
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Dietary oleacein, a secoiridoid from extra virgin olive oil, prevents collagen-induced arthritis in mice
María Ángeles
Rosillo
 *ab,
Isabel
Villegas
ab,
Victoria
Vázquez-Román
c,
José María
Fernández-Santos
c,
Juan
Ortega-Vidal
d,
Sofía
Salido
d,
María Luisa
González-Rodríguez
e and
Catalina
Alarcón-de-la-Lastra
ab
*ab,
Isabel
Villegas
ab,
Victoria
Vázquez-Román
c,
José María
Fernández-Santos
c,
Juan
Ortega-Vidal
d,
Sofía
Salido
d,
María Luisa
González-Rodríguez
e and
Catalina
Alarcón-de-la-Lastra
ab
aDepartment of Pharmacology, Faculty of Pharmacy, Universidad de, Sevilla, Calle Profesor García González 2, 41012 Seville, Spain. E-mail: rosillo@us.es; Tel: +34-954559880
bInstituto de Biomedicina de Sevilla, IBiS/Hospital Universitario Virgen del Rocío/CSIC/Universidad de Sevilla, 41013 Seville, Spain
cDepartment of Normal and Pathological Cytology and Histology, Faculty of Medicine, Universidad de Sevilla, 41012 Seville, Spain
dDepartment of Inorganic and Organic Chemistry, Faculty of Experimental Sciences, Campus de Excelencia Internacional Agroalimentario (ceiA3), Universidad de Jaén, 23071 Jaén, Spain
eDepartment of Pharmaceutical Technology, Faculty of Pharmacy, Universidad de Sevilla, 41012 Seville, Spain
First published on 11th December 2023
Abstract
Olacein (OLA), one of the main secoiridoids derived from extra virgin olive oil (EVOO), has been shown to modulate oxidative and inflammatory responses in various pathological conditions; however, its potential benefit in joint disorders such as rheumatoid arthritis (RA) is unknown. Therefore, this study was designed to evaluate the preventive role of the effects of an OLA-supplemented diet in the murine model of collagen-induced arthritis (CIA), delving into the possible mechanisms and signaling pathways involved. Animals were fed an OLA-enriched preventive diet for 6 weeks prior to CIA induction and until the end of the experimental time course. On day 43 after the first immunization, mice were sacrificed: blood was collected, and paws were histologically and biochemically processed. Dietary OLA prevented collagen-induced rheumatic bone, joint and cartilage conditions. Circulating matrix metalloproteinase (MMP)-3 and proinflammatory cytokine (IL-6, IL-1β, TNF-α, IL-17) levels were significantly decreased in the joint, as well as MMP-9 and cathepsin-K (CatK) expression in secoiridoid-fed animals. In addition, dietary OLA was able to decrease COX-2, mPGES-1 and iNOS protein expressions and, also, PGE2 levels. The mechanisms possibly involved in these protective effects could be related to the activation of the Nrf-2/HO-1 axis and the inhibition of proinflammatory signaling pathways, including JAK-STAT, MAPKs and NF-κB, involved in the production of inflammatory and oxidative mediators. These results support the interest of OLA, as a nutraceutical intervention, in the management of RA.
1. Introduction
Immune-mediated inflammatory diseases (IMIDs) are a clinically heterogeneous group of chronic and highly disabling diseases that share common inflammatory sequences and dysregulation of the immune system. IMIDs result in substantial levels of morbidity, a significant reduction in quality of life and premature deaths.1One of the most prevalent is rheumatoid arthritis (RA) which presents with symmetrical erosive synovitis, in which the granulation tissue of the synovium (pannus) degenerates, invading and eroding the cartilage and bone of the diarthrodial or synovial joints. It encompasses a wide range of features, from a progressively chronic disease with varying degrees of joint destruction to clinically evident extra-articular manifestations.2
The etiology of RA is unknown although genetic, infectious, environmental (smoking, stress and trauma, obesity, diet) and hormonal factors appear to be involved in related and complex pathways.3
While there is no consensus on the nature of the self-antigen(s) or environmental factors that initiate the dysregulation of the immune response (innate and adaptive) that originates and perpetuates the inflammatory process, it is well established that the immunocompetent cells involved are macrophages, regulatory T cells (Tregs), T cells (Th-1 and Th-17) and cytotoxic T cells that generate an imbalance in the production of Th1 cytokines: interferon (IFN)-γ, tumour necrosis factor (TNF)-α and the interleukins (IL)-1β, IL-2, IL-4, IL-6, IL-13, IL-15, IL-23, IL-27, IL-35; Th17: IL-17, IL-21 and, Th2: IL-10, IL-20, IL-22, in addition to chemokines (CXCR4, IP-10, SDF1, MCP-1) and metalloproteinases (MMP-3 and MMP-9) capable of digesting cartilage. B-lymphocytes also produce autoantibodies such as rheumatoid factor (RF) and anti-cyclic citrullinated peptide (anti-cyclic citrullinated peptide) antibodies (anti-CCP).4
The molecular signaling pathways involved include the nuclear transcription factors: nuclear transcription factor (NF-κB), forkhead box P3 (FOXP3), phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) the mitogen-activated protein kinases (MAPKs): c-Jun NH2-terminal kinase (JNK) and p38 kinase, primarily janus kinase/signal transduction and transcription activation protein (JAK/STAT) among others.5
The therapeutic approach to RA is multidisciplinary and is mainly directed at non-specific suppression of the inflammatory process with the aim of mitigating the symptoms and signs of the disease, encompassing medical, physiotherapeutic, and other methods, including nutritional therapy. In relation to pharmacotherapy, although there are currently new drugs developed for RA, such as JAK inhibitors (JAKi), which have led to a major change in treatment, no drug strategy provides an effective treatment alternative that achieves continuous disease control and is safe for patients with chronic use.6
Therefore, nutritional therapy today, including dietary modifications and the use of nutritional supplements, in addition to dietary support, can be an interesting alternative in the management of RA by generating prophylactic and therapeutic effects without the undesirable effects that accompany classical pharmacotherapy, contributing to the reduction of comorbidities and increasing the quality of life of patients. Considering that inflammation is one of the key players in the pathogenesis of RA, nutrition has been shown to influence disease flares, overall treatment, and clinical outcomes. In fact, it is suggested that nutrition may play a direct role through the intake of anti-inflammatory foods such as extra virgin olive oil (EVOO).7
The beneficial properties of EVOO are known to be due to its fatty acid composition and also its minority bioactive compounds in particular polyphenols, among which are the secoiridoids oleuropein and ligstroside aglycones, and their hydrolysis derivatives such as oleacein (OLA) and oleocanthal, among others. In this context, in the last decade, our research group has demonstrated the therapeutic effects of dietary supplementation with EVOO 8 and some of its bioactive compounds in a collagen-induced arthritis (CIA) in mice.8–10 This animal model is one of the most widely used to study and evaluate key RA pathogenic mechanisms and novel anti-arthritic drugs. It is defined by the production of CII-specific antibodies, a hallmark also described in RA.11
OLA is a diphenolic dialdehyde that is formed from the secoiridoid glucoside oleuropein (present in olive fruits) during the elaboration of olive oil. Interesting studies performed have reported that OLA may play a beneficial role against cardiovascular diseases based on the prevention of early and advanced atherosclerotic lesions.12 Also, some bioactivities of OLA like antimicrobial, antiproliferative and antisteatotic effects have been documented.13–16
Other health-promoting properties of OLA include antioxidant and anti-inflammatory activities. In fact, we have recently shown the immunomodulatory effects of OLA in LPS-activated murine immune cells by reducing oxidative damage and amelioration of the production of essential pro-inflammatory cytokines through the prevention of MAPKs and NF-κB pathways activation.17 Moreover, in the context of immune-inflammatory diseases, OLA was effective in preventing the induced immuno-inflammatory response in a murine model of encephalomyelitis, improving neuromotor disabilities and reducing central nervous system damage18 and in an animal experimental model of systemic lupus erythematosus (SLE).19
Nevertheless, to date, no data are available on the potential beneficial effects of OLA in an in vivo model of RA. Therefore, this study was conducted to assess the prophylactic effects of an OLA-enriched diet in CIA murine model. To this end, we examined the macroscopic and histological damage, the production of inflammatory and oxidative mediators and explored the possible signaling pathways implicated.
2. Materials and methods
2.1. Chemical synthesis of (−)-oleacein
The starting material for the synthesis of (–)-oleacein [(–)-oleuropein] was purchased from Sigma-Aldrich (Madrid, Spain). Solvent used for chromatography separations and synthetic procedures, such as hexane (Hex), ethyl acetate (EtOAc), ethanol (EtOH), methanol (MeOH), dimethyl sulfoxide (DMSO), chloroform (CHCl3) and diethyl ether (Et2O) were purchased from VWR (Madrid, Spain). Other reagents, solvents, material, and instruments used in this work were previously described.10A mixture of (–)-oleuropein (0.40 g) and H2O (25 mL) was subjected to microwave irradiation for 13 minutes at 180 °C. The resulting crude reaction was extracted with CHCl3 (3 × 30 mL). Organic layers were combined, dried over anhydrous Na2SO4, filtered and evaporated under reduced pressure to give a dark brown oil. The dry crude reaction was purified by fast centrifugal partition chromatography (FCPC) (Hex![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :EtOAc:EtOH:H2O, 1:1:1:1, v/v/v/v) to give (–)-oleacein (0.14 g, 58% reaction yield). The structure of this compound was confirmed by comparison of the NMR data with those previously reported.17,20 This procedure was repeated several times up to get the necessary amount of (–)-oleacein (around 1.8 g) to prepare the diets for mice.
:EtOAc:EtOH:H2O, 1:1:1:1, v/v/v/v) to give (–)-oleacein (0.14 g, 58% reaction yield). The structure of this compound was confirmed by comparison of the NMR data with those previously reported.17,20 This procedure was repeated several times up to get the necessary amount of (–)-oleacein (around 1.8 g) to prepare the diets for mice.
2.2. Diet elaboration
Diets were enriched with film-coated pellets impregned with OLA (OLA group) or non-impregned film-coated pellets (the naïve and SD-CIA group). At first, OLA (150 mg) was solved in dichloromethane (DCM) (45 mL) and added to 105 g of sucrose pellets, which were placed in a round bottom flask. The DCM was evaporated in a R-200 rotary vacuum evaporator (Büchi®, Flawil, Switzerland) for the impregnation of the pellets with OLA. After DCM evaporation, the pellets were dried in the dark, at room temperature. For film-coating, Eudragit® EPO was used. Eudragit® EPO is an excellent film-forming polymer, which protects the drug from humidity and light. Sodium dodecylsulfate (0.25 g) and stearic acid (0.4 g) were used as a stabilizer and a plasticizer, respectively. Sodium dodecylsulfate and stearic acid were dissolved in absolute ethanol (30 g) for 30 min at 40° C. Next, 2.65 g of Eudragit® EPO was added and the solution was homogenized with a T18 digital ULTRA-TURRAX® (IKA, Staufen, Germany) over a period of 90 min, forming a colloid. After homogenization, we added titanium dioxide (0.25 g) and talc (0.32 g) and blended it again in the ULTRA-TURRAX® for 15–20 min. Finally, to remove particle aggregates, the final solution was passed through a sieve (350 μm). Ultimately, 100 g of non-impregnated or OLA-impregnated pellets were coated in a fluidized bed coater system (Glatt®, Binzen, Germany) using bottom spray. The atomization pressure (1 bar), air temperature (40° C), coating solution flow rate (0.7 mL min−1), and nozzle diameter (0.5 mm) were kept constant throughout the process. The film-coated pellets were added to standard rodent powder chow (diet supplementation: 0.025%, 250 mg kg−1) and homogenized with a Turbula® 3D mixer (WAB, Basel, Switzerland). Then, compacted diets were prepared based on our previous reports.21Experimental diets were prepared following the standard reference diet of the American Institute of Nutrition (AIN). The CIA and naïve groups were fed with the standard diet (SD) supplemented with non-impregnated film-coated pellets, while the OLA-group mice received a diet enriched with OLA film-coated pellets.
2.3. Animals and diet
Thirty-six three weeks old male DBA-1/J mice (Janvier®, Le Genest St Isle, France) were accommodated under standard conditions (24–25 °C, humidity 70–75%, lighting regimen 12L/12D) in our Animal Laboratory Center. They were fed pellet diets and water ad libitum. At 4 weeks aged animals were randomized into three experimental groups: (1) Naïve group received a SD (n = 12), (2) CIA group received a SD (SD-CIA) (n = 12), and (3) CIA-OLA group which received the SD enriched with OLA film-coated pellets (0.025% (w/w)) dietary enrichment percentage selection based on our previous reports9,22 (n = 12). All diets were formulated based on the AIN standard reference diet and were prepared by mixing the respective compounds under yellow light and stored at −80 °C. Fresh diets were provided daily. The animals were fed with the corresponding diet during six weeks before the CIA induction and during the experiment time. Animal care and procedures agreed with protocols approved by Animal Ethics Committee of the Universidad de Sevilla (ethical approval number CEEA-US2022-13), and recommendations of European Union regarding animal experimentation (Directive of the European Counsel 2012/707/EU).2.4. Induction of CIA
Arthritis was induced in ten weeks old male DBA-1/J mice. Bovine type II collagen (CII) 2 mg mL−1 in dilute acetic acid (MD Bioproducts®, Zürich, Switzerland) was emulsified in equal volumes of Freund complete adjuvant (2 mg ml−1Mycobacterium tuberculosis, strain H37Ra; Difco®, Detroit, Michigan, USA). On day 0, DBA-1/J mice were immunized at the base of the tail with 100 mg of bovine CII. On day 21, mice received an intraperitoneal booster injection of 100 mg of CII dissolved in phosphate buffered saline (PBS). Mice were considered to have arthritis when significant changes in redness and/or swelling were noted in the digits or in other parts of the paws. Joint inflammation was scored visually in each paw, using a scale of 0–2 where 0 = uninflamed, 1 = mild, 1.5 = marked and 2 = severe. Scoring was performed by two independent observers without knowledge of the experimental groups.2.5. Histopathology
For histological evaluation, hind paws were removed, fixed in neutral buffered 4% formaldehyde for 24 h, and decalcified in 5% formic acid. Paws were then embedded in paraffin, sectioned at 4 μm thickness, and stained with Hematoxylin–Eosin (H&E) and Masson's Trichrome stain (TM), following standard procedures. The severity of arthritis in tarsal and metatarsophalangeal joints was evaluated and a subjective score was employed to assess the histopathological degree of the lesions.All analyses were performed at the light microscopic level under blinded conditions.
2.6. Immunohistochemical analysis
Paw sections were mounted in 3-aminopropyl triethoxy-silane-coated slides, dewaxed in xylene and hydrated through graded ethanols. For cathepsin K (CatK) immunodetection, a proteinase K (0,05 mol L−1 Tris buffer pH 7,6) digestion step for 30 min at 37 °C was carried out. Subsequently, samples were treated with 3% hydrogen peroxide to block endogenous peroxidase activity for 20 min at room temperature and incubated overnight at 4 °C in a humidified chamber, either using a rabbit-polyclonal CatK antibody (1:500; Invitrogen™, ThermoFisher Scientific, USA) or a mouse-monoclonal MMP-9 antibody (1:500; Invitrogen™, ThermoFisher Scientific, USA). Vectastain ABC-HRP Kit (Vector, USA) was used as the labeling system following manufacturer's instructions, and 3,3-diaminobenzidine tetrahydrochloride (DAB) solution (Sigma-Aldrich, Darmstadt, Germany) was used as chromogen. The specificity of the reactions was controlled by replacing the primary antibody with normal horse serum. The slides were counterstained with Harris' Haematoxylin, dehydrated and coverslipped. Thereafter, photomicrographs were taken using an Olympus photomicroscope and a Nikon DS-Fi3 camera.
2.7. Enzyme-linked immunosorbent assay (ELISA)
MMP-3 was measured in serum using an ELISA kit with 10 pg mL−1 of sensitivity (RayBio® Norcross, GA, USA). To determinate levels of pro-inflammatory cytokines, hind paws were homogenized in 1 mL of 10 mM HEPES buffer, pH 7.4. Supernatants were used for the determination of PGE2 (Cayman Chemical®, Michigan, USA), IL-1β (R&D Systems® Inc., Minneapolis, MN, USA), IL-6 (Diaclone®, Besan on Cedex, France), TNF-α and IL-17 (Peprotech®, London, UK).2.8. Western blotting
Hind paws were homogenized in liquid N2 with a phosphate buffer according to Rosillo et al., to isolate proteins.23 Protein concentration was determined with Bradford's colorimetric method.24 A total of 10 μg of proteins were separated on 10% sodium dodecylsulphate polyacrylamide gel electrophoresis, transferred to nitrocellulose membrane and, incubated overnight at 4 °C with specific primary antibodies: COX-2, iNOS, pJNK, JNK, pp38, p38, Nrf-2, pJAK-3, pSTAT-3, IκB-α, pp65 (Cell Signaling Technology®, Danvers, MA, USA), HO-1 (Enzo®, Madrid, Spain), mPGES-1 (Abcam®, Cambridge, MA, USA). Membranes were incubated for 2 h at room temperature with horseradish peroxidase-labeled secondary antibody antirabbit or antimouse (Cell Signaling Technology®, Danvers, MA, USA). Blots were analyzed with β-actin antibody (Cell Signaling Technology®, Danvers, MA, USA) to prove equal loading. The immunosignals were captured with ChemiDoc Imaging Systems (Bio-Rad® Madrid, Spain) and signals were analyzed and quantified using Image Processing and Analysis in Java (ImageJ, Softonic).2.9. Data analysis
All values are expressed as arithmetic means ± standard desviation (SD) in figures and text. Results were evaluated using Graph Pad Prism version 9 software (San Diego, CA, USA), analyzing the statistical significance by one-way analysis of variance (ANOVA), followed by Tukey's multiple comparisons test. p-values < 0.05 were considered statistically significant. Figures from densitometry experiments are representatives of different experiments performing on a different day.3. Results
3.1. Chemistry
The synthesis of (–)-oleacein (OLA) used in this work was performed in one step from commercial (–)-oleuropein under Krapcho decarbomethoxylation conditions (Scheme 1). The microwave irradiation of a mixture of (–)-oleuropein and water at 180 °C for 13 minutes, followed by the purification of the crude reaction by FCPC allowed us to get pure OLA in a 58% yield. | ||
| Scheme 1 Synthesis of (–)-oleacein (OLA) from (–)-oleuropein. | ||
This procedure is based on the attractive semi-synthesis of OLA from oleuropein reported by Vougogiannopoulou and collaborators.20 These authors refluxed oleuropein, dissolved in wet dimethylsulfoxide (DMSO), with two equivalents of an inorganic salt (NaCl) for 10 h to get OLA with a 21% yield, after purification by silica gel column chromatography. Procopio's group later improved the conversion of oleuropein changing DMSO by water and heating in a microwave reactor, with the presence of inorganic salts, to get OLA in just 20 minutes with a 48% yield.25 For that time, we were also working in the improvement of the semisynthesis of OLA from oleuropein, and that of oleocanthal from ligustroside,10 following the recommendations of a previous work aimed at adapting the classical Krapcho decarboxylation experimental conditions to an aqueous microwave scenario.26 As a result, we changed DMSO by water, and we decided not adding inorganic salts in one of our assays, finding that the reaction of semi-synthesis of OLA from oleuropein worked properly. This full conversion of oleuropein into OLA, together with the use of the FCPC technique for OLA purification, allowed us to get OLA in a better yield despite the known sensitivity of this dialdehyde.
3.2. Effects of dietary OLA on CIA-induced AR model
CIA mouse model is a widely studied autoimmune model of RA. The development of arthritis was monitored until day 42. The time course of arthritic score indicated that control CIA mice developed a progressive development of arthritis observed from day 29 (Fig. 1A). On the contrary, animals fed with OLA-diet showed a significant delayed onset (p < 0.05 vs. CIA) reducing footpad thickness and inflammation of forelegs and hind legs in comparison with SD-CIA group from days 35 to 42. Representative photographs of hind paws from the different experimental animal groups are shown in Fig. 1B. | ||
| Fig. 1 Evolution of arthritis severity in collagen–induced arthritis (CIA) mice from day 27 to 43 after second immunization. (A) Changes in paw swelling and thickness on arthritic mice. Sings was scored every two days up to the end of experiment. Naïve group: non-arthritic mouse fed with standard diet (SD); CIA control group: collagen-induced arthritis mice fed SD; OLA: arthritic mice fed with 0.025% w/w oleacein-supplemented diet. Values are presented as mean ± S.E.M. (n = 10).+p < 0.05; ++p < 0.01 vs. naïve group; *p < 0.05 vs. CIA group. (B) image of hind paws at the end of experiment (Day 43): naïve group with SD; CIA control group with SD; and OLA supplemented diet arthritic group. | ||
Besides, histological changes among control, CIA and OLA supplemented group were compared. The degree of severity of histological lesions was established as negative, mild, moderate, and severe (Table 1).
| Groups | Soft tissue inflammation | Synovial infiltration | Inflammatory Pannus | Cartilage loss and bone resorption |
|---|---|---|---|---|
| Negative (−); mild (+); moderate (++); severe (+++). | ||||
| Naïve | − | − | − | − |
| CIA | +++ | +++ | +++ | +++ |
| OLA | + | − | − | − |
In naïve mice, a thin synovial membrane that limited the synovium and articular cartilage appeared as normal. Moreover, no inflammatory signs were found in the surrounding soft tissues in any of the joints studied (Fig. 2A–C). In contrast, in CIA groups evidence of arthritic signs was observed (Fig. 2D–F): Proliferation of synovial cells leaded to a thickening of synovial membrane with formation of villi into the synovial space, cartilage erosion with pyknotic nuclei in chondrocytes as well as areas of bone resorption mixed with decalcified cartilage were also observed. Additionally, a general inflammatory pannus invasion of the joint cavity was observed, along with a general loss of the joint architecture. After OLA supplementation, inflammatory and arthritic signs were almost not detected, and synovium appearance was very similar to that observed in controls. Only in some samples, soft tissue around joints showed moderate inflammation and a mild proliferation of synovial cells was also found (Fig. 2G and H).
 | ||
| Fig. 2 Histologic analysis of tarsal and metatarsophalangeal joints from normal, CIA and OLA mice. Compared with naïve (A–C), generalized inflammation was observed in CIA group with pannus invasion (D), and loss of architecture joints (F). As shown in (D) and (E) respectively, cartilage was severely eroded (arrows), and bone resorption (head arrow) was observed. Joints in OLA supplementation group (I) were similar to controls (C) and only mild synovial membrane thickening (arrows, G) and some soft tissue inflammation foci were observed (asterisk, H). However, bone and cartilage damage were prevented in this group. Bar = 25 μm in E; bar = 50 μm in B, C, D, G, H, and I; Bar = 100 μm in A; bar = 250 μm in F. | ||
3.2. Effects of dietary OLA on immunohistochemical detection of CatK and MMP-9 in joints
Immunohistochemical studies for MMP-9 showed differences among the different experimental mice. MMP-9 immunostaining was generally positive in the surface synovial lining, small blood vessels inflammatory-cells and fibroblasts. When cartilage was affected, chondrocytes in the surface of articular cartilage were positive and when damage increased, chondrocytes inside cartilage turned positive too. In accordance with the histopathological findings described above, immunostaining for MMP-9 was found to be higher in CIA group, being in OLA group similar to that observed in controls (Fig. 3A, C and E). | ||
| Fig. 3 Immunolocalization of MMP-9 (A, C and E) and CatK antigen (B, D and F) in tarsal and metatarsal joints from naïve, control CIA and OLA mice. Synovial lining-cells (head arrows), periarticular soft-tissues (asterisk), inflammatory pannus and areas of bone and cartilage resorption were positive for both markers in control CIA and OLA mice groups. Moreover, chondrocytes in cartilage eroded zones were positive for MMP-9 (arrows, C and E). Immunostaining extension and intensity increased according to the histopathological damage. Differences among groups were more evident for CatK. Bar = 50 μm in (A–E) and 100 μm in (D). | ||
CatK is part of the family of cysteine proteases involved in many relevant mechanisms, such as the degradation activity of collagen 1 and elastin in bone resorption. Altered CatK levels are linked to various pathologies, mainly related to bone and cartilage degradation, such as osteoarthritis (associated with CatK overexpression).27 Therefore, we evaluated CatK overexpression on the paws of normal, CIA and treatment groups.
In both, control and CIA mice, a fraction of cells in the synovial membrane were immunopositive for CatK but, while in controls the immunostaining was only seen in a thin lining, in CIA group, because of its increased thickness, signal was stronger and extended to the subliming areas of synovium villi, being highly positive all along the inflammatory pannus. Moreover, bone surfaces were positive in both control and CIA mice but in the second the number of positive cells was considerably greater. Furthermore, in CIA joints, chondrocytes of the articular surface as well as the pericellular matrix exhibited cytoplasmic CatK staining. Eroded cartilage surface and resorption bone areas were also positive. In OLA supplemented group only some cells of synovial villi were immunopositive for CatK and, similarly to control animals, chondrocytes and surface cells remained negative (Fig. 3B, D and F).
3.3. Effects of dietary OLA on MMP-3 and joint inflammatory biomarkers levels
It has been reported that TNF-α, IL-1β, IL-6 and IL-17 are critical cytokines involved in the pathogenesis of RA.28 Moreover, pro-inflammatory cytokines, including TNF-α, IL-1β and IL-6, may stimulate MMP genes through the binding of several different transcription factors.30 To explore whether dietary could regulate these joint inflammatory biomarkers, we measured cytokine levels in paw homogenates and MMP-3 in serum. As shown in Fig. 4A, TNF-α, IL-1β, IL-6 and IL-17 levels were significantly increased in paw homogenates from CIA animals when compared with naïve mice (p < 0.001 vs. naïve group) suggesting its relationship with the synovial tissue inflammation. | ||
| Fig. 4 Effects of dietary OLA on MMP-3 and joint inflammatory biomarkers levels. (A) OLA diet down-regulated the levels of pro-inflammatory cytokines: TNF-α, IL-1β, IL-6 and IL-17 in tissue homogenates. (B) MMP-3 levels were decreased in serum of animals fed with OLA diet. Naïve group: non-arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values are represented as mean ± S.E.M. (n = 8), +++p < 0.001 vs. naïve group; *p < 0.05; **p < 0.01; ***p < 0.001 vs. CIA group. | ||
However, our results indicated a significant reduction of pro-inflammatory cytokines production in paw homogenates from animals fed with OLA-diet (IL-1β: p < 0.01; TNF-α: p < 0.05; IL-6: p < 0.01; IL-17: p < 0.05 vs. CIA group) (Fig. 4A).
Furthermore, circulating levels of MMP-3 were raised in SD-CIA animals, in parallel to the disease severity (p < 0.001 vs. naïve group) (Fig. 4B). On the contrary, a significant reduction of MMP-3 levels was observed in OLA-enriched diet fed animals (p < 0.001 vs. CIA group).
3.4. Effects of dietary OLA on COX-2, mPGES-1 protein expression and PGE2 production
COX-2 and mPGES1 expressions were determined by western blot in paw homogenate (Fig. 5A). Arthritic control animal group showed an overexpression of both these pro-inflammatory enzymes (COX-2: p < 0.01 and mPGES1: p < 0.01 vs. naïve group) however OLA diet was able to reduce the protein expression levels of both of them (p < 0.01 vs. CIA group) (Fig. 5B). On the other hand, levels of PGE2 were measured on the paw homogenate, OLA was capable to reduce the levels of these eicosanoid (p < 0.01 vs. CIA group) (Fig. 5B). | ||
| Fig. 5 Modulation of COX-2/mPGES-1 axis and PGE2 synthesis in dietary OLA supplemented group in hind paws. (A) COX-2 and mPGES-1 protein expression were analyzed by western blotting and quantified by densitometrical analysis. Western blot images are representative of separate experiments with similar results. (B) Quantification of PGE2 levels using ELISA kit. Naïve group: non–arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values represented mean ± S.E.M. (n = 8), ++p < 0.01; +++p < 0.001 vs. naïve group; **p < 0.01 vs. CIA group. | ||
3.5. OLA experimental diet attenuated iNOS overexpression
iNOS is considered a critical oxidative stress marker involved in nitric oxide (NO) production, a free radical inductor of oxidation.29 The protein expression of iNOS was significantly up-regulated in paws homogenates from SD-CIA mice in comparison to SD-naïve control group (p < 0.01 vs. naïve group). Whereas the protein expression of this pro-inflammatory enzyme was downregulated in paws from arthritic mice fed with OLA diet (p < 0.001 vs. CIA group) (Fig. 6). | ||
| Fig. 6 Protein expression of iNOS in hind paws homogenate. Protein expression was quantified by densitometry analysis and normalized with respect to β-actin (housekeeping). Western blot images are representative of separate experiments with similar results. Naïve group: non-arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values represented mean ± S.E.M. (n = 8), ++p < 0.01 vs. naïve group; ***p < 0.001 vs. CIA group. | ||
3.6. OLA diet induces Nrf-2/HO-1 antioxidant pathway activation
Nuclear factor erythroid 2-related factor (Nrf-2) is a transcription factor that plays a key role in cell protection against oxidative stress. It induces antioxidant enzymes that inactivate reactive oxygen species, like haem oxigenase-1 (HO-1).30The expression of the proteins HO-1 and Nrf2 were evaluated in paw homogenates by western blotting. Our data show that Nrf2 and HO-1 was significantly downregulated in CIA mice during the maintenance of chronic inflammation (Nrf2: p < 0.001; HO-1: p < 0.01 vs. naïve group); however, dietary OLA treatment induced a Nrf2 and HO-1 overexpression in comparison with CIA arthritic control group (p < 0.05 vs. CIA group) (Fig. 7).
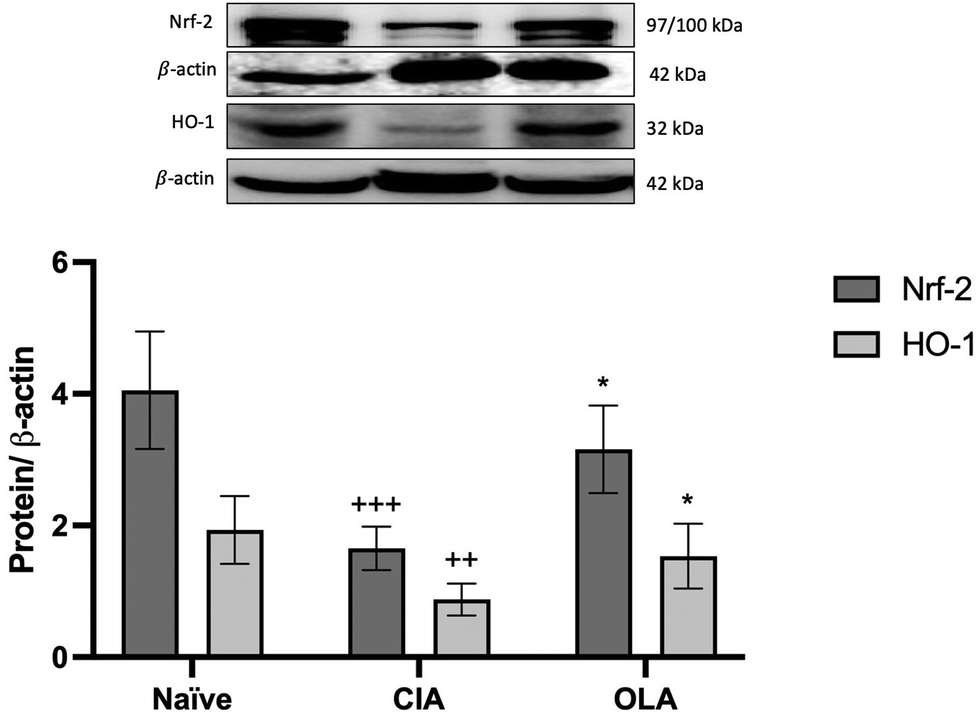 | ||
| Fig. 7 OLA dietary treatment induced up-regulation of Nrf-2/HO-1 protein expression in hind paws homogenates. Densitometry analysis was normalized with respect to β-actin (housekeeping). Western blot images are representative of separate experiments with similar results. Naïve group: non-arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values represented mean ± S.E.M. (n = 8), ++p < 0.01; +++p < 0.001 vs. naïve group; *p < 0.05 vs. CIA group. | ||
3.7. OLA diet modulates JAK/STAT signaling pathway
JAK/STAT pathway plays a crucial role in the inflammatory process of the synovium and bone destruction in RA31 and is activated by upstream cytokines, such as IL-6 and IL-17, among others.32 We evaluated pJAK-3 and pSTAT-3 protein expression by western blot from hind paw homogenates. Statistical analysis revealed a significant pJAK-3 and p-STAT-3 overexpression in CIA group when compared to the naïve control groups (pJAK-3: p < 0.001; pSTAT-3: p < 0.05 vs. naïve group) whereas nutritional therapy with OLA significantly suppressed both in arthritic CIA mice (pJAK-3: p < 0.001; pSTAT-3: p < 0.01 vs. CIA group) (Fig. 8). | ||
| Fig. 8 OLA supplemented diet regulated JAK/STAT signaling pathway in hind paws homogenates from arthritis induced mice. The expression of pJAK-3 and pSTAT-3 were measured by western blotting and normalized with respect to β-actin (housekeeping). Western blot images are representative of separate experiments with similar results. Naïve group: non-arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values represented mean ± S.E.M. (n = 8), +p < 0.05; +++p < 0.001 vs. naïve group; **p < 0.01; ***p < 0.001 vs. CIA group. | ||
3.8. Effect of dietary OLA on MAPKs signaling pathway
MAPKs signaling pathways play a key role in the establishment of inflammation process. We investigated the effects of dietary OLA treatment on MAPKs (JNK and p38) signaling pathway activation in CIA mice. We observed a significant increase on JNK and p38 phosphorylation in CIA control group in comparison with naïve control group (pJNK: p < 0.001; pp38 p < 0.01 vs. naïve group). Nevertheless, the phosphorylation degree of these MAPKs proteins was significantly ameliorated in those arthritic mice fed with the experimental diet assayed (pJNK: p < 0.001; pp38 p < 0.01 vs. CIA group) (Fig. 9). | ||
| Fig. 9 OLA diet modulated MAPKs signaling pathway in hind paws homogenates. Phosphorylated p38 and JNK was analyzed with western blot and normalized with respect to specific non-phosphorylated total proteins. Western blot images are representative of separate experiments with similar results. Naïve group: non-arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values represented mean ± S.E.M. (n = 8), ++p < 0.01; +++p < 0.001 vs. naïve group; **p < 0.01; ***p < 0.001 vs. CIA group. | ||
3.9. OLA diet modulates NF-κB signaling pathway
Finally, the effects of dietary OLA on NF-κB signaling pathway were also explored. As shown in Fig. 10, the expression of the inhibitory protein IκB-α was significantly reduced in CIA control group when compared with naïve control group (p < 0.01 vs. naïve group) which was accompanied by an over-phosphorylation of p65 protein (p < 0.05 vs. naïve group). On the contrary, dietary OLA treatment prevented the IκB-α degradation (p < 0.05 vs. CIA group) as well as the phosphorylation of p65 subunits (p < 0.05 vs. CIA group) (Fig. 10).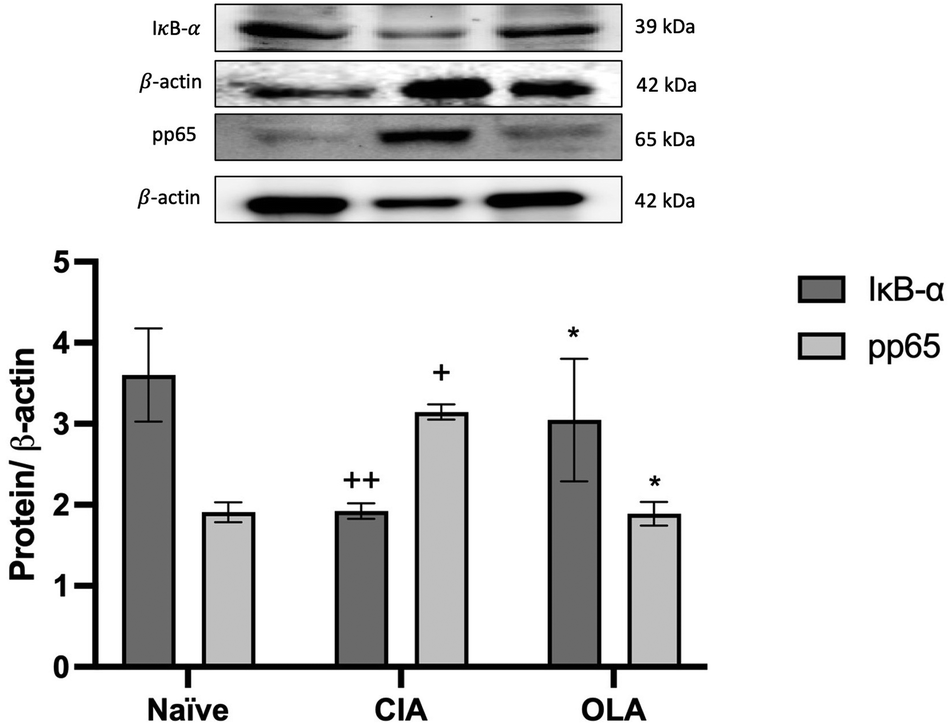 | ||
| Fig. 10 OLA diet-enriched controlled NF-κB signaling pathway preventing IκB-α degradation and p65 phosphorylation. The expression was quantified by densitometry analysis and normalized with respect to β-actin (housekeeping). Western blot images are representative of separate experiments with similar results. Naïve group: non-arthritic control animals fed with SD; CIA: control group induced arthritic animals fed with SD; OLA: induced arthritic animals fed with OLA enriched diet (0.025% w/w). Values represented mean ± S.E.M. (n = 8), +p < 0.05; ++p < 0.01 vs. naïve group; *p < 0.05 vs. CIA group. | ||
4. Discussion
Our findings have revealed, for the first time, the preventive role of OLA-enriched diet in the CIA murine model. Mice injected with collagen II developed arthritis similar in numerous features to human RA, characterized by progressive degradation of articular cartilage and bone erosion. However, dietary treatment with OLA resulted in an improvement of the signs and symptoms of the disease, this was accompanied by less histological damage as well as attenuating oxidative stress and inflammatory markers.The secretion of MMPs, cytokines and growth factors contribute to the loss of normal homeostasis in the synovial joint leading inflammation and joint damage on RA. Cytokines regulate the phenotype of effector and regulatory T-cells in the synovium. Therefore, an imbalance of cytokine network contributes to the development and progression of RA mainly an increase in the production of Th1 and Th17 cytokines and a decrease in the production of Th2 cytokines.33,34
Currently, Th17 cells have been established as having a close relationship with chronic inflammatory autoimmune diseases, as RA. Th17 cells produce cytokines such as IL-17, which stimulate joint osteoblasts, leading to the activation of receptor activator of nuclear factor-κB receptor ligand (RANKL). Subsequently, monocytes respond to RANKL and mature into osteoclasts.35 IL-17 also promotes the production of other crucial pro-inflammatoy cytokines, maily TNF-α, IL-1β and IL-6.36 In addition, TNF-α activates osteoclastogenesis, suppresses osteoblast recruitment and inhibits the expression of matrix genes.37 Besides, IL-6 produced by fibroblasts responding to IL-17 cells amplifies inflammation.38 Also, IL-6 stimulates synovial tissue in an autocrine manner, worsening the condition by amplification cycle that releases inflammatory mediators, including MMPs, induction synoviocites proliferation and osteoclast differentiation.35,39
On the other hand, the MMPs constitute a family of extracellular endopeptidases that selectively degrade the connective tissue extracellular matrix components.40 Of these, MMP-3 is a proteinase secreted by synovial fibroblasts and chondrocytes, and is responsible for the degradation of proteoglycan, various type of collagens and denatured type I and type II collagens, among others. Moreover, MMP-3 mediates the proteolytic activation of pro-MMP-9. MMP-9 promotes the recruitment of neutrophils, monocytes, T cells, and osteoclasts, also, the release of membrane- bound vascular endothelial growth factor (VEGF), thereby contributing to angiogenesis and disease progression.41
Our results are in agreement with those of previous studies and demonstrate that increased levels of Th-1 and Th-17 cytokines such as IL-1β, IL-6, IL-17, TNF-α, as well as MMP-3 and MMP-9 levels, were associated with disease onset and joint progression in CIA.42 Though, arthritic mice fed with OLA-enriched diets showed a significant reduction in serum MMP-3 levels, as well as in IL-17, TNF- α, IL-1β and IL-6 pro-inflammatory cytokines in paw homogenates, also less immunostaining for MMP-9 in tarsal and metatarsal joints sections in comparison with CIA control group, correlating in parallel with the macroscopic and histological findings. Then, these data suggest that dietary OLA exerts anti-inflammatory effects through the modulated production of these key RA biomarkers. Similar data were found in our recent study, which reported that OLA controlled the production of inflammatory mediators decreasing IL-17, TNF- α, IL-1β and IL-6 in LPS-stimulated murine peritoneal macrophages.17
Another identified biomarker of cartilage degradation is CatK, an enzyme involved in bone remodeling process and cartilage degradation. CatK is predominantly secreted by activated osteoclasts to degrade collagen and other matrix proteins during bone resorption and is overexpressed in osteoarthritic cartilage and inflamed synovial tissue.43,44 Our results showed an increased CatK immunosignal signal was extended to the subliming areas of synovium villi, being highly positive all along the inflammatory pannus in the joints of arthritic mice. However, in the OLA-supplemented group, only some cells of the synovial villi were immunopositive for CatK and, as in the control animals, chondrocytes and surface cells remained negative.
Numerous studies have reported an association between RA pathogenesis and ROS.45,46 Overproduction of NO contributes to the pathogenesis of chronic arthritis. Increased plasma nitrate/nitrite levels and increased NO production in synovial tissues induced by elevated iNOS expression have been observed in arthritis patients.47 Similarly, in collagen-induced rodent models of arthritis, overexpression of iNOS has been found in the paws of arthritic animals.9,48 On the other hand, COX-2 and mPGES-1, are enzymes responsible for the overproduction of PGE2 which is produced in response to proinflammatory cytokines through EP2/EP4 receptors, resulting in the modulation of proinflammatory cascades and suppression of osteoclastogenesis.49 However, dietary supplementation with OLA was able to reduce the overexpression of these oxidative and proinflammatory enzymes and the production of PGE2. Similar data have been observed with this secoiridoid in LPS-stimulated THP-1 macrophages, showing a reduction in COX-2, NO and PGE2 levels caused by LPS exposure. These data are also in agreement with those obtained with other olive bioactive compounds in particular: hydroxytyrosol acetate;22 the peracetylated derivative of oleuropein 9 and oleocanthal,48 among others. Therefore, the regulation of these oxidative and proinflammatory biomarkers by OLA could represent a new molecular target for OLA, in RA which has not been previously demonstrated.
On the other hand, signal transduction pathways closely involved in inflammation include the NF-κB, MAPKs, JAK-STAT pathways.5
The NF-κB signaling pathway is composed of NF-κB, NF-κB inhibitor (IκB), and IκB kinases (IKK).50 Highly activated NF-κB plays a pivotal role in the development and activation of Th-1 responses inducing the production of various pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6, thereby accelerating the progression of RA. Also, NF-κB nuclear transcription factor is in addition responsible to MAPKs for COX-2 and iNOs up-regulation.5,51
We investigated whether the activation of NF-κB signaling pathway was modulated by OLA diet. Our research showed that OLA increased the inhibitory protein IκB-α and reduced p65 phosphorylation, indicating that dietary OLA inhibited the NF-κB signaling pathway. Analogous results have been found with other olive bioactive compounds such as hydroxytyrosol acetate,22 the peracetylated derivative of oleuropein9 and oleocanthal,48 among others in arthritic mice. Additionally, our data are in agreement with Carpi et al., who demonstrated that OLA counteract obesity-associated adipocyte inflammation attenuating NF-κB activation in differentiated Simpson–Golabi–Behmel syndrome (SGBS) adipocytes52 and in lipopolysaccharide-Induced Inflammation in THP-1-derived macrophages.53
Similarly, MAPKs a family of serine/threonine protein kinases, have attracted considerable attention as potent therapeutic targets in RA. All three kinases families, JNK, p38 and extracellular signal-regulated kinase (ERK), are expressed in rheumatoid synovial tissue. They are involved in many important cell processes, mainly the regulation of cytokines and chemokines signaling, modulating the expression of several inflammation-related genes such, as iNOS, COX-2 and PGs.54
In fact, p38 and JNK MAPKs regulate MMP-3 activation in fibroblasts and osteoclast differentiation involved in cartilage destruction.55,56
Among the protein kinases of the JAK family, JAK1 and JAK3 play a particularly important role in the immune response, and their suppression may contribute to specific immune regulation.57 Besides, MAPKs phosphorylate the JAK-STAT pathway responsible for cytokine signaling leading to STAT activation and subsequent dimerization and translocation to the nucleus to bind to specific DNA sequences. STAT gene transcription is associated with persistent inflammation and severity of joint destruction in RA.8,58 Indeed, overexpression of STAT-3 has been found in synovial membranes of RA patients and contributes to chronicity in the CIA model of arthritis.8
Our findings are consistent with previous reports and show that phosphorylation of MAPKs: p38 and JNK, was increased in hind paw homogenates from CIA control mice.59 Similarly, overexpression of JAK-3 and STAT-3 proteins was also observed from arthritic mice that could be correlated with the severity of synovitis, while dietary supplementation of OLA significantly reduced both MAPKs and JAK-3/STAT-3 activation at the transcriptional level. Taken together, our data suggest that dietary OLA may ameliorate experimental arthritis by negatively interfering with MAPKs and JAK/STAT signaling pathways.
Nrf2 is a redox-sensitive transcription factor that controls the transcription of several antioxidant genes and therefore plays a critical role in pathologies involving chronic oxidative stress, including RA. Nrf2 deficiency has been documented to accelerate the effector phase of arthritis and aggravate joint disease. TNF-α, increased ROS levels, ERK and p38 MAPKs and NF-κB signaling pathways have also been described to regulate Nrf-2 activation in synovial tissues of arthritic patients. Nrf2 binds to antioxidant response elements (ARE) located in the promoter regions of detoxifying/antioxidant genes, such as HO-1. This enzyme confers protection against oxidative, inflammatory, and hypoxic stress in joint tissues by regulating synovial cell activation and metabolism.60,61 In the present study, in agreement with previous reports of our research group,8,9,22 Nrf2 and HO-1 expressions were decreased in the arthritis group. On the contrary, dietary OLA was able to recover Nrf2 and HO-1 expressions. These data are also consistent with those previously described in pristane-induced SLE mice treated with dietary OLA.19 Together, our results suggest that the protective antioxidant effect of OLA in this experimental model of CIA may be mediated through the upregulation of Nrf-2/HO-1 axis.
5. Conclusions
In summary, our study has revealed for the first time the immunomodulatory effects of dietary OLA in the CIA model, characterized by a significant reduction in serum synovial and cartilage biomarker MMP-3, also accompanied by a decrease in MMP-9 and CatK-positive cells in mice joints. Possible antioxidant and anti-inflammatory mechanisms responsible include, decrease of iNOs and COX-2 over expression, regulation of B-cell responses, activation of Nrf2/HO-1 axis and inhibition of signaling pathways such as JAK-STAT, MAPKs and NF-κB, modulating the production of inflammatory mediators, specifically, Th1 and Th17 cytokines and PGE2. It is concluded that OLA supplementation could be a new dietary strategy for nutritional therapy of RA.Author contributions
Conceptualization, C. A. L.; methodology, M. A. R.; experimentation contribution, I. V; product extraction, S. S. and J. O. V.; coated-pellet process, M. L. G. R.; histological and IHC assays, V. V. R. and J. M. F. S; software, validation and formal analysis, M. A. R.; investigation, M. A. R.; data curation, M. A. R.; writing – original draft preparation, M. A. R. and C. A. L.; writing – review and editing, C. A. L.; M. A. R. and I. V.; visualization and supervision, C. A. L. and I. V. All data were generated in-house, and no paper mill was used. All authors agree to be accountable for all aspects of work ensuring integrity and accuracy. All authors have read and agreed to the published version of the manuscript.Conflicts of interest
The authors declare no conflict of interest.Acknowledgements
The authors gratefully acknowledge the support provided by the research grant “US-1380592” (Consejería de Economía, Conocimiento, Empresas y Universidad, Junta de Andalucía, Spain). We thank the assistance of Center for Technology and Innovation Research, University of Seville (CITIUS), the Technical Scientific Instrumentation Center (CICT) of the University of Jaen and the Junta de Andalucía (CTS-259) for their financial support. M. A. Rosillo gratefully acknowledges the support provided by the from Talentos Doctores postdoctoral fellowship from the Junta de Andalucía.References
- P. Rahman, R. D. Inman, H. El-Gabalawy and D. O. Krause, Pathophysiology and Pathogenesis of Immune-Mediated Inflammatory Diseases: Commonalities and Differences, J. Rheumatol., Suppl., 2010, 85, 11–26 CrossRef.
- L. Puig, J. G. Ruiz de Morales, E. Dauden, J. L. Andreu, R. Cervera, A. Adan, S. Marsal, C. Escobar, J. Hinojosa, J. Palau, A. Arraiza, P. Casado, M. Codesido, C. Pascual, R. Saldana and A. Gil, Prevalence of ten Immune-mediated inflammatory diseases (IMID) in Spain, Rev. Esp. Salud. Publica, 2019, 93, e201903013 Search PubMed.
- A. Suzuki and K. Yamamoto, From genetics to functional insights into rheumatoid arthritis, Clin. Exp. Rheumatol., 2015, 33, S40–S43 Search PubMed.
- S. Paolino, G. Pacini, M. Patane, E. Alessandri, F. Cattelan, F. Goegan, C. Pizzorni, E. Gotelli and M. Cutolo, Interactions between microbiota, diet/nutrients and immune/inflammatory response in rheumatic diseases: focus on rheumatoid arthritis, Reumatologia, 2019, 57, 151–157 CrossRef PubMed.
- S. Liu, H. Ma, H. Zhang, C. Deng and P. Xin, Recent advances on signaling pathways and their inhibitors in rheumatoid arthritis, Clin. Immunol., 2021, 230, 108793 CrossRef PubMed.
- M. Benucci, P. Bernardini, C. Coccia, R. De Luca, J. Levani, A. Economou, A. Damiani, E. Russo, A. Amedei, S. Guiducci, E. Bartoloni, M. Manfredi, V. Grossi, M. Infantino and C. Perricone, JAK inhibitors and autoimmune rheumatic diseases, Autoimmun. Rev., 2023, 22, 103276 CrossRef PubMed.
- E. Nikiphorou and E. Philippou, Nutrition and its role in prevention and management of rheumatoid arthritis, Autoimmun. Rev., 2023, 22, 103333 CrossRef.
- M. A. Rosillo, M. Sanchez-Hidalgo, S. Sanchez-Fidalgo, M. Aparicio-Soto, I. Villegas and C. Alarcon-de-la-Lastra, Dietary extra-virgin olive oil prevents inflammatory response and cartilage matrix degradation in murine collagen-induced arthritis, Eur. J. Nutr., 2016, 55, 315–325 CrossRef PubMed.
- M. L. Castejon, C. Alarcon-de-la-Lastra, M. A. Rosillo, T. Montoya, J. G. Fernandez-Bolanos, A. Gonzalez-Benjumea and M. Sanchez-Hidalgo, A New Peracetylated Oleuropein Derivative Ameliorates Joint Inflammation and Destruction in a Murine Collagen-Induced Arthritis Model via Activation of the Nrf-2/Ho-1 Antioxidant Pathway and Suppression of MAPKs and NF-kappaB Activation, Nutrients, 2021, 13(2), 311 CrossRef CAS.
- T. Montoya, C. Alarcon-de-la-Lastra, M. L. Castejon, J. Ortega-Vidal, J. Altarejos and M. Sanchez-Hidalgo, (-)-Methyl-Oleocanthal, a New Oleocanthal Metabolite Reduces LPS-Induced Inflammatory and Oxidative Response: Molecular Signaling Pathways and Histones Epigenetic Modulation, Antioxidants, 2021, 11(1), 56 CrossRef PubMed.
- D. E. Trentham, A. S. Townes and A. H. Kang, Autoimmunity to type II collagen an experimental model of arthritis, J. Exp. Med., 1977, 146, 857–868 CrossRef CAS PubMed.
- A. Filipek, T. P. Mikolajczyk, T. J. Guzik and M. Naruszewicz, Oleacein and Foam Cell Formation in Human Monocyte-Derived Macrophages: A Potential Strategy Against Early and Advanced Atherosclerotic Lesions, Pharmaceuticals, 2020, 13(4), 64 CrossRef CAS.
- M. L. Castejon, T. Montoya, C. Alarcon-de-la-Lastra and M. Sanchez-Hidalgo, Potential Protective Role Exerted by Secoiridoids from Olea europaea L. in Cancer, Cardiovascular, Neurodegenerative, Aging-Related, and Immunoinflammatory Diseases, Antioxidants, 2020, 9(2), 149 CrossRef CAS.
- S. Cirmi, M. Celano, G. E. Lombardo, V. Maggisano, A. Procopio, D. Russo and M. Navarra, Oleacein inhibits STAT3, activates the apoptotic machinery, and exerts anti-metastatic effects in the SH-SY5Y human neuroblastoma cells, Food Funct., 2020, 11, 3271–3279 RSC.
- G. Juli, M. Oliverio, D. Bellizzi, M. E. Gallo-Cantafio, K. Grillone, G. Passarino, C. Colica, M. Nardi, M. Rossi, A. Procopio, P. Tagliaferri, P. Tassone and N. Amodio, Anti-tumor Activity and Epigenetic Impact of the Polyphenol Oleacein in Multiple Myeloma, Cancers, 2019, 11(7), 990 CrossRef CAS.
- G. E. Lombardo, S. M. Lepore, V. M. Morittu, B. Arcidiacono, C. Colica, A. Procopio, V. Maggisano, S. Bulotta, N. Costa, C. Mignogna, D. Britti, A. Brunetti, D. Russo and M. Celano, Effects of Oleacein on High-Fat Diet-Dependent Steatosis, Weight Gain, and Insulin Resistance in Mice, Front. Endocrinol., 2018, 9, 116 CrossRef PubMed.
- R. Munoz-Garcia, M. Sanchez-Hidalgo, T. Montoya, M. Alcarranza, J. Ortega-Vidal, J. Altarejos and C. Alarcon-de-la-Lastra, Effects of Oleacein, a New Epinutraceutical Bioproduct from Extra Virgin Olive Oil, in LPS-Activated Murine Immune Cells, Pharmaceuticals, 2022, 15(11), 1338 CrossRef CAS.
- B. Gutierrez-Miranda, I. Gallardo, E. Melliou, I. Cabero, Y. Alvarez, P. Magiatis, M. Hernandez and M. L. Nieto, Oleacein Attenuates the Pathogenesis of Experimental Autoimmune Encephalomyelitis through Both Antioxidant and Anti-Inflammatory Effects, Antioxidants, 2020, 9(11), 1161 CrossRef CAS.
- R. Munoz-Garcia, M. Sanchez-Hidalgo, M. Alcarranza, M. V. Vazquez-Roman, M. A. de Sotomayor, M. L. Gonzalez-Rodriguez, M. C. de Andres and C. Alarcon-de-la-Lastra, Effects of Dietary Oleacein Treatment on Endothelial Dysfunction and Lupus Nephritis in Balb/C Pristane-Induced Mice, Antioxidants, 2023, 12(6), 1303 CrossRef CAS.
- K. Vougogiannopoulou, C. Lemus, M. Halabalaki, C. Pergola, O. Werz, A. B. Smith 3rd, S. Michel, L. Skaltsounis and B. Deguin, One-step semisynthesis of oleacein and the determination as a 5-lipoxygenase inhibitor, J. Nat. Prod., 2014, 77, 441–445 CrossRef CAS PubMed.
- M. L. Castejon, M. Sanchez-Hidalgo, M. Aparicio-Soto, T. Montoya, I. Martin-LaCave, J. G. Fernandez-Bolanos and C. Alarcon-de-la-Lastra, Dietary oleuropein and its new acyl-derivate attenuate murine lupus nephritis through HO-1/Nrf2 activation and suppressing JAK/STAT, NF-kappaB, MAPK and NLRP3 inflammasome signaling pathways, J. Nutr. Biochem., 2019, 74, 108229 CrossRef CAS PubMed.
- M. A. Rosillo, M. Sanchez-Hidalgo, A. Gonzalez-Benjumea, J. G. Fernandez-Bolanos, E. Lubberts and C. Alarcon-de-la-Lastra, Preventive effects of dietary hydroxytyrosol acetate, an extra virgin olive oil polyphenol in murine collagen-induced arthritis, Mol. Nutr. Food Res., 2015, 59, 2537–2546 CrossRef CAS.
- M. A. Rosillo, M. Sanchez-Hidalgo, A. Cardeno, M. Aparicio-Soto, S. Sanchez-Fidalgo, I. Villegas and C. A. de la Lastra, Dietary supplementation of an ellagic acid-enriched pomegranate extract attenuates chronic colonic inflammation in rats, Pharmacol. Res., 2012, 66, 235–242 CrossRef CAS PubMed.
- M. M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem., 1976, 72, 248–254 CrossRef CAS.
- P. Costanzo, S. Bonacci, L. Cariati, M. Nardi, M. Oliverio and A. Procopio, Simple and efficient sustainable semi-synthesis of oleacein [2-(3,4-hydroxyphenyl) ethyl (3S,4E)-4-formyl-3-(2-oxoethyl)hex-4-enoate] as potential additive for edible oils, Food Chem., 2018, 245, 410–414 CrossRef CAS PubMed.
- J. Mason and S. Murphree, Microwave-Assisted Aqueous Krapcho Decarboxylation, Synlett, 2013, 1391–1394 CAS.
- O. Mijanovic, A. Jakovleva, A. Brankovic, K. Zdravkova, M. Pualic, T. A. Belozerskaya, A. I. Nikitkina, A. Parodi and A. A. Zamyatnin Jr., Cathepsin K in Pathological Conditions and New Therapeutic and Diagnostic Perspectives, Int. J. Mol. Sci., 2022, 23(22), 13762 CrossRef CAS.
- A. Paradowska-Gorycka, A. Grzybowska-Kowalczyk, E. Wojtecka-Lukasik and S. Maslinski, IL-23 in the pathogenesis of rheumatoid arthritis, Scand. J. Immunol., 2010, 71, 134–145 CrossRef CAS.
- A. Gul, B. Kunwar, M. Mazhar, S. Faizi, D. Ahmed, M. R. Shah and S. U. Simjee, Rutin and rutin-conjugated gold nanoparticles ameliorate collagen-induced arthritis in rats through inhibition of NF-kappaB and iNOS activation, Int. Immunopharmacol., 2018, 59, 310–317 CrossRef CAS.
- K. T. Tang, C. C. Lin, S. C. Lin, J. H. Wang and S. W. Tsai, Kurarinone Attenuates Collagen-Induced Arthritis in Mice by Inhibiting Th1/Th17 Cell Responses and Oxidative Stress, Int. J. Mol. Sci., 2021, 22(8), 4002 CrossRef CAS PubMed.
- S. A. Gonzalez-Chavez, S. M. Lopez-Loeza, S. Acosta-Jimenez, R. Cuevas-Martinez, C. Pacheco-Silva, E. Chaparro-Barrera and C. Pacheco-Tena, Low-Intensity Physical Exercise Decreases Inflammation and Joint Damage in the Preclinical Phase of a Rheumatoid Arthritis Murine Model, Biomolecules, 2023, 13(3), 488 CrossRef CAS PubMed.
- T. Thalhamer, M. A. McGrath and M. M. Harnett, MAPKs and their relevance to arthritis and inflammation, Rheumatology, 2008, 47, 409–414 CrossRef CAS.
- N. Komatsu and H. Takayanagi, Inflammation and bone destruction in arthritis: synergistic activity of immune and mesenchymal cells in joints, Front. Immunol., 2012, 3, 77 Search PubMed.
- A. Pradhan, A. Bagchi, S. De, S. Mitra, S. Mukherjee, P. Ghosh, A. Ghosh and M. Chatterjee, Role of redox imbalance and cytokines in mediating oxidative damage and disease progression of patients with rheumatoid arthritis, Free Radical Res., 2019, 53, 768–779 CrossRef CAS.
- T. Kuwabara, F. Ishikawa, M. Kondo and T. Kakiuchi, The Role of IL-17 and Related Cytokines in Inflammatory Autoimmune Diseases, Mediators Inflammation, 2017, 2017, 3908061 Search PubMed.
- C. A. Murphy, C. L. Langrish, Y. Chen, W. Blumenschein, T. McClanahan, R. A. Kastelein, J. D. Sedgwick and D. J. Cua, Divergent pro- and antiinflammatory roles for IL-23 and IL-12 in joint autoimmune inflammation, J. Exp. Med., 2003, 198, 1951–1957 CrossRef CAS PubMed.
- M. S. Nanes, Tumor necrosis factor-alpha: molecular and cellular mechanisms in skeletal pathology, Gene, 2003, 321, 1–15 CrossRef CAS PubMed.
- H. Ogura, M. Murakami, Y. Okuyama, M. Tsuruoka, C. Kitabayashi, M. Kanamoto, M. Nishihara, Y. Iwakura and T. Hirano, Interleukin-17 promotes autoimmunity by triggering a positive-feedback loop via interleukin-6 induction, Immunity, 2008, 29, 628–636 CrossRef CAS.
- J. Zwerina, K. Redlich, G. Schett and J. S. Smolen, Pathogenesis of rheumatoid arthritis: targeting cytokines, Ann. N. Y. Acad. Sci., 2005, 1051, 716–729 CrossRef CAS.
- D. Ahrens, A. E. Koch, R. M. Pope, M. Stein-Picarella and M. J. Niedbala, Expression of matrix metalloproteinase 9 (96-kd gelatinase B) in human rheumatoid arthritis, Arthritis Rheum., 1996, 39, 1576–1587 CrossRef CAS.
- M. M. Ally, B. Hodkinson, P. W. Meyer, E. Musenge, M. Tikly and R. Anderson, Serum matrix metalloproteinase-3 in comparison with acute phase proteins as a marker of disease activity and radiographic damage in early rheumatoid arthritis, Mediators Inflammation, 2013, 2013, 183653 Search PubMed.
- V. Babu, R. Singh, P. K. Kashyap, K. R. Washimkar, M. N. Mugale, S. Tandon and D. U. Bawankule, Pharmacological and Toxicological Study of Coumarinolignoids from Cleome viscosa in Small Animals for the Management of Rheumatoid Arthritis, Planta Med., 2023, 89, 62–71 CrossRef CAS PubMed.
- Y. T. Konttinen, J. Mandelin, T. F. Li, J. Salo, J. Lassus, M. Liljestrom, M. Hukkanen, M. Takagi, I. Virtanen and S. Santavirta, Acidic cysteine endoproteinase cathepsin K in the degeneration of the superficial articular hyaline cartilage in osteoarthritis, Arthritis Rheum., 2002, 46, 953–960 CrossRef CAS PubMed.
- J. Morko, R. Kiviranta, K. Joronen, A. M. Saamanen, E. Vuorio and H. Salminen-Mankonen, Spontaneous development of synovitis and cartilage degeneration in transgenic mice overexpressing cathepsin K, Arthritis Rheum., 2005, 52, 3713–3717 CrossRef CAS.
- M. J. Smallwood, A. Nissim, A. R. Knight, M. Whiteman, R. Haigh and P. G. Winyard, Oxidative stress in autoimmune rheumatic diseases, Free Radicals Biol. Med., 2018, 125, 3–14 CrossRef CAS PubMed.
- M. Veselinovic, N. Barudzic, M. Vuletic, V. Zivkovic, A. Tomic-Lucic, D. Djuric and V. Jakovljevic, Oxidative stress in rheumatoid arthritis patients: relationship to diseases activity, Mol. Cell. Biochem., 2014, 391, 225–232 CrossRef CAS.
- N. Kondo, T. Kanai and M. Okada, Rheumatoid Arthritis and Reactive Oxygen Species: A Review, Curr. Issues Mol. Biol., 2023, 45, 3000–3015 CrossRef CAS PubMed.
- T. Montoya, M. Sanchez-Hidalgo, M. L. Castejon, M. A. Rosillo, A. Gonzalez-Benjumea and C. Alarcon-de-la-Lastra, Dietary Oleocanthal Supplementation Prevents Inflammation and Oxidative Stress in Collagen-Induced Arthritis in Mice, Antioxidants, 2021, 10(5), 650 CrossRef CAS.
- J. Akaogi, T. Nozaki, M. Satoh and H. Yamada, Role of PGE2 and EP receptors in the pathogenesis of rheumatoid arthritis and as a novel therapeutic strategy, Endocr., Metab. Immune Disord.: Drug Targets, 2006, 6, 383–394 CrossRef CAS.
- M. S. Hayden and S. Ghosh, Shared principles in NF-kappaB signaling, Cell, 2008, 132, 344–362 CrossRef CAS PubMed.
- S. S. Makarov, NF-kappa B in rheumatoid arthritis: a pivotal regulator of inflammation, hyperplasia, and tissue destruction, Arthritis Res., 2001, 3, 200–206 CrossRef CAS PubMed.
- S. Carpi, E. Scoditti, M. Massaro, B. Polini, C. Manera, M. Digiacomo, J. Esposito Salsano, G. Poli, T. Tuccinardi, S. Doccini, F. M. Santorelli, M. A. Carluccio, M. Macchia, M. Wabitsch, R. De Caterina and P. Nieri, The Extra-Virgin Olive Oil Polyphenols Oleocanthal and Oleacein Counteract Inflammation-Related Gene and miRNA Expression in Adipocytes by Attenuating NF-kappaB Activation, Nutrients, 2019, 11(12), 2855 CrossRef PubMed.
- S. Cirmi, A. Maugeri, C. Russo, L. Musumeci, M. Navarra and G. E. Lombardo, Oleacein Attenuates Lipopolysaccharide-Induced Inflammation in THP-1-Derived Macrophages by the Inhibition of TLR4/MyD88/NF-kappaB Pathway, Int. J. Mol. Sci., 2022, 23(3), 1206 CrossRef CAS.
- Z. Han, D. L. Boyle, L. Chang, B. Bennett, M. Karin, L. Yang, A. M. Manning and G. S. Firestein, c-Jun N-terminal kinase is required for metalloproteinase expression and joint destruction in inflammatory arthritis, J. Clin. Invest., 2001, 108, 73–81 CrossRef CAS.
- R. K. Chauhan, P. K. Sharma and S. Srivastava, The Role of Signaling Pathway in the Biological Cause of Rheumatoid Arthritis, Curr. Drug Res. Rev., 2021, 13, 130–139 CrossRef CAS PubMed.
- T. Kanai, N. Kondo, M. Okada, H. Sano, G. Okumura, Y. Kijima, A. Ogose, H. Kawashima and N. Endo, The JNK pathway represents a novel target in the treatment of rheumatoid arthritis through the suppression of MMP-3, J. Orthop. Surg. Res., 2020, 15(1), 87 CrossRef PubMed.
- P. Xin, X. Xu, C. Deng, S. Liu, Y. Wang, X. Zhou, H. Ma, D. Wei and S. Sun, The role of JAK/STAT signaling pathway and its inhibitors in diseases, Int. Immunopharmacol., 2020, 80, 106210 CrossRef CAS PubMed.
- D. S. Aaronson and C. M. Horvath, A road map for those who don't know JAK-STAT, Science, 2002, 296, 1653–1655 CrossRef CAS PubMed.
- M. A. Rosillo, M. J. Alcaraz, M. Sanchez-Hidalgo, J. G. Fernandez-Bolanos, C. Alarcon-de-la-Lastra and M. L. Ferrandiz, Anti-inflammatory and joint protective effects of extra-virgin olive-oil polyphenol extract in experimental arthritis, J. Nutr. Biochem., 2014, 25, 1275–1281 CrossRef CAS PubMed.
- G. Manda, E. Milanesi, S. Genc, C. M. Niculite, I. V. Neagoe, B. Tastan, E. M. Dragnea and A. Cuadrado, Pros and cons of NRF2 activation as adjunctive therapy in rheumatoid arthritis, Free Radicals Biol. Med., 2022, 190, 179–201 CrossRef CAS PubMed.
- I. Rodriguez-Ramiro, S. Ramos, L. Bravo, L. Goya and M. A. Martin, Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress, Eur. J. Nutr., 2012, 51, 881–892 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2024 |