applied at a concentration causing ∼80% of the maximal relaxation. As the negative control, the rings were treated with the vehicle of the respective inhibitor in the incubation step.
Measurement of cAMP
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 cells per well, and left to grow for 2 days to reach full confluency. Cells were treated with 10 μL of inducer with or without 4-MC (both dissolved in DMSO) mixed with 390 μL of the media for 1 minute, and the reaction was stopped by the addition of 400 μL of 1% Triton X-100 in 0.1 M HCl. The suspension was incubated for 10 minutes in a 37 °C ultrasonic bath, and spun down for 2 min at 10500 g. The supernatant was stored at −80 °C until the cAMP was determined.
500g.
000 cells per well, and left to grow for 2 days to reach full confluency. Cells were treated with 10 μL of inducer with or without 4-MC (both dissolved in DMSO) mixed with 390 μL of the media for 1 minute, and the reaction was stopped by the addition of 400 μL of 1% Triton X-100 in 0.1 M HCl. The suspension was incubated for 10 minutes in a 37 °C ultrasonic bath, and spun down for 2 min at 10500 g. The supernatant was stored at −80 °C until the cAMP was determined.
500g.
The level of cAMP was assessed by ELISA (ab290713, Abcam, U.K.), according to the instructions of the manufacturer.
Molecular docking and interaction analysis
 | ||
| Fig. 2 Structures of 4-methylcatechol and the target proteins used for reverse docking. | ||
Results
Cyclic nucleotide pathways
In the first step, the possible involvement of the cyclic guanosine monophosphate (cGMP) or cyclic adenosine monophosphate (cAMP) pathways in the 4-MC-induced vasorelaxation was investigated. For this reason, SNP, a NO donor that activates soluble guanylyl cyclase (sGC), and forskolin, an activator of adenylate cyclase, were used. In the presence of 4-MC, the vasodilatory effects of both activators were potentiated (Fig. 3A and B). Due to this fact, the involvement of sGC, which produces cGMP, and PKG, which is a downstream target of cGMP, was analyzed. No significant differences were found after inhibition of sGC with ODQ (Fig. 3C), nor inhibition of different PKG isoforms; Iα by DT3 and Iα,β by Rp-8-pCPT-cGMPS (Fig. 3D). Hence, sGC and PKG are not the targets of 4-MC. To see the possible involvement of the cAMP pathway, the cAMP levels were measured in rat VSMC (ESI Fig. S1†). 4-MC was not able to modify the levels of cAMP in these cells, whether alone or in combination with forskolin, prostaglandin E2 or serotonin. As the cells had relatively high basal cAMP levels and only the combination of forskolin with theophylline (a non-selective inhibitor of phosphodiesterase) was able to markedly increase the cAMP levels, we decided to confirm these results in another model. Human platelets were selected, as they had very low basal levels of cAMP, and responded well to low concentration of forskolin. 4-MC did not modify the basal levels of cAMP in resting platelets, and it had also no impact on the raised levels of cAMP after treatment with forskolin, prostaglandin E2 or adenosine (ESI Fig. S1†). This suggests that 4-MC cannot activate adenylyl cyclase or block phosphodiesterase, in contrast to a clear impact of the non-selective phosphodiesterase inhibitor theophylline.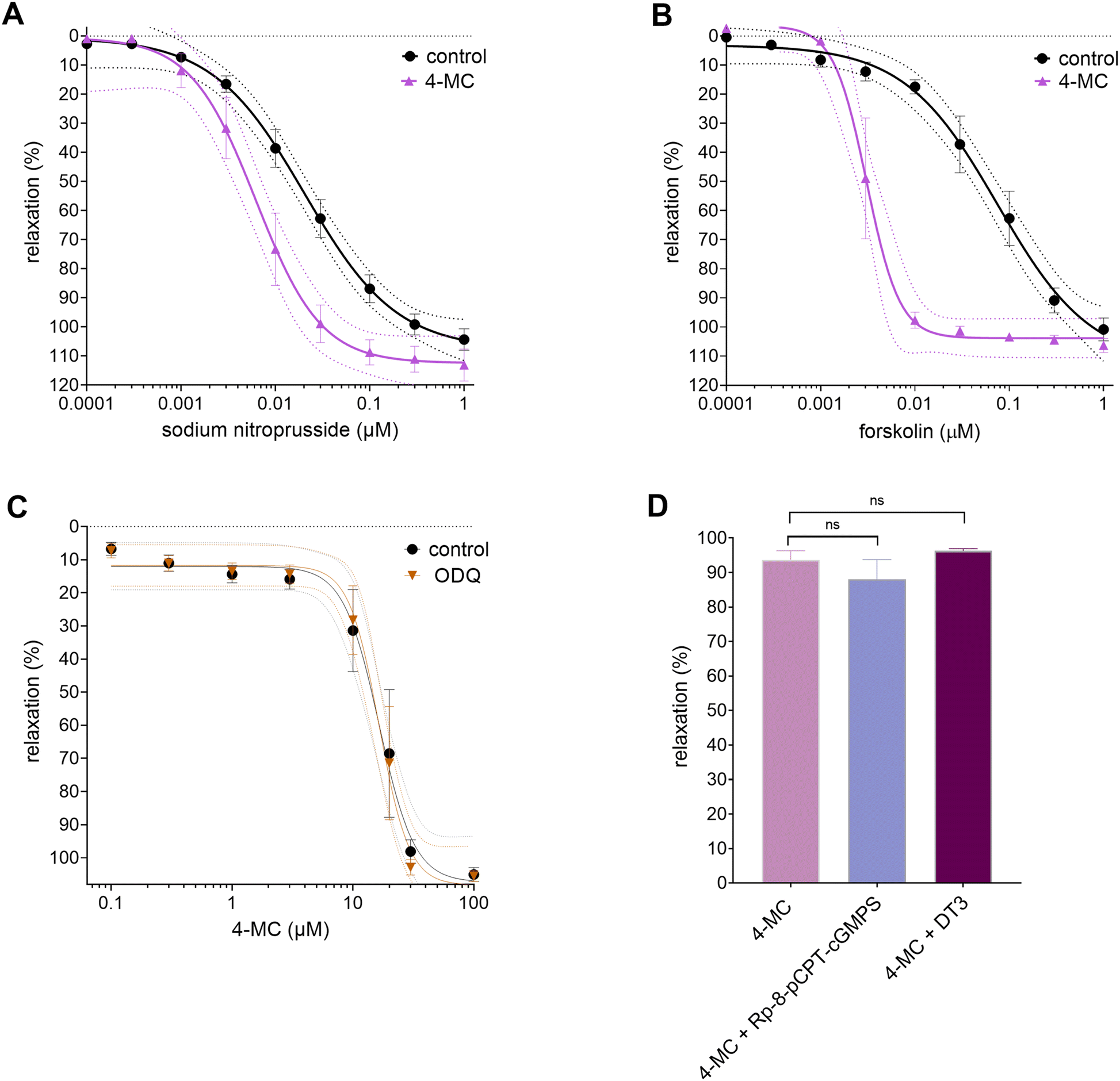 | ||
| Fig. 3 (A) Effect of 4-methylcatechol (4-MC, 15 μM) on relaxation induced by sodium nitroprusside. (B) Effect of 4-MC (15 μM) on relaxation induced by forskolin. (C) Concentration-dependent curves obtained with 4-MC in the presence and absence of the soluble guanylate cyclase inhibitor ODQ (1 μM). (D) Impact of inhibition of PKG-Iα,β isoforms by Rp-8-pCPT-cGMPS (1 μM) or DT-3 (1 μM) on 4-MC-induced vasorelaxation (70 μM, i.e., concentration able to induce ∼80% relaxation). Controls: (A and B) aortic rings were treated with cumulative concentrations of the activators sodium nitroprusside and forskolin in the absence of the tested compound 4-MC. (C) and (D) Aortic rings were treated with 4-MC in the absence of the inhibitor. Each point represents mean ± SEM from at least five separate experiments, except for (D), where n = 4 for each group. ns = not significant. | ||
As cyclic nucleotide pathways are likely not involved, attention was then given to ion channels, which are the other known downstream targets of cyclic nucleotide-mediated vasodilatory pathways.
BKCa, KATP, KIR and KV channels
Potassium channel opening is modulated by intracellular signaling pathways involving cyclic nucleotides (Fig. 4). The opening of vascular K+ channels leads to hyperpolarization of the VSMCs with consequent vasodilation. No significant differences were found before and after the inhibition of BKCa, KATP, or KIR channels with their respective inhibitors, iberiotoxin (Fig. 4A), glibenclamide (Fig. 4B), or Ba2+ (Fig. 4C). On the other hand, inhibition of KV channels by 4-aminopyridine caused about 60% reduction in the maximum relaxation caused by 4-MC (Fig. 4D). Furthermore, selective inhibition of KV7 channels with linopirdine also caused a significant but lower decrease in the vasodilatory effect, indicating that KV7 channels are partially involved in the effect of 4-MC (Fig. 4E). | ||
| Fig. 4 (A) Effect of inhibition of vascular potassium (K+) channels (BKCa, KATP, KIR and KV) on the vasodilatory activity of 4-methylcatechol (4-MC). Concentration-dependent curves for the vasorelaxation induced by 4-MC in the absence (controls) and presence of (A) iberiotoxin (20 nM), (B) glibenclamide (10 μM), (C) BaCl2 (50 μM), (D) 4-aminopyridine (3 mM), and (E) linopirdine (10 μM). Each point represents mean ± SEM from at least five separate experiments. BKCa, large conductance calcium-activated K+ channels; KATP, ATP-sensitive K+ channels; KIR, inwardly rectifying K+ channels; KV, voltage-gated K+ channels; ***p < 0.001 vs. control. | ||
L-type Ca2+ channels and SERCA
The excitability and contractility of VSMCs are regulated through the interplay between K+ and Ca2+ channels. L-type Ca2+ channels have the opposite role in mediating vasoconstriction due to VSMC depolarization. Therefore, in the next step, we sought to explore whether L-type Ca2+ channels were inhibited in the presence of 4-MC. In this case, an L-type Ca2+ channel activator, Bay K8644, was cumulatively added to the aortic rings to induce a contractile response. 4-MC did not abolish the Bay K8644-elicited vasoconstriction at any of the tested concentrations, i.e., low and high concentrations (30 and 70 μM), in contrast to the clinically used well-known L-type Ca2+ channel blocker nifedipine (Fig. 5A). A higher concentration of 4-MC even potentiated the effect of Bay K8644, rendering the possible inhibitory effect of 4-MC on the L-type Ca2+ channel highly improbable. The cytosolic calcium concentrations are also dependent on SERCA. For this reason, we decided to also test the hypothesis that 4-MC activates SERCA, as the observed vasodilation could be due to the decreasing cytosolic calcium. The pretreatment of the norepinephrine-precontracted aortic rings with the inhibitor of SERCA, thapsigargin, caused a significant but not complete decrease in the extent of relaxation induced by 4-MC, showing that this hypothesis might not be simply rejected (Fig. 5B).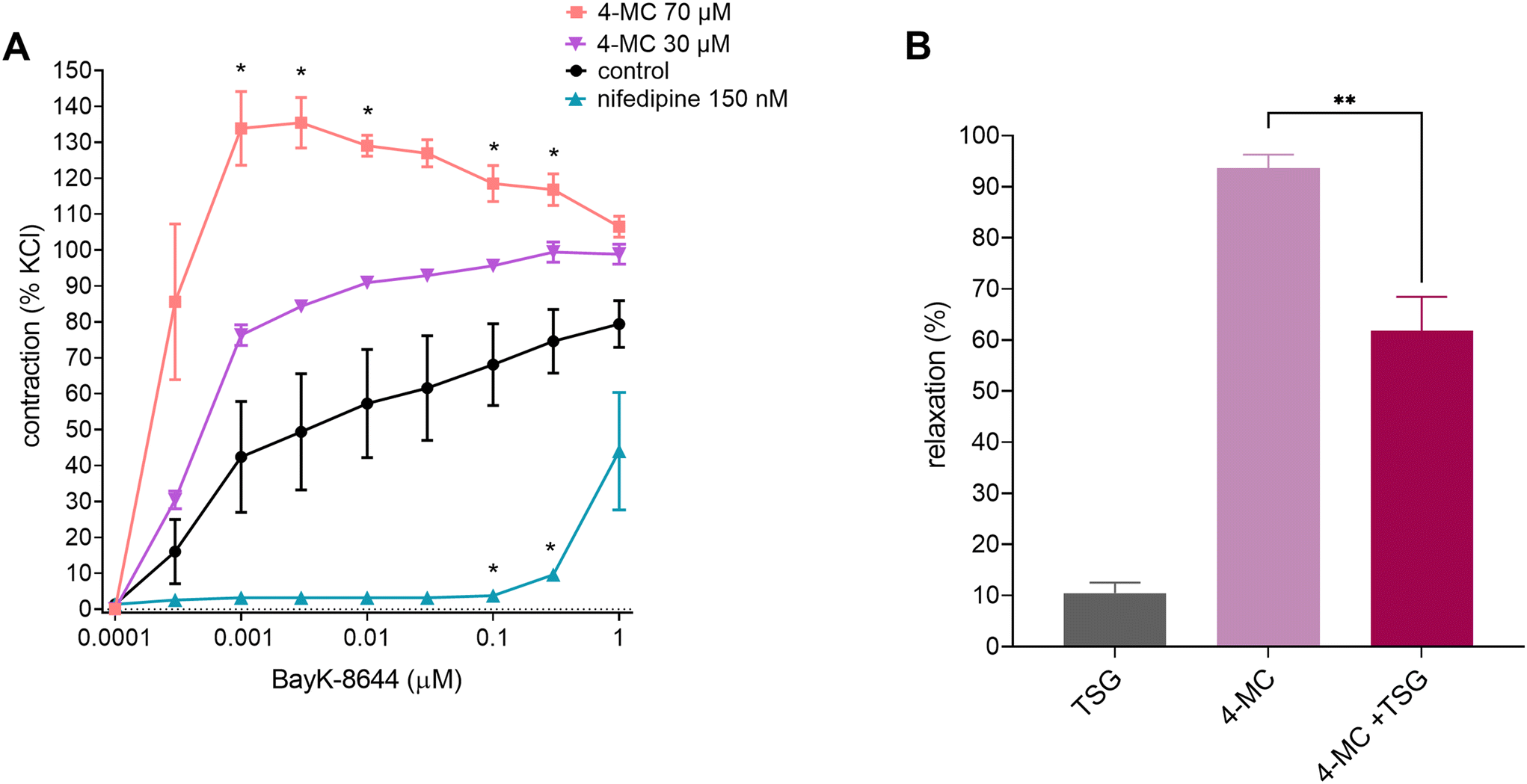 | ||
| Fig. 5 Effect of 4-methylcatechol (4-MC) on vessel tonus as a function of extracellular (A) and intracellular (B) calcium. (A) Effect of pretreatment of rat aortic rings with 4-MC (70 and 30 μM) on the Bay K8644 (100 pM–1 μM)-elicited contraction of aortic rings. Nifedipine (150 nM) and DMSO were used as the positive and negative controls, respectively. Data were calculated from the maximal contraction induced by KCl (60 mM). *p < 0.05 as compared to the negative control using two-way ANOVA, followed by the Holm-Šidák multiple comparison test. (B) Impact of the sarco/endoplasmic reticulum calcium ATPase (SERCA) inhibitor thapsigargin (TSG, 1 μM) on 4-MC (70 μM)-induced vasorelaxation. Vessels were precontracted with norepinephrine (10 μM), and the degree of vasodilation was measured thereafter. As the negative control, the rings were treated with thapsigargin (TSG) and the solvent DMSO. As the positive control, the relaxation-induced 4-MC was measured in the absence of TSG. **p < 0.01 as compared to the positive control using Student's unpaired t-test. Each point represents the mean ± SEM from at least five separate experiments, except for (B) n = 4 for control, n = 5 for 4-MC and n = 5 for 4-MC + TSG. | ||
Molecular docking and intermolecular interaction
One of the hydroxyl groups on 4-MC undergoes sulfonation in the circulation of plasma. Therefore, docking studies were performed not only on 4-MC, but also on 4-methylcatechol-1-sulfate (4-MC-1-S) and 4-methylcatechol-2-sulfate (4-MC-2-S). The docking scores of 4-MC, 4-MC-1-S and 4-MC-2-S (Table 2 and Table S1†) were lower than all reference ligands of each target protein, but showed higher ligand efficiencies than the reference ligands, particularly in the case of 4-MC. The ligand efficiency is a measurement of the binding energy per non-hydrogen atom of the ligand on the target protein, or simply the free binding energy divided by the number of heavy atoms.27 Ligand efficiency can be used to determine whether the ligand has an optimal position on the binding site of the protein through the formation of multiple contacts.27| PDB ID | Target protein | Docking score (kcal mol−1) | Docking score differences | |
|---|---|---|---|---|
| Reference ligand | 4-MC | (Reference ligand) – (4-MC) | ||
| PDB ID, Protein Data Bank Identifiers; sGC, soluble guanylate cyclase; PKGIα, protein kinase G isoform Iα; PKGIβ, protein kinase G isoform Iβ; PKA, protein kinase A; KV, voltage-gated potassium channel; AC, adenylate cyclase; SERCA, sarco/endoplasmic reticulum calcium-ATPase. | ||||
| 7D9T | sGC | −10.7 | −5.3 | 5.4 |
| 6C0T | PKGIα | −8.2 | −6.5 | 1.7 |
| 5JAX | PKGIβ | −11.6 | −6.0 | 5.6 |
| 3POO | PKA | −9.3 | −5.9 | 3.4 |
| 7TCI | KV7.1 | −8.8 | −6.0 | 2.8 |
| 7CR1 | KV7.2 | −8.1 | −6.1 | 2.0 |
| 7BYM | KV7.4 | −7.6 | −6.0 | 1.6 |
| 1CJK | AC | −9.7 | −5.6 | 4.1 |
| 6JJU | SERCA | −9.7 | −5.5 | 4.2 |
Based on the ligand efficiency value, 4-MC (>0.65) showed the potential to interact with PKGIα, PKA, KV7.1, KV7.2, and KV7.4, which meant that they can be possible 4-MC targets (Table 2).
These data agree with most of our experimental results, concluding that sGC, PKGIβ, adenylate cyclase, and possibly SERCA can be excluded from the list of potential targets of 4-MC. Interestingly, in the case of PKGIα and PKA, their ligand efficacy values were higher. However, as in the prior in vitro experiments, the inhibitor DT3 did not affect the vasodilatory effects of 4-MC. PKGIα was also ruled out as a potential target, in addition to PKA. On the other hand, our docking results further support the hypothesis that KV channels, in particular KV7.1, KV7.2, and KV7.4, might be potential targets of 4-MC (Table 2).
Molecular docking approaches allow the modelling of the interactions between ligands and proteins at the atomic level, enabling the characterization of ligand behaviour in the ligand-binding domain to elucidate fundamental biochemical processes.28 Overall, the docking poses of 4-MC, and its 1- and 2-sulfates are similar to each other, and have analogous interactions with each target protein (Fig. S2–7†). The docking of 4-MC on a binding domain of the protein that overlaps with a reference ligand reflects the higher chances of biologically relevant interactions and biochemical effects.
The best-docked poses of 4-MC on the ligand-binding domains of the abovementioned improbable targets (namely, adenylate cyclase, PKA, sGC, PKGIα, PKGIβ and SERCA) are shown as complementary results in the supplementary data (Fig. S2–S4†). Notably, at this stage of our study, the compilation of the ligand efficiency and the best docked poses confirmed, as initially hypothesized, that KV channels are involved in the vasodilatory effect of 4-MC. Although the above-reported docking scores suggested that KV7.1 could be the target, the participation of this channel is improbable because the best-docked pose of 4-MC did not overlap with the reference ligand ML277 (Fig. 6A). The molecular interactions between 4-MC or the reference ligand at the ligand binding domain of each target protein are summarized in Table S1.† Some residues involved in the interaction between 4-MC and the target protein were the same as in the interaction between the reference ligand and the protein.
 | ||
| Fig. 6 Docking pose of 4-methylcatechol (green), 4-methylcatechol-1-sulfate (yellow), 4-methylcatechol-2-sulfate (purple), and retigabine (turquoise) in the ligand binding domain of the human voltage-gated potassium channel: (A) KV7.1; (B) KV7.2; (C) KV7.4. | ||
In the case of the KV7.2 channel, the best docked pose of 4-MC overlapped with the reference ligand retigabine (Fig. 6B), and both ligands formed H-bonds with Trp236 (Fig. 7). Moreover, Phe305 and Leu299 were shared residues, albeit with different interactions involved; namely, H-bonds in the case of retigabine, and hydrophobic contacts in the case of 4-MC (Fig. 7). Concerning the KV7.4 channel, besides the observed overlapping docked poses between 4-MC and the reference ligand retigabine (Fig. 6C), similar types of interactions were found. In some cases, they were formed with shared residues. It is noteworthy that some of the residues involved, namely Trp242 and Phe246, are crucial for the activation of the KV7.4 channel.29 The best-docked pose of 4-MC in the KV7.4 binding cavity was stabilized by H-bonds and hydrophobic interactions. H-bonds were formed between the hydroxyl group of 4-MC (Fig. 7) and the amine groups of Ala241 and Phe246, and the carboxyl group of Phe310, amino acids present on the KV7.4 protein backbone. Similarly, H-bonds (Fig. 7) were established between retigabine and specific residues of the KV7.4 protein backbone: (1) an O atom and the Trp242 side chain, (2) an amine group with the Ser309 side chain, and (3) an amine group with the Leu305 and Phe311 carboxyl groups. Interestingly, due to the proximity of the benzyl groups of both ligands to the aromatic side chain of Trp242, the same π–π interaction with Trp242 was found for both 4-MC and retigabine (Fig. 7). Furthermore, retigabine and 4-MC interacted with Phe246, albeit distinctly, through π–π and H-bond interactions (Fig. 7), respectively.
 | ||
| Fig. 7 2D representation of molecular interactions between the reference ligand (ML277 for KV7.1 and retigabine for KV7.2; 7.4), 4-methylcatechol (MC), 4-methylcatechol-1-sulfate (4-MC-1-S), and 4-methylcatechol-2-sulfate (4-MC-2-S). Dashed green lines show the hydrogen bond interactions; dashed purple lines indicate the hydrophobic interactions. | ||
Discussion
In the current study, we set out to elucidate the mechanism of the vasodilatory activity of 4-MC as a follow-up to our previous research.15 To achieve a more complex picture, both in vitro and in silico approaches were employed. The obtained results apparently fit together and enabled us to overcome the inherited problems of experiments, e.g., the absence of known and selective activators or inhibitors, and docking, i.e., the need for experimental confirmation of in silico results. In reverse docking, the docking score between the ligand and the target protein serves as an evaluation criterion to rank the potential targets of the ligand. To accomplish target evaluation, reverse docking analysed whether the ligand could fit into the ligand binding site of the target protein, and evaluated the critical interactions between the ligand and protein. These two factors were used to determine the potential target of 4-MC.In the presence of 4-MC, a potentiation of the vasodilation of the rat aorta mediated by cGMP- and cAMP-pathways was observed. As the hypothesis of direct activation of sGC/PKG or impact on cAMP was subsequently ruled out, we concentrated on other downstream targets of cyclic nucleotide pathways, particularly ion channels, which are known to be involved in vasodilation/constriction of arteries. Interference with calcium influx through L-type Ca2+ channels can be excluded, but the involvement of potassium (KV) channels and specifically the KV7 subtype was observed.
Nevertheless, the mechanism of 4-MC-induced vasorelaxation is not solely based on the activation of KV7 channels. Moreover, inhibition of all KV channels by 4-AP did not fully abolish the vasodilatory effect of 4-MC (Fig. 4D). Besides that, the extent to which the selective inhibitor of KV7 channels, linopirdine, lessened the maximum vasodilatory effect of 4-MC was much lower than that caused by 4-AP (∼15% vs. ∼60% decrease, Fig. 4E and D), meaning that other subfamilies of K+ channels could be involved. In fact, the family of KV channels is highly complex and diverse with 12 subfamilies (KV1–KV12) in humans.30 In SMCs, besides the KV7 channels, KV1, KV2, and KV4 channels are also expressed.3,31 Moreover, the distribution of each subtype depends on the species and the vascular bed. To obtain a framework, KV1.2, KV1.5, KV2.1, KV7.1, KV7.4, and KV7.5 were identified in blood vessels from rodents, rabbits, dogs, and humans.32 The role of vascular KV7 channels should neither be underestimated nor overestimated. This channel family provides roughly 20% of cell ion currents (at −40 mV),33 and participates in the regulation of the resting membrane potential and the negative feedback of myogenic and vasoconstrictor-induced tone. Among the KV7 channel subtypes, KV7.4 together with KV7.5 likely play major roles.34–36 The KV7.4 channel family can be influenced by several vasoactive stimuli (e.g., activation of the cAMP/PKA pathway,37 cGMP/PKG pathway38) in several vascular beds like the renal VSMCs,37 coronary39 and cerebral arteries.40 During vasoconstriction, the activated protein kinase C was reported to inhibit the KV7 channels.41 Thus, the KV7 channels plays an important role in vascular physiology. Nevertheless, it should be kept in mind that the regulation of vascular tone is very complex and involves the interplay of numerous factors.
Complete experimental testing is currently not possible due to the lack of specific activators or inhibitors. Hence, we cannot exclude the fact that other KV channel subfamilies, not involved in this study, might also be involved. Regardless, our docking study suggested the involvement of the KV7.2 and 7.4 channels. There are additional drawbacks of experimental functional studies with activators/inhibitors, as they cannot provide direct insight into the type of 4-MC interaction with KV channels, i.e., direct or indirect. Such limitation can be resolved by supplementation with in silico data. Indeed, our in silico data showed that 4-MC could directly bind to KV channels, further supporting our hypothesis. Concerning KV7.2, H-bonds formed with the residues Phe305 and Ser303 are considered relevant for the stabilization of retigabine.42 However, the results showed that 4-MC forms hydrophobic interactions with Phe305 (Fig. 7). Moreover, KV7.2 channels are mostly expressed in the central nervous system, where together with KV7.3, they form the “M channel”.43 On the other hand, KV7.4 channels are expressed in the mammalian vasculature,44 and their involvement was most obvious and strongly supported by the following observations: (a) a very low difference in docking scores (−1.6 kcal mol−1) (Table 2), (b) the best-docked pose for 4-MC that overlapped with that of the reference ligand retigabine (Fig. 7), and (c) the intermolecular interaction analysis demonstrated that crucial residues for KV7.4 activation are involved (Fig. 7).
KV channels are major determinants of the membrane potential in VSMCs, and are thus essential for the regulation of the vascular tone. Studies have reported on alterations in the function of these channels in cardiovascular diseases. Particularly, a decrease in KV7.4 protein levels was reported in animal models of hypertension.45
Although scarce, bioavailability studies reported on human plasma maximum concentrations for the sulphated form of 4-MC ranging from ∼2 to 3.5 μM, which were attained within 6 h to 7 h after the ingestion of a mixed berry fruit purée (blueberry, blackberry, raspberry, strawberry tree fruit, and Portuguese crowberry) or cranberry juice.9,46 4-MC sulphate was also identified in human plasma after the consumption of natural and enriched apple juices.16 Therefore, relatively high plasma concentrations of 4-MC can be reached after the ingestion of foods rich in polyphenols. Thus, by targeting KV7.4 channels, 4-MC could likely contribute to the prevention of smooth muscle remodeling. Moreover, it is noteworthy that even before the consumption of the purée, this conjugate was already present in plasma from the volunteers, albeit at lower levels, i.e., ∼300 nM.46
The docking study also allows us to answer the question of whether SERCA is involved. Experimental testing showed that inhibition of SERCA by thapsigargin decreased the vasodilation caused by 4-MC (Fig. 5B). This suggested that SERCA could be a possible target. Such experimental data could not bring a clear answer, as there is a tight interplay of K+ and Ca2+ currents in modulating the VSMC contractility. The docking study does not confirm 4-MC interaction with SERCA, so the likely explanation is that an elevation in intracellular Ca2+ decreased the vasoactive effect of 4-MC mediated by K+ channels.
Nevertheless, the intriguing nature of the mechanism of 4-MC is evidenced by the fact that with increasing concentrations of 4-MC, a potentiation of the vasocontractile response induced by Bay K8644 was observed (Fig. 5A). The meaning of these findings remains to be clarified. At first sight, it seems a paradox since with increasing cytosolic Ca2+ concentrations, VSMCs become depolarized, thereby promoting the activation of KV channels. It is possible that 4-MC can somehow affect the effect of Bay K8644, which is also dependent on other conditions.47,48 Interestingly, others have reported that the parent compound, quercetin, reduces the efficacy of Bay K8644.49 Under specific conditions favouring the depolarisation of rat arteriae, both quercetin and Bay K8644 potentiated the response to calcium.50 Nevertheless, it should be also mentioned that 4-MC has a different mechanism at least in other cells, as we recently showed that its antiplatelet potential is based on interference with cyclooxygenase–thromboxane synthase coupling.51 Notably, the diverse mechanisms of action of 4-MC are reflected in pleiotropic effects that include vasodilatory and antiplatelet activities. In addition, it is important to emphasize the significance of exploring the metabolic pathways, metabolites formed, as well as their pharmacokinetics to better understand the beneficial effects of a diet rich in polyphenols. Under physiological conditions, 4-MC and other metabolites are formed, and may exert synergistic effects. In addition, the existence of large interindividual variability in the microbiota catabolic output16,52 presents a challenge for further study.
Conclusions
In summary, our findings suggest that the dietary polyphenol metabolite, 4-MC, relaxes the rat aorta through the opening of KV channels, with partial involvement of the KV7 subfamily. In silico molecular docking studies corroborated the in vitro results, and deepened our study at the molecular level. The results showed the overlapping best-docked poses of 4-MC and that of retigabine (the reference ligand) at KV7.4, as well as similar interactions with crucial residues, namely Trp242 and Phe246. However, the existence of other mechanisms apparently contributing to the relaxation of the vasculature by 4-MC is unambiguous. Given the reported pleiotropic effects of 4-MC on the cardiovascular system, further research on this metabolite and its sulfate conjugates is warranted.Author contributions
Patrícia Dias: writing – original draft, investigation & methodology, data curation, writing – review & editing; Rudy Salam: writing – original draft, investigation & methodology, data curation, writing – review & editing; Jana Pourová: supervision, investigation & methodology, writing – review & editing; Marie Vopršalová: investigation & validation; Lukáš Konečný and Eduard Jirkovský: investigation & methodology, data curation, writing – review & editing; Jurjen Duintjer Tebbens: data curation and formal analysis, supervision; Přemysl Mladěnka: supervision, writing – original draft, review & editing, critical revision of the obtained findings and manuscript content and funding acquisition.Data availability
The data for this article, including a description, are available at https://zenodo.org/records/13739740. There are supplementary data available for this paper (https://www.rsc.org/suppdata/d3/fo/d3fo04672a/d3fo04672a1.pdf) as well as research data.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by the Czech Health Research Council (grant number NU21-02-00135). P. D., R. S. and J. D. T. thank Charles University (GAUK 136120, SVV 260 663, SVV 260 666 and UNCE 24/MED/008).References
- K. T. Mills, A. Stefanescu and J. He, The global epidemiology of hypertension, Nat. Rev. Nephrol., 2020, 16, 223–237 CrossRef CAS PubMed.
- B. K. Joseph, K. M. Thakali, C. L. Moore and S. W. Rhee, Ion channel remodeling in vascular smooth muscle during hypertension: Implications for novel therapeutic approaches, Pharmacol. Res., 2013, 70, 126–138 CrossRef CAS.
- T. A. Jepps, Unravelling the complexities of vascular smooth muscle ion channels: Fine tuning of activity by ancillary subunits, Pharmacol. Ther., 2017, 178, 57–66 CrossRef CAS.
- E. A. Pereira da Silva, M. Martín-Aragón Baudel, M. F. Navedo and M. Nieves-Cintrón, Ion channel molecular complexes in vascular smooth muscle, Front. Physiol., 2022, 13, 999369 CrossRef PubMed.
- P. Mladenka, L. Zatloukalová, T. Filipský and R. Hrdina, Cardiovascular effects of flavonoids are not caused only by direct antioxidant activity, Free Radicals Biol. Med., 2010, 49, 963–975 CrossRef CAS.
- C. Del Bo, S. Bernardi, M. Marino, M. Porrini, M. Tucci, S. Guglielmetti, A. Cherubini, B. Carrieri, B. Kirkup, P. Kroon, R. Zamora-Ros, N. H. Liberona, C. Andres-Lacueva and P. Riso, Systematic Review on Polyphenol Intake and Health Outcomes: Is there Sufficient Evidence to Define a Health-Promoting Polyphenol-Rich Dietary Pattern?, Nutrients, 2019, 11, 1355 CrossRef CAS.
- S. Ghaffari and N. Roshanravan, The role of nutraceuticals in prevention and treatment of hypertension: An updated review of the literature, Food Res. Int., 2020, 128, 108749 CrossRef CAS PubMed.
- H.-H. Xiong, S.-Y. Lin, L.-L. Chen, K.-H. Ouyang and W.-J. Wang, The Interaction between Flavonoids and Intestinal Microbes: A Review, Foods, 2023, 12, 320 CrossRef CAS.
- R. P. Feliciano, A. Boeres, L. Massacessi, G. Istas, M. R. Ventura, C. Nunes dos Santos, C. Heiss and A. Rodriguez-Mateos, Identification and quantification of novel cranberry-derived plasma and urinary (poly)phenols, Arch. Biochem. Biophys., 2016, 599, 31–41 CrossRef CAS.
- X. Feng, Y. Li, M. Brobbey Oppong and F. Qiu, Insights into the intestinal bacterial metabolism of flavonoids and the bioactivities of their microbe-derived ring cleavage metabolites, Drug Metab. Rev., 2018, 50, 343–356 CrossRef CAS PubMed.
- C. Vetrani, A. A. Rivellese, G. Annuzzi, I. Mattila, E. Meudec, T. Hyötyläinen, M. Orešič and A.-M. Aura, Phenolic metabolites as compliance biomarker for polyphenol intake in a randomized controlled human intervention, Food Res. Int., 2014, 63, 233–238 CrossRef CAS.
- P. Dias, J. Pourová, M. Vopršalová, I. Nejmanová and P. Mladěnka, 3-Hydroxyphenylacetic Acid: A Blood Pressure-Reducing Flavonoid Metabolite, Nutrients, 2022, 14, 328 CrossRef CAS.
- I. Najmanová, J. Pourová and P. Mladěnka, A Mixture of Phenolic Metabolites of Quercetin Can Decrease Elevated Blood Pressure of Spontaneously Hypertensive Rats Even in Low Doses, Nutrients, 2020, 12, 213 CrossRef PubMed.
- J. Pourova, I. Najmanova, M. Voprsalova, T. Migkos, V. Pilarova, L. Applova, L. Novakova and P. Mladenka, Two flavonoid metabolites, 3,4-dihydroxyphenylacetic acid and 4-methylcatechol, relax arteries ex vivo and decrease blood pressure in vivo, Vasc. Pharmacol., 2018, 111, 36–43 CrossRef CAS.
- I. Najmanová, J. Pourová, M. Vopršalová, V. Pilařová, V. Semecký, L. Nováková and P. Mladěnka, Flavonoid metabolite 3-(3-hydroxyphenyl)propionic acid formed by human microflora decreases arterial blood pressure in rats, Mol. Nutr. Food Res., 2016, 60, 981–991 CrossRef.
- K. Trošt, M. M. Ulaszewska, J. Stanstrup, D. Albanese, C. De Filippo, K. M. Tuohy, F. Natella, C. Scaccini and F. Mattivi, Host: Microbiome co-metabolic processing of dietary polyphenols – An acute, single blinded, cross-over study with different doses of apple polyphenols in healthy subjects, Food Res. Int., 2018, 112, 108–128 CrossRef.
- L. P. Strand and R. R. Scheline, The Metabolism of Vanillin and Isovanillin in the Rat, Xenobiotica, 1975, 5, 49–63 CrossRef CAS.
- M. M. Appeldoorn, J.-P. Vincken, A.-M. Aura, P. C. H. Hollman and H. Gruppen, Procyanidin Dimers Are Metabolized by Human Microbiota with 2-(3,4-Dihydroxyphenyl)acetic Acid and 5-(3,4-Dihydroxyphenyl)-γ-valerolactone as the Major Metabolites, J. Agric. Food Chem., 2009, 57, 1084–1092 CrossRef CAS PubMed.
- M. R. Olthof, P. C. Hollman, M. N. Buijsman, J. M. van Amelsvoort and M. B. Katan, Chlorogenic acid, quercetin-3-rutinoside and black tea phenols are extensively metabolized in humans, J. Nutr., 2003, 133, 1806–1814 CrossRef CAS.
- A. Serra, A. Macià, M.-P. Romero, J. Reguant, N. Ortega and M.-J. Motilva, Metabolic pathways of the colonic metabolism of flavonoids (flavonols, flavones and flavanones) and phenolic acids, Food Chem., 2012, 130, 383–393 CrossRef CAS.
- A. Serra, A. Macià, M.-P. Romero, N. Anglés, J.-R. Morelló and M.-J. Motilva, Metabolic pathways of the colonic metabolism of procyanidins (monomers and dimers) and alkaloids, Food Chem., 2011, 126, 1127–1137 CrossRef CAS.
- M. M. Ulaszewska, K. Trost, J. Stanstrup, K. M. Tuohy, P. Franceschi, M. F.-F. Chong, T. George, A. M. Minihane, J. A. Lovegrove and F. Mattivi, Urinary metabolomic profiling to identify biomarkers of a flavonoid-rich and flavonoid-poor fruits and vegetables diet in adults: the FLAVURS trial, Metabolomics, 2016, 12, 32 CrossRef.
- S. Kim, J. Chen, T. Cheng, A. Gindulyte, J. He, S. He, Q. Li, B. A. Shoemaker, P. A. Thiessen, B. Yu, L. Zaslavsky, J. Zhang and E. E. Bolton, PubChem in 2021: new data content and improved web interfaces, Nucleic Acids Res., 2020, 49, D1388–D1395 CrossRef.
- S. K. Burley, C. Bhikadiya, C. Bi, S. Bittrich, L. Chen, G. V. Crichlow, C. H. Christie, K. Dalenberg, L. Di Costanzo, J. M. Duarte, S. Dutta, Z. Feng, S. Ganesan, D. S. Goodsell, S. Ghosh, R. K. Green, V. Guranović, D. Guzenko, B. P. Hudson, C. L. Lawson, Y. Liang, R. Lowe, H. Namkoong, E. Peisach, I. Persikova, C. Randle, A. Rose, Y. Rose, A. Sali, J. Segura, M. Sekharan, C. Shao, Y. P. Tao, M. Voigt, J. D. Westbrook, J. Y. Young, C. Zardecki and M. Zhuravleva, RCSB Protein Data Bank: powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences, Nucleic Acids Res., 2021, 49, D437–D451 CrossRef CAS PubMed.
- O. Trott and A. J. Olson, AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading, J. Comput. Chem., 2010, 31, 455–461 CrossRef CAS.
- E. F. Pettersen, T. D. Goddard, C. C. Huang, G. S. Couch, D. M. Greenblatt, E. C. Meng and T. E. Ferrin, UCSF Chimera—A visualization system for exploratory research and analysis, J. Comput. Chem., 2004, 25, 1605–1612 CrossRef CAS.
- C. H. Reynolds, S. D. Bembenek and B. A. Tounge, The role of molecular size in ligand efficiency, Bioorg. Med. Chem. Lett., 2007, 17, 4258–4261 CrossRef CAS PubMed.
- B. J. McConkey, V. Sobolev and M. Edelman, The performance of current methods in ligand-protein docking, Curr. Sci., 2002, 83, 845–856 CAS.
- T. Li, K. Wu, Z. Yue, Y. Wang, F. Zhang and H. Shen, Structural Basis for the Modulation of Human KCNQ4 by Small-Molecule Drugs, Mol. Cell, 2021, 81, 25–37.e4 CrossRef CAS PubMed.
- S. P. Alexander, J. Striessnig, E. Kelly, N. V. Marrion, J. A. Peters, E. Faccenda, S. D. Harding, A. J. Pawson, J. L. Sharman, C. Southan, J. A. Davies and C. Collaborators, THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: Voltage-gated ion channels, Br. J. Pharmacol., 2017, 174, S160–S194 CAS.
- N. R. Tykocki, E. M. Boerman and W. F. Jackson, Smooth Muscle Ion Channels and Regulation of Vascular Tone in Resistance Arteries and Arterioles, Compr. Physiol., 2017, 7, 485–581 Search PubMed.
- J. B. Stott, T. A. Jepps and I. A. Greenwood, KV7 potassium channels: a new therapeutic target in smooth muscle disorders, Drug Discovery Today, 2014, 19, 413–424 CrossRef CAS.
- R. H. Cox and S. Fromme, Functional Expression Profile of Voltage-Gated K(+) Channel Subunits in Rat Small Mesenteric Arteries, Cell Biochem. Biophys., 2016, 74, 263–276 CrossRef CAS PubMed.
- I. A. Greenwood and S. Ohya, New tricks for old dogs: KCNQ expression and role in smooth muscle, Br. J. Pharmacol., 2009, 156, 1196–1203 CrossRef CAS.
- T. A. Jepps, S. P. Olesen and I. A. Greenwood, One man's side effect is another man's therapeutic opportunity: targeting Kv7 channels in smooth muscle disorders, Br. J. Pharmacol., 2013, 168, 19–27 CrossRef CAS PubMed.
- W. F. Jackson, K(V) channels and the regulation of vascular smooth muscle tone, Microcirculation, 2018, 25 DOI:10.1111/micc.12421.
- P. S. Chadha, F. Zunke, H. L. Zhu, A. J. Davis, T. A. Jepps, S. P. Olesen, W. C. Cole, J. D. Moffatt and I. A. Greenwood, Reduced KCNQ4-encoded voltage-dependent potassium channel activity underlies impaired β-adrenoceptor-mediated relaxation of renal arteries in hypertension, Hypertension, 2012, 59, 877–884 CrossRef CAS PubMed.
- J. B. Stott, V. Barrese, T. A. Jepps, E. V. Leighton and I. A. Greenwood, Contribution of Kv7 channels to natriuretic peptide mediated vasodilation in normal and hypertensive rats, Hypertension, 2015, 65, 676–682 CrossRef CAS PubMed.
- S. Khanamiri, E. Soltysinska, T. A. Jepps, B. H. Bentzen, P. S. Chadha, N. Schmitt, I. A. Greenwood and S. P. Olesen, Contribution of Kv7 channels to basal coronary flow and active response to ischemia, Hypertension, 2013, 62, 1090–1097 CrossRef CAS PubMed.
- P. S. Chadha, T. A. Jepps, G. Carr, J. B. Stott, H. L. Zhu, W. C. Cole and I. A. Greenwood, Contribution of kv7.4/kv7.5 heteromers to intrinsic and calcitonin gene-related peptide-induced cerebral reactivity, Arterioscler., Thromb., Vasc. Biol., 2014, 34, 887–893 CrossRef CAS PubMed.
- A. R. Mackie, L. I. Brueggemann, K. K. Henderson, A. J. Shiels, L. L. Cribbs, K. E. Scrogin and K. L. Byron, Vascular KCNQ potassium channels as novel targets for the control of mesenteric artery constriction by vasopressin, based on studies in single cells, pressurized arteries, and in vivo measurements of mesenteric vascular resistance, J. Pharmacol. Exp. Ther., 2008, 325, 475–483 CrossRef CAS PubMed.
- X. Li, Q. Zhang, P. Guo, J. Fu, L. Mei, D. Lv, J. Wang, D. Lai, S. Ye, H. Yang and J. Guo, Molecular basis for ligand activation of the human KCNQ2 channel, Cell Res., 2021, 31, 52–61 CrossRef CAS PubMed.
- D. A. Brown and G. M. Passmore, Neural KCNQ (Kv7) channels, Br. J. Pharmacol., 2009, 156, 1185–1195 CrossRef CAS PubMed.
- A. L. Fosmo and Ø. B. Skraastad, The Kv7 Channel and Cardiovascular Risk Factors, Front. Cardiovasc. Med., 2017, 4, 75 CrossRef PubMed.
- T. A. Jepps, P. S. Chadha, A. J. Davis, M. I. Harhun, G. W. Cockerill, S. P. Olesen, R. S. Hansen and I. A. Greenwood, Downregulation of Kv7.4 channel activity in primary and secondary hypertension, Circulation, 2011, 124, 602–611 CrossRef CAS PubMed.
- R. C. Pimpão, M. R. Ventura, R. B. Ferreira, G. Williamson and C. N. Santos, Phenolic sulfates as new and highly abundant metabolites in human plasma after ingestion of a mixed berry fruit purée, Br. J. Nutr., 2015, 113, 454–463 CrossRef PubMed.
- G. Thomas, M. Chung and C. J. Cohen, A dihydropyridine (Bay k 8644) that enhances calcium currents in guinea pig and calf myocardial cells. A new type of positive inotropic agent, Circ. Res., 1985, 56, 87–96 CrossRef CAS PubMed.
- C. Van Hove, C. Van der Donckt, A. Herman, H. Bult and P. Fransen, Vasodilator efficacy of nitric oxide depends on mechanisms of intracellular calcium mobilization in mouse aortic smooth muscle cells, Br. J. Pharmacol., 2009, 158, 920–930 CrossRef CAS PubMed.
- S. Saponara, G. Sgaragli and F. Fusi, Quercetin antagonism of Bay K 8644 effects on rat tail artery L-type Ca(2+) channels, Eur. J. Pharmacol., 2008, 598, 75–80 CrossRef CAS PubMed.
- A. Trezza, O. Spiga, P. Mugnai, S. Saponara, G. Sgaragli and F. Fusi, Functional, electrophysiology, and molecular dynamics analysis of quercetin-induced contraction of rat vascular musculature, Eur. J. Pharmacol., 2022, 918, 174778 CrossRef CAS PubMed.
- M. Hrubša, L. Konečný, M. Paclíková, M. S. Parvin, P. Skořepa, F. Musil, J. Karlíčková, L. Javorská, K. Matoušová, L. K. Krčmová, A. Carazo, A. Šmahelová, V. Blaha and P. Mladěnka, The Antiplatelet Effect of 4-Methylcatechol in a Real Population Sample and Determination of the Mechanism of Action, Nutrients, 2022, 14, 4798 CrossRef PubMed.
- G. D. Wu, C. Compher, E. Z. Chen, S. A. Smith, R. D. Shah, K. Bittinger, C. Chehoud, L. G. Albenberg, L. Nessel and E. Gilroy, Comparative metabolomics in vegans and omnivores reveal constraints on diet-dependent gut microbiota metabolite production, Gut, 2016, 65, 63–72 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3fo04672a |
| ‡ These authors have contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2024 |