
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Total biosynthesis of fungal tetraketide pyrones†
Yunlong
Sun‡
 a,
Dongsong
Tian‡
a,
Eric
Kuhnert
a,
Geraldine
Le Goff
b,
Guillaume
Arcile
b,
Jamal
Ouazzani
b and
Russell J.
Cox
*a
a,
Dongsong
Tian‡
a,
Eric
Kuhnert
a,
Geraldine
Le Goff
b,
Guillaume
Arcile
b,
Jamal
Ouazzani
b and
Russell J.
Cox
*a
aInstitute for Organic Chemistry, Leibniz Universität Hannover, Hannover, Germany. E-mail: russell.cox@oci.uni-hannover.de
bICSN, CNRS, Gif Sur Yvette, France
First published on 23rd October 2023
Abstract
Fungal tetraketide pyrones possess important and potent bioactivities, but their detailed biosynthetic pathways are unknown and synthetic routes to their production are lengthy. Here we investigated the fungal pathways to the multiforisins and compounds related to islandic acid. Heterologous expression experiments yield high titres of these compounds and pathway intermediates. The results both elucidate the pathway and offer a platform for the total biosynthesis of this class of metabolites.
The tetraketide pyrones form a large class of bioactive fungal specialized metabolites (SMs) that feature a wide variety of oxygenation and acylation patterns. For example, multiforisin A 1A and related compounds (Fig. 1) show significant in vitro immunosuppressive activity only 10-fold less potent than the standard drug cyclosporin A.1 Related compounds such as islandic acid 2 from Penicillium islandicum possesses potent cytotoxicity vs. Yoshida sarcoma (IC50 1 μg ml−1 for the methyl ester).2,3 Similarly, allantopyrone A 3 is also a potent cytotoxin (IC50 0.32 μM) via inhibition of the NF-κB signaling pathway; and rosellisin 4 has antibacterial properties (IC50 30 ppm vs. S. aureus).4 Rosellisin 4 is known to be a polyketide derived from four acetate units5 and it seems likely that 1A, 2 and 3 arise from similar biosynthetic pathways. Production of islandic acid 2 and related compounds has been reported by multistep total chemical synthesis,6 but such methods are inefficient and production of these compounds by total biosynthesis would be advantageous. However, despite their potent and diverse bioactivities, and the presence of an intriguing EZ diene in 2 and 3, this class of bioactive tetraketide pyrones has not been deeply investigated at the biogenetic level.
 | ||
| Fig. 1 Typical Fungal tetraketide Pyrones. | ||
Fermentation of Hypomontagnella monticulosa during a series of experiments into the biosynthesis of the sporochartines,7 revealed that this organism can produce the pyrone tetraketide multiforisin H 1H.1 We screened the genome of H. monticulosa and two closely related species with the aim of discovering a biosynthetic gene cluster (BGC) which might be responsible for this pathway. We focussed on a BGC (mfn) encoding two highly-reducing polyketide synthases (hrPKS) mfnPKS1 and mfnPKS2, together with an O-acyl transferase (O-AcT, mfnL3), two cytochrome P450 monooxygenases (mfnL2 and mfnR3), an O-methyl transferase (O-MeT, mfnL1), a transcription factor (TF, mfnR1), an FAD-dependent oxidoreductase (FMO, mfnR2), and a short-chain dehydrogenase/reductase (SDR, mfnR4, Table 1).
|
|
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mfnPKS2 | mfnL3 | mfnL2 | mfnL1 | mfnPKS1 | mfnR2 | mfnR3 | mfnR4 | ilaPKS2 | ilaR2 | ilaR8 | Products | |
| Exp. | hrPKS | O-AcT | P450 | O-MeT | hrPKS | FMO | P450 | SDR | hrPKS | O-AcT | SDR | |
| a Same as expt 11, with acid workup. | ||||||||||||
| 1 | ✓ | 5 | ||||||||||
| 2 | ✓ | ✓ | 5, 6 | |||||||||
| 3 | ✓ | ✓ | 5, 6 | |||||||||
| 4 | ✓ | ✓ | 5, 7, 8 | |||||||||
| 5 | ✓ | ✓ | ✓ | 5, 9 | ||||||||
| 6 | ✓ | ✓ | ✓ | 1I, 5, 9, 10 | ||||||||
| 7 | ✓ | ✓ | ✓ | ✓ | 1I, 5, 9 | |||||||
| 8 | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 5, 9 | ||||||
| 9 | ✓ | ✓ | ✓ | ✓ | ✓ | 9, 11, 16 | ||||||
| 10 | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 5, 9, | ||||||
| 11 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 1H, 9, 12 | |||||
| 11a | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 1H, 13, 14, 15 | |||||
| 12 | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 5, 9 | ||||||
| 13 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 1H, 5, 9, 12 | |||
| 14 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1I, 5, 9 | ||||

The boundaries of this BGC were suggested by transcriptomic studies in H. monticulosa which showed this region to be transcriptionally co-regulated (see ESI,† Fig S1.1). Near identical BGCs were found in the related species H. spongiphila and H. submonticulosa (ESI,† Fig S1.2). Previous work had shown that selective gene knockout in H. monticulosa is inefficient.7 We selected to use heterologous expression in the workhorse fungal host Aspergillus oryzae to probe the biosynthesis and provide a total biosynthesis platform.8,9
An intron-free construct of mfnPKS1 was cloned into an A. oryzae expression vector. Transformation of A. oryzae yielded eight transformants, and PCR analysis confirmed six of these to contain full-length mfnPKS1. These transformants produced a new compound 5 in high titre (123 mg L−1, Fig. 2) and traces of 6 (Fig S1.16, ESI†) that were not produced by untransformed A. oryzae. Larger scale fermentation (1L) allowed purification and structure elucidation of 5 that was identified by full NMR analysis as amphicopyrone A 5.10
 | ||
| Fig. 2 LCMS results for experiments 1–11a. Diode array detector (200–600 nm) data shown. Absorbtion units are arbitrary. | ||
Next, we undertook a systematic program of gene expression of the mfn genes (Table 1, experiments 2-11, see ESI† for vector details) followed by LCMS analysis (Fig. 2). Addition of P450 monooxygenase encoding genes mfnL2 (exp. 2) or mfnR3 (exp. 3) produced small but isolatable amounts of the 8-alcohol 6 (see ESI† for full NMR characterisation of all isolated compounds) with the majority of 5 unreacted. Alcohol 6 is not on the pathway to the multiforisins, and we assumed it to be an A. oryzae shunt.
In contrast, coexpression of mfnL1 that encodes an O-MeT (exp. 4) substantially converted 5 to known methyl ether 7 (140 mg L−1) that does appear to be a pathway intermediate.10 Minor amounts of presumed shunt 8-carboxylate 8 were also observed. Compound 7 is fully converted to the known C-9 alcohol 9 (134 mg L−1) when the mfnL2 P450 monooxygenase is added (exp. 5). On the other hand, 7 is converted to 10 (majority, 31 mg L−1), 9, and some diol 1I (minority) by mfnR3 that encodes the other P450 monooxygenase (exp. 6). Expression of both P450 monooxygenases at this stage (exp. 7) leads to the expected diol multiforisin I 1I (36 mg L−1). At this point, expression of mfnR2 that encodes the FMO (exp. 8) did not appear to give any further new compounds. However, coexpression of mfnR4 that encodes the SDR, oxidises intermediate 9 to the aldehyde 11 (exp. 9, 10 mg L−1). Addition of the mfnL3-encoded O-AcT alone at this juncture gave no new compounds (exp. 10), but coexpression of mfnL3 with mfnPKS2 led to direct synthesis of multiforisin H 1H (exp. 11, 28 mg L−1). Finally, a series of thioether shunts (13–16) were isolated after acidification of the media during extraction (exp. 11a). These compounds likely arise by formation of quinomethide-type structures that are quenched by 2-hydroxyl-3-thiopropionate or 2-thioacetate.
We next searched the publically available genome of Penicillium islandicum, the producing organism of islandic acid 2, using the mfn genes as bait. This revealed a near identical BGC (ila, Table 1) encoding homologs of: mfnPKS1 (ilaPKS1), two parallel cytochrome P450s (ilaR4 & ilaR6); the O-methyltransferase (ilaR1); the transcriptional regulator (ilaR3), the SDR (ilaR7), and FMO (ilaR5). However the second PKS (ilaPKS2) and the acyl transferase (ilaR2) do not show significant similarity with the cognate proteins encoded by the mfn BGC, consistent with the formation and attachment of different PKS-derived acyl groups. Additionally the ila BGC encodes a second SDR (ilaR8) that shows low similarity to mfnR4. We hypothesised that IlaPKS2 from the proposed P. islandicum BGC might synthesise the unusual EZ triketide which may be transferred to the C-2 hydroxymethyl group by the ilaAT in analogy to the multiforisin chemistry.
In the H. monticulosa case it appears that the role of PKS2 is simply to supply an acetyl group for attachment at the 10-OH. This is consistent with the observed mutation of the active site cysteine of the MfnPKS2 KS domain to serine (ESI,† Section 1.1.5): presumably the PKS can load an acetyl starter unit but not extend it. Tang and coworkers recently described another Type I PKS component with similar activity.11
We next attempted to synthesise 2 or 3 by expressing ilaPKS2 in A. oryzae that produces multiforisin pathway intermediates. Three experiments were set up. In the first (exp. 12), A. oryzae from expt. 7 that produces 1I, 5, and 9 was cotransformed with ilaPKS2 alone. Second (exp. 13), A. oryzae from exp. 11 that produces 1I, 1H, 9, and 12 was cotransformed with ilaPKS2 and ilaR2 that encodes a putative acyl transferase. Finally (exp. 14), A. oryzae from exp. 7 that produces 1I, 5, 9 and was cotransformed with ilaPKS2, ilaR2 and ilaR7 that encodes an SDR. However, none of these transformants produced any compounds in addition to the those already produced by the A. oryzae hosts.
Yuan and coworkers recently reported a fungal BGC (amp) from Amphichorda felina that also encodes the biosynthesis of tetraketide pyrones. In comparison to the mfn and ile BGCs, the genes encoding PKS1, O-MeT and the two P450 monooxygenases are conserved. However, in their work only expression of the first catalysts from the pathway led to the production of metabolites (5 and 7). In our hands the pathway expression is more productive, showing that hydroxylation of 7 can occur in parallel, to form either or both of 9 and 10 as intermediates, before formation of multiforisin I 1I (Scheme 1).
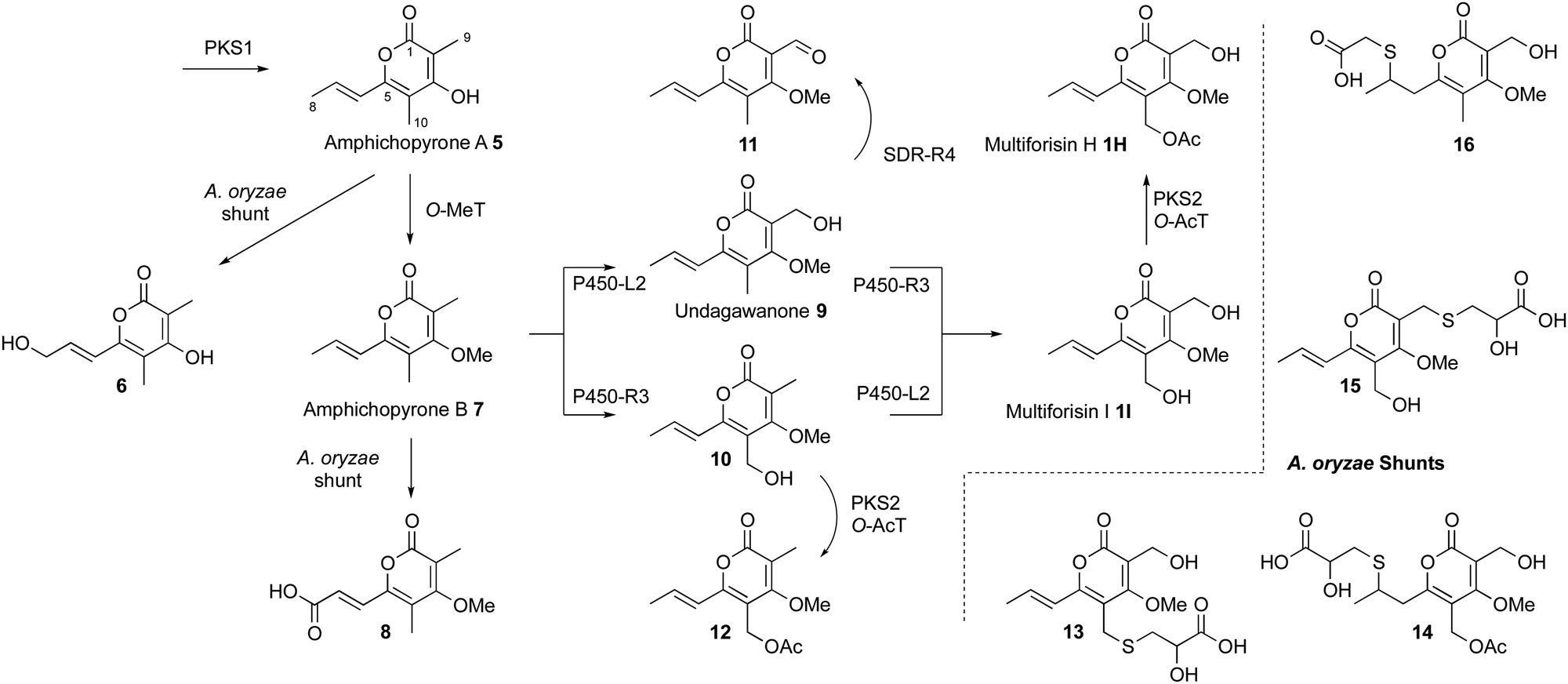 | ||
| Scheme 1 Deduced pathway for multiforisin H biosynthesis in H. spongiphila showing shunts observed in A. oryzae. | ||
Final O-acetylation is achieved by using an apparently partially active PKS and AT pair. Similar O-acylation systems are known in the cases of lovastatin12 where a diketide is added and in the case of squalestatin S113 where a tetraktetide is added. The multiforisins often feature a 9-aldehyde and this is installed by the SDR encoded by mfnR4 (e.g.11 from exp. 9).
Thus systematic heterologous expression of the mfn genes provides a convenient total biosynthesis platform for the multiforisins in good overall titres. However, attempts to redirect the pathway towards islandic acid 2 and allantopyrone 3 intermediates by adding the islandic acid triketide were unsuccessful. The IlaPKS2 has a carnitine acyl transferase (cAT) C-terminal domain. Such domains are very rare in fungal PKS, but in one case where a cAT domain has been investigated, Tang and coworkers showed that it was responsible for the reversible transfer of the completed polyketide to a polyol.14 Here we reasoned that a compound such as 1I could serve as the acceptor, but coexpression of the ilaPKS2 in cultures that produce 1I did not afford 2. Further addition of the putative AT encoded by ilaR2 did not help. We reasoned that the terminal carboxylate of 2 might be required as a recognition feature, but the ila BGC does not encode any additional monoxygenases that could oxidise position 8. The oxygenation of C-8 in A. oryzae was unreliably performed by a presumably A. oryzae native enzyme forming shunt metabolites 6 and 8. It is possible that 8-oxygenation during islandic acid biosynthesis may also be achieved by an oxygenase not encoded by the ila BGC, and that the ilaSDR-R8 may oxidise this to a carboxylic acid. However, inclusion of ilaSDR-R8 did not allow this chemistry.
Overall, we have achieved rapid total biosynthesis of a number of known and new fungal tetraketide pyrones and elucidated the pathway to the multiforisins. It should be possible to rationally engineer future members of this family using sustainable synthetic biology rather than multi-step synthetic chemistry methods.6 However, further work is required to understand the biosynthesis and attachment of the EZ triketide side chain of 2 and 3. Experiments in this direction are ongoing in our laboratories.
RJC, EK, DT and YS designed the study. DT and YS performed all bioinformatic analysis. JO, GLG and GA isolated and characterised compounds from H. spongiphila. YS and DT did all other experimental work. The MS was drafted by RJC, and all authors were involved in final editing.
The China Scholarship Council is thanked for scholarships to Y. S. (201908360165) and D. T. (201706670001). This work was also funded by the Deutsche Forschungsgemeinschaft (DFG) priority program “Taxon-Omics: New Approaches for Discovering and Naming Biodiversity” (SPP 1991), specifically CO 1328/4-2 and CO 1328/4-1. Professor Thomas Ostenfeld Larsen (Danish Technical University) is thanked for the gift of Penicillium islandicum. The publication of this article was partially funded by the Open Access Fund of the Leibniz University of Hannover.
Conflicts of interest
There are no conflicts to declare.References
- H. Fujimoto, J. Nat. Med., 2018, 72, 20–31 CrossRef CAS PubMed.
- Y. Fujimoto, H. Tsunoda, J. Uzawa and T. Tatsuno, J. Chem. Soc., Chem. Commun., 1982, 83–84 RSC.
- Y. Fujimoto, T. Kihara, K. Isono, H. Tsunoda, T. Tatsuno, K. Matsumoto and H. Hirokawa, Chem. Pharm. Bull., 1984, 32, 1583–1586 CrossRef CAS PubMed.
- M. S. R. Nair and S. T. Carey, Tetrahedron Lett., 1975, 16, 3517–3518 CrossRef.
- M. S. R. Nair, Phytochemistry, 1976, 15, 1090–1091 CrossRef CAS.
- T. Shimizu, S. Hiranuma, T. Watanabe and M. Kirihara, Heterocycles, 1994, 38, 243 CrossRef CAS.
- D.-S. Tian, E. Kuhnert, J. Ouazzani, D. Wibberg, J. Kalinowski and R. J. Cox, Chem. Sci., 2020, 11, 12477–12484 RSC.
- K. A. K. Pahirulzaman, K. Williams and C. M. Lazarus, Methods Enzymol., 2012, 517, 241–260 CAS.
- L. Kahlert, C. Schotte and R. J. Cox, Synthesis, 2021, 2381–2394, DOI:10.1055/a-1401-2716.
- S. Yuan, L. Chen, Q. Wu, M. Jiang, H. Guo, Z. Hu, S. Chen, L. Liu and Z. Gao, Mar. Drugs, 2022, 20, 294 CrossRef CAS PubMed.
- W. Cheng, M. Chen, M. Ohashi and Y. Tang, Angew. Chem., Int. Ed., 2022, 61, e202116928, DOI:10.1002/anie.202116928.
- X. Xie, K. Watanabe, W. A. Wojcicki, C. C. C. Wang and Y. Tang, Chem. Biol., 2006, 13, 1161–1169 CrossRef CAS PubMed.
- B. Bonsch, V. Belt, C. Bartel, N. Duensing, M. Koziol, C. Lazarus, A. Bailey, T. Simpson and R. J. Cox, Chem. Commun., 2016, 52, 6777–6780 RSC.
- L. Hang, M. Tang, C. J. Harvey, C. G. Page, J. Li, Y. Hung, N. Liu, M. E. Hillenmeyer and Y. Tang, Angew. Chem., Int. Ed., 2017, 56, 9556–9560 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3cc04758j |
| ‡ These authors contributed equally. |
| This journal is © The Royal Society of Chemistry 2023 |