
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceSubphthalocyanine-flipper dyads for selective membrane staining†
José
García-Calvo
 *abc,
Xiao-Xiao
Chen
d,
Naomi
Sakai
d,
Stefan
Matile
d and
Tomás
Torres
*abc
*abc,
Xiao-Xiao
Chen
d,
Naomi
Sakai
d,
Stefan
Matile
d and
Tomás
Torres
*abc
aDepartment of Organic Chemistry, Facultad de Ciencias, Universidad Autónoma de Madrid Cantoblanco, 28049-Madrid, Spain. E-mail: jose.garciac@uam.es
bInstitute for Advanced Research in Chemical Sciences (IAdChem), Universidad Autónoma de Madrid, Campus de Cantoblanco, Madrid 28049, Spain
cIMDEA-Nanociencia, c/Faraday 9, Campus de Cantoblanco, Madrid 28049, Spain
dDepartment of Organic Chemistry, University of Geneva, Geneva, Switzerland
First published on 16th January 2024
Abstract
The design, synthesis and evaluation of a subphthalocyanine-flipper (SubPc-Flipper) amphiphilic dyad is reported. This dyad combines two fluorophores that function in the visible region (420–800 nm) for the simultaneous sensing of both ordered and disordered lipidic membranes. The flipper probes part of the dyad possesses mechanosensitivity, long fluorescence lifetimes (τ = 3.5–5 ns) and selective staining of ordered membranes. On the other hand, subphthalocyanines (SubPc) are short-lifetime (τ = 1–2.5 ns) fluorophores that are insensitive to membrane tension. As a result of a Förster Resonance Energy Transfer (FRET) process, the dyad not only retains the mechanosensitivity of flippers but also demonstrates high selectivity and emission in different kinds of lipidic membranes. The dyad exhibits high emission and sensitivity to membrane tension (Δτ = 3.5 ns) when tested in giant unilamellar vesicles (GUVs) with different membrane orders. Overall, the results of this study represent a significant advancement in the applications of flippers and dyads in mechanobiology.
Introduction
Fluorescent dyads are powerful tools for photoluminescence studies. The presence of two fluorophores in close proximity allows the production of molecules with dual-tunable absorption/emission, enabling the control of fluorescence energy transfer (FRET) processes. As a result, dyads find applications in various fields, including chemical sensors,1 double photon absorption processes2 or super-resolution microscopy (SRM).3 In particular, for biological applications, dyads offer numerous advantages when studying lipidic membranes. For instance, Anderson's group reported the use of a FRET-based dyad for SRM in vesicle membranes4 and, more recently, the same group successfully developed a solvatochromic fluorescent dyad with a preference for the more disordered part of a vesicle membrane, when formed by a mixture of phospholipids.5The objective of this study was to combine fluorescent flipper probes with subphthalocyanines (SubPcs) as an environment-insensitive complement, that is to synthesize and characterize the amphiphilic SubPc-Flipper 1 (Fig. 1). This dyad represents an opportunity to improve the design of membrane selective probes capable of distinguishing changes related parameters such as membrane tension,6 polarity or water content and also retaining a high emission, independently of the membrane structure.
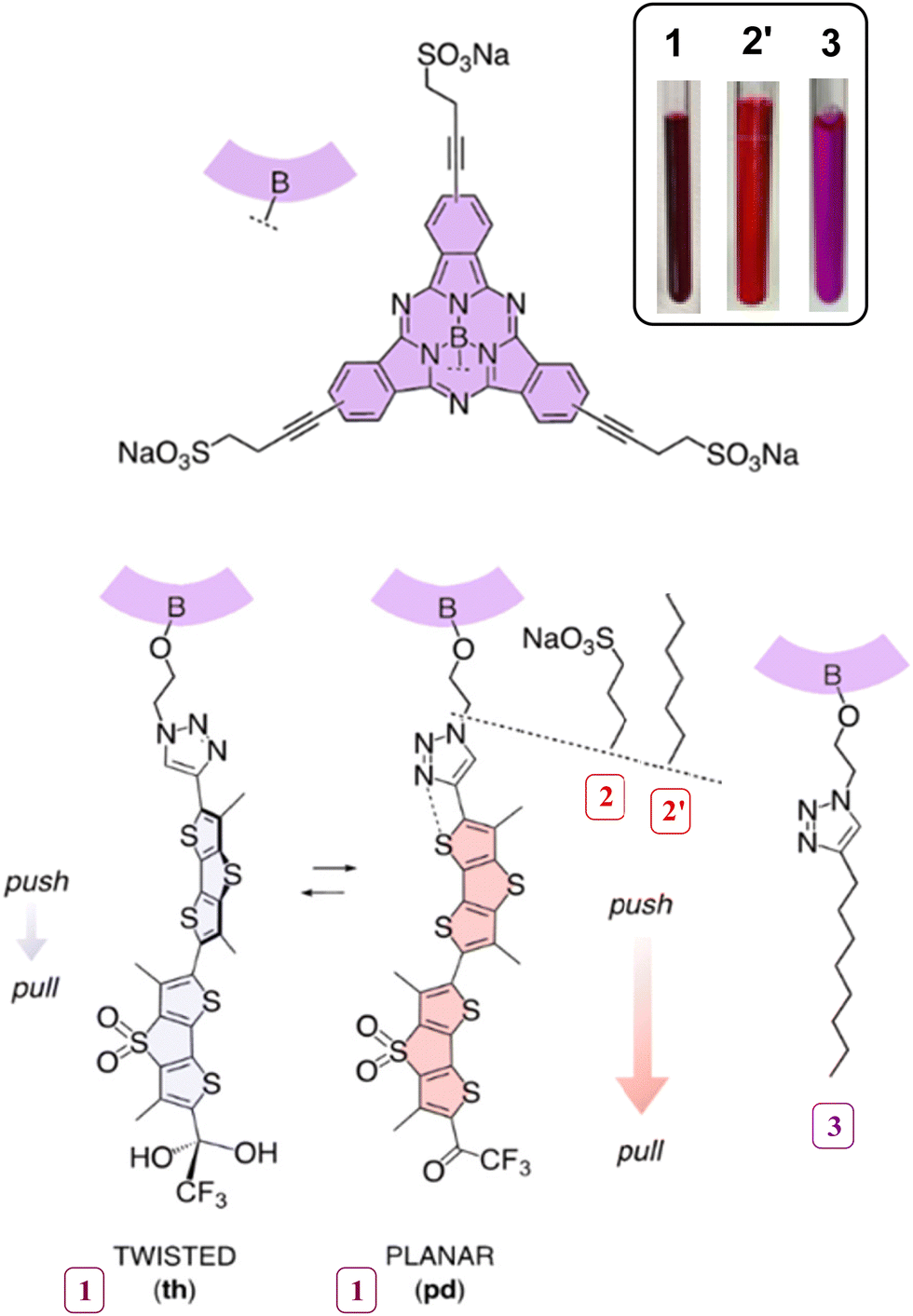 | ||
| Fig. 1 Structure of dyad subphthalocyanine (SubPc) – Flipper 1 and their separate components, namely HydroFlipper 2 and SubPc 3. Both dyad 1 and Hydroflipper 2 are dual-responsive twisted (t) and hydrated (h) probes that upon mechanical planarization (p) and chemical dehydration (d) in membranes of increasing order yield push–pull systems that red shift excitation and increase fluorescence intensity and lifetime. Environment-insensitive phthalocyanines with short lifetime are added to label disordered domains with high contrast and enhance ratiometric lifetime changes in response to tension. The picture in the right-up corner was taken from 1, 2′ and 3 dissolved in dry DMSO. | ||
Fluorescent flippers are dithienothiophene dimers (DTT-DTTO2) that provide the system with a donor/acceptor duality.7 They exhibit a fascinating photochemical behavior involving possible planarization and increased conjugation when in the excited state.8 Planarization only occurs in particular chemical environments and can be tuned by modifying the substituents of the flipper.9,10 This process results in an environment sensitive red-shifted emission; affording operational mechanophores that have been repeatedly applied in chemical biology.6,11 Consequently, derivatized flipper molecules have been utilized to stain and study the properties of different cellular membranes, with a linear correlation between fluorescence lifetimes and forces applied in the membrane.12 In this regard, a series of derivatives have been synthesized, such as the Halo-flipper13 for selective staining of organelles in modified cells, photocleavable-flippers14 for studying the endocytosis pathway or the HydroFlipper10,15 which serves as a starting point for this work.
HydroFlippers represent a new type of fluorescent flipper probes that are sensitive to both water and membrane tension. These compounds are characterized by having a trifluoroketone group, susceptible to hydration, in the terminal acceptor position (compound 2, Fig. 1). Due to this substituent, flippers are not only mechanosensitive but they may hydrate in the presence of water, forming the twisted-hydrate (th, Fig. 1), which loses its fluorescent properties. Up to now, the advantages of the hydration were explored in two different ways, taking advantage of it as a dynamic equilibrium for super-resolution microscopy,10 and using the probe as a dual reporter for the detection and differentiation of changes in membrane order vs. water content.15
Subphthalocyanines (SubPcs) are highly fluorescent molecules obtained from the cyclotrimerization of phthalonitriles in the presence of boron trihalides.16 The scope of these aromatic conical shaped molecules entangles a broad variety of applications for all kind of photophysical17 or supramolecular chemistry18 studies, as well as applications in materials chemistry; such as OLEDs19 or solar cells,20 among many others. Thanks to the high fluorescence quantum yields and the many possible routes for derivatization, they can be also a potent tool in the field of chemical biology. Thus SubPcs have a remarkable performance in optical imaging and for photodynamic therapy.21,22 Synthetically, SubPcs are mostly functionalized throughout two different approaches. On one side, the substituents of the periphery may be altered to change conjugation or add new functions, which modifies intermolecular interactions and photoluminescence properties. On the other hand, the axial substituent on the boron atom does not have a strong influence on the electronical properties, but it is used to introduce new functionalities or to control possible aggregation processes by changing the dipole moment of the molecule. Consequently, in the current case, the dyad was carefully designed to preserve the photoelectronic characteristics by introducing water-solubilizing groups at the periphery and the lipophilic flipper at axial position, which allows for the insertion in lipidic membranes and maintains the amphiphilic structure. Regarding the photophysical properties, SubPcs are particularly attractive because of their high molar extinction coefficients, as well as sharp peaks of absorption (570–580 nm) and emission (585–600 nm), with small Stokes shift, thus short lifetimes.23 Moreover, their absorption and emission bands overlap with the respective bands of HydroFlippers (420–500 nm absorption, 590–750 nm emission), allowing their co-excitation and bidirectional FRET without undesired shifts. Therefore, the combination of SubPcs and HydroFlippers in dyads (such as 1) was of interest to allow internal calibration, since the presence of the probe inside the membrane is clearly demonstrated, which complemented the environment-sensitive probe with varying fluorescence lifetime with an environment-insensitive probe of short lifetime.
The characteristics of SubPcs were of interest to label liquid-disordered (Ld) domains in multicomponent membranes and thus enhance the responsiveness for the following reasons. In fluorescence lifetime imaging microscopy (FLIM) of single-component membranes, fluorescent flippers report increasing membrane order from Ld to liquid-ordered (Lo) and solid-ordered (So) membranes with red shifts in excitation and increasing fluorescence intensity and lifetime (Fig. 2a). In FLIM images of multicomponent membranes, fluorescent flippers report increasing membrane tension as an increase in fluorescent lifetime. This response originates from tension-induced assembly of highly ordered microdomains with long-lived, highly emissive, planarized flippers (Fig. 2b). In reverse, decreasing tension decreases lifetimes because of tension-induced disassembly of microdomains with long-lived flippers (Fig. 2b). This responsiveness to tension changes is limited by the poor fluorescence intensity of short-lived twisted flippers in Ld membranes. Without contrasting emission from Ld membranes, disassembly of ordered microdomains with decreasing tension will be recorded as decreasing intensity, that is decreasing counts in FLIM histograms for long-lived planarized flippers rather than as a decrease in fluorescent lifetime. Although less significant because of overall poor intensity, it can be added that planarization of flippers in Ld domains upon lipid compression by decreasing tension increases rather than decreases lifetimes, which weakens rather than enhances the dominant response of flipper deplanarization from tension-induced domain disassembly. In HydroFlipper 2, the emission from twisted flippers 2th in Ld membranes is further weakened by hydration (Fig. 1). With SubPc-Flipper 1th in Ld membranes, fluorescent properties should thus be dominated by the short-lived, highly-emissive and tension-insensitive SubPc (Fig. 2c, purple). In Lo membranes, the fluorescence properties of SubPc-Flipper 1pd should be dominated by the long-lived, tension-sensitive flipper (Fig. 2c, red).
 | ||
| Fig. 2 (a) Mechanical flipper planarization reports increasing membrane order with red shifts and increasing intensity and lifetime. (b) Responsiveness to tension-induced domain dis/assembly is limited by insufficient short-lifetime emission from Ld domains. (c) SubPc-Flippers add tension-insensitive high-intensity short-lifetime emission to (d) label Ld domains and provide contrast to increasing emission of planar flippers in growing Lo domains with increasing tension. | ||
In multicomponent membranes, the response to high tension with many ordered microdomains should be dominated by the long-lived, tension-sensitive flipper (Fig. 2d, red). While HydroFlipper 2 turns off fluorescence, the response of SubPc-Flipper 1 to low tension with a few ordered domains should be dominated by the short-lived, tension-insensitive SubPc (Fig. 2d, purple). As a result, SubPc-Flipper 1 should label Ld domains at low lifetime with high contrast, which in turn should increase the responsiveness of the small-molecule fluorescent probe to changes in membrane tension.
Results and discussion
Synthesis
With the idea of the amphiphilic SubPc-Flipper 1 in mind, five derivatives were synthesized (compounds 2 and 2′ from Fig. 1 and 1, 3 and 4 from Scheme 1). First, following a reported procedure, HydroFlippers 2 and 2′ were synthesized. On the other hand, the amphiphilic SubPc 3 was also prepared, with an analogous structure to 1 but with an aliphatic octyl-chain instead of the flipper. Finally, a SubPc-flipper soluble in organic solvents (4) was also synthetized. | ||
| Scheme 1 When R = I: (a) 2-hydroxyethyl-1-azide, DBU, toluene, 84%; if R′ = A (b) 11, CuSO4·5H2O, sodium ascorbate, CH2Cl2:THF:H2O, rt, 12 h, 42% and (c) but-3-yne-1-sulfonate, PdCl2(PPh3)2, CuI, Et3N, THF, rt, 48 h, 19%; if R′ = B (b) 12, 1-decyne, CuSO4·5H2O, sodium ascorbate, CH2Cl2:H2O, rt, 2 h, 79%; (c) but-3-yne-1-sulfonate, PdCl2(PPh3)2, CuI, Et3N, THF, rt, 36 h, 27%. When R = H: (a) 2-hydroxyethyl-1-azide, DBU, toluene, 80%; (b) CuSO4·5H2O, sodium ascorbate, CH2Cl2:H2O, rt, 12 h, 29%. | ||
The synthesis involved two convergent routes for flipper and SubPc. For flippers 11, 2 and 2′, the reported route was followed.10 On the other hand, the synthesis of SubPc started with the previously reported triiodo-SubPc 5.24 The first step involved the reaction of the chloride in the axial position with 2-hydroxyethyl-1-azide through nucleophilic substitution in toluene and in the presence of base, resulting in the formation of SubPc 6.25 After purification and characterization, compound 6 underwent click chemistry conditions using CuSO4 and sodium ascorbate in dichloromethane. Flipper 11 was derivatized into 7 while with 1-decyne 12, SubPc 8 was obtained. Next, to make compounds 7 and 8 amphiphilic, 4-sufonate-but-1-yne was introduced throughout a triple Sonogashira coupling at the periphery of the SubPc, resulting in the formation of compounds 1 and 3, respectively. Additionally, a non-amphiphilic derivative, the dyad SubPc-Flipper 4, was also synthesized. To obtain dyad 4, a synthetically equivalent route was followed, where the azide derivative 10 was first synthesized and then, through click chemistry, dyad 4 was obtained. For further details about the synthesis and characterization, please refer to Section 2 of the ESI.†
Spectroscopical properties in solution
First, it must be taken into account that for SubPc 3 there was no significant changes in absorption or fluorescence depending on the solvent, but that was not the case for dyad 1. As it was reported for Flipper 2, and explained in the introduction, dyad 1 showed a duality between the planarized ketone (pd) and the twisted hydrate (th) (see Fig. 1). The equilibrium (pd-th) results in a blue shifted absorption/emission whenever in the hydrated form. However, amphiphiles such as 1 and 3 possess low solubility in aprotic or low-polarity solvents. Therefore, and being limited to the solubility of the amphiphiles, the photoluminescence of 1 in solution was mostly associated to the hydrate 1th (or hemiacetal in methanol solutions). For that reason, the results are also compared to the corresponding non-amphiphilic equivalents of the SubPc-Flipper 1 (4) and HydroFlipper 2 (2′); which provided a better understanding of the dyad behaviour, not only in solution but also in the following studies in phospholipid membranes (vide infra).The amphiphilic control 3 in methanol showed a sharp absorption at 577 nm (Q-band) and an emission peak at 590 nm (Fig. 3a and b, pink). Additionally, changing solvents had no significant impact on the Stokes shifts (Fig. S2b, ESI†), as it is generally reported for SubPcs.26 Conversely, the equilibria in HydroFlipper 2′pd (dioxane) to 2′th (water:dioxane, 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :25 water) produced a blue shift in the absorption band from 446 nm to 400 nm and in fluorescent emission from 619 to 537 nm (Fig. 3a and b, red). In presence of protic solvents, such as methanol or water, 1 and 4 presented an absorption band around 400 nm, associated to the forms 1th and 4th (Fig. 3a, blue and plain purple), confirming the same behaviour than for their HydroFlipper equivalents (2 and 2′). Still, in aprotic solvents, such as chloroform, dyad 4 (Fig. 3a, dashed purple) showed red shifted flipper absorption/emission (4pd), as it was the case for 2′ (Fig. 3a, dashed red). Besides, molar extinctions coefficients (ε) for dyads 1 (λ578, MeOH, 53 ± 3 mM−1 cm−1) and 4 (λ561 (CHCl3) = 63 ± 3 mM−1 cm−1, λ582 (MeOH) = 56 ± 3 mM−1 cm−1), were also matching to a theoretical sum of the components, SubPc 3 (λ577, MeOH, 61 ± 4 mM−1 cm−1) and Flipper 2′ (λ446, dioxane, 34 ± 2 mM−1 cm−1, λ399, water/dioxane, 30 ± 2 mM−1 cm−1) or 2 (λ399, MeOH, 36 ± 4 mM−1 cm−1).
:25 water) produced a blue shift in the absorption band from 446 nm to 400 nm and in fluorescent emission from 619 to 537 nm (Fig. 3a and b, red). In presence of protic solvents, such as methanol or water, 1 and 4 presented an absorption band around 400 nm, associated to the forms 1th and 4th (Fig. 3a, blue and plain purple), confirming the same behaviour than for their HydroFlipper equivalents (2 and 2′). Still, in aprotic solvents, such as chloroform, dyad 4 (Fig. 3a, dashed purple) showed red shifted flipper absorption/emission (4pd), as it was the case for 2′ (Fig. 3a, dashed red). Besides, molar extinctions coefficients (ε) for dyads 1 (λ578, MeOH, 53 ± 3 mM−1 cm−1) and 4 (λ561 (CHCl3) = 63 ± 3 mM−1 cm−1, λ582 (MeOH) = 56 ± 3 mM−1 cm−1), were also matching to a theoretical sum of the components, SubPc 3 (λ577, MeOH, 61 ± 4 mM−1 cm−1) and Flipper 2′ (λ446, dioxane, 34 ± 2 mM−1 cm−1, λ399, water/dioxane, 30 ± 2 mM−1 cm−1) or 2 (λ399, MeOH, 36 ± 4 mM−1 cm−1).
 | ||
| Fig. 3 (a) ε vs. λ and (b) normalized emission spectra of 1 (plain blue, CH3OH), 2 (plain green CH3OH), 2′ (dashed red, dioxane; plain red water/dioxane 1:25), 3 (pink, CH3OH), 4 (dashed purple, CHCl3; plain purple, CH3OH). (c) Emission intensity profile of 4 in dioxane after adding 1.5% of water (30 min, red to blue); for more details see Section S5 of the ESI.† | ||
Furthermore, 4 exhibited an environment dependent FRET response (see Fig. 3b, purple; and Fig. S4 and S5 from the ESI†). In aprotic solvents, two emission bands were observed, corresponding to the SubPc (≈570 nm) and HydroFlipper (600–700 nm), partially overlapping and showing a FRET from the HydroFlipper probe to the SubPc. Here, low fluorescence quantum yields (ϕF) were found for dyad 1th (2%, CH3OH) and 4th (2%, diox:water), which contrasted with dyad 4 in aprotic solutions (4pd, CHCl3, 17%) and SubPc 3 (14%, MeOH). As it was reported before, the equilibrium (pd-th) produced a decrease in the fluorescent quantum yield (ϕF) from 2′pd from 27% (dioxane) to 2′th 2% (water:dioxane, 4% water). Although a lower fluorescent emission was expected for 1th and 4th, the remaining SubPc emission went below the reported for the control 3, whose value is typical for other reported SubPcs.16 The lower emission of 1th and 4th may be explained by the orientation of the dipoles in protic solvents that produces a quenching by FRET from the SubPc to the low-emitting hydrated flippers. The fluorescence quenching is most noticeable when adding water to aprotic solutions (Fig. 3c) or in studies in membranes (vide infra) agreeing with the proposed explanation and results from other fluorescent dyads.5
Time dependent hydration studies were also performed, demonstrating the change from ketone form 4pd to the hydrate 4th. Dyad 4 showed a decrease in emission when water is added (Fig. 3c). It came together with a disappearance of the emission coming from the flipper and takes place in less than 30 min after adding 1.5% of water to a dioxane solution at room temperature. This is further corroborated by a small increase in the emission at 520 nm, with much lower intensity, and associated to the emission of DTT components of the flipper separately (see Fig. S6 for a comparison of the change in fluorescence at 520/620 nm, ESI†).
Fluorescence spectroscopy with LUVs
In order to compare the response to membrane tension, similar vesicle experiments to previous publications were performed.8 The amphiphilic SubPc control 3 showed the characteristic spectra with a sharp excitation maximum at 582 nm and a nearly mirror imaged emission maximizing at 592 nm (Fig. 4a). Both excitation and emission spectra were insensitive to the environment of the SubPc fluorophore. Spectra of 3 in buffer without and with LUVs composed of Ld DOPC (1,2-dioleoyl-sn-glycero-3-phosphocholine) and Lo SM/CL (egg sphingomyelin/cholesterol) 7:3 were nearly superimposable.
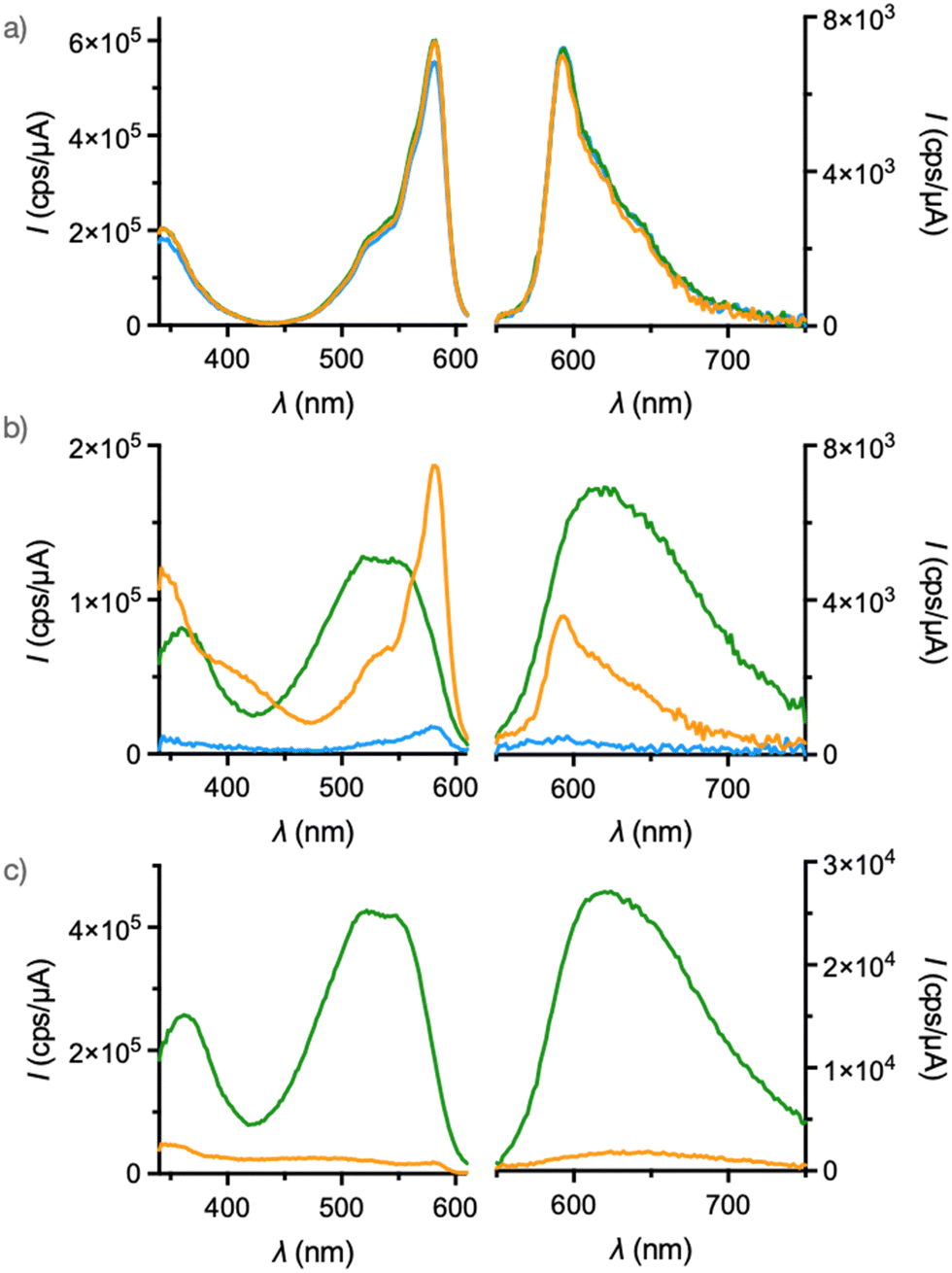 | ||
| Fig. 4 Excitation (left, λem = 650 nm) and emission spectra (right, λex = 480 nm) of 3 (a, 150 nM), 1 (b, 120 nM), and 2 (c, 100 nM) in SM/CL (7:3, green) or DOPC LUVs (gold), and buffer (blue) at 25 °C. | ||
The fluorescence spectra of HydroFlipper 2 were as reported previously.10,15 Namely, fluorescence was completely quenched in buffer, possibly due to self-assembly of the amphiphiles into micelles. Also in Ld LUVs, fluorescence was negligible because twisting and hydration into 2th both decrease fluorescence intensity (Fig. 4c, gold). In Lo LUVs, 2 was highly fluorescent. The red-shifted excitation band showed vibrational finestructure with a 0–0 transition at 560 nm, and the emission maximum at 624 nm (Fig. 4c, green). Both red shift and increasing intensity were consistent with dehydration and mechanical compression of twisted 2th into planar conformer 2pd (Fig. 1 and 2).
SubPc-Flipper 1 in water was essentially not fluorescent. This contrast to SubPc-control 3 could imply FRET from SubPc to the non-emissive flippers under these conditions, or direct self-quenching of self-assembled SubPc in micelles (Fig. 4b, blue). The spectra of 1 in Ld LUVs were dominated by contributions from SubPc (Fig. 4b, gold). The absence of flippers’ contribution in the emission spectra could be rationalized by its very weak emission in Ld membranes. On the other hand, the excitation spectra contained contributions from flippers, indicating the efficient FRET from flippers to SubPc. Compared to SubPc-control 3, the emission of 1 was weaker, probably due to FRET from SubPc to the less fluorescent flippers. However, SubPc quenching was much less pronounced in Ld LUVs than in buffer, suggesting that the two fluorophores are well separated and orthogonally oriented also in Ld LUVs. Such nearly perpendicular orientation of transition dipoles should result in less efficient FRET. This interpretation was consistent with the results obtained in solution and with fluorescence anisotropy in GUVs, described later on.
In sharp contrast, the spectra of 1 in Lo LUVs were dominated by the known characteristics of planarized flippers 2pd, with wide, slightly fine-structured excitation maxima from 500–550 nm, and broad emission maxima from 600–650 nm (Fig. 4b, green). Within these broad bands, the sharp SubPc maxima were almost invisible, only as a bathochromic shoulder of the excitation band (Fig. S8, ESI†). The identically shaped emission spectra were also obtained upon excitation at 560 nm where the absorbance of SubPc is higher, indicating a significant FRET from SubPc to planarized flippers (Fig. S7, ESI†). Fluorescence was overall weaker than 2 in Lo membrane, due probably to flippers to SubPc FRET and/or the inner filter effect.
Taken together, fluorescence spectroscopy of 1 in LUVs reported on Ld and Lo membranes with similarly high intensities and distinct signatures (Fig. 4b, green vs. gold). With SubPc-free flippers like 2, such iso-emissive labeling of membranes of different order is not possible because of insufficient intensity at high disorder (Fig. 4c, green vs. gold).
Fluorescence lifetime imaging microscopy with GUVs
FLIM images of SubPc-Flipper 1 in multi-component GUVs showed labeling of Ld and Lo domains with high intensity and high contrast (Fig. 5d). The colorful images were in stark contrast to the FLIM images of HydroFlipper 2, which labeled Lo domains almost exclusively (Fig. 5g). | ||
| Fig. 5 FLIM images of 3 (a)–(c), 1 (d)–(f) and 2 (g) in GUVs composed of DOPC/SM/CL 3:2:1 (a), (d) and (g), SM/CL 7:3 (b), (e) and DOPC (c) and (f) with enlarged parts of (d) to indicate orthogonal anisotropy (h, i, eq = equatorial, ax = axial). Scale bars = 10 μm, time bar = 0–6 ns, laser at 530 nm. | ||
The average lifetime τav = 4.9 ns of 1 in Lo domains was as long as for other flippers in planar conformation (Table 1). Nevertheless, the average lifetime τav = 1.3 ns in the bright blue Ld domains was exceptionally short (Fig. 5d, f and Table 1). It was much shorter than τav = 1.9 ns for control 3 (Fig. 5a and Table 1). This additional lifetime shortening was likely to originate from FRET from SubPc to flippers, as already observed in LUVs (Fig. 4b).
| Pc | τ L d | τ L o | ΔτLd-Lof | τ mix | τ iso |
|---|---|---|---|---|---|
|
a Fitting uncertainty ± 0.2 ns.
b Intensity averaged lifetimes (τav) in ns.
c Probes (Fig. 1).
d Lifetime τav in FLIM images of single-component Ld GUVs (DOPC)/Ld domains (τ2) of multicomponent GUVs.
e Lifetime τav in FLIM images of single-component Lo GUVs (SM/CL 7:3)/Lo domains (τ3) of multicomponent GUVs.
f Lifetime difference between FLIM images of single-component Lo and Ld GUVs/domains in multicomponent GUVs.
g Lifetime τav in FLIM images of multi-component GUVs with Lo and Ld domains (DOPC/SM/CL 3:2:1).
h Lifetime τav of the plasma membrane in FLIM images of HK cells.
i Data are from ref. 8 except for those of multicomponent GUVs.
|
|||||
| 1 | 1.4/1.3 | 4.9/4.5 | 3.5/3.2 | 1.4 | 3.7 |
| 2 | 2.1/2.2 | 4.7/4.7 | 2.6/2.5 | 4.0 | 4.6 |
| 3 | 1.9/1.9 | — | — | 1.9 | — |
The lifetime τav = 1.3 ns of 1 in the bright blue Ld domains in multicomponent GUVs was also much shorter than the τav = 2.2 ns of pale green Ld domains in multicomponent GUVs labeled with HydroFlipper 2 (Fig. 4d and g and Table 1). The difference ΔτLd-Lo = 2.5 ns between Lo and Ld domains labeled with 2 increased to a record ΔτLd-Lo = 3.2 ns for 1. At comparable intensity, this difference ΔτLd-Lo should correlate directly with responsiveness to changes in membrane tension.
The total lifetime τmix = 4.0 ns of HydroFlipper 2 in multicomponent GUVs was near the τLo = 4.7 ns of the Lo domains (Table 1). This similarity was as expected from the poor fluorescence of twisted 2th in Ld domains (Fig. 5g). In contrast, the total lifetime τmix = 1.4 ns of 1 in multicomponent GUVs was nearer to the τLd = 1.3 ns in Ld than the τLo = 4.5 ns in Lo domains (Table 1). The total lifetime obviously depends on the partitioning preference of the probes in different domains, and their areas, which are different from measurement to measurement. Nevertheless, compared to 2 much shorter τmix = 1.4 ns is unattainable without high intensity and short lifetime of SubPc enhancers added in the Ld domains (Fig. 4d). Taken together, the environment-insensitive SubPc enhancer in dyad 1 labels Ld domains in multicomponent membranes with high intensity and short lifetime, which lowers the total lifetime in multicomponent GUVs from τmix = 4.0 ns to τmix = 1.4 ns and increases the lifetime difference between Ld and Lo domains from ΔτLd-Lo = 2.5 to ΔτLd-Lo = 3.2 ns. These are significant differences that should correlate with the responsiveness of membrane tension in living cells. However, these measurements were not possible because in HeLa Kyoto (HK) cells, SubPc-Flipper 1 was internalized partially under isoosmotic and fully under hyperosmotic conditions (Fig. S11, ESI†). However, reduced lifetimes τiso = 3.7 ns of 1 in the plasma membrane only of HK cells compared to τiso = 4.6 ns of 2 were consistent with increased contributions from tension-insensitive enhancers in Ld domains and thus increased responsiveness to changes in membrane tension (Table 1). We also note that molecular design strategies to prevent probe internalization exist.
In single-component Lo GUVs, FLIM images of SubPc-Flipper 1 showed strong anisotropy with bright equatorial and dark axial regions (Fig. 5e). This anisotropy originates from the selective excitation of fluorophores oriented parallel to the polarized light, is characteristic for flipper probes in ordered membranes and consistent with tight alignment of the probes along the uniformly oriented tails of one leaflet.
In single-component Ld GUVs, FLIM images of 1 showed orthogonal anisotropy with brighter axial than equatorial regions (Fig. 5f). Originating from short-lived SubPc emission, this orthogonal anisotropy suggested that the orientation of SubPc enhancers and flippers is perpendicular to each other. With the hydrophobic flippers oriented along the lipid tails, orthogonal anisotropy aligns the trianionic SubPcs parallel with the membrane water interface (Fig. 2). The detectability of this orthogonal SubPc anisotropy was remarkable considering that in disordered membranes, flipper alignment along less constrained lipid tails is insufficient to produce significant anisotropy. It suggested that the three peripheral sulfonate anions probably all ion pair with ammonium cations in the plane of the membrane surface to ensure sufficient alignment.
In multicomponent GUVs, the anisotropy of planarized flippers in ordered domains was preserved (Fig. 5h). The failure of SubPc to color the dark axial regions in blue was consistent with efficient SubPc quenching by intramolecular FRET to flippers under these conditions, as observed already during spectroscopic analysis in LUVs or in solution. The orthogonal anisotropy of SubPc in disordered domains was also preserved in multicomponent GUVs (Fig. 5i). However, the equator regions were not darkened but colored pale green (Fig. 5i). This small increase in lifetime indicated that emission from twisted flippers compensates for orthogonal SubPc anisotropy. This behavior in Ld domains (Fig. 5i) was contrary to lacking anisotropy compensation in single-component Ld membranes (Fig. 4i) and Lo domains (Fig. 4h). It suggested that in multicomponent GUVs, flipper quenching by FRET to SubPc in Ld domains is less effective than SubPc quenching by FRET to flippers in Ld as well as Lo domains.
Conclusions
The combination of SubPc-flipper was successfully achieved, developing a new tool for staining multicomponent lipidic membranes. The photoluminescence properties of SubPc-Flipper were studied in solution, vesicle membranes, and HK cells. It has been demonstrated that in highly ordered membranes (Lo), the flipper's long fluorescence lifetimes are detected, while in highly disordered membranes (Ld), the SubPc's fluorescence lifetime is predominant. This selective labeling offers a better understanding of the partition between ordered and disordered phases in lipidic membranes.This advancement represents progress in employing dyads for fluorescence staining of membranes. It complements information from each individual reporter and has potential applications for improved selective cell membrane staining. Therefore, it is demonstrated that the use of multicomponent probes may lead to a deeper understanding of membranes providing highly attractive possibilities for chemical biology.
Author contributions
J. G-C.: conceptualization, synthesis, characterization, measurements in solution, interpretation of results and MS + SI writing. X-X. C.: vesicle and cell experiments, data treatment and interpretation and SI writing. N. S.: vesicle and cell experiments, data treatment and interpretation, MS and SI writing. S. M.: conceptualization, interpretation, resources and MS writing. T. T. conceptualization, resources, supervision and MS writing.Conflicts of interest
The University of Geneva has licensed Flipper-TR® probes to Spirochrome for commercialization.Acknowledgements
We thank the NMR, MS and Bioimaging platforms for services, and the University of Geneva, the National Centre of Competence in Research (NCCR) Chemical Biology (51NF40-185898), the NCCR Molecular Systems Engineering (51NF40-182895), as well as the Swiss NSF for financial support (Excellence Grant 200020 204175; Swiss-ERC Advanced Grant TIMEUP, TMAG-2_209190). J. G. C. acknowledges his funding from “Ayudas María Zambrano para la atracción de talento internacional” (CA3/RSUE/2021-00243) from the Spanish Government/EU funds at Universidad Autónoma de Madrid, and “Ayudas de atracción de talento de la Comunidad de Madrid 2022” (2022-T1/IND-23907) supported by Comunidad de Madrid with EU funds at IMDEA Nanociencia. T. T. acknowledges financial support from the Spanish AEI/MICINN (PID2020-116490GB-I00) and the Comunidad de Madrid and the Spanish State through the Recovery, Transformation and Resilience Plan [“Materiales Disruptivos Bidimensionales (2D)” (MAD2D-CM) (UAM1)-MRR Materiales Avanzados], and the European Union through the Next Generation EU funds. IMDEA Nanociencia acknowledges support from the “Severo Ochoa” Programme for Centres of Excellence in R&D (MINECO, Grant SEV2016-0686).Notes and references
- A. Vanessa Saura, M. Isabel Burguete, F. Galindo and S. V. Luis, Org. Biomol. Chem., 2017, 15, 3013–3024 RSC.
- D. Collado, P. Remón, Y. Vida, F. Najera, P. Sen, U. Pischel and E. Perez-Inestrosa, Chem. – Asian J., 2014, 9, 797–804 CrossRef CAS PubMed.
- E. Deniz, S. Ray, M. Tomasulo, S. Impellizzeri, S. Sortino and F. M. Raymo, J. Phys. Chem. A, 2010, 114, 11567–11575 CrossRef CAS PubMed.
- A. T. Frawley, V. Wycisk, Y. Xiong, S. Galiani, E. Sezgin, I. Urbančič, A. Vargas Jentzsch, K. G. Leslie, C. Eggeling and H. L. Anderson, Chem. Sci., 2020, 11, 8955–8960 RSC.
- A. T. Frawley, K. G. Leslie, V. Wycisk, S. Galiani, D. Shrestha, C. Eggeling and H. L. Anderson, Chem. Phys. Chem., 2023, 24, e202300125 CrossRef CAS PubMed.
- A. Colom, E. Derivery, S. Soleimanpour, C. Tomba, M. D. Molin, N. Sakai, M. González-Gaitán, S. Matile and A. Roux, Nat. Chem., 2018, 10, 1118–1125 CrossRef CAS PubMed.
- J. García-Calvo, J. López-Andarias, N. Sakai and S. Matile, Chem. Commun., 2021, 57, 3913–3916 RSC.
- M. Dal Molin, Q. Verolet, A. Colom, R. Letrun, E. Derivery, M. Gonzalez-Gaitan, E. Vauthey, A. Roux, N. Sakai and S. Matile, J. Am. Chem. Soc., 2015, 137, 568–571 CrossRef CAS PubMed.
- J. García-Calvo, J. López-Andarias, N. Sakai and S. Matile, Helv. Chim. Acta, 2022, 105, e202100238 CrossRef.
- J. García-Calvo, J. Maillard, I. Fureraj, K. Strakova, A. Colom, V. Mercier, A. Roux, E. Vauthey, N. Sakai, A. Fürstenberg and S. Matile, J. Am. Chem. Soc., 2020, 142, 12034–12038 CrossRef PubMed.
- C. Roffay, G. Molinard, K. Kim, M. Urbanska, V. Andrade, V. Barbarasa, P. Nowak, V. Mercier, J. García-Calvo, S. Matile, R. Loewith, A. Echard, J. Guck, M. Lenz and A. Roux, Proc. Natl. Acad. Sci. U. S. A., 2021, 118, e2103228118 CrossRef CAS PubMed.
- A. Goujon, A. Colom, K. Straková, V. Mercier, D. Mahecic, S. Manley, N. Sakai, A. Roux and S. Matile, J. Am. Chem. Soc., 2019, 141, 3380–3384 CrossRef CAS PubMed.
- K. Straková, J. López-Andarias, N. Jiménez-Rojo, J. E. Chambers, S. J. Marciniak, H. Riezman, N. Sakai and S. Matile, ACS Cent. Sci., 2020, 6, 1376–1385 CrossRef PubMed.
- J. López-Andarias, K. Straková, R. Martinent, N. Jiménez-Rojo, H. Riezman, N. Sakai and S. Matile, JACS Au, 2021, 1, 221–232 CrossRef PubMed.
- J. García-Calvo, J. López-Andarias, J. Maillard, V. Mercier, C. Roffay, A. Roux, A. Fürstenberg, N. Sakai and S. Matile, Chem. Sci., 2022, 13, 2086–2093 RSC.
- G. Lavarda, J. Labella, M. V. Martínez-Díaz, M. S. Rodríguez-Morgade, A. Osuka and T. Torres, Chem. Soc. Rev., 2022, 51, 9482–9619 RSC.
- H. Gotfredsen, D. Thiel, P. M. Greißel, L. Chen, M. Krug, I. Papadopoulos, M. J. Ferguson, M. B. Nielsen, T. Torres, T. Clark, D. M. Guldi and R. R. Tykwinski, J. Am. Chem. Soc., 2023, 145, 9548–9563 CrossRef CAS PubMed.
- J. Labella, G. Lavarda, L. Hernández-López, F. Aguilar-Galindo, S. Díaz-Tendero, J. Lobo-Checa and T. Torres, J. Am. Chem. Soc., 2022, 144, 16579–16587 CrossRef CAS PubMed.
- N. F. Farac, A. R. Tetreault and T. P. Bender, J. Phys. Chem. C, 2023, 127, 702–727 CrossRef CAS.
- H. Gotfredsen, T. Neumann, F. E. Storm, A. V. Muñoz, M. Jevric, O. Hammerich, K. V. Mikkelsen, M. Freitag, G. Boschloo and M. B. Nielsen, ChemPhotoChem, 2018, 2, 976–985 CrossRef CAS.
- N. M. Casellas, G. Dai, E. Y. Xue, A. Fonseca, D. K. P. Ng, M. García-Iglesias and T. Torres, Chem. Commun., 2022, 58, 669–672 RSC.
- E. Winckel, M. Mascaraque, A. Zamarrón, Á. Juarranz De La Fuente, T. Torres and A. Escosura, Adv. Funct. Mater., 2018, 28, 1705938 CrossRef.
- I. D. Burtsev, T. V. Dubinina, A. E. Egorov, A. A. Kostyukov, A. V. Shibaeva, A. S. Agranat, M. M. Ivanova, L. R. Sizov, N. V. Filatova, A. Y. Rybkin, E. V. Varakina, A. S. Bunev, A. A. Antonets, E. R. Milaeva and V. A. Kuzmin, Dyes Pigm., 2022, 207, 110690 CrossRef CAS.
- D. González-Rodríguez, T. Torres, M. Á. Herranz, L. Echegoyen, E. Carbonell and D. M. Guldi, Chem. – Eur. J., 2008, 14, 7670–7679 CrossRef PubMed.
- Ö. Göktuğ, C. Göl and M. Durmuş, J. Porphyrins phthalocyanines, 2017, 21, 539–546 CrossRef.
- F. Ma, G. Zhang, X. Wu, W. Qu, Z. Sun, X. Li and X. Yin, J. Phys. Chem. C, 2022, 126, 13496–13504 CrossRef CAS.
Footnote |
| † Electronic supplementary information (ESI) available: Detailed experiments, procedures and results are reported. See DOI: https://doi.org/10.1039/d3cp05476d |
| This journal is © the Owner Societies 2024 |