
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Lactobacillus rhamnosus GG administration partially prevents diet-induced insulin resistance in rats: a comparison with its heat-inactivated parabiotic
L.
Arellano-García
 a,
M. T.
Macarulla
abc,
A.
Cuevas-Sierra
d,
J. A.
Martínez
bd,
M. P.
Portillo
*abc and
I.
Milton-Laskibar
abc
a,
M. T.
Macarulla
abc,
A.
Cuevas-Sierra
d,
J. A.
Martínez
bd,
M. P.
Portillo
*abc and
I.
Milton-Laskibar
abc
aNutrition and Obesity Group, Department of Nutrition and Food Science, Faculty of Pharmacy and Lucio Lascaray Research Centre, University of the Basque Country (UPV/EHU), 01006 Vitoria-Gasteiz, Spain. E-mail: mariapuy.portillo@ehu.eus
bCIBERobn Physiopathology of Obesity and Nutrition, Institute of Health Carlos III, 28222 Madrid, Spain
cBIOARABA Health Research Institute, 01006 Vitoria-Gasteiz, Spain
dPrecision Nutrition and Cardiometabolic Health, IMDEA-Food Institute (Madrid Institute for Advanced Studies), Campus of International Excellence (CEI) UAM+CSIC, Spanish National Research Council, 28049 Madrid, Spain
First published on 2nd September 2023
Abstract
Insulin resistance and type 2 diabetes are obesity-related health alterations, featuring an ever-increasing prevalence. Besides inadequate feeding patterns, gut microbiota alterations stand out as potential contributors to these metabolic disturbances. The aim of this study was to investigate whether the administration of a probiotic (Lactobacillus rhamnosus GG) effectively prevents diet-induced insulin resistance in rats and to compare these potential effects with those exerted by its heat-inactivated parabiotic. For this purpose, 34 male Wistar rats were fed a standard or a high-fat high-fructose diet, alone or supplemented with viable or heat-inactivated Lactobacillus rhamnosus GG. The body and white adipose tissue weight increases, induced by the obesogenic diet, were prevented by probiotic and parabiotic administration. The trend towards higher basal glucose levels and significantly higher serum insulin concentration observed in the non-treated animals fed with the obesogenic diet were effectively reverted by both treatments. Similar results were also found for serum adiponectin and leptin, whose levels were brought back by the probiotic and parabiotic administration to values similar to those of the control animals. Noteworthily, parabiotic administration significantly reduced skeletal muscle triglyceride content and activated CPT-1b compared to the non-treated animals. Finally, both treatments enhanced Akt and AS160 phosphorylation in the skeletal muscle compared to the non-treated animals; however, only parabiotic administration increased GLUT-4 protein expression in this tissue. These results suggest that heat-inactivated Lactobacillus rhamnosus GG seem to be more effective than its probiotic of origin in preventing high-fat high-fructose diet-induced insulin resistance in rats.
1. Introduction
Obesity, considered as an abnormal or excessive fat accumulation that may impair health, has reached pandemic dimensions, whose prevalence doubling data collected in the 1980s.1 The metabolic implications of an enlarged adipose tissue mass rely on an increased risk for suffering obesity-related co-morbidities, such as insulin resistance/type 2 diabetes, cardiovascular diseases, or non-alcoholic fatty liver disease (NAFLD), especially when excessive adiposity occurs in abdominal depots.2 The close association between obesity and glucose homeostasis impairments is reflected by the high prevalence of insulin resistance and type 2 diabetes mellitus among subjects with obesity (between 51 and 99%).3 One of the main triggering factors of this metabolic disturbance is an unhealthy dietary pattern, such as the so-called western diet (WD), characterized by a high content of saturated fats and added sugars along with a low content of fruits and vegetables.4 Indeed, the high content of added fructose has been shown to be especially harmful in terms of metabolic alterations,5 enhancing the levels of reactive oxygen species (ROS) and increasing oxidative stress (OS).Altered gut microbiota composition has also been identified as an additional associated factor.6 In this line, gut microbiota, the complex community of microorganisms residing in our gastrointestinal tract (mainly made up of bacteria) plays a major role in the maintenance of homeostasis, regulating important processes, such as nutrient digestion and utilization, the synthesis of certain vitamins, regulation of immune system, or production of bioactive metabolites.7 In the case of obese subjects featuring type 2 diabetes, the gut microbiota of these individuals is characterized by a lower microbial diversity, increased Firmicutes and decreased Bacteroidetes relative abundances, a lower abundance of butyrate-producing bacteria, and an enhanced production of proinflammatory mediators.8 Noteworthy, diet is a major factor defining gut microbiota composition. Thus, diets containing high amounts of fats and added sugars have been related to impairments in gut microbiota composition.9
Due to the close relationship between obesity and insulin resistance/type 2 diabetes, the conventional treatment prescribed for these metabolic alterations is based on lifestyle modification (hypocaloric diets and the promotion of physical activity).10 Despite proven valid in some cases, the main limitation of this approach is the low adherence in patients, especially in the long term. As a consequence, the scientific community is constantly looking for additional/alternative therapeutic tools. Given the close relationship between altered gut microbiota composition in the onset and development of insulin resistance and diabetes, probiotic administration has been proposed as a potential approach to tackle these metabolic disturbances by targeting gut microbiota.11 Probiotics are live microorganisms that, when administered in appropriate amounts, exert beneficial health effects in the host.12 Several studies have addressed the usefulness of probiotic administration for type 2 diabetes management in well-designed preclinical and clinical trials.13–15 However, probiotics administration may not always be suitable since its administration to vulnerable subjects may cause systemic infections, deleterious metabolic effects, excessive immune activation on the host, or gene transferring (especially antibiotic resistance).16 Moreover, the safety and viability of microorganisms, which should be considered if probiotic administration is desirable (bacteria have to be kept alive), may not be assured due to the industrial procedures and storage conditions.
In view of the challenges that the maintenance and administration of a probiotic entails, there is major interest in the potential therapeutic effects of inactivated bacteria, also referred to as parabiotics. According to the FAO/WHO, parabiotics are defined as “inactivated (non-viable) microbial cells that, when administered in sufficient amounts, confer benefits to consumers”.17 Parabiotics alternative of interest in terms of administration risks and industrial handling. Interestingly, different studies have described the anti-inflammatory and anti-oxidative potential for heat-inactivated Lactobacillus strains in in vitro and in vivo models.18 However, the available literature regarding the potential usage of parabiotics in the management of metabolic disturbances is still scarce.
Another alternative is the use of postbiotics, which include certain probiotic-derived components, such as bacteriocins, lipoteichoic acids, surface layer protein, and secreted protein. Increasing evidence exists supporting that these postbiotics show beneficial effects on health, such as a protective role on intestinal barrier function or immunomodulation, similar to those shown by live probiotics.19,20 On the other hand, the use of postbiotics may be a better candidate for the clinical usage than the whole bacteria since it does not contain other bacterial components that may cause various side effects.21
In this scenario, the present study aims to investigate whether the administration of a probiotic (Lactobacillus rhamnosus GG) is effective in preventing diet-induced insulin resistance in rats and compare these potential effects with those exerted by its heat-inactivated parabiotic. Additionally, some putative mechanisms of action involved in the observed effects were also investigated.
2. Materials and methods
2.1. Animals, diets, and experimental design
All animal procedures were performed in accordance with the Guidelines for Care and Use of Laboratory Animals of the University of the Basque Country. The experimental protocol used in this study was approved by the Ethics Committee on Animal Experimentation (CEEA) of the University of the Basque Country (under reference M20/2021/214). The experiment was performed on thirty-four 8–9-week-old male Wistar rats (Envigo, Barcelona, Spain), which were housed in conventional polycarbonate cages (two rats per cage) and placed in an air-conditioned room (22 ± 2 °C) with a 12 h light–dark cycle. After a 6-day adaptation period, rats were allocated into four different experimental groups. One group of animals was fed a standard laboratory diet (C group; n = 8) (D10012G; Research Diets, New Brunswick, NJ, USA). The rest of the groups were fed a high-fat high-fructose diet, which provided 4.49 kcal g−1, 40% as fat, 20% as protein, and 40% as carbohydrates (10% as fructose) (D21052401; Research Diets, New Brunswick, NJ, USA) (Table 1). Animals fed with the obesogenic diet received either the diet alone (HFHF group; n = 8) or supplemented with a commercially available probiotic (Lactobacillus rhamnosus GG, Ferring Pharmaceuticals, Switzerland) (PRO group; n = 9), or with its parabiotic (non-viable, heat-inactivated probiotic) (PARA group; n = 9). The former was diluted in phosphate buffered saline (PBS) containing 5% sucrose, in an amount ensuring a dose of 109 CFU per day. As for the parabiotic, the same probiotic and sucrose-containing PBS dilution was used (ensuring a dose of 109 CFU per day), which was inactivated by heat-treatment (80 °C for 20 minutes), as explained in the literature.22–24 The animals in the C and the HFHF groups received sucrose-enriched PBS as the vehicle. Both the treatments and vehicle were given daily by oral gavage throughout the experimental period (six weeks). Animals had free access to food and water, and body weight and food intake were measured daily.| Carbohydrates | Fructose | Protein | Lipids | Total energy (kcal g−1) | |
|---|---|---|---|---|---|
| HFHF: high-fat high-fructose diet, STD: standard diet. | |||||
| STD | 63.9 | — | 20.3 | 15.8 | 3.9 |
| HFHF | 40 | 10 | 20 | 40 | 4.5 |
Basal glucose levels were assessed in the blood samples obtained from the tail vein one-week prior to sacrifice after overnight fasting using a glucometer (Medisense, Abingdon, UK) and blood glucose test strips (Optium Xceed, Abbott Diabetes Care). At the end of the experimental period, the animals were anesthetized (chloral hydrate) and sacrificed after fasting (8–12 h) by cardiac exsanguination. White adipose tissues (epididymal, perirenal, mesenteric and subcutaneous) and skeletal muscles (gastrocnemius muscle, soleus muscle and the thigh/quadriceps) were dissected, weighed, and immediately frozen in liquid nitrogen. Blood samples were centrifuged (1000g for 10 min, at 4 °C) for serum collection. All samples were stored at −80 °C until analysis.
2.2. Serum determinations
Commercially available spectrophotometric kits were used for triglyceride determination (Biosystems, Barcelona, Spain), and ELISA kits for insulin (Millipore, Darmstadt, Germany), leptin (Sigma Aldrich, St Louis, MO, USA), adiponectin (Millipore, Darmstadt, Germany), and FGF-21 (Merck KGaA, Darmstadt, Germany) measurements in serum samples.The homeostatic model assessment for insulin resistance (HOMA-IR) was calculated from basal insulin and glucose values using Matthews’ formula.23
| HOMA-IR = [fasting glucose (mmol L−1) × fasting insulin (mU L−1)]/22.5. |
The triglyceride and glucose index (TyG) was calculated as an additional marker of insulin resistance using the formula proposed by Simental-Mendía et al.24
| TyG = Ln (TG [mg dL−1] × glucose [mg dL−1]/2) |
2.3. Skeletal muscle triacylglycerol determination
Total lipids were extracted from skeletal muscle samples following the method described by Folch et al.25 Next, the lipid extract was dissolved in isopropanol and the triacylglycerol content was measured using a commercial kit (Spinreact, Barcelona, Spain).2.4. Enzymatic activities
The activity of the oxidative enzyme carnitine palmitoyl-transferase 1β (CPT-1β) was measured spectrophotometrically in the mitochondrial/peroxisomal fraction following the method described by Bieber et al.26 This enzyme regulates the entry of long chain fatty acylCoA into the mitochondria for their subsequent oxidation, and thus is commonly used as a marker of fatty acid oxidation.27 Briefly, 100–150 mg skeletal muscle samples were homogenized in 1.5 mL homogenization buffer (250 mM saccharose, 1 mM EDTA, 10 mM Tris-HCl, pH 7.4) and centrifuged at 700g for 10 min at 4 °C. Subsequently, supernatants were centrifuged at 1200g for 15 min at 4 °C. Pellets containing the mitochondrial fraction were resuspended in resuspension buffer (70 mM saccharose, 220 mM mannitol, 2 mM HEPES, pH 7.4). The protein content of the samples was determined by the Bradford method28 using bovine serum albumin as the standard. The activity of CPT-1β was assayed by measuring the released CoA-SH and expressed as nanomoles of coenzyme A formed per minute per milligram of protein.Citrate synthase (CS) activity measurement was also assessed spectrophotometrically by measuring the appearance of free CoA according to the method described by Srere.29 This enzyme is commonly referred to as a marker of mitochondrial density.30 Briefly, skeletal muscle samples were homogenized in 1.5 mL triethanolamine-HCl buffer (0.1 M). Homogenates were incubated for 2 min at 30.1 °C with acetyl-CoA, 1.01 mM DTNB, 10 mM oxalacetate, Triton X-100 (10%), and distilled water, and the absorbance was measured at 412 nm. The homogenates were then re-incubated for 5 min and readings were taken at the same wavelength. CS activity was expressed as CoA nanomoles formed per minute per milligram of protein. The protein content of the samples was determined by the Bradford method28 using bovine serum albumin as the standard.
2.5. Western blot
For insulin receptor β (IRβ), protein kinase B (Akt), Akt substrate of 160 kDa (AS160), glucose transporter 4 (GLUT-4) and alpha-tubulin (α-tubulin) protein quantification, gastrocnemius samples (200 mg) were homogenized in 1.5 mL cellular PBS (pH 7.4), containing protease inhibitors (100 mM phenylmethylsulphonyl fluoride and 100 mM iodoacetamide). The homogenates were centrifuged at 800g for 10 min at 4 °C. The protein concentration was measured by the Bradford method28 using bovine serum albumin as the standard.Immunoblot analyses were performed by loading 60 μg total protein from skeletal muscle extracts separated by electrophoresis in 7.5% or 12% SDS-polyacrylamide gels and transferred to polyvinylidene difluoride (PVDF) membranes (Merck, Darmstadt, Germany). The membranes were then blocked with 5% casein PBS-Tween buffer for 1.5 h at room temperature. Afterward, they were blotted with the appropriate antibodies overnight at 4 °C. Protein levels were measured via specific antibodies for IRβ (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1000; Santa Cruz Biotech, Dallas, TX, USA), Akt (1:1000; Santa Cruz Biotech, Dallas, TX, USA), AS160 (1:1000; Santa Cruz Biotech, Dallas, TX, USA), and GLUT-4 (1:1000; Santa Cruz Biotech, Dallas, TX, USA). Subsequently, membranes were incubated with polyclonal anti-mouse (1:5000; Santa Cruz Biotech, Dallas, TX, USA) for Akt, anti-goat (1:5000; Santa Cruz Biotech, Dallas, TX, USA) for GLUT-4 and anti-rabbit (1:5000; Santa Cruz Biotech, Dallas, TX, USA) for IRβ and AS160 antibodies for 2 h at room temperature. After antibody stripping, membranes were blocked and incubated with phosphorylated IRβ (Tyr 1162/1163) (1:1000; Santa Cruz Biotech, Dallas, TX, USA), phosphorylated Akt (Thr 473) (1:1000; Cell Signaling, Danvers, MA, USA), phosphorylated AS160 (Thr 642) (1:1000; Gene Tex, CA, USA), and α-tubulin (1:1000; Cell Signaling, Danvers, MA, USA). The bound antibodies were visualized by an ECL system (Thermo Fisher Scientific Inc., Rockford, IL, USA) and quantified by a ChemiDoc MP imaging system (Bio-Rad, Hercules, CA, USA). The measurements were normalized by either β-actin or the phosphorylated isoform of studied proteins.
:1000; Santa Cruz Biotech, Dallas, TX, USA), Akt (1:1000; Santa Cruz Biotech, Dallas, TX, USA), AS160 (1:1000; Santa Cruz Biotech, Dallas, TX, USA), and GLUT-4 (1:1000; Santa Cruz Biotech, Dallas, TX, USA). Subsequently, membranes were incubated with polyclonal anti-mouse (1:5000; Santa Cruz Biotech, Dallas, TX, USA) for Akt, anti-goat (1:5000; Santa Cruz Biotech, Dallas, TX, USA) for GLUT-4 and anti-rabbit (1:5000; Santa Cruz Biotech, Dallas, TX, USA) for IRβ and AS160 antibodies for 2 h at room temperature. After antibody stripping, membranes were blocked and incubated with phosphorylated IRβ (Tyr 1162/1163) (1:1000; Santa Cruz Biotech, Dallas, TX, USA), phosphorylated Akt (Thr 473) (1:1000; Cell Signaling, Danvers, MA, USA), phosphorylated AS160 (Thr 642) (1:1000; Gene Tex, CA, USA), and α-tubulin (1:1000; Cell Signaling, Danvers, MA, USA). The bound antibodies were visualized by an ECL system (Thermo Fisher Scientific Inc., Rockford, IL, USA) and quantified by a ChemiDoc MP imaging system (Bio-Rad, Hercules, CA, USA). The measurements were normalized by either β-actin or the phosphorylated isoform of studied proteins.
2.6. Statistical analysis
Results are presented as mean ± SEM. Statistical analysis was performed using the SPSS 24.0 software (SPSS, Chicago, IL, USA). The normal distribution of data was assessed by the Shapiro–Wilks test. Normally distributed parameters were analyzed by one-way ANOVA, followed by the Newman–Keuls post hoc test. Significance was assessed at the p < 0.05 level.3. Results
3.1. Body weight, energy intake, adipose tissue weights (total, visceral, subcutaneous) and adipose tissue index
At the end of the experimental period, the group fed on the obesogenic diet showed a significantly higher body weight gain compared to the animals receiving the standard laboratory diet. Moreover, as shown in Table 2, the energy intake of animals in the HFHF group was also higher in comparison with the C group. The increase in body weight of the HFHF rats was accompanied by a higher proportion of white adipose tissue on both the subcutaneous and visceral depots in comparison to the C group. In fact, the proportion of total white adipose tissue in relation to the final body weight of the animals, namely, the adipose index, was significantly higher in the HFHF group as compared to the C group.| C | HFHF | PRO | PARA | p value | |
|---|---|---|---|---|---|
| Values are presented as mean ± SEM. Differences among groups were determined using a one-way ANOVA, followed by the Newman Keuls post hoc test. Values not sharing a common letter are significantly different (p < 0.05). AT: adipose tissue. | |||||
| Body weight gain (g) | 140.6 ± 9.0b | 194.4 ± 7.9a | 152.5 ± 13.3b | 158.6 ± 10.9b | <0.05 |
| Energy intake (kcal d−1) | 79.1 ± 2.1b | 96.3 ± 1.6a | 90.7 ± 2.5a | 91.1 ± 2.1a | <0.01 |
| Total AT weight (g) | 34.1 ± 3.2b | 55.1 ± 4.8a | 37.0 ± 3.6b | 36.2 ± 2.7b | <0.01 |
| Visceral AT weight (g) | 22.3 ± 2.2b | 37.3 ± 3.4a | 24.3 ± 2.6b | 24.9 ± 2.2b | <0.05 |
| Subcutaneous AT weight (g) | 11.9 ± 1.8b | 17.8 ± 1.6a | 12.7 ± 1.1b | 11.3 ± 0.7b | <0.05 |
| Adipose Index (%) | 8.4 ± 0.9b | 11.7 ± 0.9a | 8.4 ± 0.6b | 8.2 ± 0.5b | <0.05 |
Regarding the effects of the treatments on somatometric variables, significantly lower body weight gain was found in the PRO and PARA groups compared to the HFHF group (p = 0.017 PRO vs. HFHF, p = 0.008 PARA vs. HFHF). Noteworthily, this effect occurred without differences among groups in terms of diet intake. Interestingly, the prevention of body weight gain was reflected in the content of total adipose tissue (visceral and subcutaneous depots), which was also lower in the treated groups compared to the HFHF group (Table 2).
3.2. Serum determinations
Markers of glycaemic control were assessed in serum samples at the end of the experimental period (Table 3). A marked tendency towards higher basal glucose levels along with a significantly higher insulin concentration was observed in the animals fed with the high-fat high-fructose diet compared to those fed with the standard diet (p = 0.073; p < 0.05 HFHF vs. C). These results suggest that insulin resistance was induced, as shown by the higher values for the HOMA-IR index (Table 3). The high-fat high-fructose feeding also resulted in higher serum triglyceride levels along with increased TyG index values compared to the C group (p < 0.05 and p < 0.01, respectively). As far as serum adipokine levels are concerned, a significant reduction in the levels of circulating adiponectin was noted in the animals receiving the obesogenic diet alone (p < 0.05 HFHF vs. C). Although no differences were found in the leptin levels between the C and the HFHF group, the adiponectin/leptin ratio (commonly used as an inverse index of insulin resistance) showed a significant reduction in the HFHF group compared to the C group (p < 0.01 HFHF vs. C). The level of the hormone fibroblast growth factor-21 (FGF-21) in the HFHF group was significantly lower than that observed in the C group (p < 0.01 HFHF vs. C).| C | HFHF | PRO | PARA | p value | |
|---|---|---|---|---|---|
| Values are presented as mean ± SEM. Differences among groups were determined using a one-way ANOVA, followed by the Newman Keuls post hoc test. Values not sharing a common letter are significantly different (p < 0.05). FGF-21: fibroblast growth factor 21. | |||||
| Basal glucose (mg dL−1) | 88.0 ± 4.7ab | 98.5 ± 4.9a | 83.5 ± 4.0b | 83.8 ± 4.8b | <0.05 |
| Insulin (mU L−1) | 18.1 ± 0.4c | 23.1 ± 1.7a | 19.6 ± 0.4b | 18.4 ± 0.2c | <0.05 |
| HOMA-IR | 4.0 ± 0.3b | 5.4 ± 0.5a | 4.2 ± 0.1b | 3.9 ± 0.2b | <0.05 |
| Triglycerides (mg dL−1) | 49.2 ± 4.9c | 81.5 ± 6.9a | 63.7 ± 1.9b | 69.4 ± 3.0ab | <0.05 |
| TyG index | 7.6 ± 0.09c | 8.3 ± 0.11a | 7.9 ± 0.04b | 8.0 ± 0.07b | <0.05 |
| Adiponectin (ng mL−1) | 41.8 ± 6.9a | 18.7 ± 4.7b | 35.1 ± 5.9a | 29.4 ± 4.5ab | <0.05 |
| Leptin (ng mL−1) | 1877 ± 530ab | 2479 ± 597a | 908 ± 133b | 1689 ± 333a | <0.05 |
| Adiponectin/leptin | 0.030 ± 0.004b | 0.009 ± 0.002c | 0.045 ± 0.007a | 0.053 ± 0.030abc | <0.01 |
| FGF-21 (pg mL−1) | 681 ± 289a | 257 ± 92c | 505 ± 230ab | 373 ± 191bc | <0.05 |
Concerning the effects of the treatments, PRO and PARA animals showed lower levels of basal glucose compared to the rats receiving the obesogenic diet alone (p = 0.017 PRO vs. HFHF; p = 0.024 PARA vs. HFHF). Regarding serum insulin levels, both treated groups showed a significant decrease in comparison to the HFHF group (p = 0.039 PRO vs. HFHF, p = 0.012 PARA vs. HFHF). Moreover, the animals receiving the parabiotic showed significantly lower levels of the same hormone compared to the animals fed with the obesogenic diet and treated with probiotic (p < 0.01) (Table 3). In addition, the elevation in the serum triglyceride levels induced by the high-fat high-fructose diet was effectively prevented by the probiotic administration (p < 0.05 PRO vs. HFHF). In the case of the animals fed the same obesogenic diet and receiving the parabiotic, a non-significant trend towards lower triglyceride levels was observed (p = 0.071). Regarding insulin resistance-related indexes, HOMA-IR was significantly lower in both treated groups compared to the rats under the obesogenic diet alone (p < 0.05 PRO vs. PARA; p < 0.01 PRO vs. PARA). In the case of the TyG index, both probiotic and parabiotic administration significantly prevented the effects induced by the obesogenic diet (Table 3).
Moreover, probiotic administration also efficiently prevented the alterations in both adiponectin and leptin serum levels compared to the non-treated group (p < 0.05 PRO vs. HFHF), and thus resulted in a significantly higher adiponectin/leptin ratio (p < 0.01). In the case of the PARA group, a tendency towards higher adiponectin levels compared to the HFHF group was observed (p = 0.061 PARA vs. HFHF), whilst no significant changes in the measured leptin were found. Therefore, the change induced by the parabiotic administration in the adiponectin/leptin ratio did not reach statistical significance (p = 0.093 PARA vs. HFHF). The lowering effect that the high-fat high-fructose diet provoked in FGF-21 serum levels was significantly prevented by the probiotic administration, whereas the parabiotic administration only tended to lower values (p = 0.055 PARA vs. HFHF).
3.3. Skeletal muscle triacylglycerol content
When skeletal muscle triacylglycerol content was evaluated, no significant changes were found in the animals receiving the obesogenic diet compared to the mice fed with the standard laboratory diet (Fig. 1). Interestingly, a significant reduction was observed in the group supplemented with the parabiotic compared to the HFHF group (p < 0.05).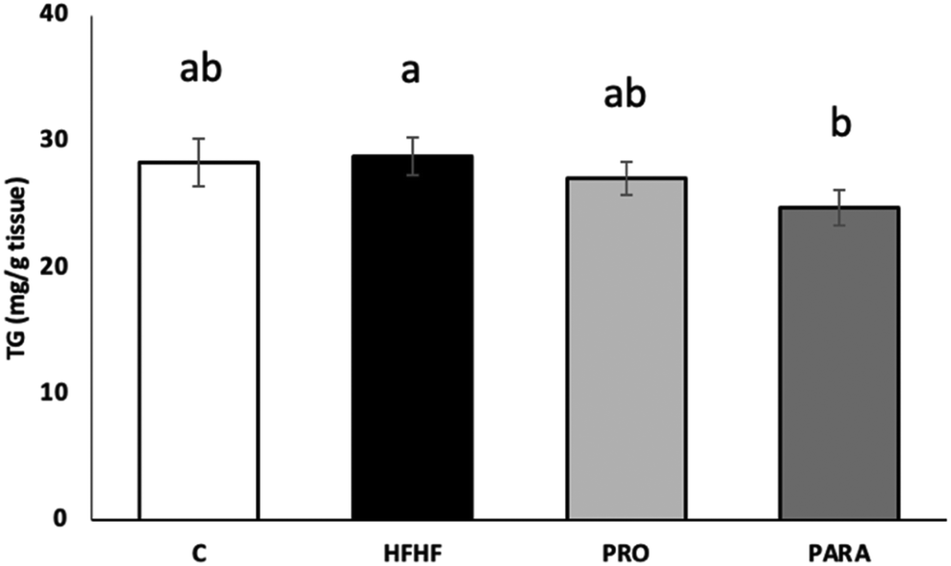 | ||
| Fig. 1 Skeletal muscle triacylglycerol content of rats fed with a control diet (C), a high-fat high-fructose diet (HFHF), and a high-fat high-fructose diet supplemented with viable or heat-inactivated 109 CFU per day of Lactobacillus rhamnosus GG (PRO and PARA, respectively). Values are presented as mean ± SEM. Differences among the groups were determined using one-way ANOVA, followed by the Newman–Keuls post hoc test. Bars not sharing common letters are significantly different (p < 0.05). | ||
3.4. Enzymatic activities
To evaluate the effects of both the high-fat high-fructose diet and the probiotic and parabiotic administration on fatty acid oxidation, the activities of two key enzymes, namely, citrate synthase (CS) and carnitine palmitoyl-transferase 1b (CPT-1b), were assessed. No significant changes were observed between the C and the HFHF groups in either of the enzyme activities (Fig. 2). | ||
| Fig. 2 CS (A) and CPT-1b (B) activities in skeletal muscle of rats fed with a control diet (C), a high-fat high-fructose diet (HFHF), and a high-fat high-fructose diet supplemented with viable or heat-inactivated 109 CFU per day of Lactobacillus rhamnosus GG (PRO and PARA, respectively). Values are presented as mean ± SEM. Differences among the groups were determined using one-way ANOVA, followed by the Newman–Keuls post hoc test. Bars not sharing common letters are significantly different (p < 0.05). | ||
As for the effects of the treatments, the probiotic group showed a non-significant yet sharp increase in the citrate synthase activity compared to the C group (p = 0.052) (Fig. 2A). In the case of CPT-1b, a trend towards a higher enzyme activity was observed in the PRO group compared to the HFHF group (p = 0.076). In the case of the PARA group, this increase reached statistical significance (p < 0.05 PARA vs. HFHF) (Fig. 2B).
3.5. Western blot analysis in skeletal muscle
To better elucidate the mechanisms underlying the effects induced by the studied treatments in glycaemic control, some of the key proteins of the insulin-signalling cascade were analyzed. Thus, the protein expression or activation (phosphorylation) of IRβ, Akt, AS160, and GLUT-4 in the skeletal muscle were assessed. Animals fed with the high-fat high-fructose diet showed a marked tendency towards a lower phosphorylation of IRβ compared to the C group (−14%; p = 0.055). This trend was also observed in Akt activation (−50%; p = 0.065). Moreover, a significant decrease in AS160 phosphorylation (p < 0.05 HFHF vs. C) was detected. Finally, although there were no significant changes in GLUT-4, the HFHF group showed a reduction of 29.3% in its protein expression levels compared to the C group (Fig. 3). | ||
| Fig. 3 Activation rate of IRβ, AKT, and AS160 and protein expression of GLUT-4 in skeletal muscle of rats fed with a control diet (C), a high-fat high-fructose diet (HFHF), and a high-fat high-fructose diet supplemented with viable or heat-inactivated 109 CFU per day of Lactobacillus rhamnosus GG (PRO and PARA, respectively). Values are presented as mean ± SEM. Differences among the groups were determined using one-way ANOVA, followed by the Newman–Keuls post hoc test. Bars not sharing common letters are significantly different (p < 0.05). # = p < 0.1 (vs. HFHF). | ||
As for the effects of the treatments, a non-statistical trend towards higher IRβ activation was observed in the PRO group (p = 0.05 PRO vs. HFHF), albeit no changes were found in the PARA group. Both treatments tended to prevent the decrease caused by the obesogenic diet in Akt phosphorylation (p = 0.088 PRO vs. HFHF; p = 0.095 PARA vs. HFHF). The lowering effect of the high-fat high-fructose diet on AS160 phosphorylation was effectively prevented by probiotic administration (p < 0.03). In the case of the PARA group, a non-significant increase in this parameter was observed compared to the HFHF group (+54%). Regarding GLUT-4 expression, probiotic treatment did not yield significant changes. However, the administration of the heat-killed bacteria significantly increased the GLUT-4 protein expression compared to the HFHF group (p < 0.05). This group also displayed a tendency towards higher expression levels compared to the animals receiving the standard laboratory diet and those treated with the viable bacteria (p = 0.057 PAR vs. C; p = 0.052 PAR vs. PRO).
4. Discussion
Obesity is not only a major health problem worldwide but also a driver of relevant metabolic impairments, such as insulin resistance, which represents a pivotal role in the onset of hallmark imbalances of metabolic syndrome.31 Unhealthy dietary patterns, characterized by a high intake of saturated fats and added sugars, have been identified as one of the main triggers of these health outcomes. Fructose has been shown to exert especially detrimental metabolic effects, contributing to insulin resistance development.32 In this study, rats receiving a high-fat high-fructose diet, which resembled the human-consumed westernized dietary pattern, showed a higher energy intake and consequently a higher body weight gain along with a greater weight of subcutaneous and visceral white adipose tissue depots, compared to the animals fed with the standard diet. The increased caloric intake observed in the HFHF group may well derive from the chronic fructose intake, which has been related to impaired hunger signals.33 Noteworthily, the administration of viable or heat-inactivated Lactobacillus rhamnosus GG resulted in a significantly lower body weight gain and adipose tissue weight gain, even though the animals receiving these treatments consumed the same obesogenic diet as the animals in the HFHF group. Indeed, these effects were found without differences in caloric intake among the groups receiving the diet rich in fat and fructose. These results are in accordance with those reported in other studies addressing the anti-obesity effects of this same probiotic.34,35 Also, a recent study has described similar results regarding the effects of probiotics in body adiposity despite having used different bacterial strains (Lactobacillus plantarum strains ATG-K2 and ATG-K6).36 Although the available literature regarding the effects of heat-inactivated probiotics is scarce so far, a recent study reported body weight gain prevention in a murine model of diet-induced obesity, highlighting the potential value of parabiotic administration for obesity prevention.37It is known that excessive fat accumulation leads to adipose tissue dysfunction, which in turn represents one of the first stages of insulin resistance development.38 Besides obesity, the high-fat high-fructose diet also led to alterations in serum markers of glucose homeostasis. In this regard, although the basal serum glucose levels observed in the animals under the high-fat high-fructose feeding were not different from those found in the control rats, their serum insulin level was significantly higher. These results are in agreement with the available literature since chronic fructose consumption is known to induce hyperinsulinemia in different animal models, leading to insulin resistance development.5,39 Indeed, the higher serum insulin level found in the HFHF group was also accompanied by higher values in indexes of insulin resistance, such as HOMA-IR and TyG. In this line, higher values of the above-mentioned indexes have also been reported in a recent clinical trial aimed at studying the effect of different fructose amount consumption on glycaemic parameters.40 Interestingly, the administration of Lactobacillus rhamnosus GG (PRO group) and inactivated bacteria (PARA group) resulted in lower blood glucose and insulin levels compared to the non-treated animals, suggesting a reduction in insulin resistance, which was confirmed by the reduction in HOMA-IR. In this context, preclinical and clinical studies have reported beneficial effects exerted by probiotics in term of glucose metabolism amelioration.41,42 Indeed, other authors have also reported changes in serum glucose levels without any difference in the insulin concentration under a different strain of probiotic Lactobacillus administration.43 As for the studies conducted to date addressing the effects of parabiotics in glycaemic control, reductions in blood glucose and insulin levels have also been described in a recent study carried out in Zucker fatty diabetic rats that were treated with a different parabiotic (heat-killed Streptococcus thermophilus) and for a longer experimental period (12 weeks).44 Thus, the results obtained in the current research along with those reported by other authors suggest that the viability of probiotic bacteria is not essential to preventing glycaemic impairments at least in rodent experimental models.
Adipose tissue dysfunction is a key signature of obesity and insulin resistance pathophysiology. In this regard, adiponectin and leptin are two well-known adipokines produced by the adipose tissue that have been related to insulin resistance.45 Indeed, direct relationships have been established between the size and/or shifts in adipose tissue weight and the circulating levels of these adipokines. In this line, the reduction in the adiponectin/leptin ratio has been considered a predictive marker of insulin resistance in obese adults.46 In this study, the high-fat high-fructose feeding induced a significant reduction in serum adiponectin levels, along with increased circulating levels of leptin compared to the control animals. These results are in good accordance with the current knowledge of these adipokines. Interestingly, the administration of viable Lactobacillus rhamnosus GG efficiently prevented the decrease in adiponectin levels produced by the obesogenic diet, an effect that has also been reported by other authors, although using a different experimental model.47 Moreover, a reduction in serum leptin levels was observed in this group, and thus, a significant increase in the adiponectin/leptin ratio compared to the non-treated animals fed with the obesogenic diet. Similar reductions in serum leptin levels were reported in a recent study using the same bacterial strain and a different experimental design, despite not having assessed further insulin resistance-related determinations.48 Although some studies have reported that heat-killed bacteria administration may restore adiponectin and leptin levels in animals displaying glycaemic control impairments,49 no such effects were observed in this study. This apparent discrepancy may well be due to the differences in the selected experimental conditions (bacterial strain and treatment length) and the animal model. Thus, a greater impairment of glucose homeostasis (such as type 2 diabetes instead of insulin resistance) might be required for parabiotics to restore serum adiponectin and leptin levels.
In order to better elucidate the mechanisms of action underlying the observed effects of the experimental diet and the treatments administered to prevent diet-induced insulin resistance, several metabolic pathway markers were studied in skeletal muscle, which accounts for up to 75% of insulin-mediated glucose transport in the body. Ectopic fat accumulation in skeletal muscle is highly related to insulin resistance, mainly by impairing the functionality of the insulin signalling cascade. In the present study, no differences were observed in gastrocnemius muscle triglyceride content between the animals fed with the standard laboratory diet and the ones receiving the obesogenic diet alone. Our results are in line with those published by other authors,50 who observed no changes in the intramyocellular lipid content of animals under a high-fructose diet. More specifically, Wang et al.51 reported that lipid and glucose metabolism alterations in the muscle occur from the eighth week onwards, suggesting that a longer experimental period may be necessary to develop these disturbances. However, parabiotic administration showed a decreased triglyceride content in the skeletal muscle of rats. Regarding mitochondrial enzyme activities, although other authors have reported no changes in the mRNA expression of Cpt1-b under heat-killed probiotic administration,52 we observed higher activity of this enzyme in the mitochondria of animals treated with the parabiotic. These data suggest that the lower muscle triglyceride content found in the parabiotic group may be linked to a higher CPT1-b enzyme activity in the skeletal muscle.
In addition, the activation rate or protein expression of several key proteins in the insulin-signalling cascade was also studied. Although no significant changes in GLUT-4 were observed in the HFHF group, the administration of the high-fat high-fructose diet caused a sharp decrease in IRβ and Akt phosphorylation, and a significant reduction in AS160 phosphorylation compared to the standard laboratory diet. These results suggest that feeding the rats with a diet rich in fat and fructose resulted in an impaired insulin signalling transduction (lower IRβ and Akt phosphorylation), paralleled with a decrease in GLUT-4 translocation to the plasma membrane (lower AS160 phosphorylation), which could explain the tendency towards the higher blood glucose levels observed in these animals despite featuring higher blood insulin concentrations. This model resembles an insulin-resistance status mainly with changes in AS160 activation rates. Other authors have also reported that either high-fructose or high-fat feeding significantly reduces the Akt and AS160 phosphorylation, respectively, in skeletal muscle.53,54 In the case of the animals fed with the obesogenic diet and treated with the probiotic, a trend towards a higher activation of IRβ and Akt along with a significant increase in AS160 phosphorylation was observed. Our results are in accordance with those published in other studies reporting a higher Akt protein expression in the skeletal muscle of diabetic mice under probiotic treatment with the microorganism used in the present study or another Lactobacillus strain (L. paracasei HII01).55,56 Regarding GLUT-4 expression, our results are not in line with studies reporting increased Glut-4 gene expression in high-fat diet fed mice and treated with a mixture of probiotics (L. plantarum MTCC5690 and L. fermentum MTCC5689) or with the same strain used in the present study.34,57 Nevertheless, it must be noted that changes in gene expression may not always translate into increased/decreased protein expression and that the experimental conditions used in those studies differ from the current study. In general, these results suggest that the translocation of the glucose transporter to the plasma membrane was activated to some extent, which in turn could result in a greater glucose uptake by the skeletal muscle. Finally, the effects induced by the administration of heat-killed bacteria in the expression/activation rate of insulin signalling proteins resembled those observed in the animals receiving the probiotic, mostly inducing changes in Akt and AS160 phosphorylation and GLUT-4 protein expression. Both Akt and AS160 showed trends towards higher phosphorylation levels compared to the animals receiving the obesogenic diet alone. Similar results have been found in other studies reporting increased Akt phosphorylation in the skeletal muscle of rats receiving heat-killed bacteria but using other species (Bifidobacterium breve B-3).58 With regard to GLUT-4, parabiotic administration was proven to be effective in increasing the protein expression level, which is in accordance with that reported by other studies wherein an increase in Glut-4 gene expression in adipose tissue under heat-killed bacteria treatment was observed.49 Nonetheless, to the best of our knowledge, no results have been published to date concerning the effects of parabiotic administration on proteins of insulin-signalling cascade in skeletal muscle. The serum glucose levels of animals receiving the probiotic or the parabiotic were similar. However, the latter showed lower levels of serum insulin. These results together with the higher expression of GLUT-4 in the skeletal muscle of rats in the PARA group may suggest that the administration of heat-inactivated bacteria is more effective in preventing the loss of insulin sensitivity under high-fat high-fructose feeding.
The results obtained in the current study demonstrate that under these experimental conditions the administration of heat-inactivated Lactobacillus rhamnosus GG is as effective as its probiotic of origin (if not better), ameliorating the impairments induced by a diet rich in saturated fat and fructose in glycaemic control. In this regard, one of the main limitations of the study is the impossibility to determine the bacterial component of the parabiotic responsible for triggering these metabolic benefits. In this line, different authors have hypothesized about the advantages of administering inactivated bacteria (featuring disrupted bacterial membranes) compared to viable probiotics since some of the bacterial products exerting benefits are located within the bacteria.59 Therefore, when administering the parabiotic these bioactive bacterial products may well be more accessible than in the probiotic (whose bacterial membranes would be intact). Moreover, despite literature regarding the potential benefits of parabiotic administration on glycaemic control disturbances still being scarce, different bacterial components (including bacterial cell wall-derived muramyl dipeptide or lipoteichoic acid, among others) are known to ameliorate insulin resistance.60,61 Indeed, potential additive or synergic effects among different bacterial bioactive components cannot be discarded either.
5. Conclusion
Altogether, the results obtained in the current study suggest that the administration of heat-inactivated Lactobacillus rhamnosus GG is more effective than its probiotic of origin, preventing insulin sensitive reduction induced under high-fat high-fructose feeding. This effect may well be mediated by both higher CPT1-b activity and enhanced GLUT-4 expression in the skeletal muscle.Author contributions
I. Milton-Laskibar, J. A. Martinez and M. P. Portillo: conceptualization; L. Arellano-García, I. Milton-Laskibar and M. T. Macarulla: investigation; L. Arellano-García and I. Milton-Laskibar: formal analysis; L. Arellano-García, I. Milton-Laskibar, M. T. Macarulla and M. P. Portillo wrote the original paper; and all the authors reviewed and edited the original paper.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This study was supported by Instituto de Salud Carlos III (CIBERobn) under grant CB12/03/30007 and The Basque Government under grant IT1482-22. Laura Isabel Arellano-García is a recipient of a doctoral fellowship from the Gobierno Vasco.References
- Y. C. Chooi, C. Ding and F. Magkos, The epidemiology of obesity, Metabolism, 2019, 92, 6–10 CrossRef CAS PubMed.
- A. G. Ampofo and E. B. Boateng, Beyond 2020: Modelling obesity and diabetes prevalence, Diabetes Res. Clin. Pract., 2020, 167, e108362 CrossRef PubMed.
- A. D. Colosia, R. Palencia and S. Khan, Prevalence of hypertension and obesity in patients with type 2 diabetes mellitus in observational studies: a systematic literature review, Diabetes, Metab. Syndr. Obes., 2013, 6, 327–338 CrossRef PubMed.
- D. Statovci, M. Aguilera, J. MacSharry and S. Melgar, The Impact of Western Diet and Nutrients on the Microbiota and Immune Response at Mucosal Interfaces, Front. Immunol., 2017, 8, 838 CrossRef.
- S. A. Hannou, D. E. Haslam, N. M. McKeown and M. A. Herman, Fructose metabolism and metabolic disease, J. Clin. Invest., 2018, 128(2), 545–555 CrossRef PubMed.
- J. Geng, Q. Ni, W. Sun, L. Li and X. Feng, The links between gut microbiota and obesity and obesity related diseases, Biomed. Pharmacother., 2022, 147, 112678 CrossRef CAS.
- E. Rinninella, P. Raoul, M. Cintoni, F. Franceschi, G. A. D. Miggiano and A. Gasbarrini, et al., What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases, Microorganisms, 2019, 7(1), 14 CrossRef CAS.
- Y. A. Kim, J. B. Keogh and P. M. Clifton, Probiotics, prebiotics, synbiotics and insulin sensitivity, Nutr. Res. Rev., 2018, 31(1), 35–51 CrossRef CAS.
- K. B. Martinez, V. Leone and E. B. Chang, Western diets, gut dysbiosis, and metabolic diseases: Are they linked?, Gut Microbes, 2017, 8(2), 130–142 CrossRef PubMed.
- J. E. Donnelly, S. N. Blair, J. M. Jakicic, M. M. Manore, J. W. Rankin and B. K. Smith, American College of Sports Medicine Position Stand. Appropriate physical activity intervention strategies for weight loss and prevention of weight regain for adults, Med. Sci. Sports Exercise, 2009, 41(2), 459–471 CrossRef PubMed.
- D. Xavier-Santos, R. Bedani, E. D. Lima and S. M. I. Saad, Impact of probiotics and prebiotics targeting metabolic syndrome, J. Funct. Foods, 2020, 64, 103666 CrossRef CAS.
- Y. Vandenplas, G. Huys and G. Daube, Probiotics: an update, J. Pediatr., 2015, 91(1), 6–21 CrossRef.
- P. S. Hsieh, H. H. Ho, S. P. Tsao, S. H. Hsieh, W. Y. Lin and J. F. Chen, et al., Multi-strain probiotic supplement attenuates streptozotocin-induced type-2 diabetes by reducing inflammation and β-cell death in rats, PLoS One, 2021, 16(6), e0251646 CrossRef CAS.
- L. B. Tonucci, K. M. Olbrich dos Santos, L. Licursi de Oliveira, S. M. Rocha Ribeiro and H. S. Duarte Martino, Clinical application of probiotics in type 2 diabetes mellitus: A randomized, double-blind, placebo-controlled study, Clin. Nutr., 2017, 36(1), 85–92 CrossRef CAS PubMed.
- F. Perraudeau, P. McMurdie, J. Bullard, A. Cheng, C. Cutcliffe and A. Deo, et al., Improvements to postprandial glucose control in subjects with type 2 diabetes: a multicenter, double blind, randomized placebo-controlled trial of a novel probiotic formulation, BMJ Open Diabetes Res. Care, 2020, 8(1), e001319 CrossRef PubMed.
- S. Doron and D. R. Snydman, Risk and safety of probiotics, Clin. Infect. Dis., 2015, 60(Suppl 2), S129–S134 CrossRef PubMed.
- N. Li, W. M. Russell, M. Douglas-Escobar, N. Hauser, M. Lopez and J. Neu, Live and heat-killed Lactobacillus rhamnosus, GG: effects on proinflammatory and anti-inflammatory cytokines/chemokines in gastrostomy-fed infant rats, Pediatr. Res., 2009, 66(2), 203–207 CrossRef CAS PubMed.
- B. H. Nataraj, S. A. Ali, P. V. Behare and H. Yadav, Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods, Microb. Cell Fact., 2020, 19(1), 1–22 CrossRef.
- J. Gao, Y. Li, Y. Wan, T. Hu, L. Liu, S. Yang, Z. Gong, Q. Zeng, Y. Wei, W. Yang and Z. Zeng, et al., A Novel Postbiotic From Lactobacillus rhamnosus GG With a Beneficial Effect on Intestinal Barrier Function, Front. Microbiol., 2019, 10, 477 CrossRef.
- S. Salva, I. Tiscornia, F. Gutiérrez, S. Alvarez and M. Bollati-Fogolín, Lactobacillus rhamnosus postbiotic-induced immunomodulation as safer alternative to the use of live bacteria, Cytokine, 2021, 146, 155631 CrossRef CAS.
- T. Matsuguchi, A. Takagi, T. Matsuzaki, M. Nagaoka, K. Ishikawa, T. Yokokura and Y. Yoshikai, Lipoteichoic acids from Lactobacillus strains elicit strong tumor necrosis factor alpha-inducing activities in macrophages through Toll-like receptor 2, Clin. Diagn. Lab. Immunol., 2003, 10(2), 259–266 CAS.
- L. Zhang, N. Li, R. Caicedo and J. Neu, Alive and dead Lactobacillus rhamnosus GG decrease tumor necrosis factor-alpha-induced interleukin-8 production in Caco-2 cells, J. Nutr., 2005, 135(7), 1752–1756 CrossRef CAS.
- D. R. Matthews, J. P. Hosker, A. S. Rudenski, B. A. Naylor, D. F. Treacher and R. C. Turner, Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man, Diabetologia, 1985, 28(7), 412–419 CrossRef CAS PubMed.
- L. E. Simental-Mendía, M. Rodríguez-Morán and F. Guerrero-Romero, The product of fasting glucose and triglycerides as surrogate for identifying insulin resistance in apparently healthy subjects, Metab. Syndr. Relat. Disord., 2008, 6(4), 299–304 CrossRef.
- J. Folch, M. Lees and G. H. Sloane Stanley, A simple method for the isolation and purification of total lipides from animal tissues, J. Biol. Chem., 1957, 226(1), 497–509 CrossRef CAS.
- L. L. Bieber, T. Abraham and T. Helmrath, A rapid spectrophotometric assay for carnitine palmitoyltransferase, Anal. Biochem., 1972, 50(2), 509–518 CrossRef CAS PubMed.
- C. R. Bruce, A. J. Hoy and N. Turner, et al., Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet-induced insulin resistance, Diabetes, 2009, 58(3), 550–558 CrossRef CAS PubMed.
- M. M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem., 1976, 72(1–2), 248–254 CrossRef CAS.
- P. A. Srere, Citrate synthase: Citrate oxaloacetate-lyase (CoA-acetylating), Methods Enzymol., 1969, 13(C), 3–11 CAS.
- S. Larsen, J. Nielsen, C. N. Hansen, L. B. Nielsen, F. Wibrand, N. Stride, H. D. Schroder, R. Boushel, J. W. Helge and F. Dela, et al., Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects, J. Physiol., 2012, 590, 3349–3360 CrossRef CAS.
- D. Lann and D. LeRoith, Insulin Resistance as the Underlying Cause for the Metabolic Syndrome, Med. Clin. North Am., 2007, 91(6), 1063–1077 CrossRef CAS PubMed.
- S. Softic, K. L. Stanhope, J. Boucher, S. Divanovic, M. A. Lanaspa and R. J. Johnson, et al., Fructose and hepatic insulin resistance, Crit. Rev. Clin. Lab. Sci., 2020, 57(5), 308–322 CrossRef PubMed.
- K. Lowette, L. Roosen, J. Tack and P. Vanden Berghe, Effects of High-Fructose Diets on Central Appetite Signaling and Cognitive Function, Front. Nutr., 2015, 2, 5 Search PubMed.
- S. W. Kim, K. Y. Park, B. Kim, E. Kim and C. K. Hyun, Lactobacillus rhamnosus GG improves insulin sensitivity and reduces adiposity in high-fat diet-fed mice through enhancement of adiponectin production, Biochem. Biophys. Res. Commun., 2013, 431(2), 258–263 CrossRef CAS PubMed.
- M. Yakovlieva, T. Tacheva, S. Mihaylova, R. Tropcheva, K. Trifonova and A. Tolekova, et al., Influence of Lactobacillus brevis, 15 and Lactobacillus plantarum 13 on blood glucose and body weight in rats after high-fructose diet, Benefic. Microbes, 2015, 6(4), 505–512 CrossRef CAS.
- M. Park, E. J. Park, S. H. Kim and H. J. Lee, Lactobacillus plantarum ATG-K2 and ATG-K6 Ameliorates High-Fat with High-Fructose Induced Intestinal Inflammation, Int. J. Mol. Sci., 2021, 22(9), 4444 CrossRef CAS PubMed.
- X. Kang, H. Liang, Y. Luo, Z. Li, F. He and X. Han, et al., Anti-adipogenesis and metabolism-regulating effects of heat-inactivated Streptococcus thermophilus MN-ZLW-002, Lett. Appl. Microbiol., 2021, 72(6), 677–687 CrossRef CAS.
- B. Ahmed, R. Sultana and M. W. Greene, Adipose tissue and insulin resistance in obese, Biomed. Pharmacother., 2021, 137, 111315 CrossRef CAS.
- E. Benetti, R. Mastrocola, M. Rogazzo, F. Chiazza, M. Aragno and R. Fantozzi, et al., High Sugar Intake and Development of Skeletal Muscle Insulin Resistance and Inflammation in Mice: A Protective Role for PPAR-δ Agonism, Mediators Inflammation, 2013, 2013, e508502 Search PubMed.
- E. Keskin and H. Yoldas Ilktac, Fructose consumption correlates with triglyceride-glucose index and glycemic status in healthy adults, Clin. Nutr. ESPEN, 2022, 52, 184–189 CrossRef PubMed.
- M. C. Hsieh, W. H. Tsai, Y. P. Jheng, S. L. Su, S. Y. Wang and C. C. Lin, et al., The beneficial effects of Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2 diabetes mellitus: a randomized, double-blinded, placebo-controlled trial, Sci. Rep., 2018, 8(1), 16791 CrossRef PubMed.
- Y. W. Tao, Y. L. Gu, X. Q. Mao, L. Zhang and Y. F. Pei, Effects of probiotics on type II diabetes mellitus: a meta-analysis, J. Transl. Med., 2020, 18(1), 30 CrossRef CAS PubMed.
- M. Toral, M. Gómez-Guzmán, R. Jiménez, M. Romero, M. Sánchez and M. P. Utrilla, et al., The probiotic Lactobacillus coryniformis CECT5711 reduces the vascular pro-oxidant and pro-inflammatory status in obese mice, Clin. Sci., 2014, 127(1), 33–45 CrossRef.
- X. Gao, F. Wang, P. Zhao, R. Zhang and Q. Zeng, Effect of heat-killed Streptococcus thermophilus on type 2 diabetes rats, PeerJ, 2019, e7117 CrossRef.
- A. Yadav, M. A. Kataria, V. Saini and A. Yadav, Role of leptin and adiponectin in insulin resistance, Clin. Chim. Acta, 2013, 417, 80–84 CrossRef CAS PubMed.
- I. Castela, C. Rodrigues, S. Ismael, I. Barreiros-Mota, J. Morais and J. R. Araújo, et al., Intermittent energy restriction ameliorates adipose tissue-associated inflammation in adults with obesity: A randomised controlled trial, Clin. Nutr., 2022, 41(8), 1660–1666 CrossRef CAS PubMed.
- S. W. Kim, K. Y. Park, B. Kim, E. Kim and C. K. Hyun, Lactobacillus rhamnosus GG improves insulin sensitivity and reduces adiposity in high-fat diet-fed mice through enhancement of adiponectin production, Biochem. Biophys. Res. Commun., 2013, 431(2), 258–263 CrossRef CAS PubMed.
- Y. C. Cheng and J. R. Liu, Effect of Lactobacillus rhamnosus, GG on Energy Metabolism, Leptin Resistance, and Gut Microbiota in Mice with Diet-Induced Obesity, Nutrients, 2020, 12, 2557 CrossRef CAS.
- Y. H. Nam, S. Yoon, J. Baek, J. H. Kim, M. Park and K. W. Hwang, et al., Heat-Killed Lactiplantibacillus plantarum LRCC5314 Mitigates the Effects of Stress-Related Type 2 Diabetes in Mice via Gut Microbiome Modulation, J. Microbiol. Biotechnol., 2022, 32(3), 324–332 CrossRef CAS PubMed.
- P. M. Nunes, A. J. Wright, A. Veltien, J. J. A. Van Asten, C. J. Tack and J. G. Jones, et al., Dietary lipids do not contribute to the higher hepatic triglyceride levels of fructose- compared to glucose-fed mice, FASEB J., 2014, 28(5), 1988–1997 CrossRef CAS PubMed.
- Y. Wang, G. Fouret, B. Bonafos, A. Blachnio-Zabielska, T. Leroy and D. Crouzier, et al., Long-term follow-up of muscle lipid accumulation, mitochondrial activity and oxidative stress and their relationship with impaired glucose homeostasis in high fat high fructose diet-fed rats, J. Nutr. Biochem., 2019, 64, 182–197 CrossRef CAS PubMed.
- A. Caimari, J. M. del Bas, N. Boqué, A. Crescenti, F. Puiggròs and E. Chenoll, et al., Heat-killed Bifidobacterium animalis subsp. Lactis CECT 8145 increases lean mass and ameliorates metabolic syndrome in cafeteria-fed obese rats, J. Funct. Foods, 2017, 38, 251–263 CrossRef CAS.
- P. Li, T. Koike, B. Qin, M. Kubota, Y. Kawata and Y. J. Jia, et al., A high-fructose diet impairs Akt and PKCζ phosphorylation and GLUT4 translocation in rat skeletal muscle, Horm. Metab. Res., 2008, 40(8), 528–532 CrossRef CAS PubMed.
- P. M. Badin, I. K. Vila, K. Louche, A. Mairal, M. A. Marques and V. Bourlier, et al., High-Fat Diet-Mediated Lipotoxicity and Insulin Resistance Is Related to Impaired Lipase Expression in Mouse Skeletal Muscle, Endocrinology, 2013, 154(4), 1444–1453 CrossRef CAS PubMed.
- K. Y. Park, B. Kim and C. K. Hyun, Lactobacillus rhamnosus GG improves glucose tolerance through alleviating ER stress and suppressing macrophage activation in db/db mice, J. Clin. Biochem. Nutr., 2015, 56(3), 240–246 CrossRef CAS PubMed.
- P. Toejing, N. Khat-Udomkiri, J. Intakhad, S. Sirilun, C. Chaiyasut and N. Lailerd, Putative Mechanisms Responsible for the Antihyperglycemic Action of Lactobacillus paracasei HII01 in Experimental Type 2 Diabetic Rats, Nutrients, 2020, 12(10), 1–18 CrossRef PubMed.
- M. Balakumar, D. Prabhu, C. Sathishkumar, P. Prabu, N. Rokana and R. Kumar, et al., Improvement in glucose tolerance and insulin sensitivity by probiotic strains of Indian gut origin in high-fat diet-fed C57BL/6J mice, Eur. J. Nutr., 2018, 57(1), 279–295 CrossRef CAS PubMed.
- K. Toda, Y. Yamauchi, A. Tanaka, T. Kuhara, T. Odamaki and S. Yoshimoto, et al., Heat-Killed Bifidobacterium breve B-3 Enhances Muscle Functions: Possible Involvement of Increases in Muscle Mass and Mitochondrial Biogenesis, Nutrients, 2020, 12(1), 219 CrossRef CAS PubMed.
- N. Piqué, M. Berlanga and D. Miñana-Galbis, Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview, Int. J. Mol. Sci., 2019, 20(10), 2534 CrossRef PubMed.
- J. F. Cavallari, M. D. Fullerton and B. M. Duggan, et al., Muramyl Dipeptide-Based Postbiotics Mitigate Obesity-Induced Insulin Resistance via IRF4, Cell Metab., 2017, 25(5), 1063–1074 CrossRef CAS.
- S. Wang, S. Ahmadi and R. Nagpal, et al., Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei, D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: from C. elegans to mice, GeroScience, 2020, 42(1), 333–352 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2023 |