
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Target 2035 – an update on private sector contributions
Suzanne
Ackloo
 *a,
Albert A.
Antolin
b,
Jose Manuel
Bartolome
c,
Hartmut
Beck
d,
Alex
Bullock
e,
Ulrich A. K.
Betz
f,
Jark
Böttcher
g,
Peter J.
Brown
h,
Menorca
Chaturvedi
i,
Alisa
Crisp
j,
Danette
Daniels
k,
Jan
Dreher
d,
Kristina
Edfeldt
l,
Aled M.
Edwards
a,
Ursula
Egner
m,
Jon
Elkins
e,
Christian
Fischer
n,
Tine
Glendorf
o,
Steven
Goldberg
p,
Ingo V.
Hartung
q,
Alexander
Hillisch
d,
Evert
Homan
r,
Stefan
Knapp
st,
Markus
Köster
i,
Oliver
Krämer
i,
Josep
Llaveria
c,
Uta
Lessel
u,
Sven
Lindemann
f,
Lars
Linderoth
o,
Hisanori
Matsui
v,
Maurice
Michel
r,
Florian
Montel
w,
Anke
Mueller-Fahrnow
m,
Susanne
Müller
st,
Dafydd R.
Owen
x,
Kumar Singh
Saikatendu
y,
Vijayaratnam
Santhakumar
a,
Wendy
Sanderson
z,
Cora
Scholten
aa,
Matthieu
Schapira
aab,
Sujata
Sharma
p,
Brock
Shireman
p,
Michael
Sundström
l,
Matthew H.
Todd
ac,
Claudia
Tredup
st,
Jennifer
Venable
p,
Timothy M.
Willson
ad and
Cheryl H.
Arrowsmith
*aae
*a,
Albert A.
Antolin
b,
Jose Manuel
Bartolome
c,
Hartmut
Beck
d,
Alex
Bullock
e,
Ulrich A. K.
Betz
f,
Jark
Böttcher
g,
Peter J.
Brown
h,
Menorca
Chaturvedi
i,
Alisa
Crisp
j,
Danette
Daniels
k,
Jan
Dreher
d,
Kristina
Edfeldt
l,
Aled M.
Edwards
a,
Ursula
Egner
m,
Jon
Elkins
e,
Christian
Fischer
n,
Tine
Glendorf
o,
Steven
Goldberg
p,
Ingo V.
Hartung
q,
Alexander
Hillisch
d,
Evert
Homan
r,
Stefan
Knapp
st,
Markus
Köster
i,
Oliver
Krämer
i,
Josep
Llaveria
c,
Uta
Lessel
u,
Sven
Lindemann
f,
Lars
Linderoth
o,
Hisanori
Matsui
v,
Maurice
Michel
r,
Florian
Montel
w,
Anke
Mueller-Fahrnow
m,
Susanne
Müller
st,
Dafydd R.
Owen
x,
Kumar Singh
Saikatendu
y,
Vijayaratnam
Santhakumar
a,
Wendy
Sanderson
z,
Cora
Scholten
aa,
Matthieu
Schapira
aab,
Sujata
Sharma
p,
Brock
Shireman
p,
Michael
Sundström
l,
Matthew H.
Todd
ac,
Claudia
Tredup
st,
Jennifer
Venable
p,
Timothy M.
Willson
ad and
Cheryl H.
Arrowsmith
*aae
aStructural Genomics Consortium, University of Toronto, Toronto, Ontario M5G 1L7, Canada. E-mail: suzanne.ackloo@utoronto.ca; aled.edwards@utoronto.ca; Cheryl.Arrowsmith@uhnresearch.ca
bProCURE, Catalan Institute of Oncology, Oncobell, Bellvitge Institute for Biomedical Research (IDIBELL), L'Hospitalet del Llobregat, Barcelona, Catalonia, Spain. E-mail: aantolin@idibell.cat
cA Division of Janssen-Cilag S.A., Janssen Research and Development, Toledo, Spain. E-mail: jbartolo@its.jnj.com; JLlaveri@ITS.JNJ.com
dResearch and Development, Bayer AG, Pharmaceuticals, 42103 Wuppertal, Germany. E-mail: hartmut.beck@bayer.com; jan.dreher@bayer.com; alexander.hillisch@bayer.com
eCenter for Medicines Discovery, Old Road Campus, University of Oxford, Roosevelt Drive, Headington, Oxford, OX3 7DQ, UK. E-mail: alex.bullock@cmd.ox.ac.uk; jon.elkins@cmd.ox.ac.uk
fMerck KGaA, Darmstadt, Germany. E-mail: ulrich.betz@merckgroup.com; Sven.Lindemann@merckgroup.com
gBoehringer Ingelheim RCV GmbH & Co KG, Vienna, Austria. E-mail: jark.boettcher@boehringer-ingelheim.com
hStructural Genomics Consortium, University of North Carolina at Chapel Hill, USA. E-mail: peter.brown@unc.edu
iBoehringer Ingelheim International, Binger Str. 173, D-55216 Ingelheim, Germany. E-mail: menorca.chaturvedi@boehringer-ingelheim.com; markus.koester@boehringer-ingelheim.com; oliver.kraemer@boehringer-ingelheim.com
jDivision of Cancer Therapeutics, The Institute of Cancer Research, London, SM2 5NG, UK. E-mail: Alisa.crisp@icr.ac.uk
kFoghorn Therapeutics, 500 Technology Square, Suite 700, Cambridge, MA 02139, USA. E-mail: DDaniels@foghorntx.com
lStructural Genomics Consortium, Department of Medicine, Karolinska University Hospital and Karolinska Institutet, Stockholm, Sweden. E-mail: kristina.edfeldt@ki.se; michael.sundstrom@ki.se
mNuvisan Innovation Campus Berlin GmbH, Müllerstraße 178, 13353, Berlin, Germany. E-mail: egner.target2035@web.de; anke.mueller-fahrnow@nuvisan.com
nDiscovery Chemistry, Merck & Co., Inc., Boston, Massachusetts 02115, USA. E-mail: christian_fischer@merck.com
oResearch & Early Development, Novo Nordisk A/S, Måløv, Denmark. E-mail: tgle@novonordisk.com; lrli@novonordisk.com
pJanssen Research and Development LLC, San Diego, California, USA. E-mail: SGoldbe1@its.jnj.com; ssharm505@ITS.JNJ.com; BShirema@its.jnj.com; JVenable@its.jnj.com
qMedicinal Chemistry, Global R&D, Merck Healthcare KGaA, Frankfurter Straße 250, 64293, Darmstadt, Germany. E-mail: ingo.hartung@merckgroup.com
rScience for Life Laboratory, Department of Oncology-Pathology, Karolinska Institutet, Stockholm, Sweden. E-mail: evert.homan@ki.se; maurice.grube@scilifelab.se
sInstitute of Pharmaceutical Chemistry, Goethe University Frankfurt, Frankfurt 60438, Germany. E-mail: knapp@pharmchem.uni-frankfurt.de; tredup@pharmchem.uni-frankfurt.de
tStructural Genomics Consortium, BMLS, Goethe University Frankfurt, Frankfurt 60438, Germany. E-mail: susanne.mueller-knapp@bmls.de
uBoehringer Ingelheim Pharma GmbH & Co. KG, Birkendorfer Str. 65, D-88397 Biberach an der Riss, Germany. E-mail: uta.lessel@boehringer-ingelheim.com
vNeuroscience Drug Discovery Unit, Research, Takeda Pharmaceutical Company Limited, Fujisawa, Kanagawa, Japan. E-mail: hisanori.matsui@takeda.com
wBoehringer Ingelheim Pharma GmbH & Co. KG, Birkendorfer Str. 65, D-88397 Biberach an der Riss, Germany. E-mail: florian.montel@boehringer-ingelheim.com
xDiscovery Network Group, Pfizer Medicine Design, Cambridge, MA 02139, USA. E-mail: Dafydd.Owen@pfizer.com
yGlobal Research Externalization, Takeda California, Inc., 9625 Towne Center Drive, San Diego, CA 92121, USA. E-mail: kumar.saikatendu@takeda.com
zJanssen Research & Development, Janssen Pharmaceutica N. V, Beerse, Belgium. E-mail: WSANDERS@its.jnj.com
aaResearch and Development, Bayer AG, Pharmaceuticals, 13353 Berlin, Germany. E-mail: cora.scholten@bayer.com
abDepartment of Pharmacology & Toxicology, University of Toronto, Toronto, Ontario M5S 1A8, Canada
acSchool of Pharmacy, University College London, London, WC1N 1AX, UK. E-mail: mattoddchem@gmail.com
adStructural Genomics Consortium, UNC Eshelman School of Pharmacy, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599, USA. E-mail: tim.willson@unc.edu
aePrincess Margaret Cancer Centre, Toronto, Ontario M5G 1L7, Canada
First published on 16th March 2023
Abstract
Target 2035, an international federation of biomedical scientists from the public and private sectors, is leveraging ‘open’ principles to develop a pharmacological tool for every human protein. These tools are important reagents for scientists studying human health and disease and will facilitate the development of new medicines. It is therefore not surprising that pharmaceutical companies are joining Target 2035, contributing both knowledge and reagents to study novel proteins. Here, we present a brief progress update on Target 2035 and highlight some of industry's contributions.
Introduction
Formulated in 2018, by public funders and scientific representatives from academia and the drug discovery industry, Target 2035 is focused on developing a pharmacological tool for each human protein by the year 2035.1–5 This is a significant challenge as illustrated by the current view of the liganded proteome (Fig. 1) especially relative to Antolin et al.'s assessment of the liganded proteome in 2018.6 We are, however, confident that, with open and unrestricted science and a global federated effort from academia and industry, we will meet this challenge.7–14 Target 2035 aims to create an open and collaborative ecosystem through forming public–private partnerships (PPPs), hosting virtual events viahttps://target2035.net,15 open science communities such as Just One Giant Lab,16 and contributing to the organization of in-person workshops such as at the International Chemical Biology Society conference (https://icbs2022.chemical-biology.org/). | ||
| Fig. 1 Current analysis of the global scope of chemical probes. The percentage of the proteome that is predicted to be ligandable has been derived from canSAR;24 it calculates the ligandability of all proteins using 3D structures from either the PDB and/or AlphaFold Protein Structure Database. The liganded proteome has been calculated assuming that a protein is liganded if it has a small molecule in public chemical databases24 with a binding or inhibition activity more potent than 10 micromolar. The human targets that can be currently studied with a chemogenomics library or a chemical probe have been calculated from public databases24 using the potency, selectivity and cell activity thresholds displayed. This analysis does not include data from patents unless they have been curated in ChEMBL25 or BindingDB.26 | ||
The goals of Target 2035 can only be accomplished by expanding hit-finding approaches beyond traditional methods. This is especially true for un/understudied proteins that have no link to disease. Traditional hit-finding methods tend to be prohibitively resource-intensive and limited to large private sector entities. During the past year Target 2035 launched two PPPs, Critical Assessment of Computational Hit-finding Experiments (CACHE),17 and Open Chemistry Networks (OCN).18 CACHE was conceived by a working group of experts from the public and private sector, including leaders of D3R, a previous similar initiative.19 Because CACHE is a prospective hit-finding exercise, where predicted compounds are procured and tested using biophysical and biochemical methods, the metric of success is hit rate, diversity, and drug likeness rather than binding pose or docking score. As such, CACHE is complementary to previous and current excellent computational chemistry benchmarking efforts such as D3R or SAMPL.19,20
In CACHE,17 computational chemists have three unique opportunities per year to benchmark their algorithms and processes. In every challenge, each participant predicts up to 100 ligands for new, expert-curated targets. The predicted compounds are purchased, evaluated experimentally in target-specific assays, and binding data returned to participants. For methods that successfully predicted binders, a second (and final) round of predictions is performed making use of the experimental results from the first round, if possible. Three benchmarking challenges (https://cache-challenge.org/) are currently underway for LRRK2 WD40 repeat domain (WDR),21 SARS-CoV-2 NSP13 RNA-binding domain, and SARS-CoV-2 NSP3 ADP-ribose binding site each with 25 participants.22
OCN is an international, open, and inclusive chemistry network that exists to bring together chemists and biochemists in the community with a common aim of developing probes to understudied targets.23 It has the potential to involve global chemistry contributors, helping to develop small molecule binders for proteins for which there may be no local expertise. Since all data are open and clearly licenced as CC-BY, contributors can use these data in their own applications for follow-on funding. The current targets include RBBP4, HIPK4, PLCZ1, NSP13, and ABHD2 each having unique needs.18 However, all projects are contingent on synthetic chemist trainees, and target champions from the drug discovery industry. The data generated may be used to facilitate funding opportunities for downstream research.
Pharmaceutical companies have collaborated effectively within PPPs, especially in the pre-competitive space8,27,28 and will continue to contribute towards Target 2035 by donating29 and co-developing chemical probes,4,30–34 and other pharmacological tools which can be used by the global research community without restrictions. In addition, industry (outside the umbrella of Target 2035) hosts open innovation programs wherein the global academic research community can access first-in-class compounds, by proposing exciting hypothesis-driven science. Noteworthy examples are the AACR-Bayer Innovation and Discovery Grants,35 Boehringer Ingelheim's OpnMe,7,36,37 Merck KGaA's Open Innovation Portal,38 Co-Create Knowledge for Pharma Innovation with Takeda Funding,39 and Novo Nordisk's Compound Sharing platform.40 In the following section, representatives from the private sector independently added granularity to their company's contributions to open science.
Summaries of private sector contributions to Target 2035
Bayer
Bayer is highly committed to contribute to open science by engaging in joint projects with academia and by donating high-quality chemical probes and chemogenomic libraries.29 Thus far, 28 chemical probes made by Bayer have been contributed to open science during long-term collaborations with Structural Genomics Consortium (SGC) and IMI projects, such as EUbOPEN.41 The majority are donated probes resulting from dedicated in-house lead optimization programs aiming at clinical candidate compounds, which speaks for their quality and relevance for the scientific community. Bayer's first donated probe in 2015 was the human neutrophil elastase (HNE) inhibitor BAY-678, profiled side by side with its close congener the clinical compound BAY 85-8501 for the treatment of pulmonary diseases.42 Bayer's latest donated probe in 2022 is the former clinical candidate compound BAY 1816032, a potent and selective BUB1 inhibitor for cancer https://www.sgc-ffm.uni-frankfurt.de/#!specificprobeoverview/BAY%201816032.Based on the recognition that “in open science everybody wins”, Bayer is increasingly interested to join efforts with academia to evaluate innovative targets. The idea is to combine the best of both worlds, by bringing together industrial lead optimization expertise with specific academic bioassay knowledge, for the benefit of all parties. Bayer's first co-developed chemical probe was the SMYD2 inhibitor BAY-598 for cancer.43 The latest example is the USP21 inhibitor BAY-80544 which is the first potent and selective probe for USP21. It will help researchers gain new insights into the intriguing disease biology of immune-oncology.
Apart from providing chemical probes, Bayer is also engaged in a variety of IMI projects contributing to open science via developing IT tools, databases, and technology platforms. Bayer shares the opinion that the ambitious goal of the Target 2035 initiative, to deliver a chemical probe for every potential target protein, can only be achieved by enhanced use of computational tools. To support this effort, Bayer co-initiated together with the SGC the CACHE17 initiative, to predict small molecules as potential target protein binders and to benchmark and ultimately improve various computational hit-finding methods.
All these manifold tools provided through open science will strongly contribute to deepen the understanding of disease-related biological processes and thus pave the way for the future development of new medicines for patients. Everybody wins.
Boehringer Ingelheim opnMe
Boehringer Ingelheim is consciously committed to continuously foster innovation through open science. To achieve this goal, we are sharing our best tool compounds and beyond via different initiatives: https://www.theSGC.org, https://EUbOPEN.org, and https://opnMe.com.7,36,37 The driver behind our open science initiatives is the recognition that, “No matter who you are, most of the brightest brains work for someone else,” and not only in your own organization. Our open science initiatives seek to lift barriers between academic researchers and high-quality, industrial technology, and, in doing so, chart a new path for pharmaceutical open innovation.Our https://opnMe.com portal is built on three initiatives:
• Molecules to order (M2O): well-characterized pre-clinical molecules delivered free-of-charge and no strings attached. Scientists who order compounds through M2O control all foreground IP rights arising from their research. As of the end of June 2022, there were 74 probe molecules available through M2O. Molecules are actively being added to the web portal at a rate of approximately one per month, with the ultimate goal of providing open access to all high-quality tool compounds.
• Molecules for collaboration (M4C): first-in-class tool compounds provided under collaboration agreements. The main aim of the M4C program is to engage outside researchers to discover new therapeutic concepts for biological targets that have no publicly reported tool compounds. Four projects initiated through M4C have returned compounds to Boehringer Ingelheim's discovery research portfolio. Although we do not reveal the compound structure or current disease areas being pursued internally, each M4C call for proposals provides researchers with both in vitro and in vivo data profiles of the compounds, thereby allowing researchers to immediately identify whether these compounds are appropriate tools for their research projects.
• opn2EXPERTS: precisely formulated questions from Boehringer Ingelheim scientists to trigger innovative science. At the time of publication of this article 20 calls have been opened to the scientific community and 457 proposals from 53 countries have been received. Despite the pandemic this program was successfully launched in 2020 and receive excellent feedbacks from the scientific community. More than 30 collaborations between academic partners or biotechnology companies and Boehringer Ingelheim have emerged from these calls.
As a member of the SGC and co-lead of the EUbOPEN project, Boehringer Ingelheim has so far contributed four co-developed – BRD9/7: BI-9564,45 NSD3: BI-9321,46 BPTF: BI-7190,47 SLC9A1: BI-962748 – and fifteen donated chemical probes.49,50
To support open science projects in earlier stages and to enable anyone interested to contribute to drug design, DrugIt51 – a public domain drug design game – has been created in collaboration with Vanderbilt University. Finally, Boehringer Ingelheim is a full member of the CACHE initiative, which provides unbiased, high-quality experimental data on computational hit-finding predictions with the aim to define state-of-the-art in silico methods for drug design. Through these open science initiatives, we believe that together we can accelerate research initiatives in areas of high unmet medical need.
Foghorn Therapeutics
Foghorn Therapeutics joined the Target 2035 initiative this year to support open science and expand the chemical probe collection with several potent small molecule inhibitors such as FHT-2344 targeting the ATPase domains of SMARCA2 and SMARCA4, also known as BRM and BRG1, respectively.52 BRM and BRG1 are core members of the mammalian SWI/SNF family of chromatin remodeling complexes (also known as BAF complexes), which regulate open and closed chromatin states and control transcriptional activation and repression.53 They are key therapeutic targets as numerous diseases have been linked to mutations or loss of BRM/BRG1 proteins, resulting in improper BAF localization and activity. The dual BRM/BRG1 probe of the ATPase domain efficiently inactivates BAF complexes, allowing for determination of relevant phenotypic outcomes in disease models which have aberrant BAF activity. These probes will not only accelerate development of potential medicines, but also enable the academic community to further understand BAF roles, activity, and important interactions.Foghorn additionally is actively developing heterobifunctional targeted protein degraders to therapeutic targets within the BAF complex or those which regulate chromatin and transcription. The Target 2035 initiative greatly enables the targeted protein degradation field by providing qualified chemical probes which can be used as starting points for development of heterobifunctional degraders. Foghorn seeks to support these efforts by contributing, in the coming years, additional probes which can specifically be applied to targeted protein degradation.
Janssen
There is an urgent need to bring in new validated targets across industry as we are faced with challenges like the recent pandemic or drug resistance in cancers or anti-infectives. Robust validation via high-quality chemical probes is an integral part of Janssen's strategy to bring new targets into the portfolio. This aligns with the Target 2035 mission to develop pharmacological tools for each protein in the proteome. Towards this mission, Janssen has donated (in the last year) six unique chemical probes,54 their negative control compounds and robust data packages for ROR gamma T,55 PDE10a, spermine oxidase, histamine receptor H4,56 CK1 Delta57 and O-GlcNAcase,58 and we remain committed to enable the community with high-quality pharmacological tools. Besides well-characterized probes, Janssen's JumpstARter Library is available to researchers to help “jump-start” hit finding projects, e.g. though our partnership with WIPO Re:Search.59 The library includes a diverse collection of 80![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 high-quality drug-like small molecules and compound fragments. Janssen is also planning to profile SGC's growing library of annotated chemical probes in high content imaging assays to create biosignatures for this library of molecules. Additionally, consortia such as the SGC allows for cross-industry sharing of probes to accelerate go/no go on targets that may be of interest to Janssen and not to other pharma partners.
000 high-quality drug-like small molecules and compound fragments. Janssen is also planning to profile SGC's growing library of annotated chemical probes in high content imaging assays to create biosignatures for this library of molecules. Additionally, consortia such as the SGC allows for cross-industry sharing of probes to accelerate go/no go on targets that may be of interest to Janssen and not to other pharma partners.
MSD
Pharmacological interrogation of the proteome with highly specific chemical probes is an important part of our strategy to support target validation at MSD, which is aligned with the Target 2035 initiative. In recent years we at MSD have developed three unique chemical probes and their negative controls through a collaboration with the SGC: MRK-740 targeting PRDM9,60 MRK-952 targeting NUDT5,61 and MRK-990 targeting PRMT9/5.62 Additionally, we have donated several potent and selective inhibitors, with their respective data packages and negative controls, from our internal collections to the community.29,50 MSD has also contributed spectrum-selective kinase inhibitors to aid the expansion of the PKIS sets.63 For the future we remain committed to promoting the use of high-quality pharmacological tools in the scientific community and will continue to enable the goals of the Target 2035 initiative through our scientific publications and disclosures of novel tool compounds with robust data packages, to support target validation.Merck KGaA Darmstadt
Merck KGaA Darmstadt sees a continuing need for high-quality chemical probes being made accessible to the scientific community as we are concerned about the still high number of target validation studies employing poorly characterized chemical probes. Resulting incorrect conclusions about the link between a target and a disease phenotype led to misallocation of research time and budget into drug discovery projects which likely will never provide benefit to patients. While during the recent decade the principles of high-quality chemical probes have been embraced by a part of the drug discovery community, we strongly believe that further efforts are necessary to reach deeper into the broader cell biology community.Therefore, we are following several approaches to contribute to the Target 2035 mission. We are actively pursuing programs to identify such high-quality chemical probes together with the SGC teams in Frankfurt and Toronto. For example, we recently jointly identified MSC2711186,64 a SRPK1/2/3 inhibitor, which will now allow researchers to explore these understudied members of the kinome. In addition, we are encouraging our internal research teams to publish high-quality chemical probes from internal programs for those targets where we believe that target biology understanding is still limited. Towards this end we recently published MSC-4381,65 a highly selective inhibitor of monocarboxylate transporter 4 (MCT4) with excellent properties for cellular as well as in vivo studies.66 This probe will allow academic labs to delineate contributions of MCT4 to lactate transport for example in the context of metabolic adaptions of tumor cells. Even more recently we disclosed MSC-4106, a valuable in vivo usable inhibitor of the Yap/TEAD interaction.67 This interaction plays a crucial role in the developmental Hippo pathway which is deregulated in certain human tumors. Until now, no well characterized probe to study the role of TEAD transcription factors was available in the public domain. We believe that while some target families are already well served with high-quality chemical probes (e.g. certain epigenetic protein families), there are also protein families which urgently need an orchestrated effort to provide such tools. Examples include DNA/RNA helicases, metallonucleases and E3 ligases.
The Merck open innovation portal offers a series of touchpoints to the scientific community to interact with Merck and to engage in one of the many open innovation offers such as e.g. a training program for students from all over the world (https://www.emdgroup.com/en/research/open-innovation/innovation-cup.html), an opportunity to apply for Merck Research Grants (https://researchgrants.merckgroup.com), or an opportunity to get Merck compounds for research.
Novo Nordisk Compound Sharing
The Novo Nordisk Compound Sharing platform40 was launched in 2020 to provide easy access to high-quality peptide and protein analogues many of which are not readily available to the scientific community. The intention with this open innovation initiative is to accelerate innovation. Sharing of compounds and knowledge that can lead to new findings is the key to discovering new modes of action and a better understanding of disease biology. Many academic groups, but also small biotech companies around the world have already tested novel scientific hypotheses that expands beyond the known biology of the compounds and are free to publish their data to the benefit of the entire scientific community.68 The list of shared compounds arises from Novo Nordisk research projects and currently comprises 20 compounds plus reference compounds including a selection of antibodies and small molecules – and the list is continuously expanded. The compounds are well-characterized and available in vitro and in vivo data as well as structural properties, handling instructions and other relevant information is readily provided on the website to ensure high-quality experiments; see an example data package for long-acting amylin analogue.69Besides sharing of compounds, Novo Nordisk is also involved in other open innovation initiatives such as innovation challenges and public–private partnerships.70 Ultimately, open innovation will contribute to accelerating novel ideas into future breakthroughs for medical solutions.
Pfizer
Taken alongside functional genomics, chemical probes shared with the scientific community can spare biological resources and accelerate the next steps for early drug targets. From the outset of Pfizer's participation and collaboration with SGC and EUbOPEN, the mission has been to develop high-quality tool compounds and negative controls that are free from restriction on use. We have 22 chemical probes that have so far been co-developed or donated.71 The initial focus of chemical probe development (ca. 2009) was on epigenetic targets. Of the initial epigenetic chemical probes, PFI-3 stands out as a molecule that invalidated the BRG1/BRM bromodomain within the catalytic subunit of the SWI/SNF complex as a potential drug target72 while the SMARCA2/4 ATPase was shown to be more fruitful.72,73 However, PFI-3 confirmed that a ligandable binding site in BRG1/BRM bromodomain existed which went on to catalyze further hypotheses around degradation of sub-unit components;74 ATPase domain inhibitors have also followed.75,76More recently the gene family focus has extended into regulation of protein abundance, for example, E3 ligases and deubiquitinases. In a pilot project on E3s, Pfizer collaborated with the SGC to develop a chemical probe PFI-7 and a handle PFI-E3H1 for GID4,77 a subunit of the CTLH complex.78 The chemical and 3D structures, in vitro and cell-based potency, and approved vectors for synthesizing proximity pharmacology reagents were deposited in the public domain in September 2021. A recent publication disclosed another potent GID4 chemical tool which has a similar binding pose to PFI-7.79 Given the success with this pilot, SGC and EUbOPEN partners have set out to identify chemical matter that can extend the pool of E3s capable of inducing protein degradation through chimeric molecules, beyond the well-known cereblon and VHL state-of-the-art. Further discovered and donated probes29,50 from Pfizer will follow throughout the duration of EUbOPEN and beyond.
Takeda
Takeda has been an active partner of open science pre-competitive alliances for more than a decade. The initial focus on generating meaningful chemical probes for understudied targets focused on epigenetic targets, notably protein arginine methyltransferases,80,81 and bromodomains.82,83 More recently, Takeda's efforts have focused on expanding our understanding of the “dark” kinome via distribution of Takeda's bespoke library of kinase inhibitors to the kinase chemogenomic set (KCGS).63 Takeda has provided potent tool agents to probe pharmacology of several important drug targets, with unrestricted use for academic research. Some notable ones include GPR40 antagonist, glutamate receptor NMDA type subunit 2A,84 O-GlcNAcase inhibitor,85 RIPK1 kinase inhibitor,86 MetAP2 inhibitor,87 BCL6 inhibitor,88 kisspeptin receptor agonist,89 matrix metalloproteinase 13 inhibitor,90 and antioxidant response element activator.91 The aspiration behind this initiative is to engage broader scientific community in expanding the biological & pharmacological knowledge around these protein targets and elucidate their potential implication in human disease.Often surprising, serendipitous breakthroughs occur due to unbiased (yet non phenotypic) screening with such chemical probes if they are available. One example was recently exemplified where a pan CLK kinase chemical probe, known as T3,92 demonstrated robust effects on splicing regulations and efficacy in multiple oncology contexts.
While most inhibitors were developed internally as part of legacy discovery programs, Takeda has also directly participated in joint academic collaborations in open science which have led to multiple chemical probes; one notable example is the SGC3027 probe for PRMT7.81 Takeda remains committed to advancement of under explored areas of biology and will continue to be an active participant of EUbOPEN and similar open science pre-competitive partnerships.
Outlook
The opinions shared by industry highlight the commitment to open science, sharing knowledge and pharmacological tools. Industry's contributions have been crucial to the success of large PPP projects such as IMI-funded ULTRA-DD (https://ultra-dd.org), ReSOLUTE (https://re-solute.eu) and EUbOPEN (https://eubopen.org). In addition, pharmaceutical companies independently host unique, open innovation programs that cater to all levels of scientists from students to entrepreneurs. During the past year, the two global initiatives CACHE17 and OCN18 would not have been launched or be sustainable without the support and insight from the pharmaceutical industry. A graphical snapshot of the open-source tools and resources described in this article are summarized (Fig. 2).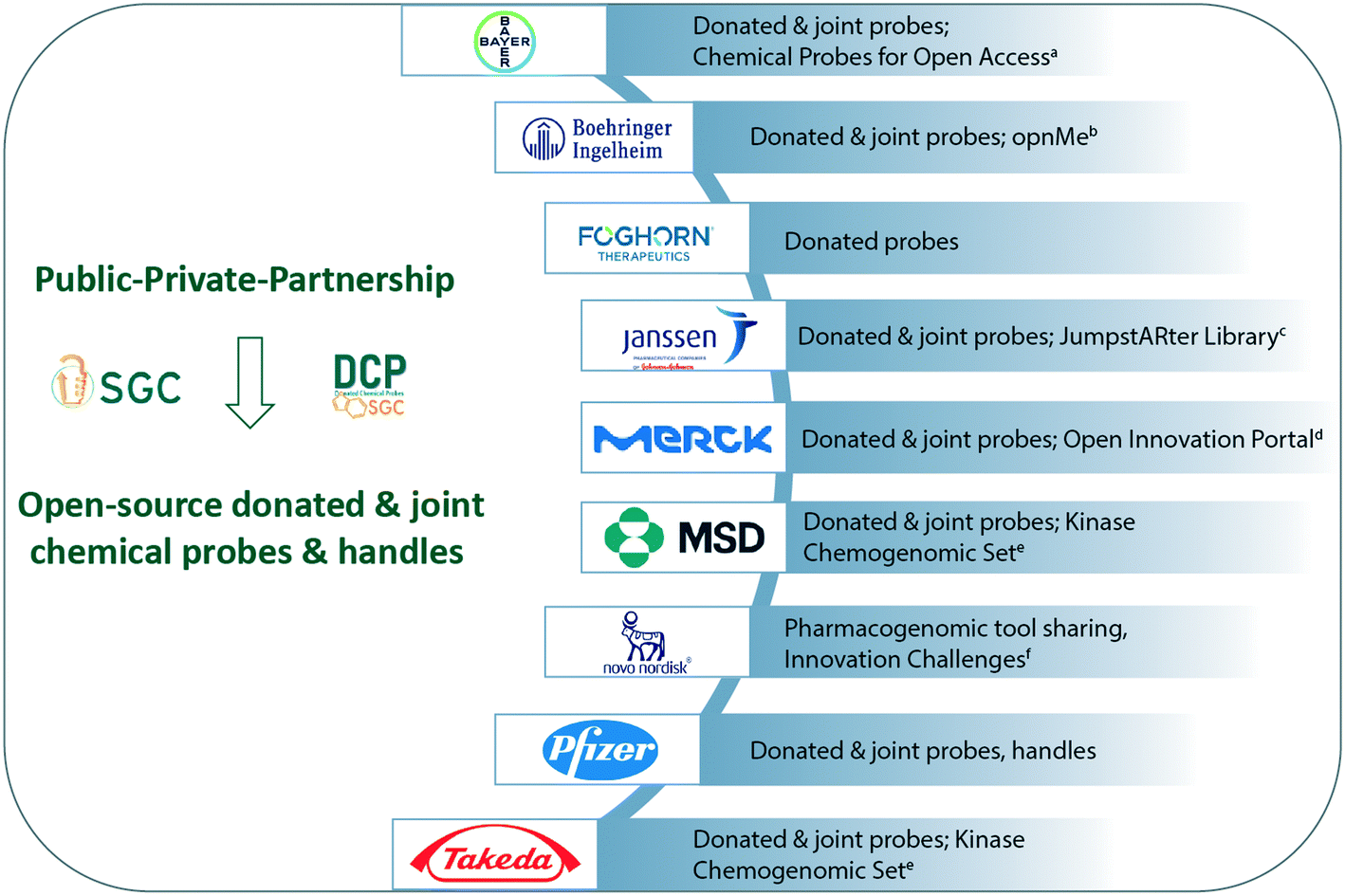 | ||
| Fig. 2 Private sector contributions to Open Science. The resources described by industry are summarized. Industry hosts a diverse set of open innovation programs, co-develop and donate pharmacogenomic tools, and often conduct off-target profiling. All chemical probes related to the SGC are listed here https://www.thesgc.org/chemical-probes, while the donated probes are listed here https://www.sgc-ffm.uni-frankfurt.de/. Between 1st May 2020 and 30th April 2025, the co-developed and donated probes and ligands will also be hosted on dedicated EUbOPEN web-pagesg,h. Where relevant the links to commercial suppliers are listed. ahttps://www.bayer.com/en/pharma/chemical-probes-open-access; bhttps://opnme.com/; chttps://www.wipo.int/edocs/pubdocs/en/wipo_pub_research_storybook_2019.pdf; dhttps://www.emdgroup.com/en/research/open-innovation.html; ehttps://ximbio.com/reagent/157681/the-kinase-chemogenomic-set-kcgs; fhttps://www.novonordisk.com/partnering-and-open-innovation/open-innovation/innovation-challenges.html; ghttps://www.eubopen.org/chemical-probes, hhttps://www.eubopen.org/chemical-handles. | ||
In the coming years, Target 2035 aims to continue to monitor and benchmark rapidly evolving technologies such as computer-aided drug design (CADD),93,94 DNA-encoded library screening followed by machine learning,95,96 affinity selection-mass spectrometry,97 automated chemical synthesis,98 and emerging applications of these technologies99–103 especially through the guidance of the private and public sectors. PPPs are a key component in developing faster, cost-effective methods to identify hits to novel targets thus accelerating the discovery of pharmacological modulators. While the private sector is predominantly focused on targets with a known disease link, government funding often supports research on a broader range of understudied proteins and protein families. For example, Genome Canada funded a two-year pilot to create drug discovery enabling reagents and know-how for the understudied WDR domain protein family. Likewise, EU funding supported the project EUbOPEN to find tool compounds for 1000 targets including targets of the ubiquitin pathway.
We hope that these accounts encourage all scientists, particularly those in the drug discovery industry, to contribute to open science projects including Target 2035.
Abbreviations
| Critical Assessment of Computational Hit-finding Experiments | CACHE |
| Computer-Assisted Drug Design | CADD |
| Enabling and unlocking Biology in the Open | EUbOPEN |
| Innovative Health Initiative | IHI |
| Innovative Medicines Initiative | IMI |
| Open Chemistry Networks | OCN |
| Public–private partnership | PPP |
| Structural Genomics Consortium | SGC |
| Unrestricted Leveraging of Targets for Research Advancement and Drug Discovery | ULTRA-DD |
| WD40 repeat | WDR |
Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors graciously thank Dr Shaunna Beedie who contributed to building the Target 2035 infrastructure. The SGC is a registered charity (number 1097737) that receives funds from Bayer Pharma AG, Boehringer Ingelheim, Canada Foundation for Innovation, Eshelman Institute for Innovation, Genentech, Genome Canada through Ontario Genomics Institute [OGI-196], Janssen, Merck KGaA (aka EMD Serono in Canada and US), Pfizer, Takeda, and the Innovative Medicines Initiative 2 Joint Undertaking (JU) under grant agreement No. 875510. The JU receives support from the European Union's Horizon 2020 research and innovation programme and EFPIA and Ontario Institute for Cancer Research, Royal Institution for the Advancement of Learning McGill University, Kungliga Tekniska Hoegskolan, Diamond Light Source Limited. This communication reflects the views of the authors, and the JU is not liable for any use that may be made of the information contained herein.References
- Target 2035, Towards medicines for all (https://www.target2035.net).
- A. J. Carter, et al., Target 2035: probing the human proteome, Drug Discovery Today, 2019, 24, 2111–2115 CrossRef CAS PubMed.
- S. Müller, et al., Target 2035 - update on the quest for a probe for every protein, RSC Med. Chem., 2022, 13, 13–21 RSC.
- C. H. Arrowsmith, et al., The promise and peril of chemical probes, Nat. Chem. Biol., 2015, 11, 536–541 CrossRef CAS PubMed.
- A. M. Edwards, et al., Too many roads not taken, Nature, 2011, 470, 163–165 CrossRef CAS PubMed.
- A. A. Antolin, et al., Objective, Quantitative, Data-Driven Assessment of Chemical Probes, Cell Chem. Biol., 2018, 25, 194–205.e5 CrossRef CAS PubMed.
- A. Schuhmacher, O. Gassmann, D. Bieniok, M. Hinder and D. Hartl, Open innovation: A paradigm shift in pharma R&D?, Drug Discovery Today, 2022, 27, 2395–2405 CrossRef PubMed.
- A. M. Davis, et al., Public-Private Partnerships: Compound and Data Sharing in Drug Discovery and Development, SLAS Discovery, 2021, 26, 604–619 CrossRef PubMed.
- S. M. Canham, et al., Systematic Chemogenetic Library Assembly, Cell Chem. Biol., 2020, 27, 1124–1129 CrossRef CAS PubMed.
- Innovative Health Initiative (https://www.ihi.europa.eu/).
- Unrestricted leveraging of targets for research advancement and drug discovery (https://www.imi.europa.eu/projects-results/project-factsheets/ultra-dd).
- Research empowerment on solute carriers (https://www.imi.europa.eu/projects-results/project-factsheets/resolute).
- G. Superti-Furga, et al., The RESOLUTE consortium: unlocking SLC transporters for drug discovery, Nat. Rev. Drug Discovery, 2020, 19, 429–430 CrossRef CAS PubMed.
- EUbOPEN: Enabling and unlocking biology in the OPEN (https://www.imi.europa.eu/projects-results/project-factsheets/eubopen).
- Target 2035 webinar recordings (https://www.target2035.net/archived-recordings).
- JOGL Space for Target 2035 (https://app.jogl.io/space/target2035).
- S. Ackloo, et al., CACHE (Critical Assessment of Computational Hit-finding Experiments): A public-private partnership benchmarking initiative to enable the development of computational methods for hit-finding, Nat. Rev. Chem., 2022, 6, 287–295 CrossRef PubMed.
- Open Chemistry Networks (https://github.com/OpenChemistry).
- C. D. Parks, et al., D3R grand challenge 4: blind prediction of protein-ligand poses, affinity rankings, and relative binding free energies, J. Comput.-Aided Mol. Des., 2020, 34, 99–119 CrossRef CAS PubMed.
- M. Amezcua, J. Setiadi, Y. Ge and D. L. Mobley, An overview of the SAMPL8 host-guest binding challenge, J. Comput.-Aided Mol. Des., 2022, 36, 707–734 CrossRef CAS PubMed.
- R. Song, Z. D. Wang and M. Schapira, Disease Association and Druggability of WD40 Repeat Proteins, J. Proteome Res., 2017, 16, 3766–3773 CrossRef CAS PubMed.
- CACHE Challenge #3: Finding Ligands Targeting the Macrodomain of SARS-CoV-2 NSP3 (https://cache-challenge.org/challenges/Finding-ligands-targeting-the-macrodomain-of-sars-cov-2-nsp3).
- M. Balasegaram, et al., An open source pharma roadmap, PLoS Med., 2017, 14, e1002276 CrossRef PubMed.
- D. Dolciami, et al., canSAR chemistry registration and standardization pipeline, Aust. J. Chem., 2022, 14, 28 Search PubMed.
- D. Mendez, et al., ChEMBL: towards direct deposition of bioassay data, Nucleic Acids Res., 2019, 47, D930–D940 CrossRef CAS PubMed.
- M. K. Gilson, et al., BindingDB in 2015: A public database for medicinal chemistry, computational chemistry and systems pharmacology, Nucleic Acids Res., 2016, 44, D1045–D1053 CrossRef CAS PubMed.
- E. R. Gold, The fall of the innovation empire and its possible rise through open science, Resour. Policy, 2021, 50, 104226 CrossRef PubMed.
- A. W. K. Yeung, et al., Open Innovation in Medical and Pharmaceutical Research: A Literature Landscape Analysis, Front. Pharmacol., 2020, 11, 587526 CrossRef PubMed.
- S. Muller, et al., Donated chemical probes for open science, eLife, 2018, 7, e34311 CrossRef PubMed.
- S. V. Frye, The art of the chemical probe, Nat. Chem. Biol., 2010, 6, 159–161 CrossRef CAS PubMed.
- A. M. Edwards, C. Bountra, D. J. Kerr and T. M. Willson, Open access chemical and clinical probes to support drug discovery, Nat. Chem. Biol., 2009, 5, 436–440 CrossRef CAS PubMed.
- J. Blagg and P. Workman, Choose and Use Your Chemical Probe Wisely to Explore Cancer Biology, Cancer Cell, 2017, 32, 268–270 CrossRef CAS PubMed.
- A. A. Antolin, P. Workman and B. Al-Lazikani, Public resources for chemical probes: the journey so far and the road ahead, Future Med. Chem., 2019, 13, 731–747 CrossRef PubMed.
- L. Liu, E. Rovers and M. Schapira, ChemBioPort: an online portal to navigate the structure, function and chemical inhibition of the human proteome, Database, 2022, baac088 CrossRef PubMed.
- Impact of AACR-Bayer Partnership (https://www.aacr.org/professionals/research-funding/grantees/impact-of-aacr-bayer-partnership/).
- A. Gollner, et al., https://opnMe.com: a digital initiative for sharing tools with the biomedical research community, Nat. Rev. Drug Discovery, 2022, 21, 475–476 CrossRef CAS PubMed.
- opnMe (https://opnme.com/).
- KGaA Open Innovation (https://www.emdgroup.com/en/research/open-innovation.html).
- Co-Create Knowledge for Pharma Innovation with Takeda Funding (https://www.takeda.com/en-au/what-we-do/research--development/cockpi-t/).
- Novo Nordisk Compound Sharing (https://www.novonordisk.com/partnering-and-open-innovation/compound-sharing.html).
- Bayer Chemical Probes (https://www.bayer.com/en/pharma/chemical-probes-open-access).
- F. von Nussbaum, et al., Freezing the Bioactive Conformation to Boost Potency: The Identification of BAY 85-8501, a Selective and Potent Inhibitor of Human Neutrophil Elastase for Pulmonary Diseases, ChemMedChem, 2015, 10, 1163–1173 CrossRef CAS PubMed.
- E. Eggert, et al., Discovery and Characterization of a Highly Potent and Selective Aminopyrazoline-Based in Vivo Probe (BAY-598) for the Protein Lysine Methyltransferase SMYD2, J. Med. Chem., 2016, 59, 4578–4600 CrossRef CAS PubMed.
- F. Göricke, et al., Discovery and Characterization of BAY-805, a Potent and Selective Inhibitor of Ubiquitin-Specific Protease USP21, J. Med. Chem., 2023, 66, 3431–3447 CrossRef PubMed.
- L. J. Martin, et al., Structure-Based Design of an in Vivo Active Selective BRD9 Inhibitor, J. Med. Chem., 2016, 59, 4462–4475 CrossRef CAS PubMed.
- J. Bottcher, et al., Fragment-based discovery of a chemical probe for the PWWP1 domain of NSD3, Nat. Chem. Biol., 2019, 15, 822–829 CrossRef PubMed.
- P. Martinelli, et al., Discovery of a Chemical Probe to Study Implications of BPTF Bromodomain Inhibition in Cellular and in vivo Experiments, ChemMedChem, 2023, e202200686 Search PubMed.
- NHE1 chemical probe (https://opnme.com/molecules/nhe1-bi-9627).
- S. Müller, et al., Donated chemical probes for open science, eLife, 2018, 7, e34311 CrossRef PubMed.
- Academia and pharma donated chemical probes (https://www.thesgc.org/donated-chemical-probes).
- Drugit (https://www.fold.it/drugit).
- SMARCA2, SMARCA4 chemical probe (https://www.sgc-ffm.uni-frankfurt.de/#!specificprobeoverview/FHT-2344).
- J. Varga, M. Kube, K. Luck and S. Schick, The BAF chromatin remodeling complexes: structure, function, and synthetic lethalities, Biochem. Soc. Trans., 2021, 49, 1489–1503 CrossRef CAS PubMed.
- Chemical probes, EUbOPEN (https://www.eubopen.org/chemical-probes).
- Amylin Analogue (https://www.novonordisk.com/partnering-and-open-innovation/compound-sharing/compound-details.316067).
- HRH4 chemical probe (https://www.eubopen.org/sites/www.eubopen.org/files/chemical-probes/EUbOPEN_ProbeSheet_JNJ-39758979_v1.pdf).
- CSNK1D, CSNK1E chemical probe (https://www.eubopen.org/sites/www.eubopen.org/files/chemical-probes/EUbOPEN_ProbeSheet_JNJ-6204_v1.).
- OGA chemical probe (https://www.eubopen.org/sites/www.eubopen.org/files/chemical-probes/EUbOPEN_ProbeSheet_JNJ-65355394).
- WIPO Re:Search Partnership Stories 2016-2019 Driving R&D for Neglected Infectious Diseases Through Global Cross-Sector Collaborations, 2019 (https://bvgh.org/wp-content/uploads/2019/11/WIPO-ReSearch-Partnership-Stories-2016-2019.pdf).
- A. Allali-Hassani, et al., Discovery of a chemical probe for PRDM9, Nat. Commun., 2019, 10, 5759 CrossRef CAS PubMed.
- NUDT5 chemical probe (https://www.thesgc.org/chemical-probes/MRK-952).
- PRMT9/5 chemical probe (https://www.thesgc.org/chemical-probes/MRK-990).
- C. I. Wells, et al., The Kinase Chemogenomic Set (KCGS): An Open Science Resource for Kinase Vulnerability Identification, Int. J. Mol. Sci., 2021, 22, E566 CrossRef PubMed.
- SRPK1/2/3 chemical probe (https://www.eubopen.org/sites/www.eubopen.org/files/chemical-probes/EUbOPEN_ProbeSheet_SRPK_v1.pdf).
- S. Müller, H. Lingard and S. Knapp, Selective Inhibition of Acetyl-Lysine Effector Domains of the Bromodomain Family in Oncology, in Nuclear Signaling Pathways and Targeting Transcription in Cancer 279-298, Humana Press, New York, NY, 2014 Search PubMed.
- T. Heinrich, et al., Discovery of 5-{2-[5-Chloro-2-(5-ethoxyquinoline-8-sulfonamido)phenyl]ethynyl}-4-methoxypyridine-2-carboxylic Acid, a Highly Selective in Vivo Useable Chemical Probe to Dissect MCT4 Biology, J. Med. Chem., 2021, 64, 11904–11933 CrossRef CAS PubMed.
- T. Heinrich, et al., Optimization of TEAD P-Site Binding Fragment Hit into In Vivo Active Lead MSC-4106, J. Med. Chem., 2022, 65, 9206–9229 CrossRef CAS PubMed.
- S. Lee, et al., Virgin β-Cells at the Neogenic Niche Proliferate Normally and Mature Slowly, Diabetes, 2021, 70, 1070–1083 CrossRef CAS PubMed.
- J. P. Daguer, et al., Identification of covalent bromodomain binders through DNA display of small molecules, Angew. Chem., Int. Ed., 2015, 54, 6057–6061 CrossRef CAS PubMed.
- Novo Nordisk Innovation Challenges (https://www.novonordisk.com/partnering-and-open-innovation/open-innovation.html).
- SGC chemical probes page (https://www.thesgc.org/chemical-probes/).
- B. S. Gerstenberger, et al., Identification of a Chemical Probe for Family VIII Bromodomains through Optimization of a Fragment Hit, J. Med. Chem., 2016, 59, 4800–4811 CrossRef CAS PubMed.
- B. Vangamudi, et al., The SMARCA2/4 ATPase Domain Surpasses the Bromodomain as a Drug Target in SWI/SNF-Mutant Cancers: Insights from cDNA Rescue and PFI-3 Inhibitor Studies, Cancer Res., 2015, 75, 3865–3878 CrossRef CAS PubMed.
- W. Farnaby, et al., BAF complex vulnerabilities in cancer demonstrated via structure-based PROTAC design, Nat. Chem. Biol., 2019, 15, 672–680 CrossRef CAS PubMed.
- A. M. Taylor, et al., GNE-064: A Potent, Selective, and Orally Bioavailable Chemical Probe for the Bromodomains of SMARCA2 and SMARCA4 and the Fifth Bromodomain of PBRM1, J. Med. Chem., 2022, 65, 11177–11186 CrossRef CAS PubMed.
- J. P. N. Papillon, et al., Discovery of Orally Active Inhibitors of Brahma Homolog (BRM)/SMARCA2 ATPase Activity for the Treatment of Brahma Related Gene 1 (BRG1)/SMARCA4-Mutant Cancers, J. Med. Chem., 2018, 61, 10155–10172 CrossRef CAS PubMed.
- GID4 chemical probe (https://www.biorxiv.org/content/10.1101/2023.01.17.524225v1.full.pdf).
- G. Onea, M. E. R. Maitland, X. Wang, G. A. Lajoie and C. Schild-Poulter, Distinct nuclear and cytoplasmic and interactomes of the mammalian CTLH E3 ligase complex, J. Cell Sci., 2022, 135, jcs259638 CrossRef CAS PubMed.
- C. K. Chana, et al., Discovery and Structural Characterization of Small Molecule Binders of the Human CTLH E3 Ligase Subunit GID4, J. Med. Chem., 2022, 65, 12725–12746 CrossRef CAS PubMed.
- K. Nakayama, et al., TP-064, a potent and selective small molecule inhibitor of PRMT4 for multiple myeloma, Oncotarget, 2018, 9, 18480–18493 CrossRef PubMed.
- M. M. Szewczyk, et al., Pharmacological inhibition of PRMT7 links arginine monomethylation to the cellular stress response, Nat. Commun., 2020, 11, 2396 CrossRef CAS PubMed.
- BRD9/7 chemical probe (https://www.thesgc.org/chemical-probes/TP-472).
- T. Lu, et al., Discovery of High-Affinity Inhibitors of the BPTF Bromodomain, J. Med. Chem., 2021, 64, 12075–12088 CrossRef CAS PubMed.
- F. Sakurai, et al., Discovery of Pyrazolo[1,5-a]pyrazin-4-ones as Potent and Brain Penetrant GluN2A-Selective Positive Allosteric Modulators Reducing AMPA Receptor Binding Activity, Bioorg. Med. Chem., 2022, 56, 116576 CrossRef CAS PubMed.
- M. Tawada, et al., Discovery of a Novel and Brain-Penetrant O-GlcNAcase Inhibitor via Virtual Screening, Structure-Based Analysis, and Rational Lead Optimization, J. Med. Chem., 2021, 64, 1103–1115 CrossRef CAS PubMed.
- M. Yoshikawa, et al., Discovery of 7-Oxo-2,4,5,7-tetrahydro-6 H-pyrazolo[3,4- c]pyridine Derivatives as Potent, Orally Available, and Brain-Penetrating Receptor Interacting Protein 1 (RIP1) Kinase Inhibitors: Analysis of Structure-Kinetic Relationships, J. Med. Chem., 2018, 61, 2384–2409 CrossRef CAS PubMed.
- C. McBride, et al., Discovery of potent, reversible MetAP2 inhibitors via fragment based drug discovery and structure based drug design-Part 2, Bioorg. Med. Chem. Lett., 2016, 26, 2779–2783 CrossRef CAS PubMed.
- T. Yasui, et al., Discovery of a novel B-cell lymphoma 6 (BCL6)-corepressor interaction inhibitor by utilizing structure-based drug design, Bioorg. Med. Chem., 2017, 25, 4876–4886 CrossRef CAS PubMed.
- T. Asami, et al., Design, synthesis, and biological evaluation of novel investigational nonapeptide KISS1R agonists with testosterone-suppressive activity, J. Med. Chem., 2013, 56, 8298–8307 CrossRef CAS PubMed.
- H. Nara, et al., Thieno[2,3-d]pyrimidine-2-carboxamides bearing a carboxybenzene group at 5-position: highly potent, selective, and orally available MMP-13 inhibitors interacting with the S1′′ binding site, Bioorg. Med. Chem., 2014, 22, 5487–5505 CrossRef CAS PubMed.
- H. Kimura, et al., BTZO-1, a cardioprotective agent, reveals that macrophage migration inhibitory factor regulates ARE-mediated gene expression, Chem. Biol., 2010, 17, 1282–1294 CrossRef CAS PubMed.
- T. Funnell, et al., CLK-dependent exon recognition and conjoined gene formation revealed with a novel small molecule inhibitor, Nat. Commun., 2017, 8, 7 CrossRef PubMed.
- W. P. Walters and R. Barzilay, Applications of Deep Learning in Molecule Generation and Molecular Property Prediction, Acc. Chem. Res., 2021, 54, 263–270 CrossRef CAS PubMed.
- B. Goldman, S. Kearnes, T. Kramer, P. Riley and W. P. Walters, Defining Levels of Automated Chemical Design, J. Med. Chem., 2022, 65, 7073–7087 CrossRef CAS PubMed.
- Unlocking the "Chemome" with DNA-Encoded Chemistry and Machine Learning (https://ai.googleblog.com/2020/06/unlocking-chemome-with-dna-encoded.html).
- Target 2035: WDR proteins as a technology testbed for illuminating the dark proteome (https://genomecanada.ca/project/target-2035-wdr-proteins-technology-testbed-illuminating-dark-prote).
- R. Prudent, D. A. Annis, P. J. Dandliker, J.-Y. Ortholand and D. Roche, Exploring new targets and chemical space with affinity selection-mass spectrometry, Nat. Rev. Chem., 2020, 5, 62–71 CrossRef.
- B. Mahjour, Y. Shen and T. Cernak, Ultrahigh-Throughput Experimentation for Information-Rich Chemical Synthesis, Acc. Chem. Res., 2021, 54, 2337–2346 CrossRef CAS PubMed.
- W. Su, et al., Triaging of DNA-Encoded Library Selection Results by High-Throughput Resynthesis of DNA-Conjugate and Affinity Selection Mass Spectrometry, Bioconjugate Chem., 2021, 32, 1001–1007 CrossRef CAS PubMed.
- A. L. Hackler, F. G. FitzGerald, V. Q. Dang, A. L. Satz and B. M. Paegel, Off-DNA DNA-Encoded Library Affinity Screening, ACS Comb. Sci., 2020, 22, 25–34 CrossRef CAS PubMed.
- W. G. Cochrane, et al., Activity-Based DNA-Encoded Library Screening, ACS Comb. Sci., 2019, 21, 425–435 CrossRef CAS PubMed.
- Y. Huang, et al., Selection of DNA-encoded chemical libraries against endogenous membrane proteins on live cells, Nat. Chem., 2021, 13, 77–88 CrossRef CAS PubMed.
- Y. Huang, Y. Li and X. Li, Strategies for developing DNA-encoded libraries beyond binding assays, Nat. Chem., 2022, 14, 129–140 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2023 |