
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAdministration time effect of dietary proanthocyanidins on the metabolome of Fischer 344 rats is sex- and diet-dependent†
Verónica
Arreaza-Gil‡
 a,
Héctor
Palacios-Jordan‡
ab,
María del Mar
Romero
cd,
Cristina
Torres-Fuentes
a,
Miguel A.
Rodríguez
b,
Xavier
Remesar
c,
José-Antonio
Fernández-López
cd and
Anna
Arola-Arnal
*ac
a,
Héctor
Palacios-Jordan‡
ab,
María del Mar
Romero
cd,
Cristina
Torres-Fuentes
a,
Miguel A.
Rodríguez
b,
Xavier
Remesar
c,
José-Antonio
Fernández-López
cd and
Anna
Arola-Arnal
*ac
aNutrigenomics Research Group, Department of Biochemistry and Biotechnology, Universitat Rovira i Virgili, 43007 Tarragona, Spain. E-mail: anna.arola@urv.cat
bEurecat, Centre Tecnològic de Catalunya, Centre for Omic Sciences (COS), Joint Unit Univeristat Rovira i Virgili-EURECAT, Unique Scientific and Technical Infrastructures (ICTS), 43204 Reus, Spain
cDepartment of Biochemistry and Molecular Biomedicine, Universitat de Barcelona, Barcelona, Spain
dCIBER Obesity and Nutrition, Institute of Health Carlos III, Av. Diagonal 643, 08028 Barcelona, Spain
First published on 11th July 2023
Abstract
Proanthocyanidins (PAs) are one of the most commonly ingested polyphenols in the human diet, with a wide range of beneficial health effects. Remarkably, PAs have been reported to influence core and peripheral clock genes expression, and their effects may change in a time-of-day dependent manner. Therefore, the aim of this study was to investigate whether the capacity of PAs to modulate the metabolome is conditioned by the time-of-day in which these compounds are consumed in a diet- and sex-dependent manner. To do this, a grape seed proanthocyanidin extract (GSPE) was administered to female and male Fischer 344 rats at ZT0 (in the morning) and ZT12 (at night) and the GSPE administration time effect was evaluated on clock genes expression, melatonin hormone and serum metabolite levels in a healthy and obesogenic context. The results showed an administration time effect of GSPE on the metabolome in a sex and diet-dependent manner. Specifically, there was an effect on amino acid, lipid and cholate metabolite levels that correlated with the central clock genes expression. Therefore, this study shows a strong influence of sex and diet on the PAs effects on the metabolome, modulated in turn by the time-of-day.
Introduction
Biological rhythms of organisms allow the adjustment of physiological and metabolic processes to daily environmental changes in order to maintain a proper energy homeostasis and a good health state.1 Among the types of biological rhythms, circadian rhythms play an important role in the regulation of several biological functions, including metabolism, endocrine system, blood pressure, gastrointestinal tract or sleep–wake cycles, in 24-hour periods.2,3Circadian rhythms are driven by central clock genes, found in the hypothalamic suprachiasmatic nucleus (SCN), and by peripheral clock tissues.4 These molecular clocks follow 24-hour cycles and consist of an autoregulatory negative transcription-translation feedback loop formed by the brain and muscle aryl hydrocarbon receptor nuclear translocator-like 1 (BMAL1) and the circadian locomotor output cycles kaput (CLOCK), which after heterodimerization activate period (Per) and cryptochrome (Cry) gene transcription. The protein products PER and CRY repress the activity of BMAL1/CLOCK dimer.5Bmal1 gene is also regulated by the inhibitor nuclear receptors REV-ERB α and β and the activator RAR-orphan receptor α and γ (RORα and -γ).6 Moreover, BMAL1/CLOCK controls the transcription of additional clock-controlled genes (CCGs) involved in numerous cellular processes.7 The central clocks synchronize the peripheral clocks by neuronal mechanisms and hormonal signals, including melatonin that indicates the time of environmental darkness,8,9 although peripheral clocks in turn are also capable of local and autonomous regulation.8 In this context, light is the main external synchronizer or zeitgeber of diurnal rhythmicity in the organism,10 but others such as diet or feeding pattern also can modulate and reprogram the diurnal oscillation of the body.11–13 Nevertheless, misaligning cues like obesogenic diets can disrupt the clock gene expressions, triggering the development of metabolic disorders.14 It is important to point out that circadian rhythms influence metabolic processes according to sex,15 and in turn their misalignment affect differently to men and women. Like this, the alteration of diurnal oscillations by shift works, promoted a higher disturbance in the energy homeostasis processes in women compared to men.16,17 Another similar study in shift workers showed increased proinflammatory gut bacteria in women in comparison with men, may leading to higher rates of obesity.16,17 Furthermore, in a mice study, Pérez-Mendoza et al.18 reported that hepatic BMAL1 expression was more altered in high fat diet-fed (HFD) female mice than in male mice, impacting differently on lipid metabolism.
Therefore, circadian rhythms are key factors in the regulation of a great number of metabolic pathways. For instance, diurnal oscillations of insulin and glucagon levels regulate glucose homeostasis.19,20 Triglycerides follow a circadian regulation achieved through the rhythmic gene expression pattern of enzymes involved in fatty acid elongation and synthesis.21,22 Similarly, cholesterol synthesis in the liver exhibits diurnal rhythmicity trough the rhythmic expression of β-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA).23 Additionally, bile acids synthesis shows a well-known rhythmicity, both in rodents and humans, by modulation of the expression of the enzyme cholesterol 7α-hydroxylase (CYP7A1).24,25 Finally, although the relationship between amino acid metabolism and circadian rhythms is less understood, it appears that circulating amino acid levels vary according to the time of the day. Thus, branched-chain amino acids (BCAAs), which are important modulators of metabolism and metabolic health, undergo diurnal oscillations via Krüppel-like factor 15 (KLF15) in muscle.26
The rhythmicity of these metabolic pathways can therefore be reflected in the concentration of metabolites in serum and investigated by metabolomics approaches. In fact, most serum metabolites show diurnal rhythmicity under normal physiological conditions.27 However, nutritional challenges like the consumption of hypercaloric diets induce a loss of serum metabolite rhythmicity, leading to the development of obesity and metabolic-related disorders.28
Proanthocyanidins (PAs) are the most abundant flavonoids in the human diet with numerous beneficial health effects,29–31 as epidemiological studies have reported.32,33 In particular, our group has extensively studied, in animal models, the effects of a grape seed proanthocyanidin-rich extract (GSPE), establishing that GSPE can effectively prevent obesity by improving lipid metabolism,34 insulin resistance,35 and decreasing oxidative stress pathways.36 Moreover, in a human clinical trial in hypercholesterolemic subjects, GSPE supplementation significantly reduced oxidized LDL, a biomarker of cardiovascular diseases, and significantly inhibited CD36 expression, a novel cardioregulatory gene, demonstrating that GSPE is a potential therapeutic tool.37 These effects are carried out through different mechanism of action: GSPE activates antioxidant enzymes such as catalase or superoxide dismutase, decreasing reactive oxygen species (ROS); efficiently regulates the activity of nuclear factor-κB (NF-κB), modulating pro-inflammatory processes; induces epigenetic changes such as DNA methylation, histone modification or regulates miRNAs; and can modulate the gut microbiota composition.29 Regarding to this last point, it is worth mentioning that the relationship between GSPE and gut microbiota is bidirectional, and that the gut microbiota undergo extensive transformation of the phenolic compounds, driving the metabolized forms most of effects of PAs.38,39 Remarkably, GSPE has also been reported as a key modulator of clock genes, being this another possible mechanism of action.40–42 Accordingly, GSPE has been found to modulate the expression of central and peripheral clock genes in healthy and obese rats,40,42 as well as to exert an effect on plasma melatonin levels in healthy rats.41 Moreover, it has been recently showed that GSPE effects in liver and adipose tissue depend on the time of administration,43,44 and that the bioavailability of GSPE in plasma is also significantly impacted by the time of its administration.45 Hence, the relationship between GSPE and circadian rhythms seems to be bidirectional since GSPE influence clock genes, but circadian rhythms also can affect its functionality. In this context, GSPE could be a potential nutraceutical that would prevent metabolic disorders depending on the time of day it is ingested, by adjusting the biological oscillation of clock genes and modulating melatonin levels, promoting good synchronization with light/dark cycles, and these changes are measurable by metabolic approaches. It is important to note that the beneficial effects of GSPE on metabolic disorders could also vary according to sex, since the circadian misalignment observed in metabolic-related diseases, influences men and women differently, as discussed above.
Therefore, the aim of this study was to investigate whether the capacity of PAs to modulate metabolism is conditioned by the time-of-day in which this compound is consumed. To this goal, the effect of GSPE administration time on central clock gene expression and serum metabolite levels was evaluated in a healthy and an obesogenic context, and in female and male rats to account for the significant sex differences in circadian rhythms.
Materials and methods
Grape seed proanthocyanidin extract (GSPE)
GSPE was provided by Les Dérives Résiniques et Terpéniques (Dax, France). According to the manufacturer, GSPE contains 21.3% of flavan-3-ols monomers, a 17.4% of dimers, a 16.3% of trimers, a 13.3% of tetramers and a 31.7% of oligomers (5–13 units). Table 1 shows the phenolic compound concentrations of GSPE analyzed by HPLC-MS/MS.46| Compound concentration (mg g−1) | Compound concentration (mg g−1) |
|---|---|
| Table adapted from Margalef et al. 2016.46 The results are expressed as mg of phenolic compound/g of GSPE (mean ± SD). PA: proanthocyanin | |
| Gallic acid | 31.07 ± 0.08 |
| Protocatechuic acid | 1.34 ± 0.02 |
| Vanillic acid | 0.77 ± 0.04 |
| PA dimer B2 | 33.24 ± 1.39 |
| PA dimer B1 | 88.80 ± 3.46 |
| PA dimer B3 | 46.09 ± 2.07 |
| Catechin | 121.32 ± 3.41 |
| Epicatechin | 93.44 ± 4.27 |
| Dimer gallate | 8.86 ± 0.14 |
| Epicatechin gallate | 21.24 ± 1.08 |
| Epigallocatechin gallate | 0.03 ± 0.00 |
| Epigallocatechin | 0.27 ± 0.03 |
| PA trimer | 4.90 ± 0.47 |
| PA tetramer | 0.05 ± 0.01 |
Animal experimental procedure
Eight-week-old female (n = 64) and male (n = 64) Fischer 344 rats (Charles River Laboratories, Barcelona, Spain) were housed in pairs in separated animal quarters at 22 °C under a light/dark cycle of 12/12 h with ad libitum access to water and food. After one week of adaptation to the facilities, animals were randomly divided into two dietary groups: (1) standard chow diet-fed group (STD) (n = 32 female rats and n = 32 male rats). The diet was composed by 72% kcal derived from carbohydrates (CH), 8% kcal derived from lipid, and 19% kcal derived from protein (Safe-A04c, Rosenberg, Germany); and (2) cafeteria-fed group (CAF) (n = 32 female rats and n = 32 male rats). CAF diet (58% kcal derived from CH, 31% kcal derived from lipid and 11% kcal derived from protein) was freshly prepared daily and included bacon biscuits with pâté, biscuits with cheese, ensaimada pastry, carrot, sweetened milk (20% sucrose w/v) and standard chow diet.After 5 weeks of dietary feeding, female and male rats in each dietary group were further randomly divided into four groups according to the GSPE administration time (n = 8). Thus, during the last 4 weeks of the experiment, animals were administered with a daily oral dose of GSPE (25 mg per kg of body weight (BW)) dissolved in vehicle (VH, condensed milk diluted in water (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :4 v:v)) when the light was turned on (zeitgeber time (ZT) 0; ZT0) or when the light was turned off (ZT12). In addition, VH-administered animals were included as controls. The experimental design is described in Fig. 1.
:4 v:v)) when the light was turned on (zeitgeber time (ZT) 0; ZT0) or when the light was turned off (ZT12). In addition, VH-administered animals were included as controls. The experimental design is described in Fig. 1.
 | ||
| Fig. 1 Animal experimental design. 8-week-female and male Fischer 344 rats were pair-housed and fed with STD or CAF diet for 9 weeks. During the last 4 weeks, rats were daily treated with an oral dose of GSPE (grape seed proanthocyanidin extract) (25 mg kg−1) dissolved in a solution of water and condensed milk (4:1, VH) either when the light was on (ZT0) or when light was off (ZT12). Animals administered with vehicle (VH) were included as controls. Sacrifice was carried out at either ZT3 or ZT15 according to the dose time administration. STD: standard chow diet; CAF: cafeteria diet. | ||
BW was recorded weekly in all groups throughout the experimental procedure. At the end of the experiment, animals were fasted for 3 hours after administration of the last dose and sacrificed by decapitation. Thereby, animals administered at ZT0 were sacrificed 3 hours after light was turned on (ZT3) and animals administered at ZT12 were sacrificed 3 hours after light was turned off (ZT15). Blood was collected from the neck and serum was obtained by centrifugation (1500g, 20 min, 4 °C). Hypothalamus samples were collected, frozen immediately in liquid nitrogen and stored at −80 °C until RNA extraction. The Animal Ethics Committee of the Universitat Rovira i Virgili (Tarragona, Spain) and the Generalitat de Catalunya approved all the procedures (license number 9495) in accordance with the EU Directive 2010/63/UE for animal.
RNA isolation and quantitative real-time PCR
Total RNA from hypothalamus samples was isolated using TriPure reagent (Roche Applied Science, Indianapolis, IN, USA) following the manufacturer's protocol. RNA concentration and quality were determined by spectrophotometry (ND-1000; Nanodrop Technologies, Wilmington, DE, USA). Reverse transcription was performed using the MMLV reverse transcriptase system (Promega, Madison, WI, USA). Real-time PCR (RT-PCR) analysis was carried out using Power SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA). The primer sequences used to amplify target gene are showed in Table 2. The cycling protocol used was 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 1 min and a final step of 95 °C for 15 s, 60 °C for 15 s and 95 °C for 15 s. Cycle threshold (Ct) values were recorded by using the ABI PRISM 7900 HT detection system (Applied Biosystems, Foster City, CA, USA) and transformed to relative gene expression values using the 2−ΔΔCt method.47 Rat peptidylprolyl isomerase a (Ppia) was used as a housekeeping gene.| Target | Forward 5′–3′ | Reverse 3′–5′ |
|---|---|---|
| Ppia: peptidylprolyl isomerase a; Bmal1: brain and muscle aryl hydrocarbon receptor nuclear translocator-like; Cry: Cryptochrome; Clock: circadian locomotor output cycles kaput; Per1/2: period circadian regulator 1/2; RORα: Orphan nuclear receptor α. | ||
| Ppia | CTTCGAGCTGTTTGCAGACAA | AAGTCACCACCCTGGCACATG |
| Bmal1 | GACTTCGCCTCCACCTGTTC | TCTTCGTCCAGCCCCATCTT |
| Cry | GTTGCCTGTTTCCTGACCCG | ATTGATGCTCCAGTCGGCGT |
| Clock | ACTCCTTCTGCCTCCTCCAG | TCCGCTGTGTCATCTTCTCA |
| Per1 | AACAACAGCCACGGTTCTCA | GCTACCACAGTCCACACAAGC |
| Per2 | TGGAGCAGGTTGAGGGCATT | GGGACACAGCCACAGCAAAC |
| RORα | GGCTTCTTCCCCTACTGTTCC | ATTGGCAGGTTTCCAGGTGC |
Melatonin analysis
Melatonin serum levels were analysed by enzyme-linked immunosorbent assay (ELISA) kit (DRG, Marburg, Germany) according to the manufacturer's instructions.Serum samples extraction for 1H NMR-based Metabolomics assays
Hydrophilic and lipophilic metabolites from serum samples were analysed by untargeted proton nuclear magnetic resonance (1H NMR). To obtain hydrophilic metabolites, 0.2 mL of serum was homogenized with a methanol:water (8:1) solution by vortexing. For lipophilic metabolites extraction, 0.1 mL of serum was homogenized with a solution of methanol:methyl tert-butyl ether:water (323:1077:180) by vortexing. All homogenates obtained were centrifuged at 15000g for 30 min at 4 °C and the supernatants were collected and dried in speed vac using nitrogen stream. Hydrophilic metabolites were reconstituted with a solution of deuterated PBS buffer (0.05 mM) and trimethylsilylpropanoic acid (TSP) (0.73 mM). Lipophilic metabolites were reconstituted with a solution of deuterated chloroform:methanol (2:1) and tertramethylsilane (TMS).
1H NMR analysis
1H NMR measurements of the hydrophilic and lipophilic extracts were performed following the protocol previously published by Palacios-Jordan et al.48 Briefly, spectra were recorded at 300 K on an Avance III 600 spectrometer (Bruker®, Germany) operating at a proton frequency of 600.20 MHz and using a 5 mm PABBO gradient probe. For metabolite identification, all acquired 1H NMR spectra were compared with pure compound references from the AMIX metabolic profiling spectra database (Bruker®), HMDB, and Chenomx NMR suite 8.4 software (Chenomx Inc., Edmonton, AN, Canada). Metabolites were assigned by 1H–1H homonuclear (COSY and TOCSY) and 1H–13C heteronuclear (HSQC) correlation in 2D NMR experiments and by correlation with pure compounds performed in-house. After pre-processing, the specific 1H NMR regions identified in the spectra were integrated using the AMIX 3.9 software package and a data matrix with metabolite concentrations was obtained.Statistical analysis
Data are shown as mean ± standard deviation (SD) and were plotted using Graphpad Prism 8.0 software (Graphpad software Inc, San Diego, CA, USA). Statistical analyses were carried out using SPSS software (IBS SPSS statistics 25, Chicago, IL, USA). Normality and homogeneity of variance were evaluated by Shapiro–Wilk and Levene's test respectively. BW gain throughout the experiment was analyzed by repeated measures ANOVA followed by LSD post hot test at each individual time point. Area under the curve (AUC) of BW gain and gene expressions were analyzed by 4-way analysis of variance (ANOVA) to evaluate sex, diet, GSPE and time of day (ZT) effects followed by LSD post hot test, as well as by 2-way ANOVA to better evaluate GSPE and ZT effects followed by LSD post hot test.Metabolomic profiles were analyzed by 4-way ANOVA to assess the sex, diet, GSPE and ZT effects and their interactions, as well as by 2-way ANOVA to better evaluate GSPE and ZT effects in each condition (Table S1†). When one or more main effects were statistically significant (p < 0.05), one-way ANOVA followed by LSD post hot test was used to determinate the differences between groups. To control family-wise type I error rate, p-values were corrected for multiple comparisons with the false discovery rate (FDR) method (p < 0.05). Principal component analysis (PCA) and heatmaps involving metabolite levels data were analyzed and plotted using MetaboAnalyst v.5.049 to determine the influence of different factors on the metabolic profiles though a multivariate approach.
Pearson's rank-order correlation analysis between gene expression and metabolite levels were carry out using Python script (PyCharm software v.2018.2.4, JetBrains s.r.o., Prague, Czech Republic) as it was previously described by our group.50
Results
Body weight changes
CAF-fed rats showed a higher BW gain and corresponding AUC compared to STD-fed rats from the second week of the experiment. However, no effect of GSPE or time-of-day was observed on the BW gain (Fig. S1†).GSPE administration time modulates hypothalamus clock genes depending on sex and diet
STD-fed females receiving VH at ZT12 and sacrificed at ZT15 showed a significant decrease of Bmal1 and an increase of Per1 and Per2 expression levels compared to those rats receiving VH at ZT0 and sacrificed at ZT3, showing these genes a time-of-day effect (Fig. 2a, c and d). However, this effect was not observed in Clock, Cry and RORα gene expressions (Fig. 2b, e and f). Interestingly, when GSPE was administered to STD-fed female rats, expressions of Clock, Per1 and Per2 were increased only at night (i.e., only at ZT12 compared to their respective control) (Fig. 2b–d). Moreover, CAF altered the expression of these genes in female rats in a time-of-day dependent manner, increasing the levels of Per2 and Cry in VH-administered rats at ZT0 and sacrificed at ZT3, while only Clock and Per2 gene expression increased in VH-administered rats at ZT12 and sacrificed at ZT15 (Fig. 2b, d and e). Additionally, CAF-fed female rats treated with GSPE at ZT0 showed lower expression levels of Bmal1, Clock, Per2 and RORα compared to STD-fed rats treated with GSPE at the same time (Fig. 2a, b, d and f). Remarkably, in contrast to STD-fed female rats, GSPE decreased the expression of Bmal1 and Cry when administered at ZT0 in CAF-fed female rats (Fig. 2a and e). Thus, GSPE effect on CAF-fed female rats was higher in the morning in comparison with STD-fed rats, which exhibited more GSPE effects at night, suggesting that the GSPE effects depend not only on the time of administration but also on the diet.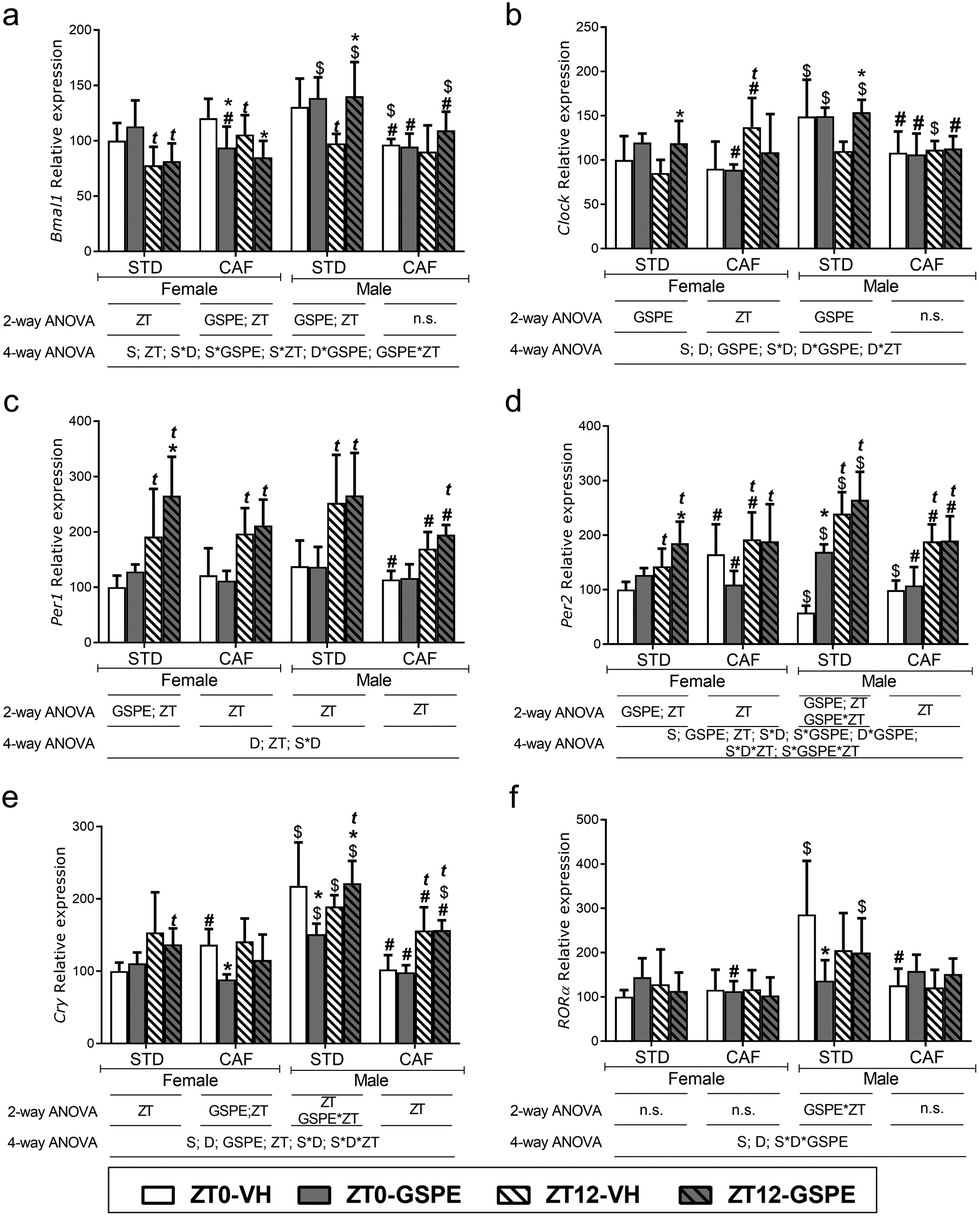 | ||
| Fig. 2 Time-of-day effect and GSPE administration time effects on hypothalamus clock genes expression. mRNA relative expression of (a) Bmal1; (b) Clock; (c) Per1; (d) Per2; (e) Cry and (f) RORα. Table below the graphs indicates significant effect and interactions among the different factors (D: diet; GSPE; ZT; S: sex) analyzed by 2-way or 4-way ANOVA followed by LSD post hoc test: *, t, # and $ indicates significant GSPE, ZT, diet and sex effect respectively (p < 0.05). Data are showed as mean ± SD (n = 6). STD: standard diet; CAF: cafeteria diet; GSPE: grape seed proanthocyanidin extract; ZT: time of the day; ZT0: VH or GSPE time administration (when light was turned on); ZT12: VH or GSPE time administration (when light was turned off); n.s: no significant differences (p > 0.05). | ||
The clock gene expressions of STD-fed males that received VH at ZT12 showed the same pattern than STD-fed females. CAF influenced the expression of clock genes by promoting, in males, the loss of the time-of-day effect in Bmal1, Clock and Per1 (Fig. 2a–c). In contrast, Per2 showed the same time effect that STD-fed males and Cry expression levels increased in rats receiving VH at ZT12 (Fig. 2d and e).
Remarkably, an overall sex effect was found on the hypothalamus clock genes, being this effect higher in STD-fed rats. In this context, when comparing STD-fed males versus STD-fed females, Bmal1 expression increased only in those rats treated with GSPE, independently of the time of administration (Fig. 2a), clock expression was higher in all STD-fed males groups, except for STD-VH-ZT12, which did not show sex differences, Per2 was increased in all STD-fed males, except for STD-VH-ZT0 males (Fig. 2b and d), Cry expression increased in all STD-fed groups, and Per1 and RORα expression only increased in the STD-VH-ZT0 group (Fig. 2c, e and f). This sex effect was lower in CAF-fed rats, and although CAF feeding in males decreased the clock gene expressions compared to STD-fed males, these levels were like those observed in CAF-fed female rats.
The GSPE effects are also influenced by sex. When GSPE was administered to STD-fed females, the extract effect was only observed at night. However, when GSPE was administered to STD-fed males, there was no clear pattern compared to STD-fed females. Thus, GSPE administration to STD-fed males resulted in increased expression levels of Bmal1 and Cry at ZT12 and Per2 at ZT0, while RORα and Cry expression levels decreased at ZT0 (Fig. 2a–f). Moreover, CAF-fed males did not show any GSPE effect at either ZT0 or ZT12 in contrast to CAF-fed females, which showed a higher GSPE effect in the morning. Therefore, GSPE influenced hypothalamus clock genes in a diet and sex-dependent manner.
Melatonin levels are affected by sex in a time-of-day-dependent manner, but not by GSPE treatment
Female rats showed a time-of-day effect on melatonin, independently of diet, increasing at ZT15 (at night) in both STD- and CAF-fed female rats (Fig. 3). An effect of diet on this hormone was also observed. Melatonin levels were higher in cafeteria-induced obese female rats at ZT15 compared to STD-fed female rats. No GSPE effect was observed in female rats (Fig. 3). | ||
| Fig. 3 Time-of-day effect and GSPE administration time effects on serum melatonin levels. Table below the graphs indicates significant effect and interactions among the different factors (D: diet; GSPE; ZT; S: sex) analysed by 2-way or 4-way ANOVA followed by LSD post hoc test: t, # and $ indicates significant ZT, diet and sex effect respectively (p < 0.05). Data are showed as mean ± SD (n = 7–8). STD: standard diet; CAF: cafeteria diet; GSPE: grape seed proanthocyanidin extract; time of day; ZT0: VH or GSPE time administration (when light was turned on); ZT12: VH or GSPE time administration (when light was turned off); n.s: no significant differences (p > 0.05). | ||
In addition, a significant sex effect was also found on melatonin levels, being higher in STD-fed males at ZT15 compared to STD-females. Interestingly, sex effect was not observed in melatonin levels in CAF-fed rats.
Regarding the time-of-day effect in males, melatonin levels showed the same pattern than those observed in females. Melatonin levels were higher at night (ZT15) in both STD- and CAF-fed males. In contrast to CAF-fed females, CAF did not increase melatonin levels in males compared to STD-fed males. However, GSPE also did not influence melatonin levels in STD- and CAF-fed males.
GSPE effects on metabolic serum profile depend on the time of administration in a sex-specific manner in standard-fed rats
CAF feeding and sex showed an impact on serum metabolic profiles. Two main clusters were found according to the sex of animals, and in turn, these were divided into two other clusters according to the type of diet. Males fed a CAF showed higher levels of lipid metabolites such as linoleic acid, monounsaturated fatty acids (MUFAs), triglycerides, oleic acid, total phospholipid, and polyunsaturated fatty acids (PUFAs), compared to both STD- and CAF-fed females (Fig. 4a). Remarkably, animals also clustered by the time of day, but no clusters were observed by GSPE administration. | ||
| Fig. 4 Effects of GSPE administration time on metabolic serum profile. (a) Heatmap of serum metabolic profiles. (b–e) Principal component analysis (PCA) showing GSPE and dose time effect in (b) STD-fed females; (c) CAF-fed females; (d) STD-fed males; and (e) CAF-fed males. STD: standard diet; CAF: cafeteria diet; GSPE: grape seed proanthocyanidin extract; time of the day; ZT0: VH or GSPE administration time (when light was turned on); ZT12: VH or GSPE administration time (when light was turned off). | ||
In addition, overall serum metabolic profiles were assessed by PCA in each diet condition and sex to further evaluate the GSPE effect on the metabolome depending on the time of administration (Fig. 4b–e). STD-fed females showed a time effect, and only rats receiving VH at ZT0 (in the morning) were clustered separately. Interestingly, STD-fed females administered with GSPE at ZT0 showed a similar metabolic profile to those administered with VH or GSPE at ZT12 (at night), suggesting that GSPE influenced the metabolic profile of STD-fed female rats mostly when it was administered in the morning (Fig. 4b). However, CAF-fed female rats did not show a defined clustering, so neither time-of-day nor GSPE effects were observed (Fig. 4c). Similarly to female rats, STD-fed males showed a time-of-day effect, clustering separately rats that received VH at ZT0 from those receiving VH at ZT12. However, in this case, GSPE administration at both ZT0 and ZT12 changed the metabolic profile of these rats, clustering separately from controls administered with VH at both ZT0 and ZT12, respectively (Fig. 4d). Moreover, CAF male rats, as in the case of CAF females, did not show time-of-day or GSPE effect since defined clusters were not observed (Fig. 4e).
GSPE modulates amino acid serum profile depending on the time of administration in a sex- and diet-dependent manner
Firstly, a sex and diet effect was observed in amino acids levels in a time of day-dependent manner: tyrosine (Tyr), phenylalanine (Phe), valine (Val), glycine (Gly) and glutamine (Gln) levels were different according to sex, showing STD-fed male higher levels of these amino acids compared to STD-fed female rats at ZT3 (Table 3). Interestingly, sex differences were lower at ZT15, showing STD-fed male lower levels of leucine (Leu), Tryptophan (Trp) and Lysine (Lys) compared to STD-fed females. CAF feeding significantly decreased some amino acid levels, being a high number of amino acids affected in males. In this context, CAF-fed females showed higher levels of alanine (Ala) and lower levels of Trp and Gly at both ZT3 and ZT15 compared to STD-fed females at the same time of day. CAF decreased Leu levels only at ZT15 in comparison with STD-fed females and increased glutamate (Glu) at ZT3. In contrast, CAF feeding decreased Leu, Lys, Phe, isoleucine (Ile), Val and Trp at both ZT3 and ZT15 compared to STD-fed males, while Gly, histidine (His) and Gln only were decreased at ZT3. Interestingly, although CAF feeding decreased amino acids levels, it did not abolish the time-of-day effect shown in rats on the STD diet (Table 3).| Female | Male | P-Valuea | FDRb | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| STD | CAF | STD | CAF | |||||||||||||||
| VH-ZT0 | GSPE-ZT0 | VH-ZT12 | GSPE-ZT12 | VH-ZT0 | GSPE-ZT0 | VH-ZT12 | GSPE-ZT12 | VH-ZT0 | GSPE-ZT0 | VH-ZT12 | GSPE-ZT12 | VH-ZT0 | GSPE-ZT0 | VH-ZT12 | GSPE-ZT12 | |||
| Data are plotted as means ± SD (arbitrary units).a P-Value by one-way ANOVA. *, t, # and $ indicate significant GPSE, ZT, diet and sex effect respectively analyzed by one-way ANOVA followed by LSD post hot test (p < 0.05). The 4-way ANOVA and 2-way ANOVA significant effects are shown in Table S1.†b FDR: false discovery rate (p < 0.05). STD: standard diet; CAF: cafeteria diet; GSPE: grape seed proanthocyanidin extract; time of the day; ZT0: VH or GSPE time administration (when light was turned on); ZT12: VH or GSPE time administration (when light was turned off). | ||||||||||||||||||
| Amino acids metabolism | ||||||||||||||||||
| Leucine | 0.07 ± 0.007 | 0.07 ± 0.009 | 0.07 ± 0.007 | 0.06 ± 0.009t | 0.08 ± 0.01 | 0.07 ± 0.006* | 0.06 ± 0.01#t | 0.05 ± 0.008*t | 0.08 ± 0.007$ | 0.07 ± 0.008 | 0.06 ± 0.02$t | 0.05 ± 0.01t | 0.07 ± 0.007#$ | 0.07 ± 0.007 | 0.04 ± 0.007#$t | 0.05 ± 0.002t | 2.97 × 10−15 | 0.01 |
| Tyrosine | 0.03 ± 0.005 | 0.03 ± 0.007 | 0.02 ± 0.003t | 0.02 ± 0.008t | 0.03 ± 0.005 | 0.03 ± 0.006 | 0.02 ± 0.005t | 0.02 ± 0.002t | 0.04 ± 0.005$ | 0.04 ± 0.004$ | 0.03 ± 0.007t | 0.03 ± 0.004t | 0.04 ± 0.007$ | 0.03 ± 0.004$ | 0.03 ± 0.004$t | 0.03 ± 0.004$t | 7.15 × 10−15 | 0.01 |
| Lysine | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.07 ± 0.008*t | 0.09 ± 0.008 | 0.09 ± 0.007 | 0.09 ± 0.01 | 0.08 ± 0.01*t | 0.08 ± 0.008 | 0.08 ± 0.01$ | 0.07 ± 0.009$t | 0.06 ± 0.009$ | 0.07 ± 0.008#$ | 0.09 ± 0.007* | 0.05 ± 0.01#t | 0.06 ± 0.004$t | 5.99 × 10−14 | 0.01 |
| Phenylalanine | 0.07 ± 0.005 | 0.07 ± 0.004 | 0.08 ± 0.01 | 0.07 ± 0.008 | 0.07 ± 0.007 | 0.07 ± 0.005 | 0.07 ± 0.005 | 0.07 ± 0.006 | 0.09 ± 0.01$ | 0.1 ± 0.02$ | 0.08 ± 0.007t | 0.09 ± 0.01*t | 0.07 ± 0.006# | 0.07 ± 0.006# | 0.07 ± 0.007# | 0.07 ± 0.003# | 2.34 × 10−12 | 0.01 |
| Isoleucine | 0.05 ± 0.004 | 0.05 ± 0.009 | 0.05 ± 0.005 | 0.04 ± 0.008*t | 0.06 ± 0.009 | 0.05 ± 0.005* | 0.04 ± 0.008t | 0.03 ± 0.006*t | 0.06 ± 0.006 | 0.05 ± 0.008 | 0.05 ± 0.009t | 0.04 ± 0.006t | 0.05 ± 0.006#$ | 0.05 ± 0.006 | 0.03 ± 0.005#$t | 0.04 ± 0.002t | 2.52 × 10−11 | 0.01 |
| Alanine | 0.21 ± 0.04 | 0.22 ± 0.03 | 0.16 ± 0.03t | 0.15 ± 0.03t | 0.25 ± 0.04# | 0.27 ± 0.04# | 0.2 ± 0.04#t | 0.18 ± 0.04t | 0.25 ± 0.02$ | 0.23 ± 0.02 | 0.15 ± 0.02t | 0.18 ± 0.03t | 0.22 ± 0.07 | 0.23 ± 0.03$ | 0.18 ± 0.06t | 0.17 ± 0.008$t | 3.54 × 10−10 | 0.02 |
| Valine | 0.09 ± 0.009 | 0.09 ± 0.02 | 0.09 ± 0.007 | 0.08 ± 0.01*t | 0.1 ± 0.02 | 0.08 ± 0.009* | 0.08 ± 0.01t | 0.07 ± 0.009*t | 0.1 ± 0.01$ | 0.09 ± 0.01 | 0.08 ± 0.02t | 0.07 ± 0.01t | 0.08 ± 0.01#$ | 0.09 ± 0.01 | 0.06 ± 0.008#$t | 0.06 ± 0.005t | 4.08 × 10−10 | 0.02 |
| Tryptophan | 0.06 ± 0.005 | 0.06 ± 0.009 | 0.06 ± 0.005 | 0.06 ± 0.008 | 0.05 ± 0.008# | 0.05 ± 0.007# | 0.05 ± 0.003# | 0.05 ± 0.007# | 0.06 ± 0.004 | 0.05 ± 0.003*$ | 0.05 ± 0.01$ | 0.04 ± 0.004*$ | 0.05 ± 0.004# | 0.05 ± 0.007 | 0.04 ± 0.006#$ | 0.04 ± 0.005 | 2.76 × 10−7 | 0.02 |
| Glycine | 0.09 ± 0.02 | 0.09 ± 0.02 | 0.1 ± 0.007 | 0.09 ± 0.02 | 0.08 ± 0.008# | 0.07 ± 0.01# | 0.08 ± 0.009# | 0.08 ± 0.007# | 0.11 ± 0.009$ | 0.11 ± 0.01$ | 0.1 ± 0.01 | 0.1 ± 0.009 | 0.08 ± 0.009# | 0.09 ± 0.01*# | 0.09 ± 0.01 | 0.1 ± 0.01$ | 1.35 × 10−6 | 0.03 |
| Histidine | 0.03 ± 0.005 | 0.03 ± 0.005 | 0.03 ± 0.002 | 0.03 ± 0.005 | 0.03 ± 0.005 | 0.03 ± 0.006 | 0.03 ± 0.004 | 0.03 ± 0.005 | 0.04 ± 0.003$ | 0.03 ± 0.004 | 0.03 ± 0.003t | 0.03 ± 0.002 | 0.03 ± 0.004# | 0.03 ± 0.002 | 0.03 ± 0.003 | 0.03 ± 0.004 | 1.00 × 10−4 | 0.03 |
| Glutamine | 0.25 ± 0.04 | 0.25 ± 0.02 | 0.3 ± 0.04t | 0.27 ± 0.02 | 0.27 ± 0.02 | 0.26 ± 0.01 | 0.29 ± 0.02 | 0.27 ± 0.02 | 0.29 ± 0.04$ | 0.28 ± 0.04$ | 0.29 ± 0.03 | 0.3 ± 0.04$ | 0.27 ± 0.03 | 0.3 ± 0.01$* | 0.29 ± 0.03 | 0.31 ± 0.02$ | 1.17 × 10−4 | 0.03 |
| Glutamate | 0.05 ± 0.01 | 0.06 ± 0.009 | 0.05 ± 0.006 | 0.05 ± 0.01 | 0.07 ± 0.008# | 0.07 ± 0.008 | 0.06 ± 0.01t | 0.05 ± 0.007t | 0.07 ± 0.006$ | 0.06 ± 0.005 | 0.05 ± 0.009t | 0.06 ± 0.01 | 0.06 ± 0.01#$ | 0.06 ± 0.006 | 0.06 ± 0.006$ | 0.06 ± 0.01 | 1.70 × 10−3 | 0.04 |
| Glycolysis | ||||||||||||||||||
| Glucose | 1.68 ± 0.26 | 1.66 ± 0.1 | 1.79 ± 0.16 | 1.68 ± 0.25 | 1.67 ± 0.2 | 1.78 ± 0.18 | 1.84 ± 0.13t | 1.81 ± 0.13 | 1.94 ± 0.12$ | 1.82 ± 0.12 | 1.88 ± 0.21 | 1.85 ± 0.12 | 1.66 ± 0.11# | 1.84 ± 0.11* | 1.89 ± 0.19t$ | 1.82 ± 0.06 | 9.05 × 10−3 | 0.04 |
| Pyruvate | 0.02 ± 0.004 | 0.03 ± 0.007 | 0.02 ± 0.007 | 0.02 ± 0.007 | 0.03 ± 0.002 | 0.03 ± 0.009 | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.003 | 0.02 ± 0.004 | 0.02 ± 0.006 | 0.02 ± 0.003 | 0.02 ± 0.007$ | 0.02 ± 0.003$ | 0.02 ± 0.007 | 0.02 ± 0.003 | 5.88 × 10−4 | 0.04 |
| Lactate | 0.84 ± 0.17 | 1.02 ± 0.42 | 0.94 ± 0.27 | 1.11 ± 0.26 | 0.87 ± 0.14 | 1.06 ± 0.37 | 1.04 ± 0.35 | 0.99 ± 0.28 | 0.93 ± 0.14 | 0.83 ± 0.19 | 1 ± 0.23 | 0.97 ± 0.23 | 0.88 ± 0.25 | 0.9 ± 0.23 | 0.92 ± 0.48 | 0.93 ± 0.37 | 9.15 × 10−1 | 0.05 |
| Lipid metabolism | ||||||||||||||||||
| ARA + EPA | 10.08 ± 0.56 | 10.77 ± 1.03 | 10.91 ± 1.37 | 10.98 ± 1.46 | 7.1 ± 0.86# | 8.29 ± 0.62*# | 8.98 ± 0.53#t | 8.13 ± 2.11# | 6.99 ± 0.53$ | 7.23 ± 0.67$ | 7.6 ± 0.43$ | 7.36 ± 0.96$ | 8.34 ± 1.18$# | 7.87 ± 1.06 | 7.75 ± 0.88 | 8.48 ± 0.61# | 2.35 × 10−17 | 0.003 |
| Triglycerides | 6.36 ± 1.16 | 7.34 ± 2.85 | 3.78 ± 0.72 | 3.65 ± 0.85 | 16.33 ± 10.38# | 15.13 ± 4.68 | 10.69 ± 5.07 | 11.66 ± 6.05 | 10.33 ± 3.58 | 10.91 ± 2.85 | 5.12 ± 2.34 | 7.86 ± 2.34 | 42.8 ± 20.92$# | 39.57 ± 8.26#$ | 27.33 ± 17.92#$t | 25.36 ± 6.46#$t | 5.65 × 10−17 | 0.004 |
| Free cholesterol | 1.5 ± 0.16 | 1.7 ± 0.1* | 1.65 ± 0.18 | 1.74 ± 0.16 | 1.13 ± 0.08# | 1.3 ± 0.1*# | 1.31 ± 0.1#t | 1.29 ± 0.25# | 1.41 ± 0.11 | 1.26 ± 0.13*$ | 1.61 ± 0.14t | 1.22 ± 0.15*$ | 1.63 ± 0.15#$ | 1.5 ± 0.17#$ | 1.47 ± 0.18#$t | 1.48 ± 0.07#$ | 3.48 × 10−16 | 0.005 |
| Oleic acid | 14.56 ± 2.67 | 25.25 ± 27.06 | 10.37 ± 1.41 | 10.2 ± 1.65 | 35.17 ± 21.03# | 33.27 ± 10.54 | 23.17 ± 9.96 | 26.55 ± 13.5 | 20.04 ± 3.99 | 19.38 ± 5.15 | 11.83 ± 2.5 | 14.27 ± 3.37 | 82.06 ± 38.98#$ | 77.02 ± 21.36#$ | 53.49 ± 36.6#$t | 48.29 ± 12.08#$t | 3.83 × 10−16 | 0.007 |
| MUFAs | 27.43 ± 1.68 | 40.45 ± 37.47 | 23.13 ± 3.55 | 21.95 ± 2.6 | 47.05 ± 26.29 | 43.8 ± 11.49 | 31.38 ± 10.68 | 31.89 ± 14.3 | 31.6 ± 6.82 | 29.17 ± 6.3 | 22.84 ± 3.78 | 23.45 ± 3.35 | 117.79 ± 54.14#$ | 111.54 ± 26.44#$ | 74.05 ± 56.17#$t | 65.47 ± 15.26#$t | 7.47 × 10−16 | 0.008 |
| Linoleic acid | 9.11 ± 0.73 | 11.54 ± 8.66 | 7.25 ± 1.25 | 6.72 ± 0.76 | 10.81 ± 5.09 | 11.15 ± 2.16 | 7.76 ± 2.8 | 6.87 ± 2.41 | 10.76 ± 3.56 | 11.39 ± 2.5 | 7.87 ± 2.08 | 10.01 ± 1.4 | 29.24 ± 13.19#$ | 28.5 ± 3.94#$ | 19.58 ± 14.11#$t | 17.19 ± 3.15$#t | 1.26 × 10−15 | 0.009 |
| Total fatty acids | 94.21 ± 6.51 | 125.82 ± 43.9* | 97.22 ± 6.26 | 104.15 ± 8.02 | 103.87 ± 30.07 | 107.7 ± 13.49 | 93.85 ± 10.92 | 104.81 ± 23.49 | 106.26 ± 5.62 | 96.08 ± 7.94 | 109.86 ± 5.54 | 87.16 ± 6.74 | 204.61 ± 65.74#$ | 178.93 ± 34.39#$ | 149.77 ± 63.15#$t | 135.87 ± 15.84#$t | 1.62 × 10−13 | 0.01 |
| Diglycerides | 0.45 ± 0.06 | 0.53 ± 0.25* | 0.57 ± 0.14 | 0.39 ± 0.15 | 0.37 ± 0.14 | 0.42 ± 0.11 | 0.37 ± 0.06# | 0.49 ± 0.26 | 0.35 ± 0.05 | 0.37 ± 0.11 | 0.33 ± 0.07$ | 0.32 ± 0.05 | 0.89 ± 0.33$# | 0.82 ± 0.17$# | 0.73 ± 0.3$# | 0.67 ± 0.08$# | 8.88 × 10−12 | 0.02 |
| DHA | 1.41 ± 0.11 | 1.42 ± 0.34 | 1.35 ± 0.21 | 1.2 ± 0.16 | 0.98 ± 0.28# | 1.12 ± 0.16# | 1.11 ± 0.26 | 0.97 ± 0.32 | 1.13 ± 0.36$ | 1.35 ± 0.27 | 0.9 ± 0.21$t | 1.09 ± 0.11 | 1.81 ± 0.48#$ | 1.5 ± 0.16*$ | 1.45 ± 0.32#$t | 1.46 ± 0.28#$ | 8.50 × 10−9 | 0.02 |
| PUFAs | 34.31 ± 1.71 | 35.21 ± 5.78 | 35.77 ± 5.74 | 34.01 ± 4.62 | 29.05 ± 4.39 | 34.27 ± 3.27 | 36.11 ± 4.99t | 32.4 ± 10.57 | 27.71 ± 4.37$ | 25.74 ± 3.86$ | 27.75 ± 2.39$ | 26.47 ± 3.49$ | 41.95 ± 7.75#$ | 37.78 ± 6.34# | 38.98 ± 6.67# | 40.46 ± 5.12#$ | 1.31 × 10−8 | 0.02 |
| Glycerol | 0.07 ± 0.02 | 0.07 ± 0.02 | 0.07 ± 0.02 | 0.08 ± 0.02 | 0.11 ± 0.03# | 0.12 ± 0.03# | 0.09 ± 0.02# | 0.09 ± 0.02t | 0.06 ± 0.01 | 0.07 ± 0.02 | 0.06 ± 0.01 | 0.07 ± 0.02 | 0.07 ± 0.02$ | 0.08 ± 0.02$ | 0.07 ± 0.02 | 0.09 ± 0.02# | 2.94 × 10−7 | 0.03 |
| Total phospholipids | 5.38 ± 0.74 | 5.81 ± 1.08 | 5.86 ± 0.97 | 5.75 ± 0.6 | 5.06 ± 0.8 | 5.73 ± 0.74 | 5.74 ± 0.82 | 5.34 ± 1.66 | 4.74 ± 0.46 | 4.07 ± 0.64$ | 4.78 ± 0.6$ | 4.64 ± 0.63$ | 7.4 ± 1.53$# | 6.62 ± 1.01# | 5.89 ± 1.46#t | 5.9 ± 0.79# | 4.82 × 10−7 | 0.03 |
| Esterified cholesterol | 9.31 ± 1.04 | 8.46 ± 0.67 | 9.47 ± 1.92 | 8.75 ± 1.09 | 6.66 ± 1.11# | 7.83 ± 0.84 | 8.28 ± 0.73#t | 6.56 ± 2.03*#t | 7.46 ± 1$ | 6.21 ± 1.12$ | 7.72 ± 1.45$ | 7.22 ± 1.62$ | 7.13 ± 1.8 | 6.52 ± 1.8$ | 6.01 ± 1.09$# | 6.49 ± 1.01 | 1.31 × 10−6 | 0.03 |
| Cholesterol | 17.21 ± 2 | 16.28 ± 1.14 | 17.97 ± 3.46 | 16.81 ± 1.9 | 12.41 ± 1.6# | 14.76 ± 1.43* | 14.81 ± 1.18#t | 12.51 ± 3.92*#t | 13.95 ± 1.43$ | 11.63 ± 1.96*$ | 14.46 ± 2.45$ | 12.77 ± 2.67*$ | 15.88 ± 2.76$ | 14.21 ± 2.79# | 12.85 ± 2.71t | 13.56 ± 1.52 | 1.46 × 10−6 | 0.03 |
| Acetate | 0.08 ± 0.01 | 0.09 ± 0.02 | 0.08 ± 0.004 | 0.08 ± 0.01 | 0.08 ± 0.009 | 0.08 ± 0.01# | 0.08 ± 0.009 | 0.08 ± 0.009 | 0.07 ± 0.008 | 0.1 ± 0.01* | 0.09 ± 0.01t | 0.07 ± 0.007*$t | 0.08 ± 0.01 | 0.07 ± 0.01# | 0.07 ± 0.01#$t | 0.07 ± 0.01$ | 3.48 × 10−5 | 0.04 |
| Choline metabolism | ||||||||||||||||||
| Choline | 0.005 ± 0.001 | 0.006 ± 0.001 | 0.007 ± 0.002t | 0.006 ± 0.001 | 0.006 ± 0.001 | 0.005 ± 0.001 | 0.006 ± 0.001 | 0.006 ± 0.001t | 0.007 ± 0.001$ | 0.006 ± 0.001 | 0.008 ± 0.001 | 0.007 ± 0.001 | 0.005 ± 0.001# | 0.005 ± 0.001 | 0.006 ± 0.001# | 0.007 ± 0.002 | 3.02 × 10−5 | 0.03 |
| Betaine | 0.25 ± 0.05 | 0.26 ± 0.04 | 0.26 ± 0.04 | 0.22 ± 0.05 | 0.2 ± 0.03# | 0.18 ± 0.03# | 0.22 ± 0.04# | 0.21 ± 0.04 | 0.28 ± 0.03 | 0.27 ± 0.04 | 0.25 ± 0.02 | 0.29 ± 0.05 | 0.21 ± 0.05# | 0.2 ± 0.03# | 0.18 ± 0.03# | 0.2 ± 0.03# | 3.02 × 10−8 | 0.03 |
| N,N-Dimethylglycine | 0.006 ± 0.001 | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.006 ± 0.001 | 0.006 ± 0.001 | 0.007 ± 0.001 | 0.006 ± 0.001 | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.006 ± 0 | 0.006 ± 0.001 | 0.006 ± 0.001 | 0.006 ± 0.0005 | 0.006 ± 0.001 | 1.27 × 10−1 | 0.05 |
| Nucleotide biosynthesis | ||||||||||||||||||
| Formate | 0.3 ± 0.03 | 0.3 ± 0.03 | 0.31 ± 0.02 | 0.28 ± 0.04 | 0.29 ± 0.03 | 0.24 ± 0.06*# | 0.3 ± 0.04 | 0.32 ± 0.03# | 0.29 ± 0.02 | 0.21 ± 0.02*$ | 0.24 ± 0.01$t | 0.13 ± 0.01*$ | 0.28 ± 0.02 | 0.29 ± 0.01#$ | 0.26 ± 0.03$ | 0.28 ± 0.03#$ | 1.18 × 10−21 | 0.001 |
| Bile salt | ||||||||||||||||||
| Cholate | 0.01 ± 0.0003 | 0.01 ± 0.001 | 0.01 ± 0.001 | 0.01 ± 0.001 | 0.01 ± 0.0003 | 0.01 ± 0.001 | 0.01 ± 0.001 | 0.01 ± 0.001 | 0.008 ± 0.001$ | 0.009 ± 0.001* | 0.008 ± 0.001$ | 0.007 ± 0.001*t$ | 0.01 ± 0.001# | 0.01 ± 0.001# | 0.01 ± 0.001# | 0.01 ± 0.001# | 7.16 × 10−4 | 0.04 |
In STD-fed females, GSPE administered at ZT0 did not exert any effect on amino acids levels. However, when GSPE was administered at ZT12, Lys, Ile and Val levels were significantly decreased compared to its respective control. GSPE in CAF-fed females significant decreased Leu, Ile and Val levels compared to CAF females received with VH at both ZT0 and ZT12. Lys levels were decreased only by GSPE at ZT12, showing a time-of-day effect not observed in control rats. On the other hand, in STD-fed males, the amino acids Leu, Tyr, Lys, Phe, Ile, Ala, Val, His and Glu were decreased at night. GSPE treatment led to a decrease of Trp at both ZT0 and ZT12, and an increase of Phe only when it was administered at ZT12. When male rats were fed a CAF diet, a similar time-of-day effect pattern was observed in amino acid levels in VH groups, significantly decreasing at night (Leu, Tyr, Lys, Ile, Ala and Val). However, GSPE effect only was found when it was administered at ZT0, increasing Lys, Gly and Gln levels (Table 3).
GSPE increases glucose levels only at ZT0 in CAF-fed males
Glucose levels did not show differences between groups in STD-fed animals. However, glucose levels increased at ZT12 in both CAF-fed females and males receiving VH compared to their respective controls at ZT0. Moreover, a GSPE effect was demonstrated at ZT0 in CAF-fed males, increasing glucose levels compared to control rats at the same time point. Regarding metabolites involved in glycolysis, no significant differences were found in pyruvate and lactate (Table 3).GSPE modulation of lipid serum profiles depend on the administration time, sex, and type of diet
An overall sex and diet effect was observed in lipid serum metabolites (Table S1†). ARA + EPA (arachidonic acid + eicosapentaenoic acid), triglycerides, free cholesterol, oleic acid, MUFAs, linoleic acid, total fatty acids, diglycerides, DHA (docosahexaenoic acid), glycerol, cholesterol and acetate levels were influenced by the sex of the rats. These lipid metabolites together with PUFAs and total phospholipids were also affected by CAF feeding, resulting in a significant time-of-day effect in CAF-fed females and increasing ARA + EPA, free cholesterol, PUFAs, esterified cholesterol and cholesterol at ZT15. However, STD-fed females did not show any differences between ZT3 and ZT15. Otherwise, STD-fed males showed a time-of-day effect increasing free cholesterol and acetate levels and decreasing DHA at ZT15. CAF-fed males also showed a high number of changes in lipid serum metabolites depending on the time of day, with lower levels of triglycerides, free cholesterol, oleic acid, MUFAs, linoleic acid, total fatty acids, DHA, total phospholipids, cholesterol, and acetate at ZT15 (Table 3).GSPE effect was different according to the diet, sex and time of administration (Table 3). When GSPE was administered at ZT0 in STD-fed females, higher levels of free cholesterol, total fatty acids and diglycerides were observed compared to its respective control rats. In CAF-female rats, GSPE administered at ZT0 increased ARA + EPA, free cholesterol and cholesterol levels. However, GSPE decreased levels of esterified cholesterol and cholesterol when it was administered at ZT12 in female CAF rats compared to its respective control. In STD-fed males GSPE significantly decreased free and total cholesterol at ZT0 and ZT12, while increased acetate at ZT0. In males CAF rats, GSPE decreased DHA only at ZT0, showing lower effect than in female CAF rats.
GSPE do not influence choline metabolism
Choline levels were higher in STD-fed females at ZT15 compared to ZT3, and no GSPE effect was observed. However, CAF-induced obese females lost the time-of-day effect observed in STD-fed females, whereas GSPE administration at ZT12 restored it, with higher levels of choline at night. In contrast, STD-fed males had no effect of either time-of-day or GSPE effect. CAF-fed males displayed lower levels of choline compared to STD-fed males. On the other hand, betaine metabolite only showed a diet effect, being its levels lower in CAF-fed animals (females and males) (Table 3).GSPE influences differently formate levels in female and male rats depending on the diet
Although no effect was observed in STD-fed females in formate levels, CAF-fed females showed a GSPE dependent effect, decreasing formate levels when GSPE was administered at ZT0. In contrast, GSPE administration in STD-fed males decreased formate levels at both ZT0 and ZT12, but no time-of-day effect was found. GSPE did not show any effect in CAF-fed males (Table 3).GSPE affects differently cholate levels depending on the time of administration in male rats on a standard diet
No effect was observed in STD- or CAF-fed rats in cholate levels, although STD-males showed lower levels of cholate than STD-fed females, showing a diet effect influenced by sex, since it was not observed in females (S × D, p < 0.05). However, in STD-fed males, GSPE increased cholate levels when administered at ZT0, whereas it decreased them when administered at ZT12. CAF-fed males showed higher levels of cholate than STD-fed males (Table 3; Table S1†).Clock genes correlate with serum melatonin and amino acids levels
To evaluate the influence of clock genes on melatonin and serum metabolite levels, correlations between these parameters were carried out. Only a significant positive correlation was observed between Per2 and melatonin (Fig. 5a; Table S2†). However, several significant correlations were found between clock genes and serum metabolite levels (Table S3†). The strongest significant correlations were observed between amino acids levels and clock genes (R = ±0.4) (Fig. 5b–e). Per2 negatively correlated with Tyr, Ala, and Leu (Fig. 5b–d) and Per1 negatively correlated with Ala (Fig. 5e). Other significant correlations (R = ±0.3) were found between clock genes and amino acids levels (Table S3†): Per2 negatively correlated with Iso, Val, Trp and His; Bmal1 positively correlated with Glut and Tyr; and Per1 negatively correlated with Lys and Tyr. Moreover, significant correlations were found between Cry and Betaine (R = 0.329) and between RORα and Glycerol (R = −0.305). Additionally, the following weaker significant correlations (R = 0.2) were observed: Cry positively correlated with choline, Gly, glucose and Phe, and negatively correlated with glycerol; Per2 positively correlated with choline, and negatively with linoleic acid. RORα positively correlated with Gly, Phe, free cholesterol and Leu. Finally, negative significant correlations were found between Bmal1 and free cholesterol, diglycerides and ARA + EPA. | ||
| Fig. 5 Correlations between hypothalamus clock genes and serum melatonin or serum metabolites levels. (a) Significant correlations between hypothalamus clock genes and serum melatonin levels, (b–e) strongest significant correlations between hypothalamus clock genes and serum metabolite levels. The analyses were performed by Pearson's rank correlation coefficient (R). | ||
Discussion
GSPE has a wide range of beneficial effects on health, one of them being the prevention of obesity and metabolic-related disorders.51,52 GSPE has also been found to be a key factor influencing clock genes.40,42 This fact is relevant since circadian rhythms are essential to maintain host homeostasis, and their alteration by misalignment cues such as obesogenic diets is associated with the development of metabolic disorders, including obesity.53 Remarkably, it has recently been reported that GSPE effects on liver and adipose tissue depended on the time of administration,43,44 and that the plasma bioavailability of GSPE also was impacted by the administration time of this proanthocyanidin-rich extract.45 Therefore, it is likely that there is a bidirectional relationship between GSPE and circadian rhythms. Hence, the current study evaluated if GSPE effects on the metabolome are conditioned by the time-of-day in both healthy and obesogenic context, investigating also the influence of the sex, which is an important factor to take into account as circadian rhythms influence females and males differently.15,16 To achieve this goal, GSPE was orally administered at 25 mg kg−1 day−1. This dose is within the estimated range of polyphenol intake in humans54–56 and is equivalent to the daily intake of 367 mg for a 70 kg person, calculated using the body surface area normalization method.57The circadian clock system can be disturbed by obesogenic diets, affecting in turn to hormone and serum metabolites rhythmicity12,58 and contributing to the development of metabolic disorders.59 Additionally, obesogenic diets do not act in the same manner in both sexes.60,61 For instance, plasma parameters associated with metabolic syndrome, such as leptin and insulin, are modulated in a sex-specific manner by obesogenic diets.62 This fact could be explained because circadian rhythms also exhibit differences between females and males, showing differences in the clock gene expressions.15 In this regard, this study showed that the clock gene expressions, as well as metabolite levels showed a time-of-day effect, differently influenced by the diet in a sex-dependent manner. These changes observed in the clock genes expressions were significantly correlated with melatonin and different serum metabolites. Melatonin levels, positively correlated with the relative expression of Per2, being in concordance with other studies in which the increase of melatonin raise the expression of Per2 in both SCN and pituitary gland.63 Furthermore, melatonin levels and Per2 expression were higher at night independently of health condition or sex, since the secretion of melatonin by the pineal gland occurs at night under the control of the light–dark cycles.64 Thus, pineal gland and central clock tissue interact with each other;63 SCN relays photoperiodic information through the retina to the pineal gland via sympathetic nervous system, and based on the information provided by the SCN, the pineal gland either up- or down-regulates the production of melatonin.65 In turn, melatonin signal impacts the function of the SCN, regulating the central clock genes and synchronize physiological activities.66 Interestingly, STD-fed males showed higher melatonin levels, and in consequence higher expression of Per2, compared to STD-fed females. These results could be associated with the fact that in women melatonin cycle has a higher amplitude and the secretion peak appears earlier than in males.67 Therefore, at ZT15 (at night) the highest peak of melatonin secretion may have already occurred in females, whereas in males this process is occurring in that point of time. Moreover, hypersecretion during metabolic syndrome, due to an increase of the sympathetic activity in the SCN,68 could explain the higher levels of this hormone showed by CAF-fed females compared to STD-fed females.
Amino acids, lipid and choline metabolism was also affected depending on the time-of-day, diet and sex. These results were expected since metabolomic profiles vary due to health status, sex and diurnal variations,69 as it is discussed above. Regarding to the sex differences, the molecular mechanisms which explain them remain largely unidentified, although it seems that sex hormones play an important role. Hence, it has been described that estrogen in women and androgen in men impact on the fatty acid, triglyceride, and cholesterol metabolism.70 Furthermore, these differences by sex could be explained by the different expressions of the enzymes for fatty acids synthesis and oxidation, which have been reported to be more active in females and in males, respectively.71 The sexual dimorphism in the amino acid metabolism has not been elucidated yet, but in the same manner, sex hormones appear to be crucial, promoting differences in musculature and in turn, in the protein metabolism.72,73 Serum metabolites were also correlated with changes in clock gene expressions. Amino acids and other metabolites such as betain, choline, DAG, glycerol and free cholesterol were significantly correlated with the central clock genes, indicating that circadian clocks can control the metabolism in a time-of-day-dependent manner.21,26 Among amino acids, it is worth highlighting the changes observed in the levels of Trp, and its negative correlation with Per2. Trp is an essential amino acid supplied by diet. It can be metabolized into different metabolites in both the gut microbiota and tissue cells, being a precursor of a large number of biologically active metabolites.74 Trp is the sole precursor of the serotonin synthesis in the brain, and subsequently, of the melatonin hormone in the pineal gland.75 However, over 90% of serotonin is located in the gastrointestinal tract, and it is produced from enterochromaffin cells. Moreover, more than 90% of total Trp is catabolized, via the kynurenine pathway, into kynurenine in the liver, which can be further metabolized into kynurenic acid and quinolinic acid (QUIN) called kynurenine (inflammatory mediators capable of reach the brain).76 Finally, QUIN can be converted into nicotinamide adenine dinucleotide (NAD) that play a key role in the energy homeostasis,76 following 24 h rhythmicity.77 Also, a small amount of Trp is converted by gut microbiota into indole and its derivatives, which have exhibited protective effects.78 In addition, Trp metabolism has been shown to follow circadian/diurnal oscillations, and Trp and their derived are believed to modulate circadian rhythms via multiple molecular mechanism.79 Along this line, our results show lower levels of Trp in healthy females and males rats at night, which could be associated with the increased levels of melatonin at that point. Furthermore, healthy males showed higher levels of Trp compared to females, showing also higher levels of melatonin as it was commented above. However, in females and obese rats, Trp levels decrease at both in the morning and at night, losing the rhythmicity. This fact could be explained by an overactivation of the kynurenine pathway since diseases characterized by a low grade-inflammation and a dysbiosis of the gut microbiota, such as obesity show higher levels of kynurenine.74 Nevertheless, the decrease of Trp in the morning was not linked to higher melatonin level in the morning. In this regard, clock genes were studied only in two time points. Therefore, a further study to evaluate rhythmicity may be necessary to obtain more differences or significant correlations in comparison with the current results. Moreover, further studies would be needed to evaluate the Trp metabolism and stablish how Trp and its derivate could modulate circadian system, promoting a good health state, evaluating also the gut microbiota composition and their produced metabolites, which is essential in the homeostasis maintenance.80,81
GSPE has been shown to modulate clock genes in a time-of-day-dependent manner in liver43 and in the adipose tissue44 in male rats to prevent the obesity development. GSPE could therefore modulate also central clock genes and its effect on the circadian clocks may influence several metabolic pathways to ameliorate obesity disease, and this could be reflected in the serum metabolic profiles. The current study shows that GSPE impacts on the central clocks and metabolome and, for the first time to our knowledge, reports that GSPE effects not only are influence by the time of day but also are strongly influenced by the health condition and sex. In this regard, these results showed that GSPE influence the expression pattern of clock genes in the hypothalamus differently in healthy and obesogenic conditions in a sex-specific manner. Particularly, GSPE influenced clock, Per1 and Per2 at night in STD females, whereas in CAF females, GSPE affected the expression of Bmal1 and Cry in the morning. Furthermore, GSPE effects on clock genes in STD males were found at both morning and night, whereas in CAF males no GSPE effects on clock genes were found. Thus, these results indicate that ability of PAs to modulate the hypothalamic clock genes depends largely on the time of administration and importantly on the diet and sex. Interestingly, in this case, GSPE did not affect melatonin hormone levels, in contrast to another acute study in which GSPE modulated plasma melatonin levels when the extract was administered in the morning.41 However, the different strain of rats used, the lower dose of GSPE administered in this study (versus 250 mg kg−1) and the different time points in which melatonin was measured, may explain this discrepancy. Furthermore, Trp levels were also not altered significantly by GSPE, which could also explain why no differences in melatonin levels are observed by GSPE. To date, a relationship between melatonin and bioactive compounds such as PAs has not yet been established, so it would be interesting to continue investigating in this field to elucidate the role of clock genes and melatonin as well as the derived-melatonin metabolites in the metabolome and whether they can be modulated by proanthocyanidin-rich extracts. Moreover, as GSPE modulates gut microbiota composition,82 it would be interesting to advance in the study of its composition and the metabolites derives from it, and how it may regulate the host homeostasis impacting on the circadian rhythms.
It has been recently reported that GSPE administration at night restore the rhythmic metabolism of Ala, aspartate (Asp) and Glu in male CAF-fed rats.83 The changes observed in amino acids levels by GSPE according to the time-of-day could be explained due to the different clock gene expressions in the hypothalamus. Amino acids and skeletal muscle clock genes are intimately linked,84 and observational studies showed a link between diurnal variations in amino acid plasma concentrations and insulin resistance and diabetes.85,86 Hence, the beneficial effect of GSPE on insulin resistance and obesity-related disorders may depends on the time of administration, sex and diet. In the same way, lipid metabolism was influenced by the time of GSPE administration according to the diet and sex. In this case, GSPE administered at night to CAF-fed females had a beneficial effect, decreasing cholesterol, total fatty acids and diglycerid levels. Interestingly, when GSPE was administered in the morning to both STD- and CAF-fed female rats, GSPE effects were detrimental, increasing the levels of these lipid metabolites. In males, the effect of GSPE administration was less remarkable. CAF-fed male rats only showing lower DHA levels in the morning, whereas GSPE decreased free and total cholesterol levels in STD-fed males when it was administered both in the morning and at night. These findings indicate that beneficial GSPE effects are strongly influenced by the time of the day in which it is administered, being sex and health status two key factors in its functionality. It is described that cholesterol is synthetized following a circadian rhythmicity in the liver thought the circadian expression of HMG-CoA.87 Moreover, enzymes that regulate fatty acids synthesis also present rhythmic expression patterns.88 Therefore, to get beneficial effects, GSPE may act on these pathways, being important the time of administration according to the circadian expression patterns of the enzymes involved. Furthermore, it has been reported that GSPE repress the expression of the clock gene Cry in the inguinal WAT in males rats only at night, which was associated with a lower adiposity in this tissue,44 suggesting that GSPE is influenced by the circadian rhythms. Cholate levels were also affected by GSPE administration time in a diet and sex-dependent manner. Cholate as a primary bile acids, is synthetized from cholesterol via two major pathways in hepatocytes.89 The key enzyme, cholesterol 7α-hydroxylase (CYP7A1), and the serum levels of total bile acids in mice have been shown to follow diurnal rhythmicity, playing the circadian rhythms a crucial role in the maintenance of bile acid homeostasis.24,90 In fact, it has been reported that the alteration of hepatic clock gene expressions by a HFD or restringing feeding, disrupts bile acid metabolism and trigger metabolic diseases.91 Moreover, CYP7A1 and transporters involved in bile acid homeostasis display significant sex differences.92 Additionally, the bile acid metabolism is closely linked with the gut microbiota composition,93 which is also influenced by many factors including biological rhythms,94,95 diet,96 or sex.97 A recent study has shown that an apple polyphenol extract may modulate bile acid metabolism and gut microbiota via clock genes regulation.98 Accordingly, it has been recently described by our group, using the same experimental design, that the bioavailability of GSPE, especially the microbiota-derived metabolites, depends on the time of administration and the sex of the animals.45 In this context, significant higher overall levels of microbial colonic metabolites were found when GSPE was administered in the morning in CAF-fed rats, increasing these colonic metabolites in females rats fed with CAF compared to male rats, and being this effect no observed in STD-fed rats.45 Hence, gut microbiota play an important role in the metabolism of these PAs, exerting their derived metabolites several functions.99–101 Furthermore, gut microbiota has been also demonstrate to follow a diurnal rhythmicity102 and to be intrinsically influence by sex,97,103 so it may be a key factor in the mechanism of GSPE. Therefore, futures studies about the gut microbiota composition and their derived colonic metabolites bioactivities would be necessary to further evaluate the effects of GSPE on the metabolome and its relationship with circadian clocks. Therefore, although further studies are needed, these findings contribute to a better understanding the mechanism by which PAs may prevent and ameliorate metabolic-related disorders, adding knowledge in the chrononutrition field, which are currently continuously growing.
Conclusions
The findings of the current study advance on research focused on targeted nutritional recommendations and chrononutrition, showing that PAs can impact on circadian rhythms system, modulating the hypothalamic clock gene expressions and the amino acid and lipid metabolism in a time-of-day-, diet- and sex-dependent manner. Although further investigation is needed to elucidate the molecular mechanisms, our work highlights the need to take into account the time of day in which bioactive compounds or functional foods are administered, as well as the health of state and the sex in the design of new experiments and in the interpretation of the results, in order to make further progress towards personalized nutrition to improve metabolic diseases.Author contributions
Verónica Arreaza-Gil: investigation, formal analysis, visualization, writing-original draft, review & editing; Héctor Palacios-Jordan: investigation, formal analysis, validation; María del Mar Romero: investigation, formal analysis, validation, writing – review & editing; Cristina Torres-Fuentes: writing – review & editing; Miguel A Rodríguez: supervision; Xavier Remesar: conceptualization, supervision, writing – review & editing; José-Antonio Fernández-López: conceptualization, supervision, writing – review & editing; Anna Arola-Arnal: conceptualization, methodology, project administration, funding acquisition, supervision, writing – review & editing.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors thank Niurka Dariela Llópiz and Rosa Pastor for their assistance and technical support. This work was supported by MCIN/AEI/10.13039/501100011033/FEDER “Una manera de hacer Europa” (AGL2016-77105-R) and grant PID2021-128813OB-I00 funded by MCIN/AEI/10.13039/501100011033; V. A.-G. was supported by the Martí i Franquès Doctoral Fellowships Programme, Universitat Rovira i Virgili (PMF-PIPF-35).References
- T. Roenneberg and M. Merrow, The Circadian Clock and Human Health, Curr. Biol., 2016, 26, R432–R443 CrossRef CAS PubMed.
- F. Levi and U. Schibler, Circadian Rhythms: Mechanisms and Therapeutic Implications, Annu. Rev. Pharmacol. Toxicol., 2007, 47, 593–628 CrossRef CAS PubMed.
- E. Poggiogalle, H. Jamshed and C. M. Peterson, Circadian Regulation of Glucose, Lipid, and Energy Metabolism in Humans HHS Public Access, Metabolism, 2018, 84, 11–27 CrossRef CAS PubMed.
- S. M. Reppert and D. R. Weaver, Coordination of circadian timing in mammals, Nature, 2002, 418, 935–941 CrossRef CAS PubMed.
- J. S. Takahashi, Transcriptional architecture of the mammalian circadian clock, Nat. Rev. Genet., 2016, 18, 164–179 CrossRef PubMed.
- N. Preitner, F. Damiola, L. Lopez-Molina, J. Zakany, D. Duboule, U. Albrecht and U. Schibler, The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator, Cell, 2002, 110, 251–260 CrossRef CAS PubMed.
- B. H. Miller, E. L. McDearmon, S. Panda, K. R. Hayes, J. Zhang, J. L. Andrews, M. P. Antoch, J. R. Walker, K. A. Esser, J. B. Hogenesch and J. S. Takahashi, Circadian and CLOCK-controlled regulation of the mouse transcriptome and cell proliferation, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 3342–3347 CrossRef CAS PubMed.
- U. Schibler, J. Ripperger and S. A. Brown, Peripheral circadian oscillators in mammals: time and food, J. Biol. Rhythms, 2003, 18, 250–260 CrossRef PubMed.
- C. Dibner, U. Schibler and U. Albrecht, The Mammalian Circadian Timing System: Organization and Coordination of Central and Peripheral Clocks, Annu. Rev. Physiol., 2010, 72, 517–549 CrossRef CAS PubMed.
- T. H. Monk, Enhancing Circadian Zeitgebers, Sleep, 2010, 33, 421 CrossRef PubMed.
- A. Hirao, Y. Tahara, I. Kimura and S. Shibata, A Balanced Diet Is Necessary for Proper Entrainment Signals of the Mouse Liver Clock, PLoS One, 2009, 4, e6909 CrossRef PubMed.
- M. Barnea, Z. Madar and O. Froy, High-fat diet followed by fasting disrupts circadian expression of adiponectin signaling pathway in muscle and adipose tissue, Obesity, 2010, 18, 230–238 CrossRef CAS PubMed.
- J. Mendoza, P. Pévet and E. Challet, High-fat feeding alters the clock synchronization to light, J. Physiol., 2008, 586, 5901 CrossRef CAS PubMed.
- A. Kohsaka, A. D. Laposky, K. M. Ramsey, C. Estrada, C. Joshu, Y. Kobayashi, F. W. Turek and J. Bass, High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice, Cell Metab., 2007, 6, 414–421 CrossRef CAS PubMed.
- J. C. Walton, J. R. Bumgarner and R. J. Nelson, Sex Differences in Circadian Rhythms, Cold Spring Harbor Perspect. Biol., 2022, 14, a039107 CrossRef CAS PubMed.
- J. Qian, C. J. Morris, R. Caputo, W. Wang, M. Garaulet and F. A. J. L. Scheer, Sex differences in the circadian misalignment effects on energy regulation, Proc. Natl. Acad. Sci. U. S. A., 2019, 116, 23806–23812 CrossRef CAS PubMed.
- C. Zhang, C. Tait, C. D. Minacapelli, A. Bhurwal, K. Gupta, R. Amin and V. K. Rustgi, The Role of Race, Sex, and Age in Circadian Disruption and Metabolic Disorders, Gastro Hep Advances, 2022, 1, 471–479 CrossRef.
- M. Pérez-Mendoza, J. B. Rivera-Zavala, A. H. Rodríguez-Guadarrama, L. M. Montoya-Gomez, A. Carmona-Castro, M. Díaz-Muñoz and M. Miranda-Anaya, Daily cycle in hepatic lipid metabolism in obese mice, Neotomodon alstoni: Sex differences, Chronobiol. Int., 2018, 35, 643–657 CrossRef PubMed.
- M. Ruiter, S. E. La Fleur, C. Van Heijningen, J. Van der Vliet, A. Kalsbeek and R. M. Buijs, The Daily Rhythm in Plasma Glucagon Concentrations in the Rat Is Modulated by the Biological Clock and by Feeding Behavior, Diabetes, 2003, 52, 1709–1715 CrossRef CAS PubMed.
- M. Perelis, B. Marcheva, K. M. Ramsey, M. J. Schipma, A. L. Hutchison, A. Taguchi, C. B. Peek, H. Hong, W. Huang, C. Omura, A. L. Allred, C. A. Bradfield, A. R. Dinner, G. D. Barish and J. Bass, Pancreatic β cell enhancers regulate rhythmic transcription of genes controlling insulin secretion, Science, 2015, 350(6261), aac4250 CrossRef PubMed.
- S. M. Bailey, U. S. Udoh and M. E. Young, Circadian regulation of metabolism, J. Endocrinol., 2014, 222(2), R75–R96 CAS.
- G. Le Martelot, T. Claudel, D. Gatfield, O. Schaad, B. Kornmann, G. Lo Sasso, A. Moschetta and U. Schibler, REV-ERBalpha participates in circadian SREBP signaling and bile acid homeostasis, PLoS Biol., 2009, 7(9), e1000181 CrossRef PubMed.
- H. Jurevics, J. Hostettler, C. Barrett, P. Morell and A. D. Toews, Diurnal and dietary-induced changes in cholesterol synthesis correlate with levels of mRNA for HMG-CoA reductase, J. Lipid Res., 2000, 41, 1048–1053 CrossRef CAS.
- J. Kovář, M. Leníček, M. Zimolová, L. Vítek, M. Jirsa and J. Piťha, Regulation of diurnal variation of cholesterol 7alpha-hydroxylase (CYP7A1) activity in healthy subjects, Physiol. Res., 2010, 59, 233–238 Search PubMed.
- M. Noshiro, E. Usui, T. Kawamoto, H. Kubo, K. Fujimoto, M. Furukawa, S. Honma, M. Makishima, K. I. Honma and Y. Kato, Multiple mechanisms regulate circadian expression of the gene for cholesterol 7alpha-hydroxylase (Cyp7a), a key enzyme in hepatic bile acid biosynthesis, J. Biol. Rhythms, 2007, 22, 299–311 CrossRef CAS PubMed.
- L. Fan, P. N. Hsieh, D. R. Sweet and M. K. Jain, Krüppel-like factor 15: Regulator of BCAA metabolism and circadian protein rhythmicity, Pharmacol. Res., 2018, 130, 123–126 CrossRef CAS PubMed.
- R. Ch, O. Chevallier and C. T. Elliott, Metabolomics reveal circadian control of cellular metabolism, TrAC, Trends Anal. Chem., 2020, 130, 115986 CrossRef CAS.
- S. Abbondante, K. L. Eckel-Mahan, N. J. Ceglia, P. Baldi and P. Sassone-Corsi, Comparative Circadian Metabolomics Reveal Differential Effects of Nutritional Challenge in the Serum and Liver, J. Biol. Chem., 2016, 291, 2812–2828 CrossRef CAS PubMed.
- C. Bladé, G. Aragonès, A. Arola-Arnal, B. Muguerza, F. I. Bravo, M. J. Salvadó, L. Arola and M. Suárez, Proanthocyanidins in health and disease, BioFactors, 2016, 42, 5–12 Search PubMed.
- C. Bladé, L. Arola and M. J. Salvadó, Hypolipidemic effects of proanthocyanidins and their underlying biochemical and molecular mechanisms., Mol. Nutr. Food Res., 2010, 54, 37–59 CrossRef PubMed.
- Z. Pons, M. Margalef, F. I. Bravo, A. Arola-Arnal and B. Muguerza, Acute administration of single oral dose of grape seed polyphenols restores blood pressure in a rat model of metabolic syndrome: role of nitric oxide and prostacyclin, Eur. J. Nutr., 2016, 55, 749–758 CrossRef CAS PubMed.
- B. V. Nemzer, F. Al-Taher, A. Yashin, I. Revelsky and Y. Yashin, Cranberry: Chemical Composition, Antioxidant Activity and Impact on Human Health: Overview, Molecules, 2022, 27, 1503 CrossRef CAS PubMed.
- Y. X. Zeng, S. Wang, L. Wei, Y. Y. Cui and Y. H. Chen, Proanthocyanidins: Components, Pharmacokinetics and Biomedical Properties, Am. J. Chin. Med., 2020, 48, 813–869 CrossRef CAS PubMed.
- A. Pascual-Serrano, A. Arola-Arnal, S. Suárez-García, F. I. Bravo, M. Suárez, L. Arola and C. Bladé, Grape seed proanthocyanidin supplementation reduces adipocyte size and increases adipocyte number in obese rats, Int. J. Obes., 2017, 41, 1246–1255 CrossRef CAS PubMed.
- G. Montagut, C. Bladé, M. Blay, J. Fernández-Larrea, G. Pujadas, M. J. Salvadó, L. Arola, M. Pinent and A. Ardévol, Effects of a grapeseed procyanidin extract (GSPE) on insulin resistance, J. Nutr. Biochem., 2010, 21, 961–967 CrossRef CAS PubMed.
- A. Caimari, J. M. Del Bas, A. Crescenti and L. Arola, Low doses of grape seed procyanidins reduce adiposity and improve the plasma lipid profile in hamsters, Int. J. Obes., 2012, 37, 576–583 CrossRef PubMed.
- D. Bagchi, C. K. Sen, S. D. Ray, D. K. Das, M. Bagchi, H. G. Preuss and J. A. Vinson, Molecular mechanisms of cardioprotection by a novel grape seed proanthocyanidin extract, Mutat. Res., Fundam. Mol. Mech. Mutagen., 2003, 523–524, 87–97 CrossRef CAS PubMed.
- M. Monagas, M. Urpi-Sarda, F. Sánchez-Patán, R. Llorach, I. Garrido, C. Gómez-Cordovés, C. Andres-Lacueva and B. Bartolomé, Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites, Food Funct., 2010, 1, 233–253 RSC.
- F. Cardona, C. Andrés-Lacueva, S. Tulipani, F. J. Tinahones and M. I. Queipo-Ortuño, J. Nutr. Biochem., 2013, 24, 1415–1422 CrossRef CAS PubMed.
- A. Ribas-Latre, L. Baselga-Escudero, E. Casanova, A. Arola-Arnal, M. J. Salvadó, C. Bladé and L. Arola, Dietary proanthocyanidins modulate BMAL1 acetylation, Nampt expression and NAD levels in rat liver, Sci. Rep., 2015, 5, 1–13 Search PubMed.
- A. Ribas-Latre, J. M. Del Bas, L. Baselga-Escudero, E. Casanova, A. Arola-Arnal, M. J. Salvadó, L. Arola and C. Bladé, Dietary proanthocyanidins modulate melatonin levels in plasma and the expression pattern of clock genes in the hypothalamus of rats, Mol. Nutr. Food Res., 2015, 59, 865–878 CrossRef CAS PubMed.
- A. Ribas-Latre, L. Baselga-Escudero, E. Casanova, A. Arola-Arnal, M. J. Salvadó, L. Arola and C. Bladé, Chronic consumption of dietary proanthocyanidins modulates peripheral clocks in healthy and obese rats, J. Nutr. Biochem., 2015, 26, 112–119 CrossRef CAS PubMed.
- R. M. Rodríguez, A. J. Cortés-Espinar, J. R. Soliz-Rueda, C. Feillet-Coudray, F. Casas, M. Colom-Pellicer, G. Aragonès, J. Avila-Román, B. Muguerza, M. Mulero and M. J. Salvadó, Time-of-Day Circadian Modulation of Grape-Seed Procyanidin Extract (GSPE) in Hepatic Mitochondrial Dynamics in Cafeteria-Diet-Induced Obese Rats, Nutrients, 2022, 14, 774 CrossRef PubMed.
- M. Colom-Pellicer, R. M. Rodríguez, È. Navarro-Masip, F. I. Bravo, M. Mulero, L. Arola and G. Aragonès, Time-of-day dependent effect of proanthocyanidins on adipose tissue metabolism in rats with diet-induced obesity, Int. J. Obes., 2022, 46, 1394–1402 CrossRef CAS PubMed.
- I. Escobar-Martínez, V. Arreaza-Gil, B. Muguerza, A. Arola-Arnal, F. I. Bravo, C. Torres-Fuentes and M. Suárez, Administration Time Significantly Affects Plasma Bioavailability of Grape Seed Proanthocyanidins Extract in Healthy and Obese Fischer 344 Rats, Mol. Nutr. Food Res., 2022, 66, e2100552 CrossRef PubMed.
- M. Margalef, Z. Pons, L. Iglesias-Carres, L. Arola, B. Muguerza and A. Arola-Arnal, Gender-related similarities and differences in the body distribution of grape seed flavanols in rats, Mol. Nutr. Food Res., 2016, 60, 760–772 CrossRef CAS PubMed.
- K. J. Livak and T. D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method, Methods, 2001, 25, 402–408 CrossRef CAS PubMed.
- H. Palacios-Jordan, M. Z. Martín-González, M. Suárez, G. Aragonès, B. Mugureza, M. A. Rodríguez and C. Bladé, The Disruption of Liver Metabolic Circadian Rhythms by a Cafeteria Diet Is Sex-Dependent in Fischer 344 Rats, Nutrients, 2020, 12, 1085 CrossRef CAS PubMed.
- Z. Pang, J. Chong, G. Zhou, D. A. De Lima Morais, L. Chang, M. Barrette, C. Gauthier, P. É. Jacques, S. Li and J. Xia, MetaboAnalyst 5.0: narrowing the gap between raw spectra and functional insights, Nucleic Acids Res., 2021, 49, W388–W396 CrossRef CAS PubMed.
- J. Ávila-Román, V. Arreaza-Gil, A. J. Cortés-Espinar, J. R. Soliz-Rueda, M. Mulero, B. Muguerza, A. Arola-Arnal, L. Arola and C. Torres-Fuentes, Impact of gut microbiota on plasma oxylipins profile under healthy and obesogenic conditions, Clin. Nutr., 2021, 40, 1475–1486 CrossRef PubMed.
- A. Pascual-Serrano, C. Bladé, M. Suárez and A. Arola-Arnal, Grape Seed Proanthocyanidins Improve White Adipose Tissue Expansion during Diet-Induced Obesity Development in Rats, Int. J. Mol. Sci., 2018, 19, 2632 CrossRef PubMed.
- X. Terra, G. Montagut, M. Bustos, N. Llopiz, A. Ardèvol, C. Bladé, J. Fernández-Larrea, G. Pujadas, J. Salvadó, L. Arola and M. Blay, Grape-seed procyanidins prevent low-grade inflammation by modulating cytokine expression in rats fed a high-fat diet, J. Nutr. Biochem., 2009, 20, 210–218 CrossRef CAS PubMed.
- A. Shetty, J. W. Hsu, P. P. Manka and W. K. Syn, Role of the Circadian Clock in the Metabolic Syndrome and Nonalcoholic Fatty Liver Disease, Dig. Dis. Sci., 2018, 63, 3187–3206 CrossRef CAS PubMed.
- F. Saura-Calixto, J. Serrano and I. Goñi, Intake and bioaccessibility of total polyphenols in a whole diet, Food Chem., 2007, 101, 492–501 CrossRef CAS.
- Y. Wang, S. J. Chung, W. O. Song and O. K. Chun, Estimation of Daily Proanthocyanidin Intake and Major Food Sources in the U.S. Diet, J. Nutr., 2011, 141, 447–452 CrossRef CAS PubMed.
- R. Zamora-Ros, C. Andres-Lacueva, R. M. Lamuela-Raventós, T. Berenguer, P. Jakszyn, A. Barricarte, E. Ardanaz, P. Amiano, M. Dorronsoro, N. Larrañaga, C. Martínez, M. J. Sánchez, C. Navarro, M. D. Chirlaque, M. J. Tormo, J. R. Quirós and C. A. González, Estimation of Dietary Sources and Flavonoid Intake in a Spanish Adult Population (EPIC-Spain), J. Am. Diet. Assoc., 2010, 110, 390–398 CrossRef CAS PubMed.
- S. Reagan-Shaw, M. Nihal and N. Ahmad, Dose translation from animal to human studies revisited, FASEB J., 2008, 22, 659–661 CrossRef CAS PubMed.
- C. M. Isherwood, D. R. Van der Veen, J. D. Johnston and D. J. Skene, Twenty-four-hour rhythmicity of circulating metabolites: effect of body mass and type 2 diabetes, FASEB J., 2017, 31, 5557–5567 CrossRef CAS PubMed.
- P. Zimmet, K. G. M. M. Alberti, N. Stern, C. Bilu, A. El-Osta, H. Einat and N. Kronfeld-Schor, The Circadian Syndrome: is the Metabolic Syndrome and much more!, J. Intern. Med., 2019, 286, 181–191 CrossRef CAS PubMed.
- M. L. Power and J. Schulkin, Sex differences in fat storage, fat metabolism, and the health risks from obesity: possible evolutionary origins, Br. J. Nutr., 2008, 99, 931–940 CrossRef CAS PubMed.
- J. R. B. Tibaes, J. Azarcoya-Barrera, B. Wollin, H. Veida-Silva, A. Makarowski, D. Vine, S. Tsai, R. Jacobs and C. Richard, Sex Differences Distinctly Impact High-Fat Diet-Induced Immune Dysfunction in Wistar Rats, J. Nutr., 2022, 152, 1347–1357 CrossRef PubMed.
- V. Carmona-Alcocer, C. Fuentes-Granados, A. Carmona-Castro, I. Aguilar-González, R. Cárdenas-Vázquez and M. Miranda-Anaya, Obesity alters circadian behavior and metabolism in sex dependent manner in the volcano mouse Neotomodon alstoni, Physiol. Behav., 2012, 105, 727–733 CrossRef CAS PubMed.
- P. C. Kandalepas, J. W. Mitchell and M. U. G. Gillette, Melatonin Signal Transduction Pathways Require E-Box-Mediated Transcription of Per1 and Per2 to Reset the SCN Clock at Dusk, PLoS One, 2016, 11, e0157824 CrossRef PubMed.
- D. J. Skene and J. Arendt, Human circadian rhythms: physiological and therapeutic relevance of light and melatonin, Ann. Clin. Biochem., 2006, 43, 344–353 CrossRef CAS PubMed.
- M. C. Tackenberg and D. G. McMahon, Photoperiodic Programming of the SCN and Its Role in Photoperiodic Output, Neural Plast., 2018, 2018, 8217345 Search PubMed.
- C. Von Gall, J. H. Stehle and D. R. Weaver, Mammalian melatonin receptors: Molecular biology and signal transduction, Cell Tissue Res., 2002, 309, 151–162 CrossRef CAS PubMed.
- M. Bailey and R. Silver, Sex differences in circadian timing systems: implications for disease, Front. Neuroendocrinol., 2014, 35, 111–139 CrossRef PubMed.
- A. Malazonia, T. Zerekidze, E. Giorgadze, N. Chkheidze and K. Asatiani, Melatonin Level Variations with Different Behavioural Risk Factors in Obese Female Patients, Open Access Maced. J. Med. Sci., 2017, 5, 613–617 CrossRef PubMed.
- L. Brennan and H. Gibbons, Sex matters: a focus on the impact of biological sex on metabolomic profiles and dietary interventions, Proc. Nutr. Soc., 2020, 79, 205–209 CrossRef CAS PubMed.
- B. T. Palmisano, L. Zhu, R. H. Eckel and J. M. Stafford, Sex differences in lipid and lipoprotein metabolism, Mol. Metab., 2018, 15, 45 CrossRef CAS PubMed.
- S. Della Torre, N. Mitro, C. Meda, F. Lolli, S. Pedretti, M. Barcella, L. Ottobrini, D. Metzger, D. Caruso and A. Maggi, Short-Term Fasting Reveals Amino Acid Metabolism as a Major Sex-Discriminating Factor in the Liver, Cell Metab., 2018, 28, 256–267 CrossRef CAS PubMed.
- L. S. Lamont, A. J. McCullough and S. C. Kalhan, Gender differences in the regulation of amino acid metabolism, J. Appl. Physiol., 2003, 95, 1259–1265 CrossRef CAS PubMed.
- K. D. Tipton, Gender differences in protein metabolism, Curr. Opin. Clin. Nutr. Metab. Care, 2001, 4, 493–498 CrossRef CAS PubMed.
- A. Agus, J. Planchais and H. Sokol, Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease, Cell Host Microbe, 2018, 23, 716–724 CrossRef CAS PubMed.
- K. Gao, C. L. Mu, A. Farzi and W. Y. Zhu, Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain, Adv. Nutr., 2020, 11, 709 CrossRef PubMed.
- M. Kiluk, J. Lewkowicz, D. Pawlak and A. Tankiewicz-Kwedlo, Crosstalk between Tryptophan Metabolism via Kynurenine Pathway and Carbohydrate Metabolism in the Context of Cardio-Metabolic Risk—Review, J. Clin. Med., 2021, 10, 10 Search PubMed.
- K. B. Koronowski, K. Kinouchi, P. S. Welz, J. G. Smith, V. M. Zinna, J. Shi, M. Samad, S. Chen, C. N. Magnan, J. M. Kinchen, W. Li, P. Baldi, S. A. Benitah and P. Sassone-Corsi, Defining the Independence of the Liver Circadian Clock, Cell, 2019, 177, 1448–1462 CrossRef CAS PubMed.
- T. Zelante, R. G. Iannitti, F. Fallarino, M. Gargaro, A. de Luca, S. Moretti, A. Bartoli and L. Romani, Tryptophan Feeding of the IDO1-AhR Axis in Host-Microbial Symbiosis, Front. Immunol., 2014, 5, 640 Search PubMed.
- P. Petrus, M. Cervantes, M. Samad, T. Sato, A. Chao, S. Sato, K. B. Koronowski, G. Park, Y. Alam, N. Mejhert, M. M. Seldin, J. M. Monroy Kuhn, K. A. Dyar, D. Lutter, P. Baldi, P. Kaiser, C. Jang and P. Sassone-Corsi, Tryptophan metabolism is a physiological integrator regulating circadian rhythms, Mol. Metab., 2022, 64, 101556 CrossRef CAS PubMed.
- Y. Tahara, M. Yamazaki, H. Sukigara, H. Motohashi, H. Sasaki, H. Miyakawa, A. Haraguchi, Y. Ikeda, S. Fukuda and S. Shibata, Gut Microbiota-Derived Short Chain Fatty Acids Induce Circadian Clock Entrainment in Mouse Peripheral Tissue, Nat. Sci. Rep., 2018, 8, 1395 Search PubMed.
- C. Torres-Fuentes, H. Schellekens, T. G. Dinan and J. F. Cryan, The microbiota–gut–brain axis in obesity, Lancet Gastroenterol. Hepatol., 2017, 2, 747–756 CrossRef PubMed.
- W. Liu, S. Zhao, J. Wang, J. Shi, Y. Sun, W. Wang, G. Ning, J. Hong and R. Liu, Grape seed proanthocyanidin extract ameliorates inflammation and adiposity by modulating gut microbiota in high-fat diet mice, Mol. Nutr. Food Res., 2017, 61, 1–14 Search PubMed.
- M. Colom-Pellicer, R. M. Rodríguez, J. R. Soliz-Rueda, L. V. M. de Assis, È. Navarro-Masip, S. Quesada-Vázquez, X. Escoté, H. Oster, M. Mulero and G. Aragonès, Proanthocyanidins Restore the Metabolic Diurnal Rhythm of Subcutaneous White Adipose Tissue According to Time-Of-Day Consumption, Nutrients, 2022, 14, 2246 CrossRef CAS PubMed.
- E. Challet, The circadian regulation of food intake, Nat. Rev. Endocrinol., 2019, 15, 393–405 CrossRef PubMed.
- O. Fiehn, W. Timothy Garvey, J. W. Newman, K. H. Lok, C. L. Hoppel and S. H. Adams, Plasma metabolomic profiles reflective of glucose homeostasis in non-diabetic and type 2 diabetic obese African-American women, PLoS One, 2010, 5, 1–10 CrossRef PubMed.
- P. Wurtz, P. Soininen, A. J. Kangas, T. Rönnemaa, T. Lehtimäki, M. Kähönen, J. S. Viikari, O. T. Raitakari and M. Ala-Korpela, Branched-chain and aromatic amino acids are predictors of insulin resistance in young adults, Diabetes Care, 2013, 36, 648–655 CrossRef CAS PubMed.
- D. Gnocchi, M. Pedrelli, M. Pedrelli, E. Hurt-Camejo and P. Parini, Lipids around the Clock: Focus on Circadian Rhythms and Lipid Metabolism, Biology, 2015, 4, 104–132 CrossRef CAS PubMed.
- G. Le Martelot, T. Claudel, D. Gatfield, O. Schaad, B. Kornmann, G. Lo Sasso, A. Moschetta and U. Schibler, REV-ERBα Participates in Circadian SREBP Signaling and Bile Acid Homeostasis, PLoS Biol., 2009, 7, e1000181 CrossRef PubMed.
- J. Y. L. Chiang, Bile acids: regulation of synthesis, J. Lipid Res., 2009, 50, 1955–1966 CrossRef CAS PubMed.
- Y. K. J. Zhang, G. L. Guo and C. D. Klaassen, Diurnal Variations of Mouse Plasma and Hepatic Bile Acid Concentrations as well as Expression of Biosynthetic Enzymes and Transporters, PLoS One, 2011, 6, e16683 CrossRef CAS PubMed.
- Y. Yang and J. Zhang, Bile acid metabolism and circadian rhythms, Am. J. Physiol.: Gastrointest. Liver Physiol., 2020, 319, G549–G563 CrossRef CAS PubMed.
- Z. Yu, J. Yang, D. Xiang, G. Li, D. Liu and C. Zhang, Circadian rhythms and bile acid homeostasis: a comprehensive review, Chronobiol. Int., 2020, 37, 618–628 CrossRef CAS PubMed.
- M. Wei, F. Huang, L. Zhao, Y. Zhang, W. Yang, S. Wang, M. Li, X. Han, K. Ge, C. Qu, C. Rajani, G. Xie, X. Zheng, A. Zhao, Z. Bian and W. Jia, A dysregulated bile acid-gut microbiota axis contributes to obesity susceptibility, EBioMedicine, 2020, 55, 102766 CrossRef PubMed.
- E. M. Teichman, K. J. O'Riordan, C. G. M. Gahan, T. G. Dinan and J. F. Cryan, Cell Metab., 2020, 31, 448–471 CrossRef CAS PubMed.
- V. Arreaza-Gil, I. Escobar-Martínez, M. Suárez, F. Bravo, B. Muguerza, A. Arola-Arnal and C. Torres-Fuentes, Gut Seasons: Photoperiod Effects on Fecal Microbiota in Healthy and Cafeteria-Induced Obese Fisher 344 Rats, Nutrients, 2022, 14, 722 CrossRef CAS PubMed.
- M. Guirro, A. Costa, A. Gual-Grau, P. Herrero, H. Torrell, N. Canela and L. Arola, Effects from diet-induced gut microbiota dysbiosis and obesity can be ameliorated by fecal microbiota transplantation: A multiomics approach, PLoS One, 2019, 14, e0218143 CrossRef CAS PubMed.
- Y. S. Kim, T. Unno, B. Y. Kim and M. S. Park, Sex Differences in Gut Microbiota, World J. Men's Health, 2020, 38, 48 CrossRef PubMed.
- Y. Cui, Y. Yin, S. Li, Z. Wu, Y. Xie, Q. Qian, H. Yang and X. Li, Apple polyphenol extract modulates bile acid metabolism and gut microbiota by regulating the circadian rhythms in daytime-restricted high fat diet feeding C57BL/6 male mice, Food Funct., 2022, 13, 2805–2822 RSC.
- C. N. Serhan, S. D. Brain, C. D. Buckley, D. W. Gilroy, C. Haslett, L. A. J. O'Neill, M. Perretti, A. G. Rossi and J. L. Wallace, Resolution of in flammation: state of the art, definitions and terms, FASEB J., 2007, 21, 325–332 CrossRef CAS PubMed.
- V. Cecarini, M. Cuccioloni, Y. Zheng, L. Bonfili, C. Gong, M. Angeletti, P. Mena, D. Del Rio and A. M. Eleuteri, Flavan-3-ol Microbial Metabolites Modulate Proteolysis in Neuronal Cells Reducing Amyloid-beta (1–42) Levels, Mol. Nutr. Food Res., 2021, 65, e2100380 CrossRef PubMed.
- P. Mena, L. Bresciani, N. Brindani, I. A. Ludwig, G. Pereira-Caro, D. Angelino, R. Llorach, L. Calani, F. Brighenti, M. N. Clifford, C. I. R. Gill, A. Crozier, C. Curti and D. Del Rio, Phenyl-γ-valerolactones and phenylvaleric acids, the main colonic metabolites of flavan-3-ols: synthesis, analysis, bioavailability, and bioactivity, Nat. Prod. Rep., 2019, 36, 714–752 RSC.
- M. Heddes, B. Altaha, Y. Niu, S. Reitmeier, K. Kleigrewe, D. Haller and S. Kiessling, The intestinal clock drives the microbiome to maintain gastrointestinal homeostasis, Nat. Commun., 2022, 13, 1–17 Search PubMed.
- B. D. Weger, C. Gobet, J. Yeung, E. Martin, S. Jimenez, B. Betrisey, F. Foata, B. Berger, A. Balvay, A. Foussier, A. Charpagne, B. Boizet-Bonhoure, C. J. Chou, F. Naef and F. Gachon, The Mouse Microbiome Is Required for Sex-Specific Diurnal Rhythms of Gene Expression and Metabolism, Cell Metab., 2019, 29, 362–382 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3fo01551c |
| ‡ These authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2023 |