
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Moving into the red – a near infra-red optical probe for analysis of human neutrophil elastase in activated neutrophils and neutrophil extracellular traps†
M.
Rodriguez-Rios
 a,
G.
Rinaldi
b,
A.
Megia-Fernandez
ac,
A.
Lilienkampf
a,
C. T.
Robb
b,
A. G.
Rossi
b and
M.
Bradley
*d
a,
G.
Rinaldi
b,
A.
Megia-Fernandez
ac,
A.
Lilienkampf
a,
C. T.
Robb
b,
A. G.
Rossi
b and
M.
Bradley
*d
aEaStCHEM School of Chemistry, University of Edinburgh, David Brewster Road, EH9 3FJ Edinburgh, UK
bUniversity of Edinburgh Centre for Inflammation Research, Queen's Medical Research Institute, 47 Little France Crescent, Edinburgh BioQuarter, Edinburgh EH16 4TJ, UK
cOrganic Chemistry Department, Faculty of Sciences, University of Granada, Avda. Fuente Nueva S/N, 18071, Spain
dPrecision Healthcare University Research Institute, Queen Mary University of London, Empire House, 67-75 New Road, London E1 1HH, UK. E-mail: m.bradley@qmul.ac.uk
First published on 5th September 2023
Abstract
Neutrophils are the first immune cells recruited for defence against invading pathogens; however, their dysregulated activation and subsequent release of the enzyme human neutrophil elastase is associated with several, inflammation-based, diseases. Herein, we describe a FRET-based, tri-branched (one quencher, three fluorophores) near infrared probe that provides an intense OFF/ON amplified fluorescence signal for specific detection of human neutrophil elastase. The probe allowed selective detection of activated neutrophils and labelling of neutrophil extracellular traps.
Neutrophils are the most abundant white cells in human blood. They are phagocytic immune cells that provide an early line of defence against pathogens,1 and apply a wide range of mechanisms to kill pathogens including phagocytosis, neutrophil degranulation, reactive oxygen radical generation,2 and neutrophil extracellular trap (NET) production during a process termed NETosis.3 In NETosis, neutrophils extrude their intracellular content (including chromatin and antimicrobial proteins) into the extracellular space, forming NETs to entrap and kill pathogens. However, despite their beneficial role in host defence, overactivation of neutrophils and dysregulated NETosis play pivotal roles in many inflammatory diseases,4 such as SARS-COV2 where NETs can cause microvascular thrombosis, leading to tissue damage, and organ failure.5 Of key relevance in neutrophil activity is human neutrophil elastase (hNE), a serine protease that is highly abundant in the granules of neutrophils. This protease is found bound to NETs, and is a validated biomarker for inflammation.6
Activity-based probes, which covalently bind to elastase, have allowed accurate cellular localisation of the enzyme and enzyme detection in gel-based assays, but most are “always on” fluorescent probes that can have high background signals when used for cell imaging microscopy and have no signal amplification.7–9 Probes based on Förster resonance energy transfer (FRET),10 on the other hand, allow signal amplification (probe turnover) and have been used for the detection of various proteases. Examples of FRET-based probes for elastase11–13 include FRET-based ratiometric probes14 and probes for elastase sensing that successfully allowed detection of activated neutrophils and visualisation of NETs.15 However, the majority of these activity and substrate-based probes for elastase lie within the “green” region (495–570 nm) of the visible spectrum where applications are limited by the high background signals commonly observed in biological tissues16 as well as by the elevated levels of tissue scattering.17 Thus, efforts on optical probe development have focused on improving the signal-to-noise ratio, often by moving the fluorescent emission of probes into the near infrared (NIR) window (650–900 nm), driving improvements in tissue penetration, reducing scattering (1/λ4) and background signal. Indeed, NIR probes are becoming common in the clinical setting with examples including the tyrosine kinase MET receptor optical probe for thyroid cancer detection,18 the FDA approved folate receptor targeting probe for ovarian cancer intraoperative imaging,19,20 and a cathepsin-activated imaging probe that has entered clinical studies for intraoperative breast cancer detection.21
Multibranched, substrate-based probes for protease detection offer a powerful means for signal amplification, with reduced background fluorescence when compared to their linear counter parts by virtue of “self-quenching”22 of identical, close proximity fluorophores and release of multiple fluorescent moieties upon cleavage.23,24 Herein, we present a tri-branched NIR probe for hNE that allows the specific detection of the enzyme in activated neutrophils and NETs. The probe was synthesised using a hybrid solid/solution-phase approach, with the final tri-branched construct assembled via copper catalysed cycloaddition chemistry.25,26
An initial design to explore the desired quencher and fluorophore targeted a classical, linear FRET peptide 1 (HNE-1F1Q) using a sulfonated Cy5 (λex/λem 640/657 nm) and QSY21 (λabs 660 nm) FRET pair on opposite ends of a hNE cleavable peptide (Glu-Glu-Ile-Nle-Arg-Arg) (Fig. 1).13,27 Probe 1 was synthesised on solid-phase using orthogonal Dde/Fmoc deprotection28 chemistry to allow selective attachment at the lysine side chain of the quencher QSY21 at the C-terminus, with coupling of the Cy5 fluorophore at the amino terminus as the final resin-based step before resin cleavage and deprotection (ESI,† Scheme S1). Surprisingly, when 1 (35 μM) was exposed to hNE, the activation proved to be very slow (>2 h, ESI,† Fig. S1), which was attributed to the bulky quencher hindering enzyme access to the cleavage site and the poor solubility of the probe (precipitation was observed in aqueous buffer even at low concentrations). Also, probe 1 did not show any increase in fluorescence when incubated with activated neutrophils.
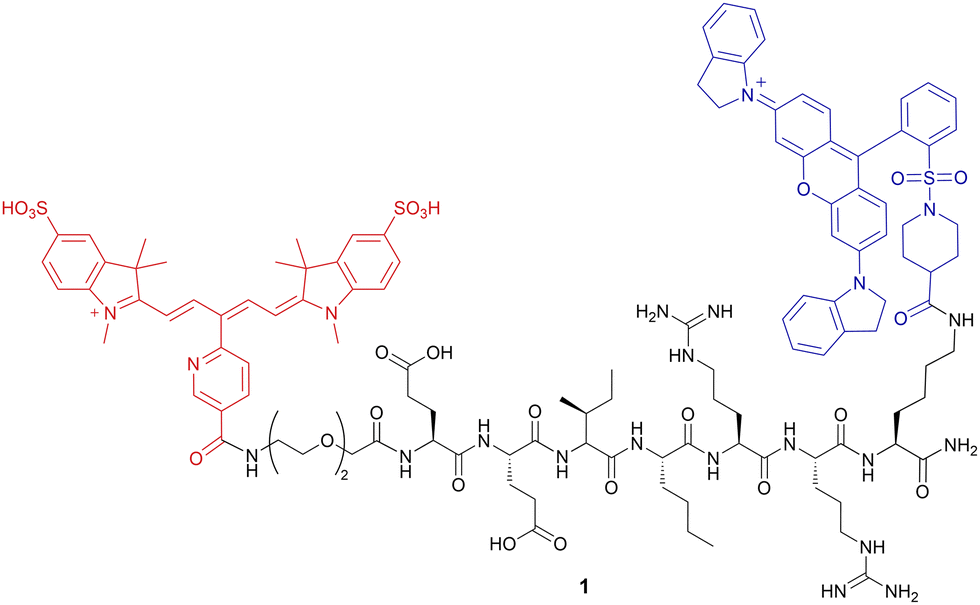 | ||
| Fig. 1 The structure of the linear FRET-probe 1 (HNE-1F1Q). hNE cleaves the probe between isoleucine and norleucine. | ||
To solve these problems, a tri-branched probe 10 was designed, in which a single QSY21 quencher was placed distally to the cleavage site (to improve cleavage kinetics) while using three copies of the sulfonated Cy5-peptide to allow “triple” levels of fluorescence amplification (Scheme 1). In this design, the quencher molecule enhances the overall silencing effect via FRET in addition to Cy5 self-quenching. Previously reported solid-phase synthetic methods used for the synthesis of the green emitting tribranched probes12 for elastase proved unsuccessful and, thus, a new convergent route was designed. The synthesis was achieved by the solid-phase synthesis of the azide-containing peptide 2 (with the N-terminus conjugated to a bis-sulfonated Cy5) on a Rink-amide linker functionalised polystyrene resin. This was followed by the convergent solution-phase attachment of 2 to the tri-branched, QSY21-alkyne scaffold 9via azide/alkyne “click” chemistry (Scheme 1).29 The tri-branched alkyne scaffold 9 was designed to have the quencher QSY21 attached to the “stem” via a bis-ethylene glycol spacer to ease its conjugation. Thus, the hindered amine of the tris-alkyne building block 630 was coupled to the spacer via a highly reactive acyl fluoride (Boc-NH-(EG)2-CO-F, 3). Once the spacer was attached, removal of the Boc group from 7 allowed facile incorporation of QSY21 via its NHS ester to give the quencher containing tris-alkyne 9. The tris-alkyne-QSY21 9 was then reacted with the azide peptide 2 (4 equiv.) at the highest reactant concentrations possible (limited by solubility – 0.6 mM and 2.5 mM for 9 and 2, respectively) using CuI/THTPA in anhydrous DMF to give the “three-fluorophores-one-quencher” probe 10 (HNE-3F1Q). Four equivalents of the azide 2 were sufficient for good conversion into the tri-branched compound, with only trace amounts of the mono and bis-functionalised intermediates ever observed by HPLC (ESI,† Fig. S2), suggesting, perhaps, that the mono and dimeric triazole species are catalysts in their own right.
 | ||
| Scheme 1 Synthesis of tri-branched probe 10. The quencher containing alkyne fragment 9 was synthesised from tris-hydroxymethyl aminomethane in six steps, with an overall yield of 20%. Azide-peptide 2 was synthesised using a Rink-amide linker functionalised polystyrene resin and assembled using standard Fmoc/tBu SPPS. The azide functionality was incorporated at the C-terminus using Fmoc-Lys(N3)-OH as the first residue and the peptide was capped with sulfonated Cy5 at its N-terminus (ESI,† Scheme S1). After the copper-catalysed click reaction, the probe was purified by semipreparative HPLC and characterised by MALDI-TOF MS and HRMS. | ||
Probe 10 (HNE-3F1Q) had a maximum excitation wavelength (λex) at 640 nm and maximum emission (λem) at 657 nm (Fig. 2a), consistent with the optical properties of the sulfonated Cy5 and, in contrast to the linear FRET probe 1, was highly water soluble. The probe was “super-silent” in its non-cleaved form, showing that a single QSY21 quencher can efficiently quench the three fluorophores within its proximity, with a rapid increase in fluorescence observed for 10 within seconds of adding hNE, (Fig. 2b), with cleavage occurring at Ile-Nle (Fig S7, ESI†). In comparison, the Cy5 tri-branched control probe 11 (HNE-3F0Q), without the QSY21 quencher, only gave a 2-fold increase in fluorescence upon elastase activation (ESI,† Fig. S3).
 | ||
| Fig. 2 (a) Fluorescence emission (λex = 640/10, λem = 680/20) of 10 (HNE-3F1Q, 12 μM) after incubation with hNE (100 nM, 1 h) (n = 3, Ctr = no enzyme); (b) time-dependent increase in fluorescence of 10 (12 μM) in the presence of hNE (100 nM). The activation was fully inhibited by the elastase inhibitor sivelestat (100 μM) (n = 3); (c) probe 10 (12 μM) was highly selective for hNE, with negligible cleavage by closely related proteases cathepsin G (100 nM) and proteinase 3 (100 nM) (n =3, Ctr = no enzyme). | ||
The KM for 10 was 5.7 μM with a turnover number (kcat/KM) of 3.5 × 104 M−1 s−1 (ESI,† Fig. S4). The cleavage kinetics of the branched probe 10 were superior to the more traditional, linear FRET peptide 1 (ESI,† Fig. S2–S6), with significantly faster cleavage (kcat/KM of 1.5 × 103 M−1 s−1). The activation of 10 was fully inhibited by the elastase inhibitor sivelestat and the probe was highly specific for elastase, with no cleavage by the closely related proteases, proteinase 3 or cathepsin G (Fig. 2c).
To evaluate if 10 (HNE-3F1Q) was able to detect hNE in neutrophils, primary human neutrophils were activated by exposure to two well stablished neutrophil activators N-formylmethionyl-leucyl-phenylalanine (fMLF) or phorbol 12-myristate 13-acetate (PMA).31–33 An increase in fluorescence from cleaved 10 was observed in activated neutrophils when compared to untreated controls that showed negligible fluorescence (Fig. 3). A clear difference between fMLF and PMA activation was observed with fMLF activated neutrophils having intact nuclei while PMA activated cells formed “cloud-like” structures, with the “DNA signal” diffusing into the extracellular environment and co-localised with the NIR signal of 10, indicating co-localisation of DNA and hNE which is a hallmark feature of NETs. When neutrophils were pre-treated with sivelestat prior to exposure to PMA, no significant increase in fluorescence was observed (Fig S8, ESI†). These results demonstrate that 10 can be used to detect hNE in activated neutrophils and specifically in NETs and that the probe is robust in the absence of the protease.
 | ||
| Fig. 3 Fluorescence microscopy images of neutrophils activated with fMLF (100 nM, 30 min) or PMA to induce NETs (10 nM, 3 h) while incubating with probe 10 (3 μM, 3 h), followed by nuclear staining with Hoechst 33342 (200 nM, 20 min). Detection wavelengths: blue channel (Hoechst) λex 405 nm, λem 420–500 nm; red channel (probe) λex 633 nm, λem 650–710 nm. | ||
Detection of hNE in activated neutrophils and NETs was possible with our NIR probe and results were consistent with those obtained with previously reported probes14,15 that used a similar substrate. However, our results contrasted with findings obtained from Kasperkiewicz9 that observed that the hNE present in NETs was not active when using phosphonate-based activity-based probes. These contrasting results could be explained by the nature of the probes, with activity-based probes unable to provide signal amplification (and having higher background fluorescence), while substrate-based probes, because they are catalytically activated by the protease, allow for much higher signals and provide lower limits of detection. In addition, activity-based probes lead to inhibition of the protease itself, which could have an effect the NETotic process. It would be interesting to perform a study where both methods of detection are compared side-by-side under the same conditions.
A tri-branched, FRET-based probe for the detection of hNE was synthesised with emission in the NIR region of the spectrum, where endogenous biomolecules have decreased absorbance, and minimal autofluorescence. The probe was synthesised using a convergent assembly method, combining solid-phase peptide synthesis and solution-phase copper catalysed azide/alkyne cycloaddition (CuAAC). Three copies of a sulfonated Cy5 N-terminal labelled elastase substrate, bearing an azide at the C-terminus, were “clicked” by CuAAC onto a tribranched scaffold bearing three terminal alkyne groups and a single copy of the quencher. This strategy allowed the quencher to be placed far from the cleavage site and proved to be key for efficient probe activation. The probe was able to rapidly detect hNE activity in vitro and was successfully used to image activated neutrophils and NETs.
We would like to thank Engineering and Physical Sciences Research Council (EPSRC, UK) for Interdisciplinary Research Collaboration grants EP/R005257/1 and Medical Research Council grant MR/N02995X/1.
Conflicts of interest
There are no conflicts to declare.Notes and references
- E. Kolaczkowska and P. Kubes, Nat. Rev. Immunol., 2013, 13, 159–175 CrossRef CAS PubMed.
- P. Lacy, Allergy, Asthma, Clin. Immunol., 2006, 2, 98 CrossRef PubMed.
- V. Brinkmann, U. Reichard, C. Goosmann, B. Fauler, Y. Uhlemann, D. S. Weiss, Y. Weinrauch and A. Zychlinsky, Science, 2004, 303, 1532–1535 CrossRef CAS PubMed.
- V. Mutua and L. J. Gershwin, Clin. Rev. Allergy Immunol., 2021, 61, 194–211 CrossRef CAS PubMed.
- Y. Zhu, X. Chen and X. Liu, Front. Immunol., 2022, 13, 838011 CrossRef CAS PubMed.
- S. J. Thulborn, V. Mistry, C. E. Brightling, K. L. Moffitt, D. Ribeiro and M. Bafadhel, Respir. Res., 2019, 20, 170 CrossRef PubMed.
- A. C. Schulz-Fincke, M. Blaut, A. Braune and M. Gutschow, ACS Med. Chem. Lett., 2018, 9, 345–350 CrossRef CAS PubMed.
- S. Y. Liu, H. Xiong, R. R. Li, W. C. Yang and G. F. Yang, Anal. Chem., 2019, 91, 3877–3884 CrossRef CAS PubMed.
- P. Kasperkiewicza, M. Porebaa, S. J. Snipasb, H. Parkerc, C. C. Winterbournc, G. S. Salvesenb and M. Drag, Proc. Natl. Acad. Sci. U. S. A., 2014, 111, 2518–2523 CrossRef PubMed.
- M. Rodriguez-Rios, A. Megia-Fernandez, D. J. Norman and M. Bradley, Chem. Soc. Rev., 2022, 51, 2081–2120 RSC.
- S. Gehrig, M. A. Mall and C. Schultz, Angew. Chem., Int. Ed., 2012, 51, 6258–6261 CrossRef CAS PubMed.
- S. Kossodo, J. Zhang, K. Groves, G. J. Cuneo, E. Handy, J. Morin, J. Delaney, W. Yared, M. Rajopadhye and J. D. Peterson, Int. J. Mol. Imaging, 2011, 2011, 581406 Search PubMed.
- B. Korkmaz, S. Attucci, M. A. Juliano, T. Kalupov, M. L. Jourdan, L. Juliano and F. Gauthier, Nat. Protoc., 2008, 3, 991–1000 CrossRef CAS PubMed.
- M. Guerra, V. S. Halls, J. Schatterny, M. Hagner, M. A. Mall and C. Schultz, J. Am. Chem. Soc., 2020, 142, 20299–20305 CrossRef CAS PubMed.
- M. R. Rios, G. Garoffolo, G. Rinaldi, A. Megia-Fernandez, S. Ferrari, C. T. Robb, A. G. Rossi, M. Pesce and M. Bradley, Chem. Commun., 2020, 57, 97–100 RSC.
- M. Monici, Biotechnol. Annu. Rev., 2005, 11, 227–256 CAS.
- J. R. Mourant, J. P. Freyer, A. H. Hielscher, A. A. Eick, D. Shen and T. M. Johnson, Appl. Opt., 1998, 37, 3586–3593 CrossRef CAS PubMed.
- P. K. C. Jonker, M. J. H. Metman, L. H. J. Sondorp, M. S. Sywak, A. J. Gill, L. Jansen, T. P. Links, P. J. van Diest, T. M. van Ginhoven, C. W. G. M. Löwik, A. H. Nguyen, R. P. Coppes, D. J. Robinson, G. M. van Dam, B. M. van Hemel, R. S. N. Fehrmann and S. Kruijff, Eur. J. Nucl. Med. Mol. Imaging, 2022, 49, 3557–3570 CrossRef CAS PubMed.
- J. L. Tanyi, H. S. Chon, M. A. Morgan, S. K. Chambers, E. S. Han, K. A. Butler, C. L. Langstraat, M. A. Powell, L. M. Randall, A. L. Vahrmeijer, I. S. Winer and R. M. Wenham, J. Clin. Oncol., 2021, 39, 5503 Search PubMed.
- S. M. Mahalingam, S. A. Kularatne, C. H. Myers, P. Gagare, M. Norshi, X. Liu, S. Singhal and P. S. Low, J. Med. Chem., 2018, 61, 9637–9646 CrossRef CAS PubMed.
- B. L. Smith, M. A. Gadd, C. R. Lanahan, U. Rai, R. Tang, T. Rice-Stitt, A. L. Merrill, D. B. Strasfeld, J. M. Ferrer, E. F. Brachtel and M. C. Specht, Breast Cancer Res. Treat., 2018, 171, 413–420 CrossRef PubMed.
- W. Bae, T.-Y. Yoon and C. Jeong, PLoS One, 2021, 16, e0247326 CrossRef CAS PubMed.
- N. Avlonitis, M. Debunne, T. Aslam, N. McDonald, C. Haslett, K. Dhaliwal and M. Bradley, Org. Biomol. Chem., 2013, 11, 4414–4418 RSC.
- T. H. Craven, N. Avlonitis, N. McDonald, T. Walton, E. Scholefield, A. R. Akram, T. S. Walsh, C. Haslett, M. Bradley and K. Dhaliwal, Sci. Rep., 2018, 8, 13490 CrossRef PubMed.
- M. Meldal and C. W. Tornøe, Chem. Rev., 2008, 108, 2952–3015 CrossRef CAS PubMed.
- C. W. Tornøe, C. Christensen and M. Meldal, J. Org. Chem., 2002, 67, 3057–3064 CrossRef PubMed.
- B. Korkmaz, S. Attucci, E. Hazouard, M. Ferrandière, M. L. Jourdan, M. Brillard-Bourdet, L. Juliano and F. Gauthier, J. Biol. Chem., 2002, 277, 39074–39081 CrossRef CAS PubMed.
- J. J. Diaz-Mochon and Mark Bradley, Org. Lett., 2004, 6, 1127–1129 CrossRef CAS PubMed.
- N. J. Agard, J. A. Prescher and C. R. Bertozzi, J. Am. Chem. Soc., 2004, 126, 15046–15047 CrossRef CAS PubMed.
- Y. M. Chabre, C. Contino-Pépin, V. Placide, T. C. Shiao and R. Roy, J. Org. Chem., 2008, 73, 5602–5605 CrossRef CAS PubMed.
- H. Takei, A. Araki, H. Watanabe, A. Ichinose and F. Sendo, J. Leukocyte Biol., 1996, 59, 229–240 CrossRef CAS PubMed.
- A. G. Mahomed and R. Anderson, Inflammation, 2000, 24, 559–569 CrossRef CAS PubMed.
- G. T. Nguyen, E. R. Green and J. Mecsas, Front. Cell. Infect. Microbiol., 2017, 7, 373 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Supporting figures and schemes, and experimental procedures. See DOI: https://doi.org/10.1039/d3cc03634k |
| This journal is © The Royal Society of Chemistry 2023 |