
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceImpact of cooking methods of red-skinned onion on metabolic transformation of phenolic compounds and gut microbiota changes†
Alice
Cattivelli‡
 a,
Lorenzo
Nissen‡
*bcd,
Flavia
Casciano
b,
Davide
Tagliazucchi
a and
Andrea
Gianotti
*bcd
a,
Lorenzo
Nissen‡
*bcd,
Flavia
Casciano
b,
Davide
Tagliazucchi
a and
Andrea
Gianotti
*bcd
aDepartment of Life Sciences, University of Modena and Reggio Emilia, Via Amendola, 2 – Pad. Besta, 42100 Reggio Emilia, Italy
bDepartment of Agricultural and Food Sciences (DISTAL), Alma Mater Studiorum – University of Bologna, Piazza Goidanich 60, 47521 Cesena (FC), Italy. E-mail: lorenzo.nissen@unibo.it; andrea.gianotti@unibo.it
cInterdepartmental Centre of Agri-Food Industrial Research (CIRI), Alma Mater Studiorum – University of Bologna, Piazza G. Goidanich 60, 47521 Cesena (FC), Italy
dCentre for Applied Biomedical Research – CRBA, Alma Mater Studiorum – Università di Bologna, Policlinico di Sant'Orsola, Via Massarenti 9, 40138 Bologna, Italy
First published on 22nd March 2023
Abstract
Herein, we investigated the stability and bioaccessibility of phenolics in differently cooked red-skinned onion (RSO) and consequently their impact on the gut microbiota and metabolism of phenolics. In fact, the different processes used to cook vegetables can modify and re-arrange the molecular profiles of bioactive compounds, such as phenolics in phenolic-rich vegetables, such as RSO. Fried and grilled RSO were compared to raw RSO and a blank control and subjected to oro-gastro-intestinal digestion and subsequent colonic fermentation. For upper gut digestion, the INFOGEST protocol was used, and for lower gut fermentation, a short-term batch model, namely, MICODE (multi-unit in vitro colon gut model), was employed. During the process, phenolic compound profile (through high-resolution mass spectrometry) and colon microbiomics (qPCR of 14 core taxa) analyses were performed. According to the results, the degradation driven by the colon microbiota of RSO flavonols resulted in the accumulation of three main metabolites, i.e., 3-(3′-hydroxyphenyl)propanoic acid, 3-(3′-hydroxyphenyl)acetic acid and 3-(3′,4′-dihydroxyphenyl)acetic acid. Also, colonic fermentation of raw onions resulted in a substantial increase in beneficial taxa, which was larger compared to the heat-treated onions, particularly Lactobacillales and beneficial clostridia. Also, a higher level of inhibition of opportunistic bacteria was seen for the raw onion samples, namely, Clostridium perfringens group and Escherichia coli. Thus, our results showed that RSO, and especially the raw one, is an excellent dietary source of flavonols that are strongly metabolized by gut bacteria and can positively modulate the gut microbiota. Although additional in vivo studies are necessary, this work is one of the first to explore how RSO processed with different cooking methods can differently impact the phenolic metabolism and microbiota composition in the large intestine of humans, fine-tuning the antioxidant nature of foods.
1. Introduction
Onion (Allium cepa L.) is one of the most consumed vegetables worldwide and is also traditionally part of the Mediterranean diet. Besides its typical taste and flavour, onion is considered a healthy food because it contains several classes of phenolic compounds such as flavonols (mainly quercetin and its glycosylated derivatives), hydroxycinnamic acids, and in red-skinned onion, anthocyanins.1–3 Recently, onion consumption has been associated with a reduction in the onset of some chronic diseases such as cardiovascular diseases, type-2 diabetes, and gastro-intestinal tract cancers.4,5Generally, onion can be consumed in the diet daily, either raw or cooked through different methods such as grilling and frying, which are the two most usual cooking procedures applied to onion. Several studies evidenced that heat treatments during cooking may modify the phenolic profile and structures of vegetables foods resulting in a different and sometimes higher bioactivity and improved bioaccessibility after in vitro gastro-intestinal digestion.1,6,7 Additionally, these modified compounds undergo an ulterior transformation when they enter the colon, where they can feed the colon microbiota to foster beneficial bacteria and inhibit opportunistic microbes.8 Unfortunately, this improvement can also have the opposite impact on the colon microbiota, adversely affecting sensitive beneficial bacteria, such as bifidobacteria.9 Moreover, it has been shown that both food composition and thermal treatments alter the composition of the gut microbiota.10 In addition, previous studies highlighted that different cooking procedures may affect the profile and concentration of phenolic metabolites derived from the gut microbiota metabolic activity during in vitro fermentation of several vegetables.10–13
The two-way interaction between the gut microbiota and phenolic compounds is considered pivotal in determining the beneficial effect of phenolic compounds in human health.14 In this sense, the colon bacteria can metabolize phenolic compounds in low molecular weight phenolic metabolites, which are better absorbed and often more active than the parent compounds as well as phenolic compounds may modulate the gut microbiota population.15,16 In particular, phenolic compounds may increase the relative abundance of beneficial bacteria involved in the protection of the intestinal epithelium (such as Lactobacillus and Bifidobacterium species) or that displaying anti-inflammatory effects (such as Faecalibacterium prausnitzii and Akkermansia muciniphila).10,17–19 In turn, colonic bacteria transform the structure of parent phenolic compounds in metabolites with improved biological activities and more bioavailable to the host.10,15,20
In vitro gut models are considered suitable to study the complex relationships between the chemical constituents of food and the colon microbiota because they can clarify the effect of prebiotics such as phenolic compounds on the human gut microbiota through the analysis of the shift in the core microbial groups or selected species, as well as assessing the modification in the produced metabolites by individual bacterial species.21 In this study, we employed a gut model based on healthy adults, coupling the INFOGEST static protocol to simulate the oro-gastro-duodenal digestion22 to the colon model MICODE (Multi-Unit in vitro Colon Model) to simulate proximal colonic fermentation, previously used with success to study food matrices rich in phenolic compounds.8–10 Therefore, the aim of this study was to assess the bioaccessibility of phenolic compounds of both raw and cooked (grilled and fried) red-skinned onion as well as to elucidate the principal metabolic pathways and produced metabolites after in vitro colonic fermentation. Furthermore, the changes in fecal microbiota groups and selected bacterial taxa were also investigated.
2. Materials and methods
2.1. Red-skinned onion (RSO) thermal treatments and in vitro gastro-intestinal digestion
Red-skinned onion (RSO) was subjected to two distinct thermal treatments to mimic the most common home cooking methods to which onion is subjected, i.e., grilling and frying. The cooking procedures and the cooking parameters were fully described previously.1 Thereafter, raw (RRSO), grilled (GRSO) and fried (FRSO) red-skinned onion samples were subjected to in vitro gastro-intestinal digestion using the INFOGEST protocol22 as previously described for onion.1 Finally, an aliquot of sample was collected at the end of the digestion procedure, centrifuged (10 min, 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g, 4 °C) and the supernatant stored at −80 °C for further analysis, whereas the remaining material (denser portion) was used for the in vitro colonic fermentation trials as described in Section 2.2. The cooking treatments and digestions were carried out in triplicate. Afterwards, for colonic in vitro fermentation, the triplicates of a given digestate were pooled.
000g, 4 °C) and the supernatant stored at −80 °C for further analysis, whereas the remaining material (denser portion) was used for the in vitro colonic fermentation trials as described in Section 2.2. The cooking treatments and digestions were carried out in triplicate. Afterwards, for colonic in vitro fermentation, the triplicates of a given digestate were pooled.
2.2. Colonic fermentation
2.3. High-resolution mass spectrometry analysis of phenolic compounds and fecal metabolites
:70:2, v/v/v) solution. Then, the mixtures were incubated for 30 min at 37 °C, and after centrifugation (20 min, 6000g, 4 °C), the supernatants were collected and analysed by mass spectrometry for the identification and quantification of the phenolic compounds.
The phenolic compounds in the in vitro digested samples and faecal metabolites were extracted as previously described.12,13 Briefly, 500 μL of extraction solution (water/methanol/formic acid; 19.9:80:0.1, v/v/v) was mixed with 500 μL of digested or fermented samples. After vortexing for 1 min, the mixtures were centrifuged (10 min; 12000g; 4 °C). Then the supernatants were used for mass spectrometry identification and quantification of phenolic compounds and metabolites.
2.4. Microbiota analyses
| qPI = (Bifidobacteriaceae/Eubacteria) − (Enterobacteriaceae/Eubacteria) + (Lactobacillales/Eubacteria) − (Clostridium group I/Eubacteria). |
2.5. Data processing and statistical analysis
The shifts in the abundance of the bacterial groups were calculated as log2(F/C) using the raw data36 obtained from sextuplicates. Also, based on the raw data, the qPI was obtained after normalization with the mean centering method. Normality and homoscedasticity of the datasets were considered with Levene's and Shapiro–Wilk's test, respectively. To compare the “time points” and “samples” with each other, the two factors used to categorize the MANOVAs were “time” and “food matrix”, respectively. MANOVA for “time” and “food matrix” as categorical descriptors and followed by a Duncan's multiple range post-hoc test was applied using Statistica v 8.0 (Tibco, Palo Alto, CA, USA). The differences were considered significant with p < 0.05. To address specific correlations among bacteria and metabolites, two independent datasets were merged and computed by Spearman rank analysis and visualized with a two-way joining heatmap, including Pearson dendrograms with complete linkage for both variables.8–10 The dataset of correlations was performed with Statistica v. 8.0 (Tibco, USA) and the heatmap was produced with the Expression tool at https://www.heatmapper.ca37 (last accessed on 15/12/2022).
3. Results and discussion
3.1. Effect of cooking and in vitro digestion on red-skinned onion phenolic compounds bioaccessibility
A total of 54 phenolic compounds belonging to different classes were identified and quantified in the red-skinned onion (RSO) samples (Table 1). The most representative class of phenolic compounds in RSO were flavonols, and in particular quercetin and quercetin-derivatives. In the raw sample (RRSO), flavonols represented the 85.3% of total phenolic compounds. Among the flavonols, the quercetin-derivatives accounted for 74.2% of the total phenolic compounds. The individual compound found at the highest concentration in RRSO was quercetin that represented the 50.1% of the total phenolic compounds. Two glycosylated-derivatives of quercetin, namely, quercetin-4′-O-glucoside and quercetin-3-O-glucoside-4′-O-glucoside were also present in an appreciable amount in RRSO, representing the 12.3% and 10.9% of total phenolic compounds, respectively. The second most important class of phenolic compounds in RRSO was represented by anthocyanins, which accounted for 11.2% of total phenolic compounds. A similar phenolic profile in RRSO was previously reported.2,38| Compound | RRSO | FRSO | GRSO | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Before digestion | After digestion | BI (%) | Before digestion | After digestion | BI (%) | Before digestion | After digestion | BI (%) | |
| Different superscript letters (a–c) within the same row mean significant different (p < 0.05) values; n.d. = not detected compound; n.f. = newly formed compound; RRSO = raw red-skinned onion; GRSO = grilled red-skinned onion; and FRSO = fried red-skinned onion. | |||||||||
| Hydroxycinnamic acids | |||||||||
| Di-hydro-coumaric acid | n.d. | 0.30 ± 0.02a | n.f. | 0.17 ± 0.01b | 0.35 ± 0.03a | 205.9 | 0.13 ± 0.01c | 0.29 ± 0.02a | 223.1 |
| Caffeic acid | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.03 ± 0.00 | n.d. | 0 |
| Sinapic acid | 0.08 ± 0.00a | 0.03 ± 0.00b | 37.5 | 0.02 ± 0.00b | 0.14 ± 0.01c | 700.0 | 0.04 ± 0.00b | 0.03 ± 0.00b | 67.7 |
| Ferulic acid-4-O-hexoside | 0.18 ± 0.01a | 0.18 ± 0.01a | 100.0 | 0.35 ± 0.03b | 0.11 ± 0.01c | 31.4 | 0.19 ± 0.01a | 0.14 ± 0.01d | 75.0 |
| Sinapic acid-O-hexoside isomer 1 | 0.70 ± 0.03a | 0.40 ± 0.02b | 57.1 | 2.05 ± 0.15c | 1.86 ± 0.09d | 90.7 | 0.40 ± 0.02b | 0.35 ± 0.03b | 87.5 |
| Sinapic acid-O-hexoside isomer 2 | 1.59 ± 0.08a | 0.12 ± 0.01b | 7.5 | 1.42 ± 0.08a | 0.73 ± 0.03c | 51.4 | 0.53 ± 0.03d | 0.15 ± 0.01b | 28.3 |
| Sinapic acid-O-hexoside isomer 3 | 1.00 ± 0.10a | 0.15 ± 0.01b | 15.0 | 1.09 ± 0.09a | 0.54 ± 0.04c | 49.5 | 0.10 ± 0.00d | n.d. | 0 |
| Sinapic acid-O-hexoside isomer 4 | 0.45 ± 0.04a | 0.08 ± 0.00b | 17.8 | 0.49 ± 0.03a | 0.54 ± 0.04a | 110.2 | n.d. | n.d. | n.d. |
| Total hydroxycinnamic acids | 4.00 ± 0.14 a | 1.26 ± 0.03 b | 31.5 | 5.59 ± 0.20 c | 4.27 ± 0.12 a | 76.4 | 1.42 ± 0.04 d | 0.96 ± 0.04 | 67.6 |
| Hydroxybenzoic acids | |||||||||
| Hydroxybenzoic acid isomer 1 | 0.08 ± 0.01a | 0.04 ± 0.00b | 50.0 | 0.17 ± 0.01c | 0.12 ± 0.01d | 70.6 | 0.14 ± 0.01d | 0.12 ± 0.01d | 85.7 |
| Hydroxybenzoic acid isomer 2 | 0.04 ± 0.00a | 0.06 ± 0.00a | 150.0 | 0.05 ± 0.00a | 0.06 ± 0.00a | 120.0 | 0.04 ± 0.00a | 0.06 ± 0.00a | 150.0 |
| Protocatechuic acid | 0.18 ± 0.01a | 1.00 ± 0.09b | 555.6 | 0.22 ± 0.01c | 0.86 ± 0.05b | 390.9 | 0.24 ± 0.01c | 1.26 ± 0.07d | 525.0 |
| Di-hydroxybenzoic acid | 0.15 ± 0.01a | 0.04 ± 0.00b | 26.7 | 0.16 ± 0.01a | 0.05 ± 0.00b | 31.3 | 0.17 ± 0.01a | 0.03 ± 0.00b | 17.6 |
| Vanillic acid | 0.12 ± 0.01a | 0.01 ± 0.00b | 8.3 | 0.07 ± 0.00c | n.d. | 0 | 0.12 ± 0.01a | n.d. | 0 |
| Di-hydroxybenzoic acid-hexoside isomer 1 | 0.14 ± 0.01a | 3.51 ± 0.25b | 2507.1 | 0.26 ± 0.01c | 1.45 ± 0.09d | 557.7 | 0.51 ± 0.04e | 2.35 ± 0.12f | 460.8 |
| Di-hydroxybenzoic acid-hexoside isomer 2 | 0.03 ± 0.00a | 0.08 ± 0.00a | 266.7 | 0.05 ± 0.00a | 0.05 ± 0.00a | 100.0 | 0.07 ± 0.00a | 0.07 ± 0.00a | 100.0 |
| Di-hydroxybenzoic acid-hexoside isomer 3 | 0.03 ± 0.00a | n.d. | 0 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Total hydroxybenzoic acids | 0.77 ± 0.02 a | 4.74 ± 0.27 b | 615.6 | 0.98 ± 0.02 c | 2.59 ± 0.10 d | 264.3 | 1.29 ± 0.04 e | 3.89 ± 0.14 f | 308.7 |
| Flavonols | |||||||||
| Quercetin | 74.14 ± 3.61a | n.d. | 0 | 50.01 ± 2.57b | 23.98 ± 1.77c | 48.0 | 56.59 ± 3.10b | 0.38 ± 0.01 | 0.67 |
| Quercetin-3-O-hexoside | n.d. | n.d. | n.d. | 0.04 ± 0.00a | n.d. | 0 | 0.04 ± 0.00a | n.d. | 0 |
| Quercetin-3-O-glucoside | 0.96 ± 0.05a | 0.61 ± 0.03b | 63.5 | 0.79 ± 0.01c | 1.90 ± 0.09d | 240.5 | 1.87 ± 0.11d | 1.76 ± 0.07 | 94.1 |
| Quercetin-4′-O-glucoside | 18.28 ± 1.03a | 17.50 ± 1.11a | 95.7 | 45.11 ± 1.80b | 18.88 ± 1.21a | 41.9 | 50.60 ± 3.24b | 10.60 ± 0.81c | 20.9 |
| Quercetin-7-O-hexoside-4′-O-hexoside | 0.03 ± 0.00a | 0.17 ± 0.01b | 566.7 | 0.14 ± 0.01b | 0.17 ± 0.01b | 121.4 | 0.22 ± 0.01c | 0.07 ± 0.00a | 31.8 |
| Quercetin-3-O-glucoside-4′-O-glucoside | 16.21 ± 0.99a | 15.93 ± 0.99a | 98.3 | 31.12 ± 1.51b | 23.13 ± 1.56c | 74.3 | 46.65 ± 2.39d | 39.76 ± 2.09e | 85.2 |
| Quercetin-tri-O-hexoside | 0.05 ± 0.00a | 0.04 ± 0.00a | 80.0 | 0.12 ± 0.00b | 0.06 ± 0.00a | 50.0 | 0.25 ± 0.02c | 0.07 ± 0.00a | 28.0 |
| Quercetin-O-hexoside-O-malonyl-hexoside isomer 1 | 0.04 ± 0.00a | 0.04 ± 0.00a | 100.0 | 0.03 ± 0.00a | 0.03 ± 0.00a | 100.0 | 0.05 ± 0.00a | 0.05 ± 0.00a | 100.0 |
| Quercetin-O-hexoside-O-malonyl-hexoside isomer 2 | 0.05 ± 0.01a | 0.03 ± 0.00a | 60.0 | 0.04 ± 0.00a | 0.02 ± 0.00a | 50.0 | 0.06 ± 0.00a | 0.04 ± 0.00a | 66.7 |
| Quercetin-7-O-acetyl-hexoside | 0.07 ± 0.00a | 0.03 ± 0.00a | 42.9 | 0.05 ± 0.00a | 0.03 ± 0.00a | 60.0 | 0.06 ± 0.00a | 0.03 ± 0.00a | 50.0 |
| Quercetin-O-hexoside-O-acetyl-hexoside | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.01 ± 0.00 | 0.01 ± 0.00 | 100.0 |
| Isorhamnetin | 9.14 ± 0.52a | n.d. | 0 | 8.66 ± 0.51a | 8.30 ± 0.60a | 95.8 | 9.78 ± 0.61a | n.d. | 0 |
| Isorhamnetin-3-O-hexoside | 0.09 ± 0.00a | 0.13 ± 0.01b | 144.4 | 0.20 ± 0.01c | 0.12 ± 0.01b | 65.0 | 0.27 ± 0.01d | 0.16 ± 0.01b | 66.7 |
| Isorhamnetin-4′-O-hexoside | 5.63 ± 0.20a | 3.49 ± 0.14b | 62.0 | 9.73 ± 0.63c | 5.48 ± 0.35a | 56.3 | 5.84 ± 0.67a | 2.68 ± 0.16d | 45.9 |
| Isorhamnetin-O-hexoside-O-pentoside | 0.02 ± 0.00a | n.d. | 0 | 0.03 ± 0.00a | 0.02 ± 0.00a | 66.7 | 0.02 ± 0.00a | 0.02 ± 0.00a | 100.0 |
| Isorhamnetin-3-O-hexoside-4′-O-hexoside | 0.40 ± 0.02a | 0.73 ± 0.03b | 182.5 | 1.43 ± 0.08c | 1.38 ± 0.06c | 96.5 | 1.40 ± 0.06c | 1.04 ± 0.03d | 74.3 |
| Kaempferol-7-O-hexoside-isomer 1 | 0.35 ± 0.02a | n.d. | 0 | 0.32 ± 0.02a | 0.03 ± 0.00b | 9.4 | 0.91 ± 0.07c | 0.63 ± 0.04d | 69.2 |
| Kaempferol-7-O-hexoside-isomer 2 | 0.01 ± 0.00a | 0.01 ± 0.00a | 100.0 | 0.02 ± 0.00a | 0.01 ± 0.00a | 50.0 | 0.01 ± 0.00a | 0.01 ± 0.00a | 100.0 |
| Kaempferol-3-O-hexoside | 0.10 ± 0.00a | 0.05 ± 0.00b | 50.0 | 0.07 ± 0.00b | 0.03 ± 0.00a | 42.9 | 0.04 ± 0.00a | 0.02 ± 0.00a | 50.0 |
| Kaempferol-O-hexoside-O-hexoside isomer 1 | 0.71 ± 0.05a | 0.01 ± 0.00b | 1.4 | 0.33 ± 0.02c | 0.03 ± 0.00b | 9.1 | 0.81 ± 0.06a | 0.02 ± 0.00b | 2.5 |
| Kaempferol-O-hexoside-O-hexoside isomer 2 | 0.08 ± 0.00a | 0.05 ± 0.00b | 62.5 | 0.05 ± 0.00b | 0.04 ± 0.00b | 80.0 | 0.04 ± 0.00b | 0.03 ± 0.00b | 75.0 |
| Total flavonols | 126.36 ± 3.92 a | 38.82 ± 1.50 b | 30.7 | 148.29 ± 3.58 c | 83.65 ± 2.74 d | 56.4 | 175.52 ± 5.16 e | 57.39 ± 2.25 f | 32.7 |
| Other flavonoids | |||||||||
| Epicatechin-O-hexoside isomer 1 | 0.02 ± 0.00a | n.d. | 0 | 0.01 ± 0.00a | n.d. | 0 | 0.01 ± 0.00a | n.d. | 0 |
| Epicatechin-O-hexoside isomer 2 | 0.01 ± 0.00a | n.d. | 0 | n.d. | n.d. | n.d. | 0.01 ± 0.00a | n.d. | 0 |
| Epicatechin-O-hexoside isomer 3 | 0.01 ± 0.00a | n.d. | 0 | n.d. | n.d. | n.d. | 0.01 ± 0.00a | n.d. | 0 |
| Taxifolin-O-hexoside isomer 1 | 0.06 ± 0.00a | 0.01 ± 0.00b | 16.7 | 0.09 ± 0.00a | 0.05 ± 0.00a | 55.6 | 0.02 ± 0.00b | 0.01 ± 0.00b | 50.0 |
| Taxifolin-O-hexoside isomer 2 | 0.16 ± 0.01a | n.d. | 0 | 0.20 ± 0.01a | 0.08 ± 0.00b | 40.0 | 0.11 ± 0.01b | n.d. | 0 |
| Taxifolin-O-hexoside isomer 3 | 0.11 ± 0.01a | n.d. | 0 | 0.11 ± 0.01a | n.d. | 0 | 0.15 ± 0.01b | n.d. | 0 |
| Taxifolin-O-hexoside isomer 4 | 0.06 ± 0.00a | 0.01 ± 0.00b | 16.7 | 0.10 ± 0.01c | 0.04 ± 0.00a | 40.0 | 0.04 ± 0.00a | 0.01 ± 0.00b | 25.0 |
| Total other flavonoids | 0.43 ± 0.01 a | 0.02 ± 0.00 b | 4.7 | 0.51 ± 0.02 c | 0.17 ± 0.00 d | 33.3 | 0.35 ± 0.01 e | 0.02 ± 0.00 b | 5.7 |
| Anthocyanins | |||||||||
| Cyanidin-3-O-hexoside isomer 1 | 1.86 ± 0.15a | 0.02 ± 0.00b | 1.1 | 1.40 ± 0.11c | 0.11 ± 0.01d | 7.9 | 2.11 ± 0.18a | 0.09 ± 0.01d | 4.3 |
| Cyanidin-3-O-hexoside isomer 2 | 0.09 ± 0.01a | 0.05 ± 0.00b | 55.6 | 0.06 ± 0.00b | 0.02 ± 0.00c | 33.3 | 0.06 ± 0.00b | 0.02 ± 0.00c | 50.0 |
| Peonidin-3-O-hexoside | 0.13 ± 0.01a | n.d. | 0 | 0.19 ± 0.02b | 0.02 ± 0.00c | 10.5 | 0.16 ± 0.01b | 0.09 ± 0.00d | 56.3 |
| Cyanidin-O-malonyl-hexoside | 8.99 ± 0.74a | 0.19 ± 0.02b | 2.1 | 6.15 ± 0.51c | 0.86 ± 0.07d | 14.0 | 7.51 ± 0.48c | 0.43 ± 0.03e | 5.7 |
| Peonidin-O-malonyl-hexoside | 0.16 ± 0.01a | 0.01 ± 0.00b | 6.3 | 0.14 ± 0.01a | 0.03 ± 0.00b | 21.4 | 0.14 ± 0.01a | 0.01 ± 0.00b | 7.1 |
| Cyanidin-O-hexoside-O-hexoside isomer 1 | 0.10 ± 0.01a | 0.01 ± 0.00b | 10.0 | 1.01 ± 0.09c | n.d. | 0 | 0.21 ± 0.02d | 0.03 ± 0.00b | 14.3 |
| Cyanidin-O-hexoside-O-hexoside isomer 2 | 1.63 ± 0.12a | 0.03 ± 0.00b | 1.8 | 0.04 ± 0.00b | 0.09 ± 0.01c | 225.0 | 1.52 ± 0.11a | 0.07 ± 0.00c | 4.6 |
| Cyanidin-O-hexoside-O-hexoside isomer 3 | 0.05 ± 0.00a | 0.05 ± 0.00a | 100.0 | 0.03 ± 0.00a | 0.03 ± 0.00a | 100.0 | 0.04 ± 0.00a | 0.03 ± 0.00a | 75.0 |
| Cyanidin-O-hexoside-O-malonyl-hexoside isomer 1 | n.d. | n.d. | n.d. | 0.05 ± 0.00a | 0.01 ± 0.00b | 20.0 | 0.10 ± 0.01c | 0.01 ± 0.00b | 10.0 |
| Cyanidin-O-hexoside-O-malonyl-hexoside isomer 2 | 3.49 ± 0.29a | 0.06 ± 0.00b | 1.7 | 1.55 ± 0.10c | 0.20 ± 0.01d | 12.9 | 2.58 ± 0.19e | 0.16 ± 0.01d | 6.2 |
| Total anthocyanins | 16.50 ± 0.82 a | 0.42 ± 0.02 b | 2.5 | 10.62 ± 0.54 c | 1.37 ± 0.07 d | 12.9 | 14.43 ± 0.56 a | 0.94 ± 0.03 e | 6.5 |
| Total | 148.06 ± 4.01 a | 45.26 ± 1.52 b | 30.6 | 165.99 ± 3.62 c | 92.05 ± 2.75 d | 55.5 | 193.01 ± 5.19 e | 63.21 ± 2.25 | 32.7 |
Both applied thermal treatments induced an increase in the amount of extractable total phenolic compounds in RSO, mainly as a consequence of the matrix softening effect and the loss of water, which occurred during cooking, as previously suggested.1,2 Specifically, grilling resulted in a 30.4% increase in total phenolic compounds, whereas the recorded increase was 12.1% in fried RSO. The increase in phenolic compounds in both the grilled (GRSO) and fried (FRSO) red-skinned onion is attributed to the increase in the concentration of quercetin-4′-O-glucoside and quercetin-3-O-glucoside-4′-O-glucoside. Conversely, the concentration of quercetin decreased in both GRSO and FRSO in comparison with the raw sample. Similar results were previously reported.1,2,39,40
Both grilling and frying also positively affected the release of onion phenolic compounds during in vitro gastro-intestinal digestion (Table 1). The highest amount of released phenolic compounds after in vitro digestion was found for the FRSO sample (92.05 ± 2.75 μmol per 100 g), which also showed the highest bioaccessibility index (55.5%). The RRSO and GRSO samples displayed a similar bioaccessibility index for total phenolic compounds (30.6% and 32.7%, respectively) although in vitro digestion of GRSO resulted in a significantly higher amount of bioaccessible phenolic compounds compared to RRSO (63.21 ± 2.25 and 45.26 ± 1.52 μmol per 100 g, respectively).
The highest bioaccessibility index of FRSO was mainly due to the high bioaccessibility of quercetin (48.0%), which was present in a trace amount or not detected in the in vitro digested RRSO and GRSO. As previously suggested, the highest concentration of quercetin in in vitro digested FRSO compared to RRSO and GRSO may be due to the presence of oil, which favoured the solubility and stability of quercetin during digestion.2,41 Similar behaviour was also observed for the aglycone isorhamnetin, which displayed a bioaccessibility index close to 100% in FRSO, whereas not detected in the digested RRSO and GRSO. Considering the behaviour of the other two most important flavonols, quercetin-3-O-glucoside-4′-O-glucoside was characterized by a high bioaccessibility index in all the samples (from 74.3% to 98.3% in FRSO and RRSO, respectively), whereas quercetin-4′-O-glucoside only showed a high bioaccessibility index in RRSO (Table 1). In all the RSO digested samples, the lowest bioaccessibility index was found for anthocyanins (from 2.5% in RRSO to 12.9% in FRSO), whereas the class of hydroxybenzoic acids was always characterized by a bioaccessibility index of 100%. This indicates the occurrence of the oxidative degradation of quercetin-mono-glucosides and quercetin during in vitro digestion, which resulted in the production of protocatechuic acid and di-hydroxybenzoic acid-hexosides.1,42,43
3.2. Red-skinned onion phenolic compound metabolism during in vitro colonic fermentation
The absorption and bioavailability of quercetin and glycosylated quercetin-derivatives have been fairly studied in the past few years.44,45 In general, quercetin-mono-glucosides (such as quercetin-4′-O-glucoside and quercetin-3-O-glucoside) are quite well absorbed in the small intestine, reaching 50% of the ingested dose.46 The absorption pathway involves the hydrolysis of the glucose moiety by brush-border glucosidases (such as lactase phlorizin hydrolase) and the absorption of the quercetin aglycone, which is sufficiently hydrophobic to cross the membrane of enterocytes by passive diffusion.47 Differently, the bioavailability of quercetin aglycone is much lower than that of the glucoside-derivatives (about 20% of the ingested doses) mainly due to its low bioaccessibility and solubility in the intestinal fluid.48 Therefore, most of the quercetin and its derivatives present in onion are supposed to reach the colon, where they are thoroughly metabolized by the colon microbiota. The fecal metabolism of quercetin (and its derivatives) involves deglycosylation reaction, followed by C-ring fission, which leads to the production of several low-molecular weight phenolic acids.49,50 Although there are some in vitro studies on the elucidation of colonic quercetin metabolism, there are no studies in the literature on the investigation of the effect of cooking on the colonic metabolism of onion quercetin and the changes in the microbiota. Therefore, the in vitro digested RSO samples were further subjected to in vitro faecal fermentation and the generated metabolites were identified and quantified by means of high-resolution mass spectrometry at BL, T1 and EP of fermentation (Tables 2–4).| Compound | Time points of colonic fermentation | ||
|---|---|---|---|
| BL | T1 | EP | |
| Different superscript letters (a–c) within the same row mean significant different (p < 0.05) values; n.d. = not detected compound; BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); and EP = endpoint (27.48 h). | |||
| Hydroxybenzoic acid isomer 1 | 0.47 ± 0.02a | 1.18 ± 0.09b | 1.17 ± 0.11b |
| Hydroxybenzoic acid isomer 2 | n.d. | 0.71 ± 0.05a | 0.70 ± 0.05a |
| 3-(3′-Hydroxyphenyl)acetic acid | 7.51 ± 0.61a | 7.93 ± 0.59a | 12.49 ± 1.00b |
| 3-(4′-Hydroxyphenyl)acetic acid | n.d. | n.d. | n.d. |
| 3,4-Dihydroxybenzoic acid | 1.89 ± 0.11a | 1.98 ± 0.15a | 2.13 ± 0.11a |
| Dihydroxybenzoic acid isomer 1 | 0.31 ± 0.01a | n.d. | n.d. |
| Dihydroxybenzoic acid isomer 2 | n.d. | n.d. | n.d. |
| 3-(3′-Hydroxyphenyl)propanoic acid | 183.31 ± 11.29a | 32.66 ± 2.36b | 39.50 ± 1.63c |
| 3-(4′-Hydroxyphenyl)propanoic acid | 0.20 ± 0.01a | 2.69 ± 0.19b | 6.12 ± 0.48c |
| 4-Hydroxy-3-methoxybenzoic acid | 0.21 ± 0.01a | 0.27 ± 0.02b | 0.29 ± 0.01b |
| 3-(3′,4′-Dihydroxyphenyl)acetic acid | 0.68 ± 0.02a | 1.79 ± 0.16b | 3.05 ± 0.11c |
| 3′,4′-Dihydroxycinnamic acid | 0.41 ± 0.02a | 0.27 ± 0.02b | 0.29 ± 0.02b |
| (Dihydroxyphenyl)propanoic acid | 0.22 ± 0.01a | n.d. | 0.05 ± 0.00b |
| 3-(3′,4′-Dihydroxyphenyl)propanoic acid | 0.14 ± 0.01a | 1.25 ± 0.09b | 0.19 ± 0.01c |
| 3-(4′-Hydroxy-3′-methoxyphenyl)acetic acid | 0.02 ± 0.00a | 0.02 ± 0.00a | 0.08 ± 0.00b |
| 3-(4′-Hydroxy-3′-methoxyphenyl)propanoic acid | 0.68 ± 0.04a | 0.66 ± 0.05a | 1.16 ± 0.09b |
| Luteolin | 7.51 ± 0.51a | 3.96 ± 0.28b | 3.39 ± 0.21b |
| Quercetin | 96.41 ± 5.66a | 1.09 ± 0.10b | n.d. |
| Isorhamnetin | 17.39 ± 1.08a | 0.43 ± 0.03b | 0.20 ± 0.02c |
| Total | 317.36 ± 12.70a | 56.89 ± 2.47b | 70.81 ± 1.99c |
| Compound | Time points of colonic fermentation | ||
|---|---|---|---|
| BL | T1 | EP | |
| Different superscript letters (a–c) within the same row mean significant different (p < 0.05) values; n.d. = not detected compound; BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); and EP = endpoint (27.48 h). | |||
| Hydroxybenzoic acid isomer 1 | 0.44 ± 0.03a | 1.24 ± 0.10b | 1.08 ± 0.09b |
| Hydroxybenzoic acid isomer 2 | 0.11 ± 0.00a | 1.01 ± 0.10b | 1.09 ± 0.09b |
| 3-(3′-Hydroxyphenyl)acetic acid | 7.83 ± 0.58a | 11.69 ± 0.99b | 12.84 ± 1.13b |
| 3-(4′-Hydroxyphenyl)acetic acid | n.d. | 15.53 ± 0.81a | 8.58 ± 0.72b |
| 3,4-Dihydroxybenzoic acid | 1.45 ± 0.11a | 6.83 ± 0.49b | 1.52 ± 0.11a |
| Dihydroxybenzoic acid isomer 1 | 0.24 ± 0.02a | n.d. | n.d. |
| Dihydroxybenzoic acid isomer 2 | n.d. | n.d. | n.d. |
| 3-(3′-Hydroxyphenyl)propanoic acid | 170.43 ± 12.84a | 49.70 ± 2.92b | 44.20 ± 2.66b |
| 3-(4′-Hydroxyphenyl)propanoic acid | 0.17 ± 0.01a | 4.13 ± 0.28b | 4.13 ± 0.36b |
| 4-Hydroxy-3-methoxybenzoic acid | 0.19 ± 0.01a | 0.24 ± 0.02a | 0.19 ± 0.01a |
| 3-(3′,4′-Dihydroxyphenyl)acetic acid | 1.16 ± 0.09a | 10.68 ± 0.61b | 14.40 ± 0.97c |
| 3′,4′-Dihydroxycinnamic acid | 0.37 ± 0.01a | 0.41 ± 0.03a | 0.35 ± 0.03a |
| (Dihydroxyphenyl)propanoic acid | 0.29 ± 0.02a | 2.01 ± 0.11b | 0.61 ± 0.04c |
| 3-(3′,4′-Dihydroxyphenyl)propanoic acid | 0.12 ± 0.00a | 0.09 ± 0.00b | 0.15 ± 0.01c |
| 3-(4′-Hydroxy-3′-methoxyphenyl)acetic acid | 0.01 ± 0.00a | 0.11 ± 0.00b | 0.12 ± 0.01b |
| 3-(4′-Hydroxy-3′-methoxyphenyl)propanoic acid | 0.13 ± 0.01a | 0.63 ± 0.05b | n.d. |
| Luteolin | 7.97 ± 0.60a | 7.62 ± 0.45a | 4.39 ± 0.31b |
| Quercetin | 121.16 ± 8.22a | 12.44 ± 1.02b | n.d. |
| Isorhamnetin | 37.99 ± 2.18a | 4.48 ± 0.36b | 1.58 ± 0.09c |
| Total | 350.06 ± 15.42a | 128.84 ± 3.41b | 95.23 ± 3.09c |
| Compound | Time points of colonic fermentation | ||
|---|---|---|---|
| BL | T1 | EP | |
| Different superscript letters (a–c) within the same row mean significant different (p < 0.05) values; n.d. = not detected compound; BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); and EP = endpoint (27.48 h). | |||
| Hydroxybenzoic acid isomer 1 | 0.47 ± 0.03a | 1.08 ± 0.08b | 1.15 ± 0.10b |
| Hydroxybenzoic acid isomer 2 | n.d. | 1.14 ± 0.09a | 2.21 ± 0.12b |
| 3-(3′-Hydroxyphenyl)acetic acid | 8.45 ± 0.78a | 11.86 ± 1.28b | 13.53 ± 0.97b |
| 3-(4′-Hydroxyphenyl)acetic acid | n.d. | n.d. | n.d. |
| 3,4-Dihydroxybenzoic acid | 1.74 ± 0.14a | 2.22 ± 0.11b | 2.12 ± 0.09b |
| Dihydroxybenzoic acid isomer 1 | 0.99 ± 0.10a | n.d. | n.d. |
| Dihydroxybenzoic acid isomer 2 | 0.31 ± 0.03a | n.d. | n.d. |
| 3-(3′-Hydroxyphenyl)propanoic acid | 203.89 ± 11.64a | 76.01 ± 4.58b | 91.32 ± 6.64c |
| 3-(4′-Hydroxyphenyl)propanoic acid | 0.20 ± 0.01a | 1.49 ± 0.09b | 1.88 ± 0.12c |
| 4-Hydroxy-3-methoxybenzoic acid | 0.27 ± 0.02a | 0.27 ± 0.02a | 0.31 ± 0.02a |
| 3-(3′,4′-Dihydroxyphenyl)acetic acid | 0.94 ± 0.08a | 2.56 ± 0.19b | 3.37 ± 0.19c |
| 3′,4′-Dihydroxycinnamic acid | 0.40 ± 0.03a | 0.32 ± 0.02b | 0.29 ± 0.02b |
| (Dihydroxyphenyl)propanoic acid | 0.28 ± 0.02a | 1.85 ± 0.11b | 1.54 ± 0.10c |
| 3-(3′,4′-Dihydroxyphenyl)propanoic acid | 0.15 ± 0.01a | 0.03 ± 0.00b | 0.02 ± 0.00b |
| 3-(4′-Hydroxy-3′-methoxyphenyl)acetic acid | 0.02 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a |
| 3-(4′-Hydroxy-3′-methoxyphenyl)propanoic acid | 0.20 ± 0.01a | n.d. | n.d. |
| Luteolin | 7.81 ± 0.57a | 6.64 ± 0.46b | 5.97 ± 0.32b |
| Quercetin | 144.91 ± 7.31a | 8.50 ± 0.71b | n.d. |
| Isorhamnetin | 18.92 ± 1.04a | 1.23 ± 0.09b | 0.32 ± 0.03c |
| Total | 389.95 ± 13.82a | 115.21 ± 4.84b | 124.04 ± 6.73b |
In all the RSO samples, the quercetin-glucosides and isorhamnetin-glucosides were rapidly hydrolysed to the corresponding aglycones given that they almost completely disappeared already at the BL of fermentation. These results are in agreement with previous studies that showed the complete hydrolysis of O-glucosides of flavonols in the first hours of colon fermentation.39,51 In addition to quercetin and isorhamnetin aglycones, luteolin aglycone also appeared at the BL of fermentation in all the RSO samples. Given that luteolin aglycone and luteolin-derivatives were not identified in the RSO samples, we speculate that the luteolin aglycone was derived from quercetin dehydroxylation at the C3 position in the C-ring, as already suggested.39 The concentration of quercetin and isorhamnetin aglycones strongly decreased at T1 of fermentation and quercetin was no longer detectable at the EP of fermentation (Tables 2–4).
The metabolite found at the highest concentration in all the RSO samples at the BL of fermentation was 3-(3′-hydroxyphenyl)propanoic acid, which represented 57.8%, 48.7% and 52.3% of the total metabolites in RRSO, GRSO and FRSO, respectively. The additional metabolites identified in an appreciable amount at the BL of fermentation were 3-(3′-hydroxyphenyl)acetic acid, 3,4-dihydroxybenzoic acid and 3-(3′,4′-dihydroxyphenyl)acetic acid. At the BL of fermentation, only quantitative differences in the amount of total metabolites were found among the samples. The highest concentration of metabolites was found in GRSO, followed by FRSO and RRSO. In all the RSO samples, the concentration of metabolites at the BL of fermentation strongly exceeded the amount of parent phenolic compounds identified in onion, suggesting the presence of a high amount of bound phenolic compounds in onion, which can be released from fiber or other macromolecules by the action of the gut bacteria.
At the end of the fermentation process, a decline in the concentration of phenolic metabolites was observed and GRSO once again was the sample releasing the highest amount of phenolic metabolites. In all the samples, 3-(3′-hydroxyphenyl)propanoic acid was still the metabolite present at the highest concentration although with a different incidence depending on the sample. In GRSO, 3-(3′-hydroxyphenyl)propanoic acid accounted for 73.6% of the total metabolites, whereas in FRSO and RRSO its incidence was lower (46.4% and 55.8%, respectively). In addition, the other important metabolites were found in different proportions depending on the thermal treatment at the end of the fermentation process. RRSO was the richest in terms of percentage of incidence of 3-(3′-hydroxyphenyl)acetic acid (17.6% of total phenolic compounds) followed by FRSO (13.4% of total phenolic compounds) and GRSO (10.4% of total phenolic compounds). Moreover, in the FRSO sample, the metabolite 3-(3′,4′-dihydroxyphenyl)acetic acid was found in high proportion, representing the 15.1% of total metabolites, whereas it represented only 2.7% and 4.3% of the total metabolites in GRSO and RRSO, respectively. Generally, the three metabolites 3-(3′-hydroxyphenyl)propanoic acid, 3-(3′-hydroxyphenyl)acetic acid and 3-(3′,4′-dihydroxyphenyl)acetic acid tended to accumulate in a high amount in the fermented samples but with a different proportion depending on the sample. Accordingly, cooking may modulate the metabolite profile after fermentation both from a quantitative and qualitative point-of-view.
Fig. 1 displays the colonic microbiota metabolic pathway of red-skinned onion flavonols. The first step is the de-glycosylation of quercetin-glucosides and isorhamnetin-glucosides into the corresponding aglycones quercetin and isorhamnetin.39,51 The appearance of luteolin, which was not present as aglycone or any derivatives, suggested the possible de-hydroxylation of quercetin.39 The next step in the flavonols metabolism is ring fission, leading to the production of 3-(3′,4′-dihydroxyphenyl)propanoic acid in the case of quercetin and luteolin or 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid in the case of isorhamnetin.49 Ring fission of quercetin may also result in the production of 3,4-dihydroxybenzoic acid.52 The metabolic intermediate 3-(3′,4′-dihydroxyphenyl)propanoic acid may have different fates, as depicted in Fig. 1. The major metabolic pathway for this intermediate was dehydroxylation to 3-(3′-hydroxyphenyl)propanoic acid, which was the major metabolite found at any time in each RSO sample during fermentation.10 Alternatively, 3-(3′,4′-dihydroxyphenyl)propanoic acid may undergo an α-oxidation step, producing 3-(3′,4′-dihydroxyphenyl)acetic acid, which can be further de-hydroxylated to 3-(3′-hydroxyphenyl)acetic acid or subjected to a second α-oxidation step, releasing 3,4-dihydroxybenzoic acid.13 Additional metabolic pathways involve the direct β-oxidation of 3-(3′,4′-dihydroxyphenyl)propanoic acid, producing 3,4-dihydroxybenzoic acid.53 All these metabolites were found in a high amount in the fermented RSO samples, suggesting that the described pathways are predominant during the fermentation of flavonols. Regarding isorhamnetin metabolism, the C-ring fission product 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid may undergo β-oxidation or two steps of α-oxidation to produce 4-hydroxy-3-methoxybenzoic acid. Otherwise, 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid may be demethoxylated, producing 3-(4′-hydroxyphenyl)propanoic acid, which can subsequently be subjected to α-oxidation, generating 3-(4′-hydroxyphenyl)acetic acid.
 | ||
| Fig. 1 Proposed metabolic pathways for phenolic compounds microbial metabolism after in vitro gastro-intestinal digestion and colonic fermentation of red-skinned onion. | ||
However, all these metabolites were found in a low concentration in the fermented samples, suggesting a possible alternative metabolic pathways for 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid. Indeed, this last intermediate may be demethylated, resulting in the formation of 3-(3′,4′-dihydroxyphenyl)propanoic acid.10,54,55
Phenolic metabolites generated by the action of gut microbiota are generally considered to be better absorbed and responsible for the beneficial effects following the consumption of phenolic compound-rich food.15 Several in vitro and in vivo studies demonstrated the biological potential of these metabolites. For example, 3-(3′-hydroxyphenyl)propanoic acid displayed in vitro anti-inflammatory and anti-diabetic activities.55–57 Furthermore, 3-(3′-hydroxyphenyl)propanoic acid and 3-(3′-hydroxyphenyl)acetic acid exhibited a blood pressure-reducing effect ex vivo and in vivo.58,59 Moreover, 3-(3′,4′-dihydroxyphenyl)acetic acid was reported to inhibit colon cancer cell proliferation more efficiently than 3-(3′-hydroxyphenyl)propanoic acid and 3-(3′-hydroxyphenyl)acetic acid.60 Finally, all three metabolites showed neuroprotective activity in vitro.61
3.3. Shift of total bacteria and microbiota eubiosis
qPCR absolute quantification targeted 14 different bacterial taxa (Table S1†) related to the core microbiota, including total Eubacteria and the two main phyla of the human healthy colon, i.e., Bacteroidetes and Firmicutes. Considering the total Eubacteria, compared to the abundances at the baseline (BL) and besides the values of the blank control (BC), just RRSO at T1 and EP was significantly overrepresented up to 1.43 folds more at the EP (p < 0.05) (Table 5). Conversely, both samples of cooked onions, independently from the time points of fermentation, were underrepresented, in particular, GRSO had slight not significant reduction (p > 0.05), but FRSO had a 1.68 fold significant reduction. These evident results indicated that the cooked onion samples, similar to the BC, were limited or not able to support the growth of total colon Eubacteria. The quantification of the Bacteroidetes phylum showed significant changes at each time point for all the samples tested, including the BC, with respect to the BL (p < 0.05) (Table 5). In particular, fermentation of RRSO generated the lowest underrepresentation of Bacteroidetes, up to 1.54-fold less at the EP, and FRSO generated the highest underrepresentation, up to 6.44-fold less at the EP. In detail, FRSO brought a reduction of 4.18 times higher than that brought by RRSO. The relatively low loads of Bacteroidetes after FRSO fermentation indicate that this cooking process can particularly reduce the availability of fiber for fermentation, given that many genera of this phylum are fibrolytic.62 Considering Firmicutes (Table 5), overrepresentations were seen for each sample at both time points of fermentation, although almost all not significantly with respect to the BL (p > 0.05). The sole time sample and time point that scored a slight increase, but significant in abundance was RRSO at the EP with just 1.00log2(FC). This latter increase was significantly higher than that of GRSO and FRSO at the EP (p > 0.05), but not than the BC (p > 0.05). These outputs indicate opposite scenarios depending on which group of Firmicutes was the most fostered, where the positive outlook could be due to the increase in Lactobacillales, while the negative due to the increase in Clostridiaceae.
| Target & sample | Quantifications at baseline (cells per mL) | Changes in log2(F/C) | MANOVAa | |||
|---|---|---|---|---|---|---|
| BL raw valuesb | BL ± SDc | T1 | EP | p (time) | −log10(p) | |
| Upperscore letters (A–C) are for statistical difference within “food matrix” and lowerscore letters (a–c) are for statistical difference within “time “, according to Duncan's multiple range post hoc test (p < 0.05); −log10(p) = significance of log2(F/C); BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); EP = endpoint (27.48 h); RRSO = raw red skinned onion; GRSO = grilled red skinned onion; FRSO = fried red skinned onion; and BC = blank control.a Multivariate ANOVA model with category “time” on column and category “food matrix” on rows (p < 0.05).b BL raw values are the mean of sextuplicates from each sample at the beginning of its colonic fermentations.c BL ± SD is BL mean ± standard deviation and obtained from BL raw values. | ||||||
| Eubacteria | ||||||
| RRSO | 5.13 × 108 | 7.42 × 108 ± 2.08 × 108b |
0.47abA | 1.43aA | 0.001469 | 2.833075 |
| GRSO | 6.30 × 108 | 7.42 × 108 ± 2.08 × 108a |
−0.36abB | −0.81bB | 0.000122 | 3.930721 |
| FRSO | 8.59 × 108 | 7.42 × 108 ± 2.08 × 108a |
−0.55abB | −1.68bB | 0.000970 | 3.013253 |
| BC | 9.67 × 108 | 7.42 × 108 ± 2.08 × 108a |
−0.71aB | −4.21bC | 0.002812 | 2.551055 |
| p (food matrix) | 0.726777 | 0.002947 | 0.000004 | |||
| Firmicutes | ||||||
| RRSO | 4.80 × 107 | 5.25 × 107 ± 6.86 × 106b |
0.66abA | 1.00aA | 0.029213 | 1.534484 |
| GRSO | 6.20 × 107 | 5.25 × 107 ± 6.86 × 106 | 0.27B | 0.28B | 0.272088 | 0.565290 |
| FRSO | 4.69 × 107 | 5.25 × 107 ± 6.86 × 106b |
0.39aAB | 0.27aB | 0.000182 | 3.750942 |
| BC | 5.32 × 107 | 5.25 × 107 ± 6.86 × 106b |
0.44abA | 0.76aAB | 0.001091 | 2.962952 |
| p (food matrix) | 0.589376 | 0.000126 | 0.000002 | |||
| Bacteroidetes | ||||||
| RRSO | 2.37 × 108 | 1.74 × 108 ± 6.44 × 107a |
−1.36bA | −1.54bA | 0.017556 | 1.755573 |
| GRSO | 2.21 × 108 | 1.74 × 108 ± 6.44 × 107a |
−6.05bC | −5.94bB | 0.002734 | 2.563179 |
| FRSO | 1.05 × 108 | 1.74 × 108 ± 6.44 × 107a |
−6.30bC | −6.44bB | 0.002727 | 2.564311 |
| BC | 1.35 × 108 | 1.74 × 108 ± 6.44 × 107a |
−3.46bB | −4.16bB | 0.003134 | 2.503885 |
| p (food matrix) | 0.972251 | 0.000018 | 0.000003 | |||
As a general parameter for microbiota eubiosis, we chose the Firmicutes/Bacteroidetes (F/B) ratio (Table 6), where a value of over two is usually linked to microbiota dysbiosis.8,10,63–66 The fecal samples at the baseline had an F/B ratio of 0.30 and this eubiosis condition was strongly maintained after 24 h just by RRSO (0.88), while the cooked onion samples failed. In particular, GRSO had a rise in the ratio up to 1.90, and FRSO overcame significantly with respect to the BL the dysbiotic value of 2.00 (p < 0.05). These results indicate that during the 24 h of fermentation, just the raw onion samples did not perturb the colon core microbiota of the healthy donors and could provide a nutritionally adapted substrate. Oppositely, fermentation of FRSO triggered a dysbiosis condition, which mirrored the lower Bacteroidetes content.
| Target & sample | Firmicutes to Bacteroidetes ratio (F/B) | MANOVAa | ||
|---|---|---|---|---|
| BL ± SDb | T1 | EP | p (time) | |
| Upperscore letters (A–C) are for statistical difference within “food matrix” and lowerscore letters (a–c) are for statistical difference within “time”, according to Duncan's multiple range post hoc test (p < 0.05). BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); EP = endpoint (27.48 h); RRSO = raw red skinned onion; GRSO = grilled red skinned onion; FRSO = fried red skinned onion; and BC = blank control.a Multivariate ANOVA model with category “time” on column and category “food matrix” on rows (p < 0.05).b BL ± SD is BL mean ± standard deviation and is obtained from BL raw values. | ||||
| RRSO | 0.30 ± 0.11b | 0.51 ± 0.45ab | 0.88 ± 0.77aC | 0.002296 |
| GRSO | 0.30 ± 0.11 | 1.64 ± 0.25 | 1.90 ± 0.69B | 0.230897 |
| FRSO | 0.30 ± 0.11b | 1.96 ± 0.90a | 2.05 ± 2.20aB | 0.004958 |
| BC | 0.30 ± 0.11c | 1.46 ± 0.15b | 4.08 ± 0.45aA | 0.016104 |
| p (food matrix) | 0.124932 | 0.328928 | 0.020129 | |
3.4. Shift in selected beneficial bacterial taxa
To evidence the shifts in colon core microbiota, we quantified the abundance of selected bacterial taxa known to be active responders to fiber substrate fermentation and modulators of host wellbeing (Table 7). The first set was related to beneficial microbes, while the second set was related to opportunistic and dysbiotic microbes. Lactobacillales and Bifidobacteriaceae with the exceptions of few species are related to the host's wellbeing and eubiosis of the colon microbiota.67Lactobacillales also include Lactobacillaceae and Enterococcaceae. These three families are implicated in the biotransformation of phenolic compounds, for example, Lactobacillaceae can increase the content of epicatechin as well as gallic, caffeic, and ferulic acids68 and Enterococcaceae can promote O-deglycosylation and C-ring fission of several flavonols,17 whereas Bifidobacteriaceae can metabolize homovanillic acids69 and hydrolyze chlorogenic acids.70 However, in our experiments, only the class of Lactobacillales was fostered by any onions fermentation, while targets such Bifidobacteriaceae and also specific Bifidobacterium longum underwent a large depletion (Table 7). In detail, the Lactobacillales were overrepresented in any sample, and still the raw onions almost doubled in abundance than both cooked onions. Thus, all the onion substrates once fermented by the colon microbiota increased the abundance of Lactobacillales significantly or not with respect to the BL and the BC. Similarly, in the case of long-term fermentation of phenolic compounds on gut models, other authors found that Enterococcaceae was the most fostered family.34,71| Targets & samples | Quantifications at baseline (cells per mL) | Changes in log2(F/C) | MANOVAa | |||
|---|---|---|---|---|---|---|
| BL raw valuesb | BL ± SDc | T1 | EP | p (time) | −log10(p) | |
| Upperscore letters (A–C) are for statistical difference within “food matrix” and lowerscore letters (a–c) are for statistical difference within “time”, according to Duncan's multiple range post hoc test (p < 0.05). −log10(p) = significance of log2(F/C); BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); EP = endpoint (27.48 h); RRSO = raw red skinned onion; GRSO = grilled red skinned onion; FRSO = fried red skinned onion; and BC = blank control.a Multivariate ANOVA model with category “time” on column and category “food matrix” on rows (p < 0.05).b BL raw values are the means of sextuplicates from each sample at the beginning of its fermentation.c BL ± SD is BL mean ± standard deviation and is obtained from BL raw values. | ||||||
| Lactobacillales | ||||||
| RRSO | 7.70 × 105 | 8.79 × 105 ± 1.32 × 105b |
1.05a | 1.90aA | 0.014742 | 1.831452 |
| GRSO | 1.05 × 106 | 8.79 × 105 ± 1.32 × 105b |
0.46ab | 0.97aAB | 0.000859 | 3.065816 |
| FRSO | 7.81 × 105 | 8.79 × 105 ± 1.32 × 105 | 0.38 | 1.03AB | 0.084676 | 1.072238 |
| BC | 9.16 × 105 | 8.79 × 105 ± 1.32 × 105 | −0.49 | −0.65B | 0.924597 | 0.034047 |
| p (food matrix) | 0.709415 | 0.369291 | 0.004476 | |||
| Bifidobacteriaceae | ||||||
| RRSO | 1.22 × 105 | 5.08 × 104 ± 5.24 × 104a |
−5.17bC | −5.06bC | 0.031567 | 1.500769 |
| GRSO | 5.77 × 104 | 5.08 × 104 ± 5.24 × 104a |
−5.11bC | −5.26bC | 0.012853 | 1.891000 |
| FRSO | 1.55 × 104 | 5.08 × 104 ± 5.24 × 104a |
−2.40bB | −3.27bB | 0.000041 | 4.388389 |
| BC | 7.85 × 103 | 5.08 × 104 ± 5.24 × 104 | 0.19A | 0.28A | 0.777184 | 0.109475 |
| p (food matrix) | 0.683959 | 0.000962 | 0.014112 | |||
| Bacteroides–Prevotella–Porphyromonas (BPP) group | ||||||
| RRSO | 2.34 × 107 | 2.09 × 107 ± 3.41 × 106a |
−0.34bA | −0.98bA | 0.001932 | 2.714073 |
| GRSO | 2.39 × 107 | 2.09 × 107 ± 3.41 × 106a |
−3.45bB | −3.17bB | 0.000005 | 5.324022 |
| FRSO | 1.95 × 107 | 2.09 × 107 ± 3.41 × 106a |
−2.58bB | −3.69bB | 0.000008 | 5.081399 |
| BC | 1.67 × 107 | 2.09 × 107 ± 3.41 × 106a |
−1.32bA | −1.10bA | 0.015634 | 1.805925 |
| p (food matrix) | 0.509081 | 0.000009 | 0.033771 | |||
| Clostridium IV (CLOS IV) group | ||||||
| RRSO | 1.25 × 107 | 1.79 × 107 ± 6.42 × 106 | 0.25B | 5.65A | 0.072745 | 1.138197 |
| GRSO | 2.72 × 107 | 1.79 × 107 ± 6.42 × 106b |
1.76aA | 2.53aB | 0.012853 | 1.891000 |
| FRSO | 1.57 × 107 | 1.79 × 107 ± 6.42 × 106b |
1.16bA | 2.99aB | 0.000511 | 3.291724 |
| BC | 1.61 × 107 | 1.79 × 107 ± 6.42 × 106a |
−2.28bC | −1.35bC | 0.021311 | 1.671394 |
| p (food matrix) | 0.643268 | 0.008103 | 0.009921 | |||
| Bifidobacterium longum | ||||||
| RRSO | 1.76 × 106 | 2.53 × 106 ± 1.91 × 106a |
−0.17Ab | −0.69Bb | 0.014742 | 1.831452 |
| GRSO | 5.04 × 106 | 2.53 × 106 ± 1.91 × 106b |
0.39abA | 0.71aA | 0.000859 | 3.065816 |
| FRSO | 5.25 × 105 | 2.53 × 106 ± 1.91 × 106 | 0.04A | 0.39A | 0.084676 | 1.072238 |
| BC | 2.78 × 106 | 2.53 × 106 ± 1.91 × 106 | −2.59B | −3.55C | 0.924597 | 0.034047 |
| p (food matrix) | 0.036929 | 0.042285 | 0.004473 | |||
| Akkermansia muciniphila | ||||||
| RRSO | 6.04 × 105 | 5.26 × 105 ± 8.23 × 104b |
0.51aA | 0.81aA | 0.000201 | 3.695973 |
| GRSO | 5.85 × 105 | 5.26 × 105 ± 8.23 × 104b |
0.45abA | 0.95aA | 0.000181 | 3.743186 |
| FRSO | 4.80 × 105 | 5.26 × 105 ± 8.23 × 104a |
0.06aA | −0.53bAB | 0.018797 | 1.725910 |
| BC | 4.34 × 105 | 5.26 × 105 ± 8.23 × 104a |
−4.47bB | −3.88bB | 0.000004 | 5.370729 |
| p (food matrix) | 0.645485 | <0.000001 | <0.000001 | |||
| Faecalibacterium prausnitzii | ||||||
| RRSO | 1.06 × 107 | 2.22 × 107 ± 7.79 × 106 | 0.59A | 0.93A | 0.272796 | 0.564162 |
| GRSO | 2.52 × 107 | 2.22 × 107 ± 7.79 × 106 | −0.32B | −1.60B | 0.098563 | 1.006284 |
| FRSO | 2.59 × 107 | 2.22 × 107 ± 7.79 × 106a |
−1.45bBC | −2.50bB | 0.019097 | 1.719023 |
| BC | 2.71 × 107 | 2.22 × 107 ± 7.79 × 106a |
−1.82bC | −4.74cC | 0.011480 | 1.940064 |
| p (food matrix) | 0.300806 | 0.001150 | 0.007717 | |||
Alternatively, according to our results, Bifidobacteriaceae and the B. longum taxa were not fostered by any of the onion substrates. In particular, the fact that a significant reduction with respect to the BL was achieved only on the onion substrates and not in the BC indicates that the high flavonol content of the samples somehow inhibited the sensitive taxon of Bifidobacteriaceae (Table 7). It has been reported that quercetin is inhibitory with respect to bifidobacteria,72 while other studies in similar in vitro environments reported that of this vital group was fostered.50 This scenario on beneficial core microbiota suggests that Lactobacillales are more specific in metabolizing onion flavonol compounds than Bifidobacteriaceae, and the latter group is more sensible to some detrimental compounds generated by the pyrolysis of cooked foods. It was reported that Bifidobacteriaceae are somehow inhibited by a certain class of phenolic compounds, e.g., that from black tea71 rather than others, e.g. that from olives.8 Additionally, it is known that Lactobacillales can resist a high content of phenolic compounds73 because they possess an enzymatic arsenal with a wide target range, enabling the de-esterification, hydrolysis, or conversion of phenolics to individual acids (gallic, quinic, caffeic, p-coumaric, ferulic, dihydrocaffeic, dihydroferulic acid, vinylcatechol, and vinylguaiacol).73,74 Also, Date and colleagues75 reported that onion can foster the growth of three probiotic lactobacilli after in vitro fermentation with fecal inoculum. Among the beneficial taxa, Clostridium group IV and Faecalibacterium prausnitzii are reported to be involved in the biotransformation of phenolic compounds.76 According to our results (Table 7), Clostridium group IV increased significantly at the EP in all the onion fermentations, but not in the BC. However, its resident F. prausnitzii was only fostered by the raw onions and limited by the cooked samples, even if less than twice than the BC.
The Clostridium group IV and its resident F. prausnitzii are important SCFA producers and their presence is associated with a healthy gut. A decrease in the amount of these bacteria is observed in patients with inflammatory bowel disease.77 Consistent with our results, these taxa were reported to increase after a dietary clinical trial with phenolic compounds from red wine.78,79
Lastly, Akkermansia muciniphila has been reported to be elevated in the presence of phenolic compounds, similar to some phenolic compounds acting as prebiotics.16 According to our results (Table 7), this taxon was fostered both at T1 and at EP by the RRSO and GRSO substrates but not significantly, while it was limited at the EP by both FRSO and by BC, which in comparison three times the strength by the latter. Another commensal group that is involved mainly in fiber degradation is the Bacteroides–Prevotella group, covered by the BPP (Bacteroides–Prevotella–Porphyromonas) target. Generally, these bacteria are sensible to both fibers80 and phenolic compounds. For example, Tamargo and colleagues34 observed a significant increase in this group after phenolic compound fermentation on a gut model. Alternatively, according to our results, but in line with our findings about the superior phylum of Bacteroidetes, the BPP group was significantly underrepresented at the end of all the fermentations, in a strength larger than the BC (Table 7). These results can be applied as indicators of health-related effects when RSO is consumed and can be translated to the in vivo condition.
3.5. Shift in selected opportunistic bacterial taxa
Considering the opportunistic bacterial taxa, we targeted Enterobacteriaceae, Clostridium group I, the Atopobium–Collinsella–Eggerthella (ATOP) group, and Escherichia coli (Table 8). Within these groups, we found the most important features on the modulation of gut microbiota by onion fermentation. Indeed, Enterobacteriaceae at both time points were inhibited just by RRSO, but the other samples limited their growth more than that in the BC. A similar trend was also detected by the resident Escherichia coli taxon. Clostridium group I was reduced almost in all cases at any time point, except for the BC, but significantly just for RRSO at the EP (p < 0.05). Lastly, considering the ATOP group, slight increases were observed just in the BC, while the onion samples were able to largely reduce the ATOP group loads up to growth inhibition due to the mean reduction of 4.53-fold less for any of the onion substrates with respect to the BL. Additionally, the RRSO and GRSO were better performers in action than FRSO. These groups engaged in the biotransformation of phenolic compounds because Atopobium parvulum, for example, possess tannases,81 while Atopobium spp. is correlated with protocatechuic acid production after colonic fermentation of berry phenolic compounds.82 However, according to our results, this scenario is oppositely described, giving the onion samples high capability in opposing opportunistic bacteria, such as Atopobium and Eggerthella. This feature is of high relevance given that it indicates that the consumption of RSO can protect the intestinal tract from colonization by opportunistic bacteria and pathogens, limiting possible dysbiosis.| Targets & sample | Quantifications at baseline (cells per mL) | Changes in log2(F/C) | MANOVAa | |||
|---|---|---|---|---|---|---|
| BL raw valuesb | BL ± SDc | T1 | EP | p (time) | −log10(p) | |
| Upperscore letters (A–C) are for statistical difference within “food matrix” and lowerscore letters (a–c) are for statistical difference within “time”, according to Duncan's multiple range post hoc test (p < 0.05); −log10(p) = significance of log2(F/C); BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); EP = endpoint (27.48 h); RRSO = raw red skinned onion; GRSO = grilled red skinned onion; FRSO = fried red skinned onion; and BC = blank control.a Multivariate ANOVA model with category “time” on column and category “food matrix” on rows (p < 0.05).b BL raw values are the means of sextuplicates from each sample at the beginning of its fermentations.c BL ± SD is BL mean ± standard deviation and is obtained from BL raw values. | ||||||
| Enterobacteriaceae | ||||||
| RRSO | 1.42 × 106 | 1.44 × 106 ± 2.98 × 105 | −0.69B | −0.87B | 0.199488 | 0.700083 |
| GRSO | 1.81 × 106 | 1.44 × 106 ± 2.98 × 105b |
0.90aAB | 0.72aB | 0.014941 | 1.825625 |
| FRSO | 1.08 × 106 | 1.44 × 106 ± 2.98 × 105b |
1.13abAB | 1.84aAB | 0.000012 | 4.914587 |
| BC | 1.44 × 106 | 1.44 × 106 ± 2.98 × 105b |
2.61aA | 3.62aA | 0.000011 | 4.956323 |
| p (food matrix) | 0.977079 | <0.000001 | <0.000001 | |||
| Clostridium group I (CLOS I) | ||||||
| RRSO | 8.91 × 103 | 1.51 × 104 ± 9.51 × 103 | 0.11 | −1.35 | 0.142556 | 0.700083 |
| GRSO | 5.19 × 103 | 1.51 × 104 ± 9.51 × 103 | −0.12 | −0.02 | 0.844734 | 1.825625 |
| FRSO | 2.17 × 104 | 1.51 × 104 ± 9.51 × 103a |
−0.10b | −0.81b | 0.038646 | 4.914587 |
| BC | 2.46 × 104 | 1.51 × 104 ± 9.51 × 103b |
0.16b | 1.09a | 0.033585 | 4.956323 |
| p (food matrix) | 0.262495 | 0.084737 | 0.086195 | |||
| Atopobium–Collinsella–Eggerthella (ATOP) group | ||||||
| RRSO | 1.22 × 105 | 5.08 × 104 ± 5.24 × 104 | −5.17B | −5.06B | 0.072155 | 1.141731 |
| GRSO | 5.77 × 104 | 5.08 × 104 ± 5.24 × 104 | −5.11B | −5.26B | 0.071840 | 1.143634 |
| FRSO | 1.55 × 104 | 5.08 × 104 ± 5.24 × 104 | −2.40B | −3.27B | 0.079729 | 1.098386 |
| BC | 7.85 × 103 | 5.08 × 104 ± 5.24 × 104 | 0.19A | 1.18A | 0.152951 | 0.815447 |
| p (food matrix) | 0.046520 | 0.000043 | <0.000001 | |||
| Escherichia coli | ||||||
| RRSO | 8.70 × 102 | 5.25 × 103 ± 5.37 × 103 | 0.30B | 0.02C | 0.829838 | 0.081006 |
| GRSO | 4.43 × 102 | 5.25 × 103 ± 5.37 × 103 | 0.55B | 1.61B | 0.279017 | 0.554369 |
| FRSO | 1.09 × 104 | 5.25 × 103 ± 5.37 × 103 | 2.81AB | 2.22B | 0.093314 | 1.030053 |
| BC | 8.82 × 103 | 5.25 × 103 ± 5.37 × 103b |
3.92aA | 4.57aA | 0.005090 | 2.293282 |
| p (food matrix) | 0.040281 | 0.000067 | 0.001695 | |||
3.6. qPCR prebiotic index
According to the results for qPI (Table 9), the fermented substrate with the best prebiotic activity at the EP of fermentation was RRSO, followed by GRSO and FRSO. Thus, FRSO had the lowest prebiotic index, except for that of the negative control (BC), although there were no significant differences among RSO samples. The different prebiotic indices of the samples can be attributed to the degradation of the structural composition of the onion fibers after cooking or transformation of the onion flavanols with prebiotic capacity. Similar to most known prebiotics (e.g., fructooligosaccharides), it is recognized that soluble fibers are excellent substrates for the production of postbiotics (e.g., short chain fatty acids) in the large intestine.24 Additionally, in line with the latest definition of prebiotics,83 there is evidence that some phenolics can foster the growth of beneficial bacteria, thus possessing a prebiotic nature.16 Anyhow, the prebiotic potential generated by any of the onion samples can be ascribed to the reduction and limitation of opportunistic growth rather than the fostering of beneficial bacteria. A previous study documented the prebiotic potential of onion, which could foster the growth of three main beneficial lactobacilli in just 12 h of faecal in vitro fermentation.75 Therefore, the relevance of our results is the confirmation of onion prebiotic activity even after cooking and intestinal digestion.| Sample | qPCR prebiotic index (qPI)a | MANOVAb | |
|---|---|---|---|
| T1 | EP | p (time) | |
| Upperscore letters (A–C) are for statistical difference within “food matrix” and lowerscore letters (a–c) are for statistical difference within “time”, according to Duncan's multiple range post hoc test (p < 0.05); BL = baseline (3.48 h); T1 = intermediate time point (21.48 h); EP = endpoint (27.48 h); RRSO = raw red skinned onion; GRSO = grilled red skinned onion; FRSO = fried red skinned onion; and BC = blank control.a Normalized means.b Multivariate ANOVA model with category “time” on column and category “food matrix” on rows (p < 0.05). | |||
| RRSO | 0.821 ± 0.01bAB | 0.932 ± 0.05aA | <0.000001 |
| GRSO | 0.840 ± 0.07A | 0.857 ± 0.041AB | 0.431179 |
| FRSO | 0.829 ± 0.09A | 0.814 ± 0.011AB | 0.084396 |
| BC | 0.749 ± 0.014aB | 0.664 ± 0.291bB | 0.000047 |
| p (food matrix) | <0.000001 | 0.000002 | |
3.7. Correlations among phenolics and microbial taxa
Spearman rank correlations (p < 0.05), two-joining-way heatmaps, and Pearson cluster analysis were performed by comparison of two different normalized datasets, each derived from values of absolute quantification (microbes and phenolic compounds) (Fig. 2). According to the left-side dendrograms, two main clusters were acknowledged. Subsequently, the two key clusters were sorted in two different subgroups obtained by the dendrograms related to phenolics. In the resulting joint cluster 1A, the most significant positive correlations were held by Bacteroides and associated BPP group, by Firmicutes and associated Clostridium group IV and associated F. prausnitzii, with respect to five different phenolics. All these taxa shared significant correlations with two common variables, i.e. hydroxybenzoic acid isomer 1 and 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid. In cluster 1B, the same phenolics variables had negative correlations, but the overall significance was small. In particular, ATOP had significant negative correlations towards 4-hydroxy-3-methoxybenzoic acid, E. coli towards hydroxybenzoic acid isomer 1 and 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid, and Enterobacteriaceae towards the former two and 3-(3′,4′-dihydroxyphenyl)propanoic acid. Suggestions indicate that all these metabolites putatively produced by beneficial F. prausnitzii play a central role in the containment of opportunistic taxa, as in the RRSO samples. In this regard, our outputs are consistent with a recent result demonstrating the antibiofilm and antiproliferative action of hydroxybenzoic acids versus E. coli and other Enterobacteriaceae.84 Also, in the literature, it has been reported that species from Clostridium group IV can hydrolyze ester bonds, C-ring cleavage, and dehydroxylation of anthocyanins and proanthocyanidins, releasing hydroxybenzoic acids.75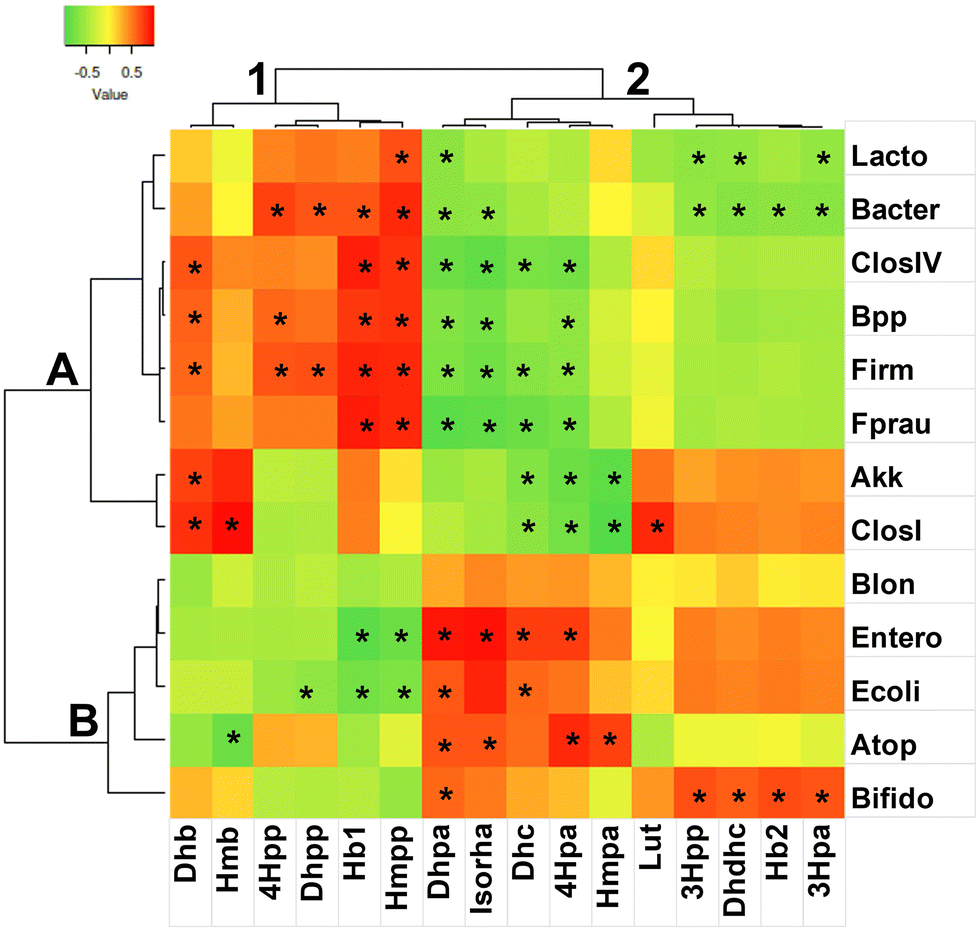 | ||
| Fig. 2 Two-way joining heatmap of Spearman-rank correlations. (A, B, 1 and 2) Cluster labels from Pearson dendrograms with centroid linkage. (*) Significant correlations (p > 0.05). X axis labels: Hb1 = hydroxybenzoic acid isomer 1; Hb2 = hydroxybenzoic acid isomer 2; 3Hpa = 3-(3′-hydroxyphenyl)acetic acid; 4Hpa = 3-(4′-hydroxyphenyl)acetic acid; Dhb = 3,4-dihydroxybenzoic acid; 3Hpp = 3-(3′-hydroxyphenyl)propanoic acid; 4Hpp = 3-(4′-hydroxyphenyl)propanoic acid; Hmb = 4-hydroxy-3-methoxybenzoic acid; Dhpa = 3-(3′,4′-dihydroxyphenyl)acetic acid; Dhc = 3′,4′-dihydroxycinnamic acid; Dhdhc = di-hydro-dihydroxycinnamic acid; Dhpp = 3-(3′,4′-dihydroxyphenyl)propanoic acid; Hmpa = 3-(4′-hydroxy-3′-methoxyphenyl)acetic acid; Hmpp = 3-(4′-hydroxy-3′-methoxyphenyl)propanoic acid; Lut = luteolin; Isorha = isorhamnetin. Y label; Lacto = Lactobacillales; Bacter = Bacteroidetes; ClosIV = Clostridium IV group; BPP = Bacteroides–Prevotella–Porphyromonas group; Firm = Firmicutes; Fprau = Faecalibacterium prausnitzii; Akk = Akkermansia muciniphila; ClosI = Clostridium group I; Blon = Bifidobacterium longum; Entero = Enterobacteriaceae; Ecoli = Escherichia coli; ATOP = Atopobium–Collinsella–Eggerthella group; Bifido = Bifidobacteriaceae. | ||
Cluster 2 accounted for a higher number of significant negative correlations compared to cluster 1. Indeed, subcluster 2A was defined by nine variables with significant negative correlations differently shared among seven bacterial variables. In particular, Firmicutes, Clos IV group and F. prausnitzii were negatively correlated with four common variables, namely, 3-(3′,4′-dihydroxyphenyl)acetic acid, isorhamnetin, 3′,4′-dihydroxycinnamic acid, and 3-(4′-hydroxyphenyl)acetic acid. Conversely, in subcluster 2B, the four former phenolics showed positive correlations with respect to the family Enterobacteriaceae, and two of these molecules (3-(3′,4′-dihydroxyphenyl)acetic acid and 3′,4′-dihydroxycinnamic acid) also with respect to E. coli. It is possible that 3′,4′-dihydroxycinnamic acid and 3-(4′-hydroxyphenyl)acetic acid produced or transformed by Enterobacteriaceae could be responsible for the depletion of A. muciniphila during the fermentation of the FRSO samples. Even if it is generally appreciated in the literature that most phenolics protect the gut epithelial barrier function and foster A. muciniphila and other beneficial microbes,85,86 it has also been reported that cinnamic compounds, e.g., cinnamaldehyde, produce side effects on commensals of the gut microbiota.87
4. Conclusions
Heat treatment increased the extractability of red-skinned onion phenolic compounds and their release during in vitro gastro-intestinal digestion. The grilled and fried red-skinned onions presented the highest amount of phenolic compounds following in vitro gastro-intestinal digestion and colonic fermentation. Additionally, specific beneficial taxa of the colon microbiota used in these experiments could transform and modify the chemical structure of onion phenolic compounds employing specific metabolic pathways, consequently gaining energy to multiply and foster in the colon niche. The degradation driven by the colon microbiota of RSO flavonols resulted in the accumulation of three main metabolites, i.e., 3-(3′-hydroxyphenyl)propanoic acid, 3-(3′-hydroxyphenyl)acetic acid and 3-(3′,4′-dihydroxyphenyl)acetic acid, with some qualitative and quantitative differences among the samples.Colonic fermentation of raw onions resulted in a substantial increase in beneficial taxa, which was larger compared to the heat-treated onions, particularly for Lactobacillales and beneficial clostridia. Also, a higher level of inhibition of opportunistic bacteria was seen for the raw onion samples, namely, the Clostridium perfringens group and E. coli. Eventually, the high flavonol content of the onion raw samples was found to be responsible for this effect, and also negatively seen for the depletion of Bifidobacteriaceae, which has been reported to be sensitive to quercetin, slowing metabolic pathways and cell wall synthesis.50
Therefore, consuming raw onions is preferable compared to grilled or fried onions in terms of amount of bioaccessible phenolic compounds and metabolites, which also have prebiotic activity fostering the growth of beneficial colonic bacteria and restraining the growth of opportunistic bacteria. Also, the cooked onions failed to maintain an eubiotic condition in the colon, triggering an increase in the F/B ratio to the level of a dysbiotic situation. The results reported in the present study showed that red-skinned onion, and especially raw, is an excellent dietary source of flavonols that are strongly metabolized by gut bacteria and can modulate the gut microbiota, promoting the growth of bacterial groups associated with a healthy status. However, more in vivo studies are necessary to validate the observed in vitro effect.
Institutional review board statement
The study was conducted according to the guidelines of the ethics procedures required at the University of Bologna.Informed consent statement
Informed consent has been obtained from the patient(s) to publish this paper.Data availability
Upon request to the corresponding author.Author contributions
Conceptualization, L. N., D. T., and A. G.; methodology, A. C., L. N., D. T., and A. G.; software, L. N., F. C., and A. C.; validation, L. N., D. T. and A. G.; formal analysis, A. C., F. C. and L. N.; investigation, A. C., L. N., F. C., D. T. and A. G.; resources, D. T., and A. G.; data curation, A. C., L. N., F. C., D. T., and A. G.; writing—original draft preparation, A. C., L. N., F. C., D. T., and A. G.; writing—review and editing, A. C., L. N., F. C., D. T., and A. G.; visualization A. C., L. N., F. C.; supervision, L. N., D. T., and A. G.; project administration, D. T. and A. G.; funding acquisition, D. T. and A. G. All authors have read and agreed to the published version of the manuscript.Conflicts of interest
The authors declare no conflicts of interest.Acknowledgements
The authors acknowledge the Fondazione Cassa di Risparmio di Modena for funding the HPLC-ESI-Q Exactive Hybrid Quadrupole-Orbitrap Mass Spectrometer System at the Centro Interdipartimentale Grandi Strumenti (CIGS). The authors also acknowledge the personnel of Centre for Applied Biomedical Research – CRBA, Alma Mater Studiorum – Università di Bologna, Policlinico di Sant'Orsola, Bologna, Italy.References
- A. Cattivelli, A. Conte, S. Martini and D. Tagliazucchi, Influence of cooking methods on onion phenolic compounds bioaccessibility, Foods, 2021, 10, 1023 CrossRef CAS PubMed.
- A. Cattivelli, A. Di Lorenzo, A. Conte, S. Martini and D. Tagliazucchi, Red-skinned onion phenolic compounds stability and bioaccessibility: A comparative study between deep-frying and air-frying, J. Food Compos. Anal., 2023, 115, 105024 CrossRef CAS.
- D. Kothari, W. D. Lee and S. K. Kim, Allium flavonols: Health benefits, molecular targets, and bioavailability, Antioxidants, 2020, 9, 888 CrossRef CAS PubMed.
- Z. Bahadoran, P. Mirmiran, A. A. Momenan and F. Azizi, Allium vegetable intakes and the incidence of cardiovascular disease, hypertension, chronic kidney disease, and type 2 diabetes in adults: a longitudinal follow-up study, J. Hypertens., 2017, 35(9), 1909–1916 CrossRef CAS PubMed.
- Q. Wan, N. Li, L. Du, R. Zhao, M. Yi, Q. Xu and Y. Zhou, Allium vegetable consumption and health: An umbrella review of meta-analyses of multiple health outcomes, Food Sci. Nutr., 2019, 7(8), 2451–2460 CrossRef CAS PubMed.
- S. Martini, A. Conte, A. Cattivelli and D. Tagliazucchi, Domestic cooking methods affect the stability and bioaccessibility of dark purple eggplant (Solanum melongena) phenolic compounds, Food Chem., 2021, 341, 128298 CrossRef CAS PubMed.
- A. Cattivelli, A. Conte, S. Martini and D. Tagliazucchi, Cooking and in vitro digestion modulate the anti-diabetic properties of red-skinned onion and dark purple eggplant phenolic compounds, Foods, 2022, 11, 689 CrossRef CAS PubMed.
- L. Nissen, F. Casciano, E. Chiarello, M. Di Nunzio, A. Bordoni and A. Gianotti, Colonic in vitro model assessment of the prebiotic potential of bread fortified with polyphenols rich olive fiber, Nutrients, 2021, 13, 787 CrossRef CAS PubMed.
- L. Nissen, M. C. Valerii, E. Spisni, F. Casciano and A. Gianotti, Multiunit in vitro colon model for the evaluation of prebiotic potential of a fiber plus D-limonene food supplement, Foods, 2021, 10, 2371 CrossRef CAS PubMed.
- L. Nissen, A. Cattivelli, F. Casciano, A. Gianotti and D. Tagliazucchi, Roasting and frying modulate the phenolic profile of dark purple eggplant and differently change the colon microbiota and phenolic metabolites after in vitro digestion and fermentation in a gut model, Food Res. Int., 2022, 160, 111702 CrossRef CAS PubMed.
- E. De Santiago, C. I. R. Gill, I. Carafa, K. M. Tuohy, M. P. De Peña and C. Cid, Digestion and colonic fermentation of raw and cooked Opuntia ficus-indica cladodes impacts bioaccessibility and bioactivity, J. Agric. Food Chem., 2019, 67, 2490–2499 CrossRef CAS PubMed.
- I. Juániz, I. A. Ludwig, L. Bresciani, M. Dall'Asta, P. Mena, D. Del Rio, C. Cid and M. P. De Peña, Catabolism of raw and cooked green pepper (Capsicum annuum) (poly)phenolic compounds after simulated gastrointestinal digestion and faecal fermentation, J. Funct. Foods, 2016, 27, 201–213 CrossRef.
- I. Juániz, I. A. Ludwig, L. Bresciani, M. Dall'Asta, P. Mena, D. Del Rio, C. Cid and M. P. De Peña, Bioaccessibility of (poly)phenolic compounds of raw and cooked cardoon (Cynara cardunculus L.) after simulated gastrointestinal digestion and fermentation by human colonic microbiota, J. Funct. Foods, 2017, 32, 195–207 CrossRef.
- J. C. Espín, A. González-Sarrías and F. A. Tomás-Barberán, The gut microbiota: A key factor in the therapeutic effects of (poly)phenols, Biochem. Pharmacol., 2017, 139, 82–93 CrossRef PubMed.
- D. Carregosa, C. Pinto, M. Á. Ávila-Gálvez, P. Bastos, D. Berry and C. N. dos Santos, A look beyond dietary (poly)phenols: The low molecular weight phenolic metabolites and their concentrations in human circulation, Compr. Rev. Food Sci. Food Saf., 2022, 21, 3931–3962 CrossRef CAS PubMed.
- M. C. Rodríguez-Daza, E. C. Pulido-Mateos, J. Lupien-Meilleur, D. Guyonnet, Y. Desjardins and D. Roy, Polyphenol-Mediated Gut Microbiota Modulation: Toward Prebiotics and Further, Front. Nutr., 2021, 8, 689456 CrossRef PubMed.
- T. Corrêa, M. M. Rogero, N. Hassimotto and F. M. Lajolo, The two-way polyphenols-microbiota interactions and their effects on obesity and related metabolic diseases, Front. Nutr., 2019, 6, 188 CrossRef PubMed.
- K. Kasprzak-Drozd, T. Oniszczuk, M. Stasiak and A. Oniszczuk, Beneficial effects of phenolic compounds on gut microbiota and metabolic syndrome, Int. J. Mol. Sci., 2021, 22, 3715 CrossRef CAS PubMed.
- P. Mena and L. Bresciani, Dietary fibre modifies gut microbiota: what's the role of (poly)phenols?, Int. J. Food Sci. Nutr., 2020, 71, 783–784 CrossRef CAS PubMed.
- M. Koudoufio, Y. Desjardins, F. Feldam, S. Spahis, E. Delvin and E. Levy, Insight into polyphenol and gut microbiota crosstalk: are their metabolites the key to understand protective effects against metabolic disorders?, Antioxidants, 2020, 9, 982 CrossRef CAS PubMed.
- L. Nissen, F. Casciano and A. Gianotti, Intestinal fermentation in vitro models to study food-induced gut microbiota shift: An updated review, FEMS Microbiol. Lett., 2020, 367, fnaa097 CrossRef CAS PubMed.
- M. Minekus, M. Alminger, P. Alvito, S. Ballance, T. Bohn and C. Bourlieu, et al., A standardised static in vitro digestion method suitable for food – An international consensus, Food Funct., 2014, 5, 1113–1124 RSC.
- M. L. Connolly, K. M. Tuohy and J. A. Lovegrove, Wholegrain oat-based cereals have prebiotic potential and low glycaemic index, Br. J. Nutr., 2012, 108, 2198–2206 CrossRef CAS PubMed.
- S. Oba, T. Sunagawa, R. Tanihiro, K. Awashima, H. Sugiyama and T. Odani, et al., Prebiotic effects of yeast mannan, which selectively promotes Bacteroides thetaiotaomicron and Bacteroides ovatus in a human colonic microbiota model, Sci. Rep., 2020, 10(1), 1–11 CrossRef PubMed.
- M. E. Arnal, S. Denis, O. Uriot, C. Lambert, S. Holowacz and F. Paul, et al., Impact of oral galenic formulations of Lactobacillus salivarius on probiotic survival and interactions with microbiota in human in vitro gut models, Benefic. Microbes, 2021, 12, 381–396 CrossRef CAS PubMed.
- L. Nissen, C. Aniballi, F. Casciano, A. Elmi, D. Ventrella, A. Zannoni, A. Gianotti and M. L. Bacci, Maternal amoxicillin affects piglets colon microbiota: microbial ecology and metabolomics in a gut model, Appl. Microbiol. Biotechnol., 2022, 106(22), 7595–7614 CrossRef CAS PubMed.
- F. Casciano, L. Nissen, E. Chiarello, M. Di Nunzio, A. Bordoni and A. Gianotti, In vitro assessment of the effect of lactose-free milk on colon microbiota of lactose intolerant adults, Int. J. Food Sci. Technol., 2022 DOI:10.1111/ijfs.16253.
- X. Wang, G. R. Gibson, M. Sailer, S. Theis and R. A. Rastall, Prebiotics inhibit proteolysis by gut bacteria in a host diet-dependent manner: A three-stage continuous in vitro gut model experiment, Appl. Environ. Microbiol., 2020, 86, e02730–e02719 CAS.
- S. Martini, D. Tagliazucchi, G. Minelli and D. P. Lo Fiego, Influence of linseed and antioxidant-rich diets in pig nutrition on lipid oxidation during cooking and in vitro digestion of pork, Food Res. Int., 2020, 137, 109528 CrossRef CAS PubMed.
- C. D. Kay, M. N. Clifford, P. Mena, G. J. McDougall, C. Andres-Lacueva, A. Cassidy, D. Del Rio and N. Kuhnert, et al., Recommendations for standardizing nomenclature for dietary (poly)phenol catabolites, Am. J. Clin. Nutr., 2020, 112, 1051–1068 CrossRef PubMed.
- S. A. Tanner, A. Zihler Berner, E. Rigozzi, F. Grattepanche, C. Chassard and C. Lacroix, In vitro continuous fermentation model (PolyFermS) of the swine proximal colon for simultaneous testing on the same gut microbiota, PLoS One, 2014, 9(4), e94123 CrossRef PubMed.
- S. Westfall, N. Lomis and S. A. Prakash, Polyphenol-rich prebiotic in combination with a novel probiotic formulation alleviates markers of obesity and diabetes in Drosophila, J. Funct. Foods, 2018, 48, 374–386 CrossRef CAS.
- I. Tsitko, F. Wiik-Miettinen, O. Mattila, N. Rosa-Sibakov, T. Seppänen-Laakso and J. Maukonen, et al., A small in vitro fermentation model for screening the gut microbiota effects of different fiber preparations, Int. J. Mol. Sci., 2019, 20, 1925 CrossRef CAS PubMed.
- A. Tamargo, C. Cueva, M. Silva, N. Molinero, B. Miralles, B. Bartolomé and M. V. Moreno-Arribas, Gastrointestinal co-digestion of wine polyphenols with glucose/whey proteins affects their bioaccessibility and impact on colonic microbiota, Food Res. Int., 2022, 111010 CrossRef CAS PubMed.
- M. Modesto, I. Stefanini, M. R. D'Aimmo, L. Nissen, D. Tabanelli, M. Mazzoni, P. Bosi, G. P. Strozzi and B. Biavati, Strategies to augment non-immune system based defence mechanisms against gastrointestinal diseases in pigs, NJAS – Wagening. J. Life Sci., 2011, 58, 149–156 CrossRef.
- M. I. Love, W. Huber and S. Anders, Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2, Genome Biol., 2014, 15, 1–21 CrossRef PubMed.
- S. Babicki, D. Arndt, A. Marcu, Y. Liang, J. R. Grant, A. Maciejewski and D. S. Wishart, Heatmapper: web-enabled heat mapping for all, Nucleic Acids Res., 2016, 44, W147–W153 CrossRef CAS PubMed.
- M. Marotti and R. Piccaglia, Characterization of flavonoids in different cultivars of onion (Allium cepa L.), J. Food Sci., 2002, 67, 1229–1232 CrossRef CAS.
- I. Juániz, I. A. Ludwig, E. Huarte, G. Pereira-Caro, J. M. Moreno-Rojas, C. Cid and M. P. De Peña, Influence of heat treatment on antioxidant capacity and (poly)phenolic compounds of selected vegetables, Food Chem., 2016, 197, 466–473 CrossRef PubMed.
- S. Harris, N. Brunton, U. Tiwari and E. Cummins, Human exposure modelling of quercetin in onions (Allium cepa L.) following thermal processing, Food Chem., 2015, 187, 135–139 CrossRef CAS PubMed.
- N. Ortega, J. Reguant, M. P. Romero, A. Macià and M. J. Motilva, Effect of fat content on the digestibility and bioaccessibility of cocoa polyphenol by an in vitro digestion model, J. Agric. Food Chem., 2009, 57, 5743–5749 CrossRef CAS PubMed.
- J. Fuentes, E. Atala, E. Pastene, C. Carrasco-Pozo and H. Speisky, Quercetin oxidation paradoxically enhances its antioxidant and cytoprotective properties, J. Agric. Food Chem., 2017, 65, 11002–11010 CrossRef CAS PubMed.
- M. Rogozinska and M. Biesaga, Decomposition of flavonols in the presence of saliva, Appl. Sci., 2020, 10, 7511 CrossRef CAS.
- D. Del Rio, A. Rodriguez-Mateos, J. P. Spencer, M. Tognolini, G. Borges and A. Crozier, Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases, Antioxid. Redox Signaling, 2013, 18(14), 1818–1892 CrossRef CAS PubMed.
- A. F. Almeida, G. I. A. Borge, M. Piskula, A. Tudose, L. Tudoreanu, K. Valentová, G. Williamson and C. N. Santos, Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation, Compr. Rev. Food Sci. Food Saf., 2018, 17, 714–731 CrossRef CAS PubMed.
- M. R. Olthof, P. C. Hollman, T. B. Vree and M. B. Katan, Bioavailabilities of quercetin-3-glucoside and quercetin-4′-glucoside do not differ in humans, J. Nutr., 2000, 130(5), 1200–1203 CrossRef CAS PubMed.
- A. J. Day, F. J. Canada, J. C. Diaz, P. A. Kroon, W. R. McLauchlan and C. B. Faulds, et al., Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase, FEBS Lett., 2000, 468, 166–170 CrossRef CAS PubMed.
- P. C. H. Hollman, J. H. M. Devries, S. D. Vanleeuwen, M. J. B. Mengelers and M. B. Katan, Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers, Am. J. Clin. Nutr., 1995, 62(6), 1276–1282 CrossRef CAS PubMed.
- A. Serra, A. Macia, M. P. Romero, J. Reguant, N. Ortega and M. J. Motilva, Metabolic pathways of the colonic metabolism of flavonoids (flavonols, flavones and flavanones) and phenolic acids, Food Chem., 2012, 130(2), 383–393 CrossRef CAS.
- I. Fernández-Jalao, C. Balderas, M. V. Calvo, J. Fontecha, C. Sánchez-Moreno and B. De Ancos, Impact of High-Pressure Processed Onion on Colonic Metabolism Using a Dynamic Gastrointestinal Digestion Simulator, Metabolites, 2021, 11, 262 CrossRef PubMed.
- E. Hein, K. Rose, G. Van't Slot, A. W. Friedrich and H. Humpf, Deconjugation and degradation of flavonol glycosides by pig cecal microbiota characterized by fluorescence in situ hybridization (FISH), J. Agric. Food Chem., 2008, 56(6), 2281–2290 CrossRef CAS PubMed.
- A. R. Rechner, M. A. Smith, G. Kuhnle, G. R. Gibson, E. S. Debnam, S. K. Srai, P. Moore and C. A. Rice-Evans, Colonic metabolism of dietary polyphenols: Influence of structure on microbial fermentation products, Free Radical Biol. Med., 2004, 36(2), 212–220 CrossRef CAS PubMed.
- A. Cortés-Martín, M. V. Selma, F. Tomás-Barberán, A. González-Sarrías and J. C. Espín, Where to look into the puzzle of polyphenols and health? The postbiotics and gut microbiota associated with human metabotypes, Mol. Nutr. Food Res., 2020, 64, e1900952 CrossRef PubMed.
- S. H. Duncan, W. R. Russel, A. Quartieri, M. Rossi, J. Parkhill, A. W. Walker and H. J. Flint, Wheat bran promotes enrichment within the human colonic microbiota of butyrate-producing bacteria that release ferulic acid, Environ. Microbiol., 2016, 18, 2214–2225 CrossRef CAS PubMed.
- W. R. Russell, L. Scobbie, A. Chesson, A. J. Richardson, C. S. Stewart, S. H. Duncan, J. E. Drew and G. G. Duthie, Anti-inflammatory implications of the microbial transformation of dietary phenolic compounds, Nutr. Cancer, 2008, 60, 636–642 CrossRef CAS PubMed.
- E. Fernández-Millán, S. Ramos, C. Alvarez, L. Bravo, L. Goya and M. A. Martín, Microbial phenolic metabolites improve glucose-stimulated insulin secretion and protect pancreatic beta cells against tert-butyl hydroperoxide-induced toxicity via ERKs and PKC pathways, Food Chem. Toxicol., 2014, 66, 245–253 CrossRef PubMed.
- M. Larrosa, C. Luceri, E. Vivoli, C. Pagliuca, M. Lodovici, G. Moneti and P. Dolara, Polyphenol metabolites from colonic microbiota exert anti-inflammatory activity on different inflammation models, Mol. Nutr. Food Res., 2009, 53, 1044–1054 CrossRef CAS PubMed.
- I. Najmanová, J. Pourová, M. Vopršalová, V. Pilařová, V. Semecký, L. Nováková and P. Mladěnka, Flavonoid metabolite 3-(3-hydroxyphenyl)propionic acid formed by human microflora decreases arterial blood pressure in rats, Mol. Nutr. Food Res., 2016, 60, 981–991 CrossRef PubMed.
- P. Dias, J. Pourová, M. Vopršalová, I. Nejmanová and P. Mladěnka, 3-Hydroxyphenylacetic acid: a blood pressure-reducing flavonoid metabolite, Nutrients, 2022, 14, 328 CrossRef CAS PubMed.
- K. Gao, A. Xu, C. Krul, K. Venema, Y. Liu, Y. Niu, J. Lu, L. Bensoussan, N. P. Seeram, D. Heber and S. M. Henning, Of the major phenolic acids formed during human microbial fermentation of tea, citrus, and soy flavonoid supplements, only 3,4-dihydroxyphenylacetic acid has antiproliferative activity, J. Nutr., 2006, 136, 52–57 CrossRef CAS PubMed.
- D. Carregosa, R. Carecho, I. Figueira and C. N. Santos, Low-molecular weight metabolites from polyphenols as effectors for attenuating neuroinflammation, J. Agric. Food Chem., 2020, 68, 1790–1807 CrossRef CAS PubMed.
- A. Nakajima, T. Sasaki, K. Itoh, T. Kitahara, Y. Takema and K. Hiramatsu, et al., A soluble fiber diet increases Bacteroides fragilis group abundance and immunoglobulin A production in the gut, Appl. Environ. Microbiol., 2020, 86(13), e00405–e00420 CrossRef CAS PubMed.
- M. Blaut, M. D. Collins, G. W. Welling, J. Doré, J. van Loo and W. de Vos, Molecular biological methods for studying the gut microbiota: the EU human gut flora project, Br. J. Nutr., 2002, 87, S203–S211 CrossRef CAS PubMed.
- A. Koliada, G. Syzenko, V. Moseiko, L. Budovska, K. Puchkov and V. Perederiy, et al., Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population, BMC Microbiol., 2017, 17, 1–6 CrossRef PubMed.
- S. Zhou, Y. Wang, J. J. Jacoby, Y. Jiang, Y. Zhang and L. L. Yu, Effects of medium-and long-chain triacylglycerols on lipid metabolism and gut microbiota composition in C57BL/6J mice, J. Agric. Food Chem., 2017, 65, 6599–6607 CrossRef CAS PubMed.
- J. E. Martinez, D. D. Kahana, S. Ghuman, H. P. Wilson, J. Wilson and S. C. Kim, et al., Unhealthy lifestyle and gut dysbiosis: A better understanding of the effects of poor diet and nicotine on the intestinal microbiome, Front. Endocrinol., 2021, 12, 649 Search PubMed.
- M. E. Sanders, D. J. Merenstein and G. Reid, et al., Probiotics and prebiotics in intestinal health and disease: from biology to the clinic, Nat. Rev. Gastroenterol. Hepatol., 2019, 16, 605–616 CrossRef PubMed.
- L. Liu, C. Zhang, H. Zhang, G. Qu, C. Li and L. Liu, Biotransformation of polyphenols in apple pomace fermented by β-glucosidase-producing Lactobacillus rhamnosus, L08, Foods, 2021, 10, 1343 CrossRef CAS PubMed.
- M. Boto-Ordóñez, M. Urpi-Sarda, M. I. Queipo-Ortuño, S. Tulipani, F. J. Tinahones and C. Andres-Lacueva, High levels of Bifidobacteria are associated with increased levels of anthocyanin microbial metabolites: A randomized clinical trial, Food Funct., 2014, 5, 1932–1938 RSC.
- Y. Iqbal, J. J. Cottrell, H. A. Suleria and F. R. Dunshea, Gut microbiota-polyphenol interactions in chicken: A review, Animals, 2020, 10, 1391 CrossRef PubMed.
- R. A. Kemperman, G. Gross, S. Mondot, S. Possemiers, M. Marzorati, T. Van de Wiele, J. Doré and E. E. Vaughan, Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome, Food Res. Int., 2013, 53, 659–669 CrossRef CAS.
- J. Firrman, L. Liu, L. Zhang, G. A. Argoty, M. Wang and P. Tomasula, et al., The effect of quercetin on genetic expression of the commensal gut microbes Bifidobacterium catenulatum, Enterococcus caccae and Ruminococcus gauvreauii, Anaerobes, 2016, 42, 130–141 CrossRef CAS PubMed.
- S. Khubber, F. J. Marti-Quijal, I. Tomasevic, F. Remize and F. J. Barba, Lactic acid fermentation as a useful strategy to recover antimicrobial and antioxidant compounds from food and by-products, Curr. Opin. Food Sci., 2022, 43, 189–198 CrossRef CAS.
- P. Filannino, R. Di Cagno and M. Gobbetti, Metabolic and functional paths of lactic acid bacteria in plant foods: get out of the labyrinth, Curr. Opin. Biotechnol., 2018, 49, 64–72 CrossRef CAS PubMed.
- Y. Date, Y. Nakanishi, S. Fukuda, Y. Nuijima, T. Kato, M. Umehara, H. Ohno and J. Kikuchi, In vitro evaluation method for screening of candidate prebiotic foods, Food Chem., 2014, 152, 251–260 CrossRef CAS PubMed.
- A. Braune and M. Blaut, Bacterial species involved in the conversion of dietary flavonoids in the human gut, Gut Microbes, 2016, 7, 216–234 CrossRef CAS PubMed.
- M. Zhang, L. Zhou, Y. Wang, R. G. Dorfman, D. Tang, L. Xu and Y. Pan, et al., Faecalibacterium prausnitzii produces butyrate to decrease c-Myc-related metabolism and Th17 differentiation by inhibiting histone deacetylase 3, Int. Immunol., 2019, 31, 499–514 CrossRef CAS PubMed.
- I. Moreno-Indias, L. Sanchez-Alcoholado, P. Pérez-Martínez, C. Andres-Lacueva, F. Cardona, F. J. Tinahones and M. I. Q. Ortuño, Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients, Food Funct., 2016, 7, 1775–1787 RSC.
- M. Wiciński, J. Gębalski, E. Mazurek, M. Podhorecka, M. Śniegocki, P. Szychta, E. Sawicka and B. Malinowski, The influence of polyphenol compounds on human gastrointestinal tract microbiota, Nutrients, 2020, 12, 350 CrossRef PubMed.
- O. Patrascu, F. Béguet-Crespel, L. Marinelli, E. Le Chatelier, A. L. Abraham, M. Leclerc and C. Klopp, et al., A fibrolytic potential in the human ileum mucosal microbiota revealed by functional metagenomic, Sci. Rep., 2017, 7, 40248 CrossRef CAS PubMed.
- N. Jiménez, L. Santamaría, M. Esteban-Torres, B. de las Rivas and R. Muñoz, Contribution of a tannase from Atopobium parvulum DSM 20469T in the oral processing of food tannins, Food Res. Int., 2014, 62, 397–402 CrossRef.
- G. Williamson and M. N. Clifford, Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols, Biochem. Pharmacol., 2017, 139, 24–39 CrossRef CAS PubMed.
- G. R. Gibson, R. Hutkins, M. E. Sanders, S. L. Prescott, R. A. Reimer and S. J. Salminen, et al., Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics, Nat. Rev. Gastroenterol. Hepatol., 2017, 14, 491–502 CrossRef PubMed.
- A. N. Tamfu, O. Ceylan, G. Cârâc, E. Talla and R. M. Dinica, Antibiofilm and anti-quorum sensing potential of cycloartane-type triterpene acids from cameroonian grassland propolis: phenolic profile and antioxidant activity of crude extract, Molecules, 2022, 27(15), 4872 CrossRef CAS PubMed.
- F. F. Anhê, G. Pilon, D. Roy, Y. Desjardins, E. Levy and A. Marette, Triggering Akkermansia with dietary polyphenols: A new weapon to combat the metabolic syndrome?, Gut Microbes, 2016, 7(2), 146–153 CrossRef PubMed.
- X. Wang, Y. Qi and H. Zheng, Dietary Polyphenol, Gut Microbiota, and Health Benefits, Antioxidants, 2022, 11(6), 1212 CrossRef CAS PubMed.
- D. P. Singh, P. Khare, V. Bijalwan, R. K. Baboota, J. Singh and K. K. Kondepudi, et al., Coadministration of isomalto–oligosaccharides augments metabolic health benefits of cinnamaldehyde in high fat diet fed mice, BioFactors, 2017, 43(6), 821–835 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3fo00085k |
| ‡ These authors contributed equally. |
| This journal is © The Royal Society of Chemistry 2023 |