
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceValorisation of xylose to renewable fuels and chemicals, an essential step in augmenting the commercial viability of lignocellulosic biorefineries
Vivek
Narisetty
a,
Rylan
Cox
ab,
Rajesh
Bommareddy
 c,
Deepti
Agrawal
d,
Ejaz
Ahmad
e,
Kamal Kumar
Pant
f,
Anuj Kumar
Chandel
g,
Shashi Kant
Bhatia
h,
Dinesh
Kumar
i,
Parmeswaran
Binod
j,
Vijai Kumar
Gupta
k and
Vinod
Kumar
*af
c,
Deepti
Agrawal
d,
Ejaz
Ahmad
e,
Kamal Kumar
Pant
f,
Anuj Kumar
Chandel
g,
Shashi Kant
Bhatia
h,
Dinesh
Kumar
i,
Parmeswaran
Binod
j,
Vijai Kumar
Gupta
k and
Vinod
Kumar
*af
aSchool of Water, Energy and Environment, Cranfield University, Cranfield MK43 0AL, UK. E-mail: Vinod.Kumar@cranfield.ac.uk; Tel: +44 (0)1234754786
bSchool of Aerospace, Transport and Manufacturing, Cranfield University, Cranfield MK43 0AL, UK
cDepartment of Applied Sciences, Northumbria University, Newcastle upon Tyne NE1 8ST, UK
dBiochemistry and Biotechnology Area, Material Resource Efficiency Division, CSIR- Indian Institute of Petroleum, Mohkampur, Dehradun 248005, India
eDepartment of Chemical Engineering, Indian Institute of Technology (ISM), Dhanbad 826004, India
fDepartment of Chemical Engineering, Indian Institute of Technology Delhi, New Delhi 110016, India
gDepartment of Biotechnology, Engineering School of Lorena (EEL), University of São Paulo, Lorena 12.602.810, Brazil
hDepartment of Biological Engineering, College of Engineering, Konkuk University, Seoul 05029, Republic of Korea
iSchool of Bioengineering & Food Technology, Shoolini University of Biotechnology and Management Sciences, Solan 173229, Himachal Pradesh, India
jMicrobial Processes and Technology Division, CSIR-National Institute for Interdisciplinary Science and Technology (CSIR-NIIST), Thiruvananthapuram 695 019, Kerala, India
kScotland's Rural College, Barony Campus, Dumfries DG1 3NE, UK
First published on 26th October 2021
Abstract
Biologists and engineers are making tremendous efforts in contributing to a sustainable and green society. To that end, there is growing interest in waste management and valorisation. Lignocellulosic biomass (LCB) is the most abundant material on the earth and an inevitable waste predominantly originating from agricultural residues, forest biomass and municipal solid waste streams. LCB serves as the renewable feedstock for clean and sustainable processes and products with low carbon emission. Cellulose and hemicellulose constitute the polymeric structure of LCB, which on depolymerisation liberates oligomeric or monomeric glucose and xylose, respectively. The preferential utilization of glucose and/or absence of the xylose metabolic pathway in microbial systems cause xylose valorization to be alienated and abandoned, a major bottleneck in the commercial viability of LCB-based biorefineries. Xylose is the second most abundant sugar in LCB, but a non-conventional industrial substrate unlike glucose. The current review seeks to summarize the recent developments in the biological conversion of xylose into a myriad of sustainable products and associated challenges. The review discusses the microbiology, genetics, and biochemistry of xylose metabolism with hurdles requiring debottlenecking for efficient xylose assimilation. It further describes the product formation by microbial cell factories which can assimilate xylose naturally and rewiring of metabolic networks to ameliorate xylose-based bioproduction in native as well as non-native strains. The review also includes a case study that provides an argument on a suitable pathway for optimal cell growth and succinic acid (SA) production from xylose through elementary flux mode analysis. Finally, a product portfolio from xylose bioconversion has been evaluated along with significant developments made through enzyme, metabolic and process engineering approaches, to maximize the product titers and yield, eventually empowering LCB-based biorefineries. Towards the end, the review is wrapped up with current challenges, concluding remarks, and prospects with an argument for intense future research into xylose-based biorefineries.
1. Introduction
Biomass is a potential alternative to non-renewable and non-sustainable fossil fuels causing massive harm to the atmosphere through colossal carbon emission and generation of pollutants.1 Analogous to a petroleum refinery, a biorefinery processes biomass into multiple products with a green and sustainable approach leading to low carbon biomanufacturing technologies.1,2 First generation biorefineries making use of edible feedstocks such as sugar, starch, and vegetable oils for generating biofuels are well established, but pose a significant concern and are a regular subject of the food vs. fuel debate.3 On the other hand, second generation biorefineries based on non-edible feedstocks such as lignocellulosic biomass (LCB) do not interfere in any food chain and offer a clear value proposition for the production of bulk and speciality chemicals. LCB is the most abundant feedstock on the planet (∼200 billion tonnes) with a significant contribution stemming from post-harvest agricultural residues. It is composed of lignin (15–20%), the outermost protective layer, cellulose (40–50%), the inner amorphous and crystalline component of the secondary wall, and hemicellulose (25–30%) microfibrils that connect the outermost and inner cellulose layers (Fig. 1A).4 Cellulose is a linear homo-polymer of D-glucose units connected by β-1,4-glycosidic bonds, and hemicellulose is a complex hetero-polymer containing D-xylose, L-arabinose, D-glucose, L-galactose, D-mannose, D-glucuronic acid and D-galacturonic acid (Fig. 1B). Hemicelluloses constitute 26% dry weight in hard woods, 22% in soft woods, and up to 25% in agro-residues with various polymeric forms such as xylan, arabinoxylan, xyloglucan, and glucuronoxylan.5,6 To utilize this three-dimensional polymeric structure as the feedstock for fermentative production of value-added chemicals, the polymer is converted into simple fermentable sugars. However, the major limitation is that most of the microorganisms are incapable of metabolizing all the fermentable sugars present in LCB, especially pentoses. The pentose sugars are present in the hemicellulosic fraction with xylan as the major polysaccharide which is composed of β-1,4-linked xylose residues. The depolymerization of the hemicellulosic fraction generates a mixture of sugars containing ∼90% xylose. In fact, xylose is the second most abundant sugar available after glucose in LCB (Fig. 1A).7 Despite this, the application of xylose as a potential feedstock is overlooked for biorefineries and it is discarded as waste or incinerated for energy purposes. This is due to a lack of efficient fermentation systems, as many of the microorganisms do not have a native pathway for metabolizing xylose. In addition, uptake of xylose is suppressed in the presence of glucose due to carbon catabolite repression.8 That is why the number of literature reports using glucose as a substrate for bioproduction is much larger in comparison to that using xylose. However, while exploiting biochemical platforms, the techno-commercial success of an LCB-based biorefinery largely thrives on the revival of the carbohydrate economy, which in turn is dependent on efficient depolymerization of both the structural polysaccharides to simple sugars and their subsequent valorisation to various commercially important products either through chemical or biotechnological routes.9,10 Therefore, efficient conversion of xylose is necessary and it is imperative to find robust microbial systems for metabolizing xylose for simultaneous assimilation of glucose and xylose for the pragmatic development of profitable LCB-based biorefineries.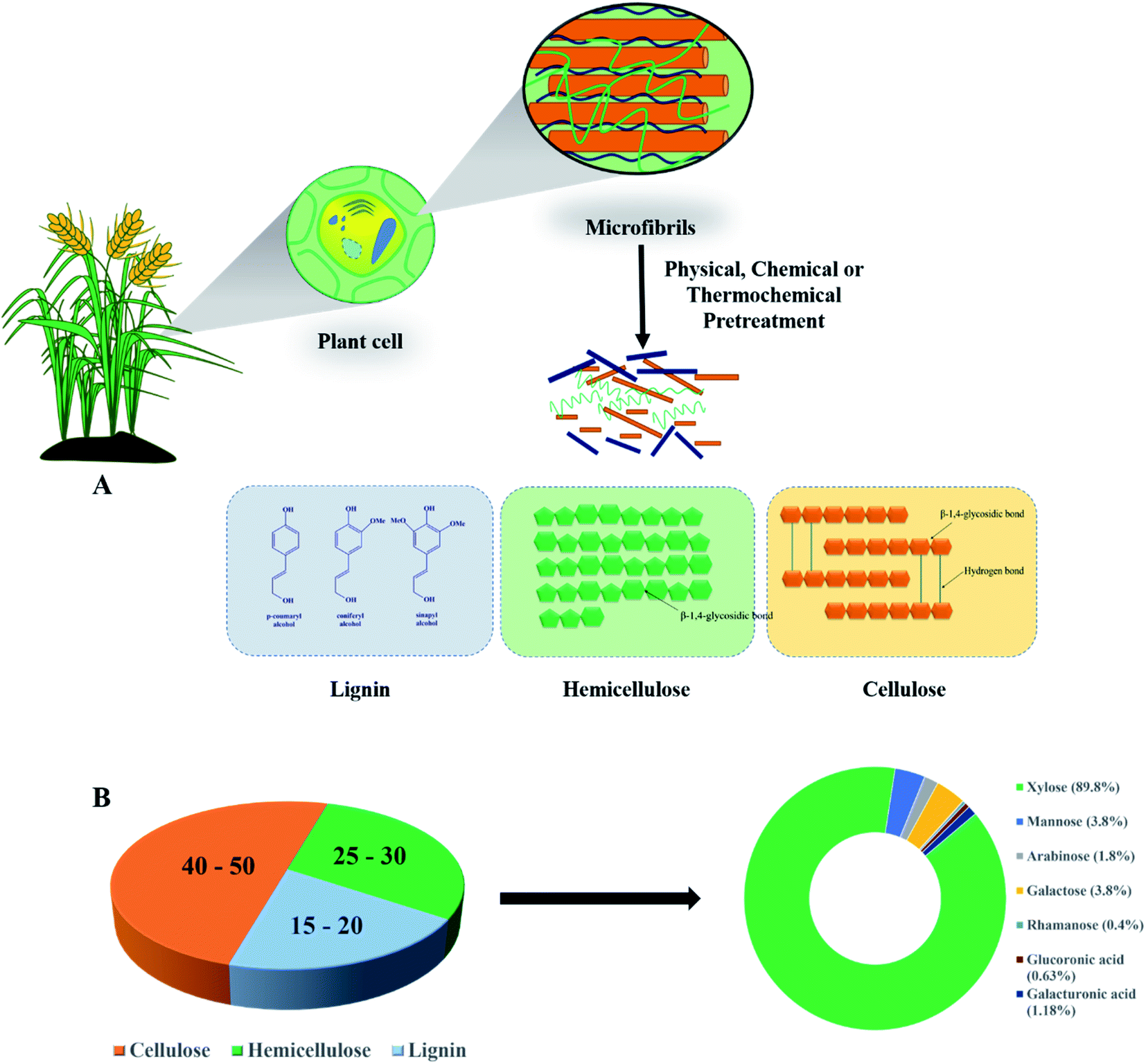 | ||
| Fig. 1 (A) Structural components of lignocellulosic biomass (LCB). (B)Illustration of the composition of individual subunits of LCB and compositions of sugar and sugar acids in the hemicellulosic fraction. | ||
Considering the challenges associated with xylose utilization, the current review (i) covers the efficient pretreatment processes assisting in xylan extraction from different LCB residues, (ii) discusses the bottlenecks impeding xylose assimilation and strategies to overcome them, (iii) describes the major native and engineered microbial cell factories available for efficient bioconversion of xylose to chemical building blocks, (iv) includes implementation of elementary flux mode analysis to understand the optimal pathway for xylose utilization to produce biomass and end metabolites with a case study of succinic acid, and (v) briefly covers alternative chemical catalysis of xylose for manufacturing value-added products. Finally, the limitations and future perspectives for constructing microbial cell factories to effectively utilize xylose and produce a wide array of products are included.
1.1 Pretreatment strategies for the extraction of fermentable sugars from LCB
Recalcitrance is a natural and intrinsic feature of any LCB, originating from its three principal constituents, cellulose, hemicellulose, and lignin, that chemically interact to form a complex network popularly known as a lignin–carbohydrate complex (LCC).11,12 During biorefining via a biochemical route, pretreatment is an imperative module that disrupts the lignocellulosic matrix by breaking LCC linkages leading to delignification and partial or complete hydrolysis of xylan, thereby improving the surface characteristics of biomass and enhancing the accessibility of cellulose for enzymatic hydrolysis. Invariably, most of the traditional pretreatment strategies primarily result in lignin removal, releasing fermentable sugars from the thermolabile hemicellulosic/xylan fraction, or are focused on selective delignification enriching the biomass in glucan and xylan fractions.13| Type of LCB | Type of pretreatment | Pretreatment conditions | Biomass composition (%) | Removal (%) | References | ||
|---|---|---|---|---|---|---|---|
| Before pretreatment | After pretreatment | Xylan | Lignin | ||||
| a MW: maple wood; SCB: sugarcane bagasse; SG: switchgrass; WS: wheat straw; CC: corn cob; SS: sugarcane straw; DA: dilute acid; SE: steam explosion; LHW: liquid hot water; HT: hydrothermal; Gln: glucan; Xln: xylan; HC: hemicellulose; KL: Klason lignin: TL: acid soluble and insoluble lignin; #XMG: xylan, mannan and galactan. | |||||||
| Poplar | DA | Temp: 170 °C; time: 8.5 min; H2SO4: 0.5% (w/w) | Gln-57.9; HC-17.5; KL-24.6 | Gln-74.2; HC-<2.0; KL-25 | 99 | — | 198 |
| MS | HT pretreatment at low acid | Temp: 180 °C; 10 min; H2SO4: 0.3% (w/w) | Gln-41.9; #XMG-22.1; KL-22.0 | Gln-64.4; #XMG-5.0; KL-29.3 | 86.4 | 20.2 | 199 |
| CS | DA in steam gun | Temp: 160 °C; time: 5 min; H2SO4: 2% (w/w) | Gln-34.0; Xln-22.0; KL-12.3 | Gln-57.4; Xln-3.2; KL-24.8 | ∼92.8 | ∼<1.5 | 200 |
| SCB | DA | Temp: 140 °C; time: 8 min; H3PO4: 0.2% (w/v) | Gln- 40.1; HC-27.5; TL-18.5 | Gln-58.5; HC-1.8; TL-29.05 | 96.5 | 14.8 | 201 |
| SG | DA | Temp: 160 °C; time: 30 min; H2SO4: 1% (w/w) | Gln-33.5; Xln-22.7; KL-16.3 | Gln-53.2; Xln-0.8; KL-33.3 | 98.6 | 18.3 | 202 |
| WS | DA | Temp: 140 °C; time: 90 min; H2SO4: 0.5% (w/w) | Gln-43.2; Xln-24.4; KL-20.8 | Gln-59.1; Xln-2.4; KL-30.7 | 91.5 | — | 203 |
| CS | HT | Temp: 180 °C | Gln-36.1; Xln-21.4; TL-13.6 | Gln-33.0; Xln: 5.4; TL-13.5 | 74.9 | — | 200 |
| CC | HT | Temp: 207 °C | Gln-28.8; Xln-29.6; KL-18.6 | Gln-54.5; Xln-10.2; KL-21.8 | 80.4 | 33.1 | 204 |
| SS | LHW | Temp: 220 °C; time: 5 min | Gln-33.13; HC-26.2; KL-18.2 | Gln-56.7; Xln-2.0; KL-37.0 | 96.5 | 6.6 | 205 |
| SCB | H3PO4 catalysed SE | Temp: 195 °C; time: 7.5 min; H3PO4: 0.95% (w/w) | Gln-31.8; Xln-12.2; KL-24.3 | Gln-49.7; Xln-2.3; KL-31.9 | 90.6 | 14.4 | 206 |
| H2SO4 catalysed SE | Temp: 195 °C; time: 7.5 min; H2SO4: 0.2% (w/w) | Gln-49.4; Xln-3.3; KL-31.5 | 86.6 | 12.1 | |||
| LCB type | Reactor type | Reaction conditions | Biomass composition (%) | Composition of pre-hydrolysate | References | ||
|---|---|---|---|---|---|---|---|
| Untreated | Pretreated | Sugars | Non-sugar component | ||||
| a CC: corn cobs; SCB: sugarcane bagasse; RS: rice straw; WS: wheat straw; EG: Eucalyptus grandis; CS: corn stover; MA: Miscanthus; Gln: glucan; Xln: xylan; HC: hemicellulose; KL: Klason lignin; TL: acid soluble and insoluble lignin; C5: pentose sugars; C6: hexose sugars; XOS: xylooligosaccharides; GOS: glucooligosaccharides; DA: dilute acid; TPL: total phenolics; TA: total aromatics; wt%: Wt in g/100 g biomass. | |||||||
| CC | Screw steam explosive extruder | Pressure: 15.5 bar | Gln-42.23 | Xylose: 27.5 wt% | Acetic acid: 1.1 wt% | 207 | |
| Time: 5.5 min | HC-39.01 | XOS: 2.4 wt% | TP: 1.7 wt% | ||||
| H2SO4: 2.4% (w/w) + steam explosion | KL-14.42 | Glucose: 3.9 wt% | Furfural: 0.5 wt% | ||||
| Arabinose: 3.7 wt% | 5 HMF: 0.2 wt% | ||||||
| SCB | 350-L SS reactor with stirrer & thermal oil heating | Temp: 120 °C | Gln-45.1 | Gln-54.6 | C5: 17.4 g L−1 | Acetic acid: 2.3 g L−1 | 208 |
| Time: 10 min | HC-26.9 | HC-10 | TA: 7.5 g L−1 | ||||
| H2SO4: 1% (w/v) | KL-22.2 | KL-32 | C6: 1.6 g L−1 | Furfural: 0.8 g L−1 | |||
| 5 HMF: 0.2 g L−1 | |||||||
| WS | Continuous pretreatment reactor (250 kg day−1) | Temp: 160 °C | Gln-47.1 | Gln-63.1 | Xylose: 29.2 g L−1 | Acetic acid: 1.9 g L−1 | 209 |
| Pressure: 5.2 bar | |||||||
| Time: 10 min | HC-24.3 | HC-1.0 | Glucose: 8.4 g L−1 | Furfural: 0.9 g L−1 | |||
| H2SO4: 0.5% (v/v) | KL-28.5 | KL-35.8 | Arabinose: 2.6 g L−1 | 5 HMF: 0.6 g L−1 | |||
| EG | 150 L horizontal Andritz reactor | Temp: 180 °C; time: 15 min; H2SO4: 2.4% (w/w) + steam explosion | Gln-38.5 | Gln-55.5 | 92% xylan recoverable and 74% as xylose | Acetic acid: 2.9 wt% | 210 |
| Xln-11.0 | Xln: 0.8 | Furfural: 0.9 wt% | |||||
| KL-25.2 | KL-37.1 | 5 HMF: 0.2 wt% | |||||
| RS | Continuous pretreatment reactor (250 kg day−1) | Temp: 162 °C; time: 10 min | Gln-37 | Gln-51.8 | 100 g xylose in hydrolysate/kg initial dry substrate | Acetic acid: 2 g L−1 | 211 |
| Final H2SO4: 0.35% (w/w) | Xln-20 | Xln-3.6 | Furfural: 1.2 g L−1 | ||||
| Preasoaking in acid: 0.5 h acid | TL-13.4 | TL-28.8 | 5 HMF: 1.1 g L−1 | ||||
| WS | Steam explosion in a 30L rig | Pressure: 12 bar; time: 12 min; final H3PO4: 1.2% (w/v); acid pre-soaked biomass introduced | Gln-41.6 | Xylose: 17.7 wt% | — | 212 | |
| Xln-30.3 | |||||||
| TL-19.3 | |||||||
| CS | Gln-38.5 | Xylose: 13.9 wt% | — | ||||
| Xln-24.3 | |||||||
| TL-18.3 | |||||||
| MS | Gln-47.0 | Xylose: 14.7 wt% | — | ||||
| Xln-25.1 | |||||||
| TL-26.15 | |||||||
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 ethanol–water mix and 80 mM H2SO4 at 150 °C for 70 min, ∼82.7 wt% lignin yield was obtained in the liquid fraction leading to the generation of a highly digestible cellulose-rich pulp.23 Earlier, Smit and Huijen evaluated seven different feedstocks: wheat straw, corn stover, beechwood, poplar, birchwood, spruce, and pine for mild organosolv pretreatment with 50% acetone and <50 mM H2SO4 at 140 °C for 2 h. Irrespective of biomass type, 87–97% xylan hydrolysis was observed. Poor delignification yields were obtained only in spruce and pine, while glucan recoveries ranged between 68 and 94%. Later the group precipitated the dissolved lignin by diluting with water, leading to effective fractionation of all three components of different LCBs.24
:1 ethanol–water mix and 80 mM H2SO4 at 150 °C for 70 min, ∼82.7 wt% lignin yield was obtained in the liquid fraction leading to the generation of a highly digestible cellulose-rich pulp.23 Earlier, Smit and Huijen evaluated seven different feedstocks: wheat straw, corn stover, beechwood, poplar, birchwood, spruce, and pine for mild organosolv pretreatment with 50% acetone and <50 mM H2SO4 at 140 °C for 2 h. Irrespective of biomass type, 87–97% xylan hydrolysis was observed. Poor delignification yields were obtained only in spruce and pine, while glucan recoveries ranged between 68 and 94%. Later the group precipitated the dissolved lignin by diluting with water, leading to effective fractionation of all three components of different LCBs.24
Recently, Xu et al. devised a mild technique for hemicellulose extraction from poplar wood with a binary solvent system containing formic acid and water. Pretreatment at 90 °C for 4 h resulted in 73.1% xylose yield while the solvent was recovered by fractional distillation and recycled back for a second round of pretreatment.25 The following section emphasizes the use of novel solvents for complete LCB fractionation. Chen et al. used 1% H2SO4 with 75% choline chloride to fractionate cellulose of switchgrass from lignin and xylan fractions. Treatment with this acidified deep eutectic solvent (DES) at 120 °C for 25 min removed 76% of the xylan fraction along with 51.1% delignification. Five cycles of recycling and reuse of this acidified liquor enriched the hydrolysed xylan and lignin fraction. Later, the group used xylose-rich liquor for furfural production at 160 °C for 15 min with 2% w/v AlCl3 and recovered lignin.26 Very recently, a biphasic acidic water/phenol system was used for the fractionation of Populus wood chips.27 This unique biphasic system enriched the water-soluble phase with 77% xylose and negligible by-products when the chips were subjected to 120 °C for an hour. In contrast, the phenolic phase contained 90% dissolved lignin (90%), leaving solids retaining 96% of the original cellulosic fraction.27 Likewise, a novel biphasic system comprising 2-phenoxyethanol and acidified water (70:30) was used to fractionate rice straw.28 Pretreatment at 130 °C for 2 h led to cellulose-rich (86.48% retained) biomass, facilitated by 92.1 and 63.16% removal of hemicellulose and lignin fractions, respectively. Later, 92.6% pure lignin was recovered by simple precipitation and 81.83% of xylan/xylose enriched in the aqueous phase.28 Yang et al. evaluated the effect of p-toluenesulfonic acid (p-TsOH) on the fractionation of three feedstocks: corncobs, wheat straw, and miscanthus. Pretreatment at 80 °C for 10 min resulted in significant removal of lignin and xylan, leaving a cellulose-rich pulp. Later, spent liquor was diluted to precipitate lignin, and the reusability of p-TsOH was shown to be ∼5 times higher.29 A similar attempt was made by yet another green hydrotrope, maleic acid (MA), for the effective fractionation of birchwood.30 At 100 °C and 50 wt% MA, 94.5% of the cellulosic fraction was obtained as a solid after 30 min. Lignin was precipitated by dilution, and the solubilised xylan was converted to furfural with ∼70% yield. Furthermore, MA displayed ∼3 times recyclability with comparable performance.30 Earlier, the cosolvent enhanced lignocellulose fractionation (CELF) method was developed for pre-treating corn stover using 0.5% H2SO4 and tetrahydrofuran (THF) in the ratio of 1:1. The dilute acid hydrolysed xylan to xylose which later dehydrated to furfural, while THF led to lignin dissolution enriching the cellulosic biomass. Later, the group separated furfural from THF. The latter was recovered by vacuum distillation and recycled, leaving lignin as a powder.31
2. Chemo-catalytic transformation of xylose to high-value chemicals
Chemo-catalytic routes are the conventional processes for the conversion of petroleum derivatives into bulk, fine and speciality chemicals. Like biological routes, several chemical routes exist via which xylose can be converted to a wide range of products such as furfural, furfuryl alcohol, xylitol, levulinic acid, levulinic ester, and other value-added chemicals, as shown in Fig. 2.32,33 In general, xylose conversion proceeds either via hydrogenation reaction in the presence of a metal catalyst to yield xylitol or isomerization reaction in the presence of a Lewis acid catalyst to produce xylulose. Xylulose further dehydrates to yield furfural in the presence of a Brønsted acid catalyst. Notably, furfural's estimated global market size was valued at $1.2 billion in 2019 and is expected to grow further to $2 billion by 2027 which makes it the most attractive and widely produced product from xylose (https://www.alliedmarketresearch.com/press-release/furfural-market.html, accessed on 29-03-2021). Xylose can be converted to furfural via an enol route, β-elimination, tautomerization and several other routes in the presence of a homogeneous or a heterogeneous catalyst, and the catalyst must have acidic properties.34 Therefore, a wide range of homogeneous mineral acids such as sulfuric acid, hydrochloric acid, nitric acid, phosphoric acid, acetic acid, and formic acid have been used for xylose conversion to furfural.35 It is noteworthy that 60–70% of the total furfural produced globally is used for manufacturing furfuryl alcohol.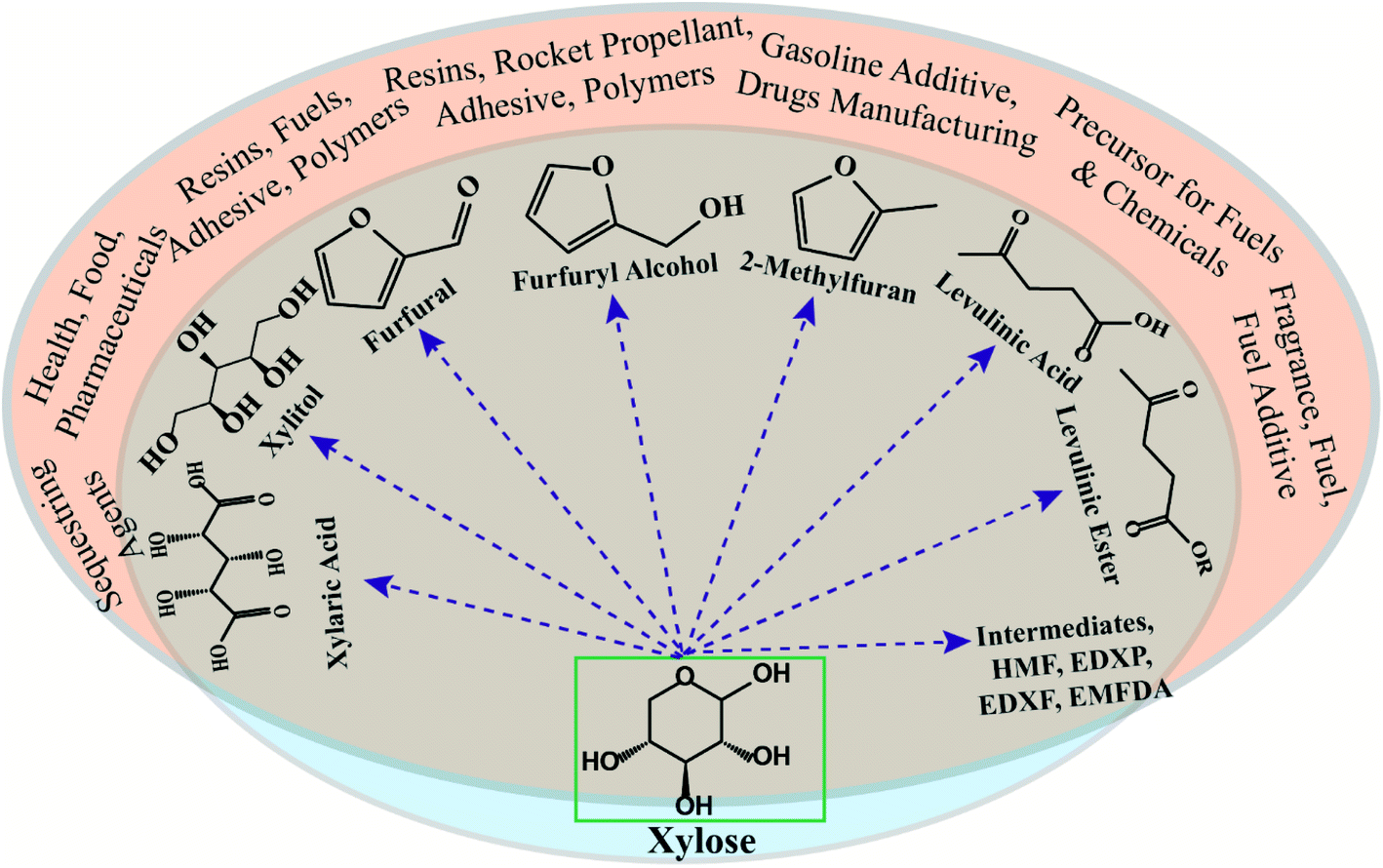 | ||
| Fig. 2 Catalytic routes for xylose conversion to value-added chemicals. | ||
Interestingly, xylose can also be directly converted to furfuryl alcohol via the hydrogenation route by using a metal catalyst. The development of such processes may minimize the conventional multistep and tedious method of converting xylose to furfural and then hydrogenating it to produce furfuryl alcohol. In this regard, the Zhu group reported 87.2% furfuryl alcohol yield in the presence of a Cu/ZnO/Al2O3 catalyst in a continuous fixed-bed reactor at a temperature of 150 °C.36 Furthermore, the authors observed that increasing the reaction temperature to 190 °C alters the xylose conversion pathway to yield 86.8% 2-methyl furan.36 Therefore, the final product from xylose conversion via hydrogenation can be altered by tuning the operating parameters and catalytic materials. For example, Li and co-workers have carried out the xylose conversion to levulinic acid and levulinic esters in a high-pressure hydrogenation reactor using a Pd/Al2O3 catalyst. The conversion yields of levulinic acid and levulinic esters achieved were 40 and 10%, respectively.37
It is also worth mentioning that hydrogenation of xylose at an industrial scale is being done since the 1970s to produce an essential chemical, xylitol.38 In general, the xylitol production process takes place in the presence of a metal catalyst and hydrogen source at 353–413 K temperature and 1–8 MPa pressure for 15–360 minutes of reaction time.39 The xylose to xylitol conversion is a surface controlled reaction; therefore, the interaction between adsorbed/unadsorbed xylose and chemisorbed hydrogen and the catalyst surface dictates the process's overall yield. In contrast, the product xylitol does not desorb easily from the catalyst surface, thereby causing the catalyst's saturation.40 Interestingly, some of these metal catalysts can also be used to convert xylose into xylaric acid via oxidation reaction. For example, Saha and co-workers have observed 60% xylaric acid yield in the presence of a Pt/C catalyst via oxidation reaction. However, limited data are available for such reactions. The xylose-based conversion via chemical routes suffers from lower yields and furthermore, the use of acidic catalysts and reaction operation at higher temperatures and pressures make the process environmentally unfriendly. Although many chemical processes such as xylitol production are running at the commercial scale, the long-term sustainability is doubtful due to the high cost of production and environmental incompatibility.
3. Xylose metabolism: genetics and biochemistry of enzymes and their regulation
Xylose valorisation through biotechnological intervention has the potential to become the most popular route for producing various bio-based chemicals and fuels. A diverse group of microbes such as bacteria, yeast, and fungi are known to assimilate xylose naturally through different metabolic pathways leading to formation of a range of products such as xylitol, 2,3-butanediol, ethanol-n-butanol, lactic acid, succinic acid, etc. A considerable knowledge on these pathways can provide guidance in constructing efficient xylose assimilatory strains.3.1 Xylose assimilation
The process of D-xylose assimilation is quite different from that of D-glucose assimilation, which is metabolized through the Embden–Meyerhof–Parnas (EMP) pathway. D-Xylose undergoes isomerization or reduction and subsequent oxidation to form D-xylulose. D-Xylulose is the key intermediate for the pentose phosphate pathway, and upon phosphorylation is converted into xylulose-5-phosphate (X5P), which is funnelled to the central carbon metabolism to generate C3–C7 metabolites (Fig. 3). These metabolites can be either precursors or intermediates for the EMP pathway, and biosynthesis of amino acids and nucleotides.41,42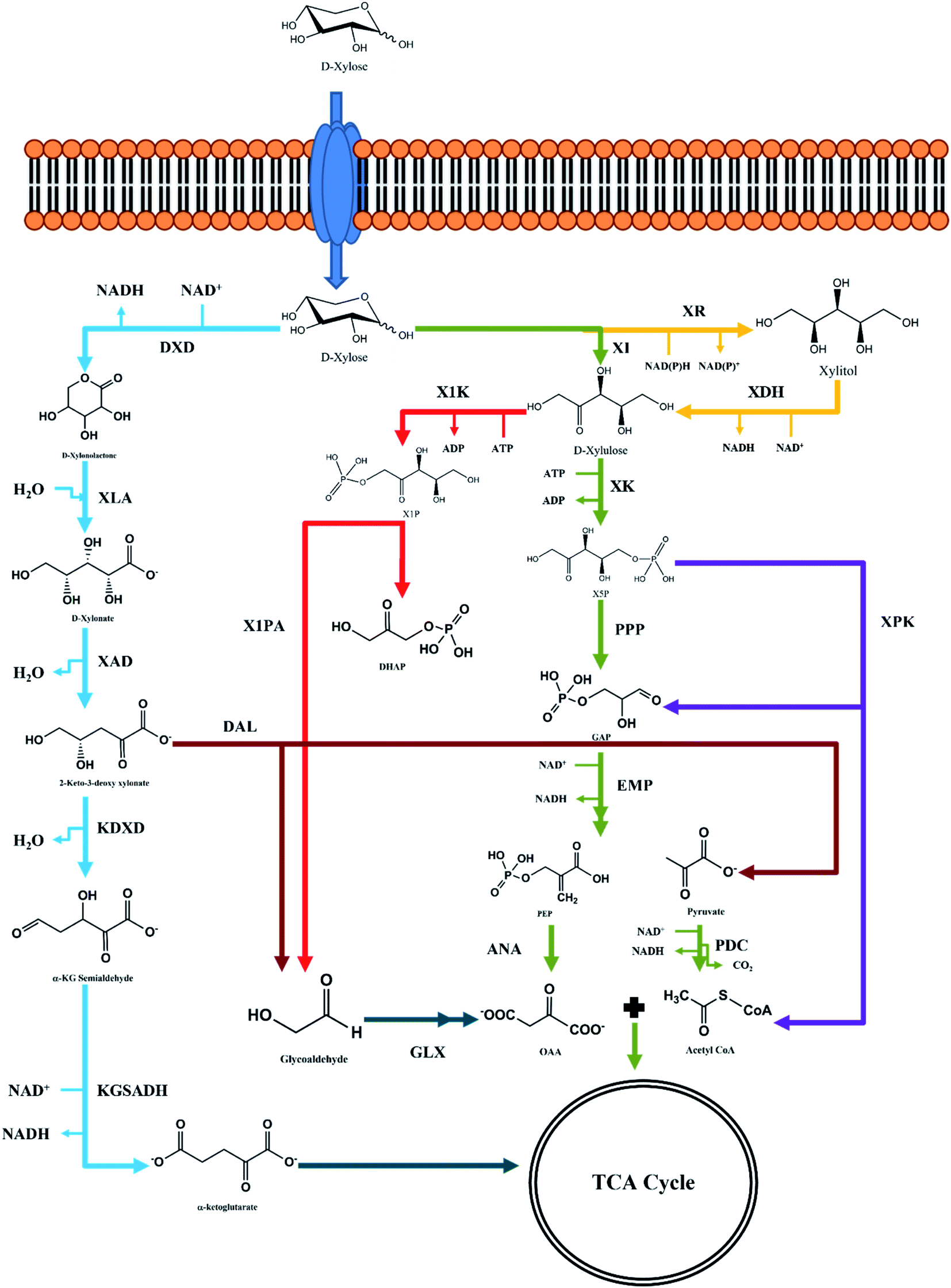 | ||
| Fig. 3 Illustration of xylose transport inside the microbial cell and further dissimilatory pathways directing the carbon flux into central carbon metabolism. Abbreviations: DXD: xylose dehydrogenase; XLA: xylonate synthase; XAD: xylonate dehydratase; KDXD: 2-keto-3-deoxyxylonate dehydratase; KGSADH: ketoglutarate semialdehyde dehydrogenase; XI: xylose isomerase; XR: xylose reductase; XDH: xylitol dehydrogenase; X1K: xylulose-1-kinase; X1PA: xylulose-1-phosphate aldolase; DAL: 2-keo-3-deoxyxylonate aldolase; GLX: glyoxylate shunt; XLK: xylulose-5-kinase; XPK: phosphoketolase; PPP: pentose phosphate pathway; EMP: Embden–Meyerhof–Parnas pathway; Weimberg pathway (blue); isomerase pathway (green); Dahms pathway (brown); XR–XDH pathway (yellow); phosphoketolase pathway (violet); synthetic pathway (red). | ||
Xylose isomerases (EC. 5.3.1.5) (D-xylose → D-xylulose) encoded by the XylA gene are metal dependent enzymes classified into two different classes, I and II. These two enzyme classes differ in length of the polypeptide chain, where class II enzymes have an additional 34 amino acid residues on the N-terminus compared to class I. The catalytic activity of XI is conserved at two sites of histidine residues H101 and H271 and induced in the presence of xylose. XI mediates the synthesis of xylulose via a hybrid shift mechanism for ring opening to form an open chair conformation.44 The substrate binding at the active site was observed by fluorescence quenching at two conserved regions W29 and W188 with the tryptophan residue at W29 being essential for catalytic activity. The genome mining and sequencing in the thermophilic Bacillus coagulans strain identified that the XylA gene consisting of 1338 base pairs encodes for 50 KDa class II protein with 445 amino acids. The amino acid identity of B. coagulans XI gave a homology of 65, 64, 58, 48 and 25% with Lactobacillus brevis, L. pentosus, L. lactis, Piromyces sp. E2, and Streptomyces albus, respectively.45 Thermostable XI with maximum enzyme activity at 85 °C and neutral pH was isolated from thermophilic strains like Thermoanaerobacterium ethanolicus.46 Similarly, the Streptomyces sp. F-1 strain, a new isolate, has two copies of XylA genes, and the biochemical characterization presented a significant difference in their optimal temperature. The protein coded from XylA1 and XylA2 displayed maximum activity at 60 and 75 °C, respectively.47 The structural characteristics and enzyme kinetics of XI are well investigated. An interesting feature of XI is that the enzyme operates with high activity within a broad temperature range of 30–85 °C. However, it is sensitive to pH change and the maximum specific activity was observed at a physiological pH range of 6.0–8.0 which declined rapidly under strong acidic or alkaline conditions.46,48 It was also observed that the divalent metal ions are a pre-requisite for the activation and stabilization of the enzyme activity. The presence of Mg2+, Co2+, or Mn2+ has a profound positive effect compared to other divalent metal ions, while Ni2+ has been found to be inhibitory for the enzyme. These metallic cofactors also protect the enzyme from thermal denaturation.45
Xylulokinase (XK) (EC 2.7.1.17) is a substrate (D-xylulose) specific kinase enzyme catalysing the phosphorylation reaction D-xylulose + ATP → D-xylulose-5-phopsphate + ADP. XK in B. coagulans is a 56 kDa protein consisting of 1536 bp with 511 amino acids. The amino acid identity of XK from B. coagulans revealed a sequence homology of 56, 49, 38 and 25% with L. pentosus, L. lactis, E. coli, and Scheffersomyces stipitis, respectively. However, the homology between xylulokinase of B. coagulans and L. brevis was only 19%. During the activity measurement at different pHs, the maximum activity of XK was observed at an optimal pH and temperature of 7 and 85 °C, respectively, while the enzyme lost 20% and >50% of the activity when the pH was reduced to 6.0 or increased to 8.0, respectively. Similar to XI, the divalent ions Co2+, Mn2+, and Fe2+ enhanced the activity of XK.45
Xylose reductase (XR; EC 1.1.1.21) [D-xylose + NAD(P)H → xylitol + NAD(P)+] is a 36 KDa protein containing 322 amino acids and is a member of aldoketoreductase family 2 (AKR2). AKRs are superfamilies of enzymes that catalyse the reversible reduction of aldehydes or ketones to their respective alcohols utilizing NADPH as a cofactor. XR is a highly important enzyme when the desired product is xylitol, a molecule with nutritional and pharmaceutical value. Son et al. (2018) reported the crystal structure of XR from S. stipitis. XR is a dimer with two polypeptide chains made of 15 α-helices and 10 β-strands each with conserved catalytic sites at Asp43, Tyr48, Lys77 and His110.51 The literature describes the flexibility of XR in using NADPH as well as NADH as a cofactor. The Km values of NADPH and NADH for XR in S. stipitis are 0.0277 and 0.136 mM, respectively, indicating more affinity and specificity for NADPH than NADH. Although the physiological function of XR is to reduce D-xylose, the Km value of D-xylose for XR is very high (39.4 mM), indicating that a high level of xylose is needed to drive xylose metabolism efficiently. The structural conformation of the enzyme displays the presence of a hydrophobic binding pocket. It could be one of the reasons for the low affinity of XR towards xylose which has a high degree of hydrophilicity due to the presence of five hydroxyl groups. Similar to S. stipitis, a dimeric XR structure with Km of 87 mM for D-xylose has been elucidated in Candida tenuis.56 Recently, new xylose utilizing yeasts of Spathaspora sp. were characterized to have high XR activity which were mostly NADPH dependent, except three species, Sp. arborariae, Sp. gorwiae, and Sp. passalidarum.57,58 The genome mining resulted in two putative XR genes, where SpXYL1.2 has relatively higher XR activity with NADH, and the strain could assimilate xylose effectively under anaerobic conditions. In Sp. arborariae, XR accepts NADH and NADPH as cofactors, with an affinity (Km) of 12.8 (NADH) and 26.1 (NADPH) μM, respectively. In the presence of xylose as a substrate, the affinity was observed to strengthen with Km of 29.5 (NADH) and 57.5 (NADPH) mM, respectively.53,57
Xylitol dehydrogenase (XDH; EC 1.1.1.9) or xylulose reductase (xylitol + NAD+ → D-xylulose + NADH) mediates the conversion of xylitol to D-xylulose and is a well characterized enzyme encoded by a nucleotide sequence of 1089 bp, and the operon reading frame codes for a protein containing 363 amino acids, with an approximate mass of 38.5 KDa. The XDH mediates the oxidation of xylitol using NAD+ as a cofactor.59,60 The NAD+-dependent XDH is a homotetramer which forms heteronuclear multi-metal protein with 1 mol of Zn2+ and 6 mol of Mg2+ ions per mol of 37.4 kDa protomer (structural subunit of an oligomeric protein) with Km of 39 μM for xylitol. The XDH enzyme displayed a half-life of 300 h in 50 mM Tris buffer at pH 7.5. Metal ions like Co2+, Mn2+, and Zn2+ exert an inhibitory effect on the enzyme and the activity completely ceases at 5.0 mM concentration of these metal ions. But complete dissociation of Zn2+ from the enzyme was observed to inactivate XDH completely.61
The BLAST analysis of the P. fragi genomic database revealed that Caulobacter crescentus, Burkholderia xenovorans, and Chromohalobacter salexigens have possible genes mediating the Weimberg pathway. In 2007, Craig Stephens and associates observed the expression of XylXABCD genes, when a freshwater bacterium, C. crescentus, was grown on D-xylose as the sole carbon and energy source. Recently, it has been found that Pseudomonas taiwanensis VLB120 can assimilate D-xylose through the Weimberg pathway, but the initial oxidation and hydrolysis reactions are mediated by glucose dehydrogenase (EC 1.1.5.2) and gluconolactonase (EC 3.1.1.17) instead of DXD and XLA. Similar behaviour was also observed in P. putida strains wherein DXD and XLA are involved in gluconic acid production.65 Further, determination of kinetic parameters Km and Vmax revealed two rate limiting reactions in the Weimberg pathway mediated by Mn2+ dependent XDH and NAD+ dependent KGSADH.66 Therefore, D-xylose assimilation by the Weimberg pathway requires external supplementation of metal ions (Mn2+) and availability of NAD+ for complete conversion of D-xylose into α-ketoglutarate.
3.2 Xylose operon
The genes responsible for the xylose transport and assimilation are clustered into an open reading frame called a xylose (xyl) operon. In bacteria, xylose metabolism is mediated through the XI pathway.71 The genes responsible for the metabolism of xylose were observed to be organized into two major transcriptional units XylAB and XylFGHR with promoters PA and PE, respectively. It was observed that the transcriptional activation is induced by xylose and repressed by glucose, i.e. as long as glucose is available, the xylose assimilation will be suppressed.72 The transcriptional units XylAB and XylFGHR were observed to be located at 80 min on the chromosome map.73XylAB consists of genes encoding for XI (XylA) and XK (XylB), respectively. XylFGHR, a high affinity ABC type transporter for transport of xylose, consists of four subunits where XylH acts as the transmembrane transporter, XylF binds to xylose and XylG is an ATP binding protein that mediates the phosphorylation of xylose. Subsequently, the transported xylose is acted upon by XI and XK, to form xylulose-5-phosphate. The XylR gene has been observed to be constitutively expressed under a weak promoter PR regardless of xylose or glucose availability. The repressor tends to bind two DNA binding regions IA and IE, upstream to the transcriptional promoter's consensus sequences and adjacent to the RNA polymerase binding site. In the presence of xylose, XylR forms a dimer with the xylose substrate, and causes activation of two promoters PA and PE, resulting in simultaneous transcription of XylFGH and XylAB genes. Any mutation in the repressor protein XylR was observed to abolish the expression of PA and PE promoters.74,75 In E. coli the xylose transport into the microbial cell is mediated by low affinity transporter xylE, and the expression was observed to be 10-fold higher when the external medium was supplemented with xylose.76 Though the xylose dissimilation follows the XI pathway in Gram-positive bacteria, the regulation was observed to be different in few enteric bacteria like B. subtilis and Lactobacillus strains, where the XylR gene displays repressive behaviour rather than acting as a transcriptional activator. The XylR of B. subtilis and Lactobacillus strains is not homologous to its counterpart in E. coli and binds to a palindromic sequence upstream to the transcriptional start codon, repressing the transcription activation and the repression effect is relieved in the presence of xylose.77 In Staphylococcus xylosus, three open reading frames containing 4520 nucleotide bases were annotated as XylR, XylA and XylB genes. The BLAST studies of the XylA gene presented 65% and 51% similarity with B. subtilis and E. coli, respectively.78In the genera Clostridia, C. acetobutylicum is the representative strain for investigating different metabolic activities. The whole genome sequence is available along with the required genetic tools for strain engineering. Genome mining through a subsystem-based approach revealed the presence of a novel XI (CAC2610), and further characterization indicated that it is not homologous to known XI (XylA) genes. Along with XI, XK (XylB, CAC2612), a xylose proton symporter (XylT, CAC1345), and a transcriptional regulator (XylR, CAC3673) were also sequenced and characterized. The C. acetobutylicum strains are well known to utilize a broad range of monosaccharides, disaccharides, starches, and other polysaccharides like xylan and xyloglucan. Xylan and xyloglucan are the major components of the hemicellulosic fraction of plants. The depolymerization of xyloglucan and xylan results in α- and β-xylosides, respectively. These xylosides are transported into the cell and further degraded into xylose. The genetic make-up for utilizing these xylosides was mainly observed in firmicutes like Bacillus, Lactobacillus and Clostridium spp. In B. subtilis and C. acetobutylicum, xylose operon contains two clusters, XylAB and XynTB. The XynT gene encodes the ABC transporter that transports β and α-xylosides into the microbial cell and further the XynB gene converts xyloside into D-xylose.73,79
In yeast and fungi, xylose is sequentially metabolised through three cytosolic enzymes, XR, XDH and XK, to convert it to xylulose-5-phosphate. S. stipites the most efficient xylose fermenting yeast was characterized to reveal the genes encoding XR (Xyl1), XDH (Xyl2), and XK (Xyl3) enzymes. The genes are either co-localized or distributed in the genome; for example in S. stipitis, Xyl1 was observed on chromosome (Chr) V, Xyl2 on Chr I, and Xyl3 on Chr VIII. Although S. cerevisiae strains are not native xylose utilizing strains, a putative XDH (Xyl2) gene was identified on Chr XV, and XK on Chr VII.
3.3 Carbon catabolite repression or the glucose effect: the natural phenomenon arresting the simultaneous conversion of mixed sugars
Microorganisms cultured on mixed sugar substrates display a pattern of two successive exponential phases during growth, called diauxic growth. The occurrence of this growth pattern is due to utilization of the preferred substrate which suppresses the uptake of other carbon sources present in the medium and this phenomenon is known as carbon catabolite repression (CCR) or the glucose repression effect.80,81 The diauxic growth significantly affects the utilization of mixed sugars and increases the length of fermentation (decreased productivity).LCB or agricultural residues as the feedstock for the production of biofuels and bioproducts has received considerable interest.6 It is not just surplus agro-residual biomass, utilizing LCB as feedstock addresses various environmental concerns and the food vs. feed debate with first generation starchy feedstocks. As LCBs are polymers of celluloses and hemicelluloses, hydrolysates derived after pretreatment and saccharification contain a mixture of hexoses (mostly glucose) and pentoses (mostly xylose).41,80 The growth of microorganisms on hydrolysates containing mixed sugars results in suppression of pentose sugar utilisation. The mechanism and the strategies to overcome the limitation are discussed in this section. Aidelberg and co-workers observed a hierarchical fashion of utilization of hexoses and pentoses. When the microorganism is grown on mixed sugars (glucose, arabinose and xylose) as substrates, the most preferred carbon source has an inhibitory effect on other sugars; for example, glucose represses the uptake of arabinose and xylose, and upon glucose depletion, the next preferred substrate is arabinose, and the xylose utilization mechanism is still inhibited.82 From the literature, it was explained that the mechanism occurs due to two reasons: (i) inhibition of expression of the genes involved in the non-glucose sugars by 3′,5′-cyclic adenosine monophosphate (cAMP). cAMP is a secondary messenger, derivative of ATP, synthesized by an adenylate cyclase enzyme. In bacteria, cAMP levels depend on the type of growth medium. Intracellular transportation of glucose inhibits the adenylate cyclase enzyme and decreases cAMP and cAMP receptor protein (CRP), which inhibits the transcription of the xyl operon;83 (ii) inhibition of xylose transport mediated by dephosphorylated PTC component EIIAglc that binds to the cognate sugar transporter and prevents the transport, and the mechanism is inducer exclusion (Fig. 4A).84 However, in the phosphorylated form, the EIIAglc component activates the adenylate cyclase, which improves intracellular cAMP levels. Improved cAMP levels bind with CRP to form a complex, and the active cAMP–CRP complex could bind to the ORF and express the permeases and other genes involved in the metabolism of non-glucose sugars (Fig. 4B).85
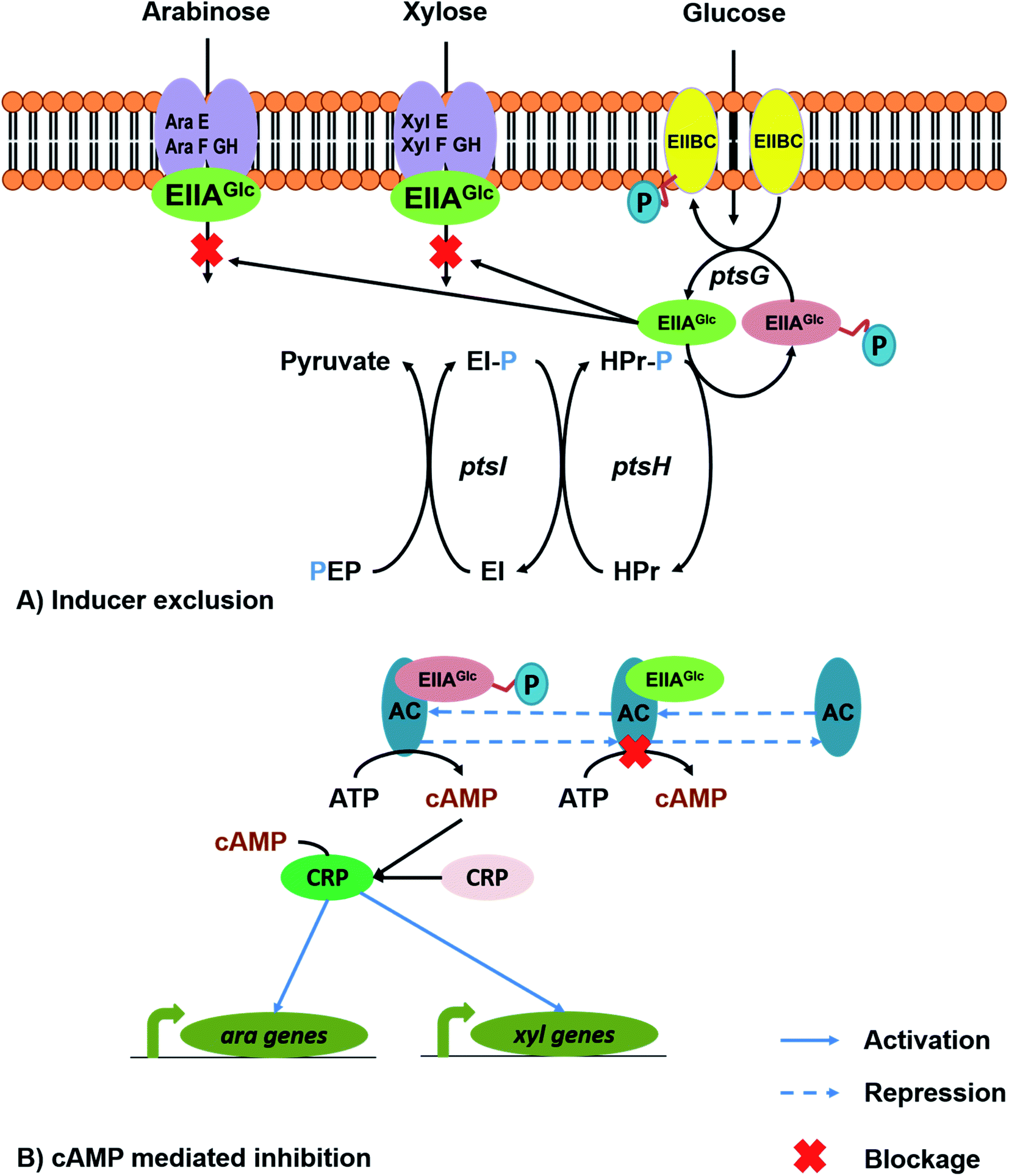 | ||
| Fig. 4 Illustration of possible mechanisms for carbon catabolite repression: (A) inducer exclusion and (B) cAMP mediated inhibition. | ||
In B. megaterium, the glucose mediated xylose repression was 14-fold, and the XylR gene which regulates the transcription initiation by binding to promoters of XylAB and XylFGH was modified by incorporating a kanamycin resistance gene resulting in lowering the repression to 8-fold, and deletion of 184 bp at the 5′-end of the XylR gene further reduced repression by 2-fold.85,86 An alternative CCR mechanism called feedback inhibition was observed in a few Gram-positive bacteria, in which catabolite control is exerted by catabolite control protein A (ccpA). The ccpA is a dimeric transcriptional regulator, expressed constitutively regardless of the carbon source. In the presence of glucose and other glycolytic intermediates like fructose 1,6-bisphosphate, the histone protein (HPr) component of the enzyme phosphorylation cascade (PTS enzyme I, HPr, and Enzyme II), the major facilitator of sugars, is phosphorylated at the serine residue (HPr-Ser46-P) instead of the histidine residue. The phosphorylated HPr binds to catabolite control protein (ccpA), and the complex binds to catabolite repressive element (CRE) within the transcriptional or coding sequence upstream of the promoter region by blocking the transcription of pentose sugars.87 In C. acetobutylicum, deletion of ccpA and the enzyme II complex was attempted but it resulted in an impaired growth rate and failure in metabolic flux.87 But in B. subtilis, deletion of CRE protein could overcome the degree of repression from 13- to 2.5-fold.88 The successful development of the CCR negative strain would be of high value with the capacity to assimilate glucose and non-glucose sugars simultaneously causing an increase in the yield and productivity.
4. Hurdles requiring debottlenecking for efficient xylose metabolism
4.1 Transport of xylose into the microbial cell
Xylose metabolism is well investigated and characterised in bacteria, fungi, yeast, and few archaea. In these prokaryotic and eukaryotic organisms, the xylose metabolism is either native or heterologously expressed to shape the cellular metabolism to rely on xylose as a sole carbon and energy source. Before the start of xylose metabolism, the sugar must be transported into the microbial cell (Fig. 5). The possible mechanisms for transport of sugars through the membrane are passive diffusion, facilitated diffusion and active transport.89 Passive diffusion is the simplest process that occurs based on the concentration gradient of the substrate between the intracellular environment and the extracellular medium, while in facilitated diffusion a carrier protein mediates the transfer based on the concentration gradient. In active transport, the translocation of sugar through the transmembrane proteins happens with energy expenditure. Usually, transmembrane proteins that span across the outer membrane mediate the translocation from the extracellular space to the intracellular environment. These transmembrane proteins belong to the major facilitator superfamily (MFS), and are divided into three classes based on the functionality, (i) uniporters, which transport a single substrate across the membrane; (ii) symporters, which transport one substrate coupled with a charged molecule; and (iii) antiporters, by which two different substrates are translocated in the opposite directions.90 | ||
| Fig. 5 Transport of xylose into the microbial cell and further flux into central carbon metabolism. | ||
In bacteria there are three possible mechanisms known for the transport of xylose into the microbial cell, (i) the H+/Na+-symporter, which is identified in E. coli,91Salmonella typhimurium, B. megaterium, L. brevis, and B. subtilis, (ii) PEP:carbohydrate phosphotransferase system, which is identified in E. coli and uses PEP as the source of energy, and (iii) ATP driven ABC transportation periplasmic binding protein, identified in E. coli and few other Bacillus spp. Facilitated diffusion is not well known in bacterial populations, except the glycerol facilitator protein (GlpF) in E. coli and glucose transporter (GlfZ) in Zymomonas mobilis. In lactic acid bacteria, the phosphoenol pyruvate (PEP) D-mannose phosphotransferase system (PTS) with two integral membrane proteins EIICMan and EIIDMan, and cytoplasmic phosphorylation proteins EIIAMan and EIIBMan is observed to have a significant role in xylose transport.89 The endogenous transporters in the microbial cells have an affinity for glucose from low (Km 50–100 mM) to high (Km 1–2 mM), whereas for xylose the numbers can be up to 10 times higher.92 For example, Glf, a promiscuous glucose-facilitated diffusion protein from Z. mobilis expressed in E. coli, has lower affinity towards xylose (Km 40 mM) than glucose (Km 4.1 mM), and Vmax was observed to be two-fold higher for glucose, resulting in delayed xylose uptake and assimilation, when expressed in E. coli cells.91
There are two mechanisms identified in yeast for xylose transport, (i) membrane potential due to proton symport, or (ii) facilitative diffusion through low affinity transporters. Scheffersomyces stipitis, a well-known xylose metabolizing strain has low and high affinity carriers to mediate xylose transport, whereas in C. shehatae, facilitated diffusion and a low affinity symport mechanism was observed.93,94 The sugar binding pockets in the transmembrane proteins have residues specific for sugar, and any alteration or changes in the glucose specific residues can alter the D-glucose uptake rate and increase the endogenous xylose transport. Farwick et al. (2014) implemented this method in altering the amino acid sequence which interacts with the C6-hydroxymethyl group of D-glucose, but the deletion of those specific amino acids led to deleterious effects on the transport of glucose as well as xylose.92 Wang and associates have studied the effect of 28 different site directed mutations on the xylose uptake rate and metabolism in the Mgt05196p transporter of Meyerozyma guilliermondii. The substitution mutations at Phe432Ala and Asn360Ser on Mgt05196p improved the xylose uptake, but diminished the glucose uptake, whereas the N360F mutation specifically enhanced the xylose transport without any glucose inhibition.95
Evolution is the natural mechanism of adapting to new environmental or physiological conditions. To create an order of natural evolution, a new approach called adaptive laboratory evolution (ALE) was attempted to increase the efficiency of xylose transporters XUT3 and GXS1 from S. stipitis and C. intermedia, respectively.96 The XUT3 is one of the seven high affinity xylose (XUT) transporters annotated in S. stipitis with similar specificity towards both glucose and xylose. The GXS1 in C. intermedia is a broad range sugar transporter with specificity to carbon sources like glucose, arabinose, ribose, and xylose.96 In this directed evolutionary approach, the substitution mutations Phe40Val, Phe465Ser, and Phe500Ser in GXS1, and Leu122Val, Phe343Leu, Gln345Arg, Ala298Thr, Tyr304Phe, and Lys542Arg in XUT3 influenced the xylose uptake. The heterologous expression of these mutated transporters in S. cerevisiae resulted in a 70% increase in the specific growth rate on xylose.96 The mutant transporters also displayed a phenomenal alteration in the diauxic growth and the evolved strain could simultaneously utilize xylose and glucose. In the sugar transporters, motif G–G/F-XXX-G is a conserved sequence present on the transmembrane component. After site-directed mutagenesis or ALE, a modified motif GGFIMG with larger side chains restricting the pore size for glucose transport and allowing smaller xylose molecules was identified. The alteration in the motif sequence because of point mutations increased the pentose specificity to the binding site by decreasing the pore size and transporting xylose efficiently compared to glucose.97 However, bioprospecting for a novel xylose specific transporter or modification of an existing transporter to overcome the CCR induced by glucose will be beneficial and could significantly improve simultaneous glucose and xylose consumption.
4.2 Availability of redox cofactors and homeostasis
Redox homeostasis is an important consideration in microbial cell factories as it affects a wide range of genes, cellular functions and metabolite profiles, and redox balancing plays a critical role in coupling catabolism and anabolism. The co-factors involved in maintaining homeostasis are NADH and NADPH, which usually act as electron carriers and are involved in respiratory chain reactions (catabolism) and cell synthesis (anabolism), respectively. NADH is the predominant redox product of catabolism while NADPH has a greater role in anabolism with a major fraction coming from the pentose phosphate pathway and a delicate balance in the intracellular level of these cofactors is required to ascertain an optimal metabolic output. The NADH/NAD+ ratio which reflects the intracellular redox state of a living cell and is influenced by various factors such as the physiological state of the cell, oxidation state of the substrate, the nature and presence of electron acceptors, and enzymes requiring redox factors.98–101 The cells often start side reactions leading to byproduct formation which contributes towards the redox homeostasis. For example, during ethanol fermentation by S. cerevisiae, the yeast starts parallel formation of acetate and glycerol, NADH consuming reactions.In the case of change in the substrate from glucose to xylose, a myriad of changes were observed in the metabolism, and the responses observed were increased amino acid concentrations, increased TCA cycle intermediates, and reduction in sugar phosphates and reducing equivalents or redox cofactors.102 One of the challenges with xylose metabolism is maintaining redox homeostasis. After the first 3–4 specific steps of xylose metabolism, it is connected to central carbon metabolism. The majority of this problem stems from the first two steps where xylose is isomerised to xylulose via XR and XDH. The higher preference of XR towards NADPH generates NADP+ while the second step requires NAD+. The diminished synthesis of reducing equivalents and uneven demand during the xylose assimilation results in cofactor imbalance, which in turn affects the ATP yield and metabolic fluxes. The different cofactor preference of these two enzymes results in NAD+ deficiency resulting in accumulation of the intermediate xylitol. The condition mostly prevails under anaerobic or oxygen limited conditions, where NADH cannot be oxidized to NAD+, due to the absence of oxygen, the final electron acceptor.103 This problem could be overcome by a continuous supply of NADPH and NAD+.
To prevent the xylitol accumulation and further direct the flux of xylose carbon to central carbon metabolism, NADH oxidase (NOX) can be used. NOX catalyses a water forming reaction using oxygen as the electron acceptor (NADH + H+ + 0.5O2 → NAD+ + H2O), thereby regenerating the NAD+ molecules. The xylose assimilation through the XR–XDH pathway linked with NADH oxidase could enable redox homeostasis. Zhang et al. (2012) constructed a cycle of regeneration using NOX, for regeneration of NAD+.104 In a similar approach, NOX from L. lactis was heterologously overexpressed in S. cerevisiae harbouring XR–XDH from S. stipites which resulted in a 69.6% decrease in xylitol accumulation, and more carbon flux was directed towards ethanol leading to an improvement of 39.3% in molar yields.105
Scheffersomyces stipitis has the ability to ferment xylose under anaerobic conditions, and thus under unfavourable conditions such as redox imbalance, the accumulated NADH can be utilized by NADH-dependent XR and circumvents the pathway.50 In yeasts such as S. stipitis, XR has affinity for NADH as well as NADPH, and hence using advanced genetic engineering techniques, the cofactor specificity of XR in the required host strain can be altered, so that the cofactor requirement for the first two steps can be compensated internally and the continuous availability of cofactors can lead to improved xylose uptake and fermentation efficiency. A mutant XR enzyme K270M from S. stipitis with lower specificity to NADPH was expressed in S. cerevisiae and the cultivation of the strain with mutated XR on xylose showed a 16-fold reduction in NADPH and 4.3-fold increase in NADH specificity.93 After a site directed mutagenesis approach on the XR enzyme in C. tenuis, a 170-fold change in cofactor preference from NADPH to NADH was observed in a Lys274Arg and Asn276Asp double mutant. When the mutant XR along with XDH from Galactocandida mastotermitis was expressed in S. cerevisiae, a 42% increase in ethanol, and a 52 and 57% decrease in xylitol and glycerol yields was observed, respectively, with xylose as the sole carbon source.106 Similarly, a double mutant of S. stipitis XR (Arg276His and Lys270Arg/Asn270Asp) showed a decrease in catalytic efficiency and increase in Km values towards NADPH, resulting in enhanced XR dependence on NADH. The strain expressing NADH-dependent XR efficiently utilized xylose, resulting in a 20% increase in the ethanol level and 52% decrease in xylitol accumulation.107 A wild type NADH specific XR was identified from C. parapsilosis, in which the conserved motifs have arginine residues instead of lysine. Later, the structural integrity of NADH specific XR of S. stipitis was altered (Lys270Arg), and the S. cerevisiae strain expressing this modified XR diverted the flux of carbon towards ethanol with reduced xylitol accumulation.108
Besides overexpression of NOX and change in cofactor preference, a number of other approaches have been employed to alleviate the problem of redox imbalance. Under anaerobic conditions, NADH molecules are oxidized through a transhydrogenase shunt with malic enzyme (MAE1) (malate + NADP+ → pyruvate + NADPH), malate dehydrogenase (MDH2) (oxaloacetate + NADH → malate + NAD+), and pyruvate carboxylase (PYC2) (pyruvate + ATP → oxaloacetate + ADP) that can regulate the redox balance in S. cerevisiae. In a combinatorial cassette along with xylose metabolizing (XR, XDH, and XK) genes, two different strains were constructed. Strain 1 expressing MAE1 with Xyl genes was observed to improve the xylose uptake and caused an increment in the NADPH/NADP+ ratio. The co-expression of MAE1 and MDH2 along with Xyl genes resulted in a 1.25-fold increase in ethanol titers due to regeneration of cofactors required for the 1st and 2nd steps of the XR–XDH pathway.109 Alternative to multiple gene overexpression, a native NADH kinase (NADH + NADP+ → NADPH + NAD+) enzyme was overexpressed replenishing the NADPH and NAD+ cofactors.110 In another study, NADPH dependent glutamate dehydrogenase (GDH1) (α-ketoglutarate + NH4+ + NADPH → glutamate + NADP+) was deleted in S. cerevisiae and NAD+ dependent glutamate dehydrogenase (glutamate + NAD+ + ATP + H2O → α-ketoglutarate + NH4+ + NADH + ADP) was overexpressed resulting in increased ethanol production and reduced xylitol accumulation.111 Recently, two NADH oxidation approaches were demonstrated in L. lactis, where external supplementation of hemin112 and flavinium113 catalyses the oxidation of NADH molecules in the presence of O2. Although the mechanism was demonstrated in L. lactis, this in situ regeneration of reducing cofactors could be of wide significance from an industrial perspective.
Like NADH, NADPH is also a crucial electron donor in various metabolic pathways. Celton and associates reported that S. cerevisiae cells growing on pentose sugars respond to an increase in NADPH demand by directing the carbon flux through the pentose phosphate pathway (PPP) and acetate synthesis pathway, as well as transforming NADH to NADPH in the cytosol via the transhydrogenase cycle. The enzymes involved in the regeneration of NADPH and NAD+ cofactors are glucose-6-phosphate dehydrogenase (Reaction 1), 6-phosphogluconate dehydrogenase (Reaction 2), and transhydrogenase (Reaction 3).
| Glucose-6-phosphate + NADP+ = 6-phosphoglucono-δ-lactone + NADPH(PPP) | (1) |
| 6-Phosphogluconate + NADP+ = ribulose-5-phosphate + NADPH(PPP) | (2) |
| NADH + NADP+ = NAD+ + NADPH (transhydrogenase cycle) | (3) |
In the case of higher xylose concentrations and increased uptake, the demand for NAD(P)H will be further increased, a predicted glycerol–DHA cycle has been reported to exchange NADH and NADP+ for NAD+ and NADPH, at the expense of an ATP molecule. In the glycerol–DHA cycle, dihydroxyacetone phosphate (DHAP) is reduced to glycerol by NADH-dependent glycerol-3-phosphate dehydrogenase (Reaction 4), glycerol is then oxidised to dihydroxyacetone (DHA) by NADP+ dependent glycerol dehydrogenase (Reaction 5) and finally DHA is phosphorylated to DHAP at the expense of one ATP molecule (Reaction 6).114 Thus, the glycerol–DHA cycle generates both the redox factors (NADPH and NAD+) required for the XR–XDH pathway.
| DHAP + NADH = glycerol + NAD+ | (4) |
| Glycerol + NADP+ = DHA + NADPH | (5) |
| DHA + ATP = DHAP + ADP | (6) |
| NADH + NADP+ + ATP = NAD+ + NADPH + ADP | (7) |
5. Xylose as an alternative carbon source for microbial growth and product development
The bioconversion of xylose into value-added chemicals has received a lot of attention in recent years. Many naturally occurring or engineered microbial strains have been discovered or designed to synthesize various industrially important chemicals and fuels using xylose as a sole carbon and energy source. Table 3 summarizes various chemicals that can be produced from xylose through the biological route and their commercial applications.| Chemical | Commercial applications |
|---|---|

|
Solvent, automotive gasoline, alcohol beverages, distilled spirits, hand sanitizers and medical antiseptics |
|
Polymeric monomers, paints, adhesives, inks, coatings, and food additives |
|
Plastics, cosmetics, and solvents |
|
Food, beverages, polyesters, textiles, and pharmaceuticals |
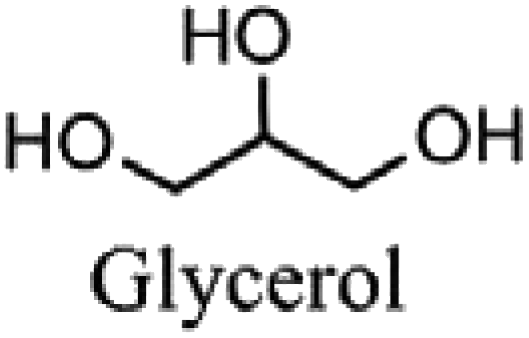
|
Pharmaceutical, food, polymer, humectant, solvent, lubricant, personal care, and household products |
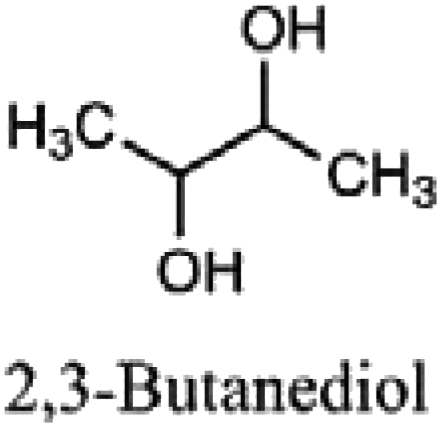
|
Polymers, solvents, fine chemicals, lactones, fuel additives |
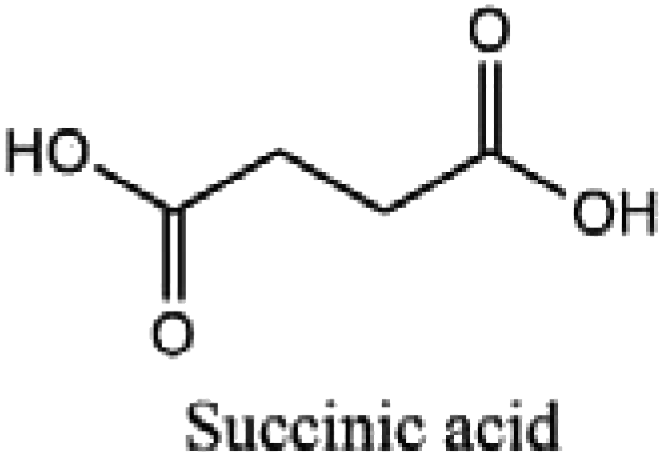
|
Pharmaceutical products, surfactants, detergents, plastics and food grade ingredients |

|
Lubricants, brake fluids, synthetic rubber, polymers, and automotive fuels |
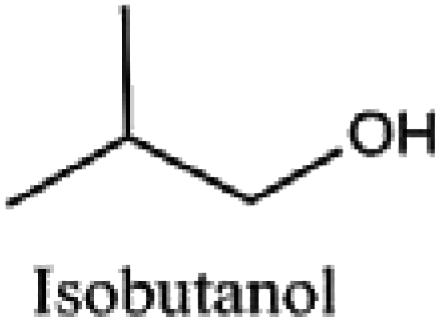
|
Coatings, chemical derivatives, paints, fuel additive, and solvents |
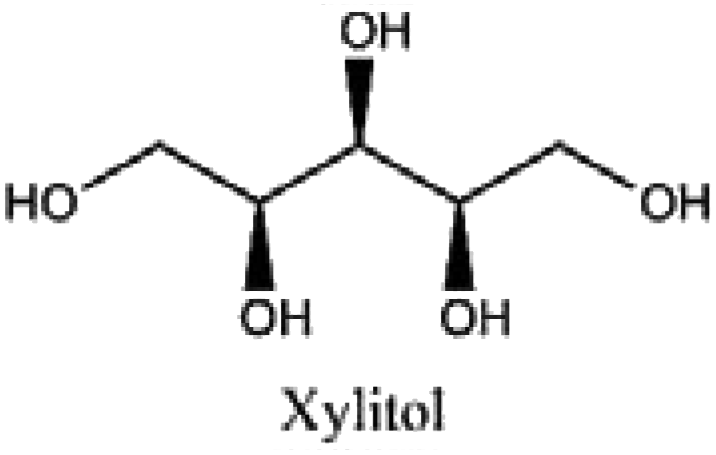
|
Confectionery, chewing gums, syrups, and odontological and pharmaceutical products |
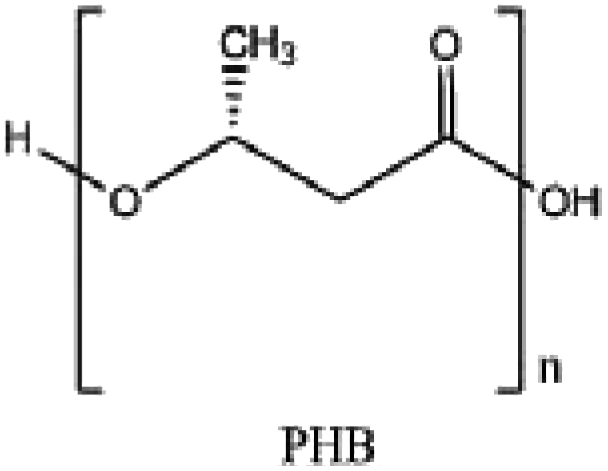
|
Biodegradable plastics |
5.1 Xylitol
Xylitol (C5H12O5), a platform chemical, is a five-carbon sugar alcohol with a wide spectrum of applications in personal care, food, confectionary, and pharmaceutical industries.115 Xylitol is equivalent to common table sugar with a lower calorific value (2.4 vs. 4 calories per gram), lower glycaemic index (7 vs. 60–70%) and insulin independent metabolism. In 2016, the global xylitol market was worth US$ 725.9 million with a production capacity of 190.9 thousand metric tons. It has been forecasted that with the increased global market demand and compound annual growth rate (CAGR) of 5.7%, the production capacity should be increased to 265.5 thousand metric tons (US$ 1 billion) by 2022.116 The commercial production of xylitol is performed via a chemical route by catalytic dehydrogenation of pure xylose, involving expensive Ni based catalyst, sulfuric acid, calcium oxide, phosphoric acid and activated charcoal treatments at high pressure (5000 kPa) and temperature (140 °C).117 The process is uneconomical due to the requirement of pure xylose as the substrate, the process generates heavy metal pollutants, and there is a high risk associated with operating conditions and environmental pollution.117 An alternative route for the production of xylitol is the biological process, wherein whole/immobilized cells expressing XR or cell-free extracts with XR activity act as biocatalysts (Fig. 3), producing xylitol from pure and crude renewable sources rich in xylose. The process offers the advantages of mild operation conditions and non-requirement of purified xylose.118Xylose is a native substrate for xylitol which is accumulated due to lack of synchronisation between steps catalysed by XR and XDH. Various microorganisms having natural xylitol producing ability include bacteria, yeast, and fungi. Among them yeasts are predominant such as Candida athensensis, C. boidinii, C. guilliermondii, Debaryomyces hansenii, C. tropicalis, C. magnolia, and S. stipitis that can accumulate xylitol with significant yields and productivities (Table 4). Later with the introduction of heterologous pathway engineering, S. cerevisiae, Kluyveromyces and other Candida spp. have been engineered to accumulate xylitol. Usually bacterial systems present the XI pathway for xylose assimilation, but a few bacterial strains like Bacillus coagulans, Cellulomonas cellulans, Corynebacterium glutamicum, Corynebacterium ammoniagenes, Enterobacter liquefaciens, Mycobacterium smegmatis, and Serratia marcescens present the XR–XDH pathway for producing xylitol.119 A new bacterial isolate, Pseudomonas putida, was screened for xylitol production and characterized to have XR activity of 48.7 IU mg−1. The strain accumulated 35.2 g L−1 xylitol with a productivity of 0.98 g L−1 h−1 when cultured on xylose under optimized growth conditions.118
| Product | Pretreatment | Form of substrate | Additional steps prior to fermentation | Microorganism | Mode of fermentation | Titer (g L−1) | Yield (g g−1) | Productivity (g L−1 h−1) | References |
|---|---|---|---|---|---|---|---|---|---|
| a SCB: sugarcane bagasse; CC: corn cobs; CS: corn stalk; EG: elephant grass; SHF: separate hydrolysis and fermentation; SScF: simultaneous saccharification and fermentation; GM: genetically modified. | |||||||||
| Xylitol | Hydrothermally pretreated SCB | Xylose-rich hydrolysate | None | Pichia fermentans | Batch | 79 | 0.67 | 0.47 | 55 |
| HNO3 pretreated CC | Xylose-rich hydrolysate | Detoxification & decolorization | Candida tropicalis MTCC 6192 | Batch | 34.35 | 0.62 | 0.26 | 213 | |
| Pure xylose | Pure xylose + pure glycerol | None | Yarrowia lipolytica Polt | Batch | 53.2 | 0.97 | 0.36 | 121 | |
| Pure xylose | Pure xylose + crude glycerol | None | Y. lipolytica Polt | Batch | 50.5 | 0.92 | 0.35 | 121 | |
| Steam exploded poplar | Xylose-rich hydrolysate | Detoxification | C. guilliermondii | Batch | 28.78 | 0.59 | 0.81 | 214 | |
| Steam exploded Eucalyptus globulus chips | Xylose-rich hydrolysate | Detoxification | Kluyveromyces marxianus NRRL Y-6373 | Batch | 28.05 | 0.68 | 0.16 | 215 | |
| H2SO4 pretreated SS | Xylose-rich hydrolysate | None | Corynebacterium glutamicum Cg-ax3 | Fed-batch | 27.0 | 2.25 | 216 | ||
| Hydrothermally pretreated CC | Whole slurry of corn cobs | None | Saccharomyces cerevisiae PE-2 (GM) | Fed-batch | 29.61 | 0.93 | 0.54 | 115 | |
| CC | Corn cob hydrolysate | Detoxification | S. cerevisiae IS5-d | Fed-batch | 143 | 0.93 | 1.83 | 217 | |
| S. cerevisiae 2bpgi | Fed-batch | 162 | 2.13 | ||||||
| Lactic acid | HCl pretreatedSCB | Xylose-rich hydrolysate | None | Bacillus coagulans DSM ID 14-300 | Batch | 55.99 | 0.87 | 1.70 | 218 |
| H2SO4 pretreated SCB | Xylose-rich hydrolysate | None | Lactobacillus pentosus ATCC 8041 | Batch | 42.4 | 0.71 | 1.02 | 125 | |
| Batch SScFa | 64.8 | 0.93 | 1.01 | ||||||
| DryPB pretreated CS | Carbohydrate-rich biomass | None | Pediococcus acidilactici ZY1 | Fed-batch SScF | 97.3$ | 0.926# | 1.01 | 219 | |
| NaOH pretreated CS | Carbohydrate-rich & delignified biomass | Washed | P. acidilactici PA204 | Fed-batch SScF | 104 | 0.69 | 1.24 | 220 | |
| NaOH pretreated CC | Carbohydrate-rich & delignified biomass | None | K. marxianus YKX071 (GM) | Fed-batch SScF | 101.7 | 0.56 | 1.10 | 221 | |
| Organosolv pretreated beechwood | Cellic CTec2 & HTec2 hydrolysed mixed sugars | None | B. coagulans DSM ID 14-298 | Batch | 88.09 | 0.674 | 2.93 | 222 | |
| Succinic acid | NaOH pretreated SCB | Delignified carbohydrate-rich biomass hydrolysed by cellulclast 1.5L | None | Y. lipolytica PSA02004 | Batch | 33.2 | 0.58 | 0.33 | 223 |
| Sulfuric acid pretreated CS | Mixed hydrolysate (81% xylose; 14% glucose) | None | Escherichia coli BA408 | Batch | 23.1 | 0.85 | 0.24 | 224 | |
| Sulfuric acid pretreated EG | Mixed hydrolysate (69% xylose; 29.4% glucose) | None | E. coliM6PM | Dual phase (aerobic growth and anaerobic production phase) | 30.03 | 0.64 | 0.41 | 225 | |
| Acid sulphite pulping of Eucalyptus globulus | Xylose-rich spent sulphite liquor | Nanofiltration using a vibratory shear-enhanced processing filtration unit | Actinobacillus succinogenes DSM 22257 | Continuous (dilution rate 0.2 h−1) | 19 | 0.48 | — | 226 | |
| Basfia succiniciproducens DSM 22022 | 22 | 0.55 | — | ||||||
| CS hydrolysate | Glucose: xylose (70:30) |
C. glutamicum CGS5 (GM) | Two stage (initial growth and production stage) | 98.6 | 0.87 | 4.29 | 227 | ||
| Dilute acid pretreated CC | Xylose rich hydrolysate | Detoxification | C. glutamicum NC-2 (GM) | Two stage (initial growth and production stage) | 40.8 | 0.69 | 0.85 | 228 | |
| SCB | Hydrolysate (glucose: xylose; 2:1) |
None | E. coli BA305 | Fed-batch | 39.3 | 0.97 | 0.327 | 229 | |
| 2,3 Butanediol | Hydrothermally pretreated SCB | Xylose-rich hydrolysate | None | Mutant of E. ludwigii | Fed-batch | 32.3 | 0.36 | 0.84 | 148 |
| Detoxification | 63.5 | 0.33 | 0.43 | ||||||
| KOH pretreated corn cobs followed by hemicellulose precipitation | Enzymatically derived xylose-rich hydrolysate | None | K. oxytoca XF7 | Batch | 12.18 | 0.97 | 0.50 | 230 | |
| H2SO4 pretreated soybean hulls | Xylose-rich hydrolysate | None | Pantoea agglomerans BL1 | Batch | 20.5 | 0.50 | 0.55 | 231 | |
| None | K.pneumoniae BLh-1 | 21.9 | 0.40 | 0.30 | |||||
| H2SO4 pretreated soybean hulls | Xylose-rich hydrolysate | None | P. agglomerans BL1 | Batch | 14.02 | 0.53 | 1.17 | 232 | |
| H2SO4 pretreated wheat straw | Xylose-rich hydrolysate + exogenous glucose | None | Paenibacillus polymyxa DSM 365 | Batch | 23.4 | 0.27 | 0.28 | 233 | |
| Sulphite pretreated oil palm empty fruit bunches | Hydrolysed slurry + xylose-rich spent liquor | None | K. pneumoniae PM2 | SHF | 75.03 | 0.43 | 0.78 | 234 | |
| Corn cobs pretreated with mixed acids | Xylose-rich hydrolysate | None | Enterobacter cloacae M22 | Batch | 23.2 | 0.44 | 235 | ||
| Hot water extracted Birchwood | Xylose-rich hydrolysate | Detoxification by ultra-filtration | B. vallismortis B-14891 | Batch | 22.7 | 0.43 | 0.63 | 236 | |
| Ethanol | NaOH pretreated SCB | Xylose-rich hydrolysate | None | Scheffersomyces stipitis Y-7124 | Sequential fed-batch with cell-recycle | 18.52 | 0.32 | 0.36 | 158 |
| NaOH pretreated SCB | Xylose-rich hydrolysate | None | Spathaspora passalidarum Y-27907 | Sequential fed-batch with cell-recycle | 23.3 | 0.46 | 0.81 | 158 | |
| Steam exploded WS | Whole biomass slurry | None | S. cerevisiae F12 (GM) | Batch | 23.7 | 0.43 | 0.7 | 237 | |
| 1.5% v/v H2SO4 pretreated SCB | Xylose-rich hydrolysate | Detoxification by Ca(OH)2 | Spathaspora passalidarum Y-27907 | Batch | 19.4 | 0.4 | 0.8 | 238 | |
| n-Butanol | Hydrothermally pretreated SCB | Xylose-rich hydrolysate fortified with xylose | None | C. saccharoperbutylacetonicum DSM 14923 | Batch | 5.8 | 0.22 | 0.08 | 239 |
| Hot water extracted debarked hybrid poplar | Xylose-rich hydrolysate | Detoxification | C. acetobutylicum ATCC 824 | SScF | 6.8 | 0.16 | 0.07 | 240 | |
| Xylan | Pure xylan | None | C. acetobutylicum NJ4 | Batch-CBP involving co-culturing with hemicellulase secreting Thermoanaerobacterium thermosaccharolyticum M5 | 13.28 | 0.19 | 0.08 | 241 | |
| Hot water extraction and steam treatment with chips (60% aspen and 40% maple) | Xylose-rich hydrolysate | Laccase mediated detoxification | C. acetobutylicum | Batch | 4.17 | — | — | 242 | |
One of the most critical environmental parameters to be considered during processing of yeast for xylitol production is dissolved oxygen concentration. Since oxygen limited conditions usually favour xylitol formation, conversion of NADH to NAD+ is hampered and the reduced availability of NAD+ impedes xylitol to xylulose conversion, resulting in the accumulation of xylitol.59 The XR catalysing reduction of xylose to xylitol is NAD(P)H dependent, and hence to improve NAD(P)H levels, overexpression of glucose-6-phosphate dehydrogenase (glucose-6-phosphate + NADP+ → 6-phosphogluconolactone + NADPH) from the PP pathway in S. cerevisiae resulted in a xylitol titer and productivity of 196.3 g L−1 and 4.27 gL−1h, respectively.120 A new isolate, P. fermentans, subjected to chemical mutagenesis resulted in a strain with improved XR activity (34%) and reduced XDH (22.9%) activity. The fed-batch fermentation using a mutant strain of P. fermentans produced 98.9 g L−1 xylitol with conversion yield of 0.67 g g−1 using pure xylose as a substrate. Further, using non-detoxified xylose rich pre-hydrolysate from sugarcane bagasse, the strain amassed 79.0 g L−1 xylitol with an overall yield of 0.54 g g−1 respectively.55S. stipitis is well known for its high xylose utilization rate, but as xylose assimilation leads to high ethanol production, the host is not suitable for xylitol production. Yarrowia lipolytica, an oleaginous yeast is well known for production of lipids and TCA cycle intermediates. The yeast has a cryptic xylose metabolic pathway or inactive xylose assimilatory enzymes. As a result of it, Y. lipolytica is unable to grow on xylose as a sole carbon source but can biotransform xylose into xylitol when cultivated in xylose along with other carbon sources like glucose or glycerol. The Y. lipolytica Polt strain accumulated 53.2 g L−1 xylitol with a yield of 0.97 g g−1 using pure glycerol and xylose as carbon sources, where glycerol was used for biomass production. Similar results were obtained when pure glycerol was substituted with crude glycerol from the biodiesel industry (titer: 50.5 g L−1; yield: 0.92 g g−1).121
Other than the environmental characteristics, basic mechanisms like substrate and product mediated growth inhibition limits the final product titers in biological processes. Xylitol, a polyhydroxy compound, can interfere with the membrane fluidity of the host cell's membrane, disrupting growth and increasing xylitol accumulation. Nystatin, a membrane porin agent that can increase the permeability of the lipid membranes to ions, water and non-electrolytes, was used to increase the xylitol transport from the cell. The C. tropicalis ATCC 13803 strain cultured along with nystatin could accumulate 197 g L−1 xylitol with 0.75 g g−1 and 3.9 g L−1 h−1 yield and productivity, respectively.122
5.2 Lactic acid
Lactic acid (LA) or 2-hydroxyproponoic acid is an optically active compound and exists in L and D forms. Being a platform chemical, LA has diverse industrial applications in food, cosmetics, polymers and packaging. The most valued application of LA as a monomer is in the production of poly lactic acid (PLA), an alternative to commercial petrochemical polymers. The market price for food grade LA is approximately $1400–$1600 per metric ton. It has been estimated that the global market size of LA would reach ∼$8.7 billion by 2025, with a CAGR of 18.7%.123Various microorganisms like bacteria, fungi and yeast have been employed for the production of LA using xylose as the sole carbon and energy source (Table 4). Lactic acid bacteria (LAB) are considered predominant for industrial scale LA production. Based on the end products, LAB strains can be divided into two categories: homofermentative (Lactobacillus delbrukii, L. acidophilus, L. plantarum, and L. helveticus) and heterofermentative (L. brevis, L. diolivorans, L. fermentum, and L. reuteri) for lactic acid fermentation. In homofermentative bacteria, the carbon flux is directed only to lactic acid (LA) with no by-product formation, whereas in case of heterofermentative bacteria, a mixture of LA, acetic acid and ethanol are obtained with LA as the main product. One of the most common routes for LA production is the simple reduction reaction of pyruvate obtained from the EMP and/or PP pathway catalysed by the NADH-dependent lactate dehydrogenase enzyme (Fig. 6A). Lactobacillus pentoses and L. brevis can naturally ferment xylose through the pentose phosphate and phosphoketolase pathways, producing LA and a mixture of acetic and LA, respectively.124 Wischral et al. (2019) investigated different Lactobacillus spp., for fermentation of xylose-rich hemicellulosic hydrolysates and identified the L. pentoses strain efficiently utilizing the xylose-rich SCB hydrolysate obtained from combined alkali and acid pretreatment and accumulated 65 g L−1 LA with yield and productivity of 0.93 g g−1 and 1.01 g L−1 h−1, respectively.125 Similarly an engineered E. coli JU15 strain was supplemented with 32 g L−1 xylose and 42 g L−1 glucose, a simulated corn stover hydrolysate, resulting in 40 g L−1 LA with a conversion yield of 0.60 gLA gsugar−1.126Pediococcus acidilactici is a facultative anaerobic lactic acid producing strain with specificity to a wide range of substrates including xylose, but the strain P. acidilactici TY112 was not able to utilise xylose. Qui and associates blocked the PK pathway by deleting the pkt gene, and overexpressed heterologous xylose assimilation pathway genes xylA and xylB along with transketolase (tkt) (X5P + E4P → F6P + G3P) and transaldolase (tal) (G3P + S7P → F6P + E4P) genes, directing the carbon flux from the phosphoketolase to the pentose phosphate pathway. The recombinant strain accumulated 130.8 g L−1 LA with 0.68 g g−1 yield in simultaneous saccharification and fermentation mode using dilute acid pretreated wheat straw as feedstock.124 The group have employed a similar overexpression and deletion strategy in the D-LA producing P. acidilactici ZP26 strain, and the modified strain was adapted under laboratory conditions with xylose as the sole carbon source. The adapted strain P. acidilactici ZY15 accumulated 97.3 g L−1 D-LA with 0.92 g g−1 conversion yield through a simultaneous saccharification and co-fermentation strategy using dry dilute acid pretreated and detoxified corn stover feedstock.127 However the CCR limits the performance of various strains on LCB hydrolysates as they consist of both hexose and pentose sugars, and the E. coli JH15 strain was engineered to overcome CCR by deletion of the ptsG gene which encodes for IIBCglc (a PTS enzyme for glucose transport). The engineered strain produced 83 g L−1 D-LA with 0.83 g g−1 yield and 0.86 g L−1 h−1 productivity from co-fermentation with a mixture containing glucose and xylose in a 1:1 ratio.128
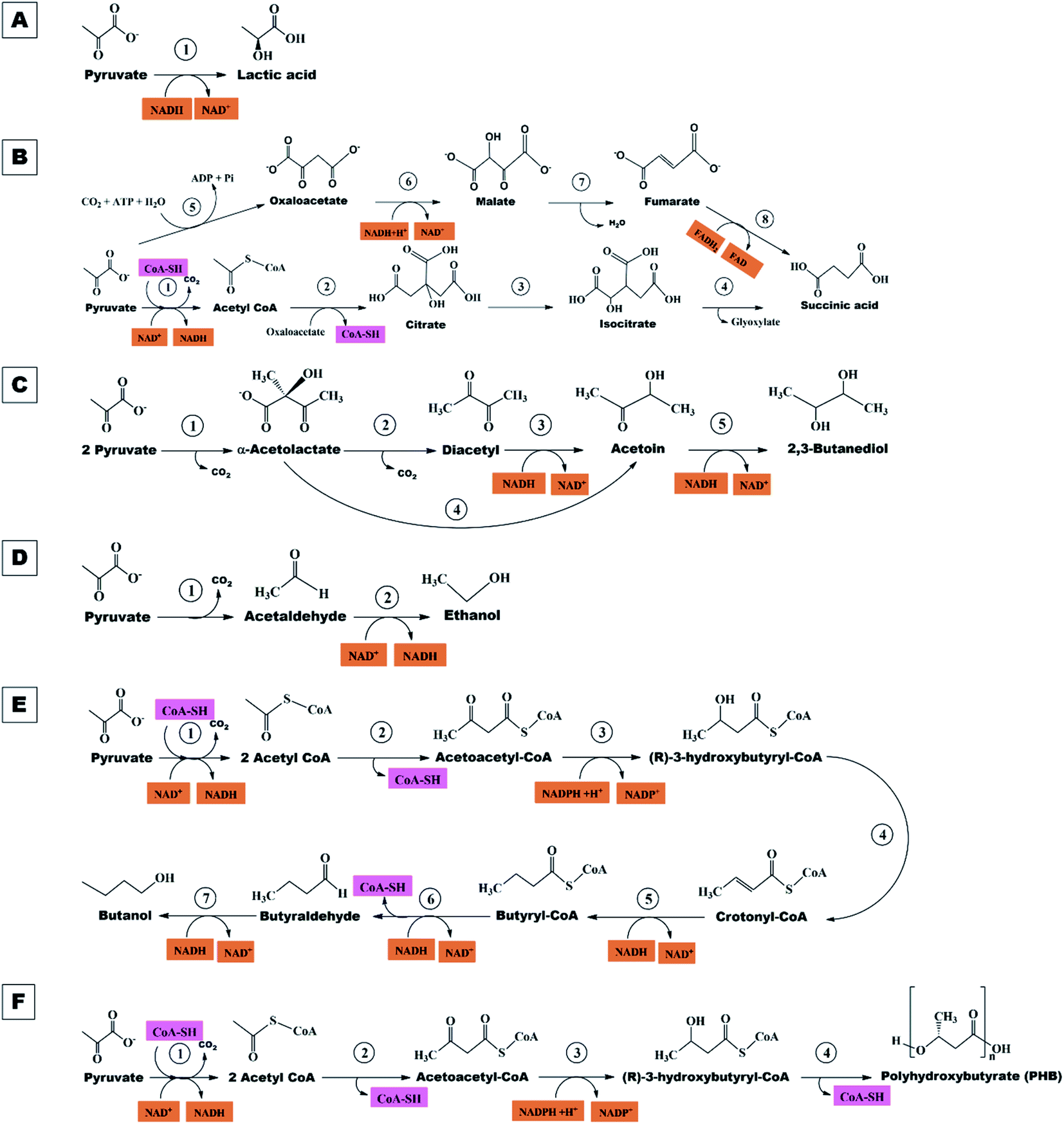 | ||
| Fig. 6 Biochemical pathways: (A) lactic acid, enzymes and genes involved: (1) lactate dehydrogenase (ldh). (B) Succinic acid, enzymes and genes involved: 1, acetyl-CoA synthetase (acs); 2, citrate synthase (gltA); 3, aconitase (acnAB); 4, isocitrate lyase (aceA); 5, pyruvate carboxylase (pyc); 6, malate dehydrogenase (mdh); 7, fumarase (fh); 8, fumarate reductase (frd) or succinate dehydrogenase (sdh). (C) 2,3-Butanediol, enzymes and genes involved: 1, α-acetolactate synthase (alsS); 2, spontaneous reaction; 3, diacetyl reductase (butA); 4, α-acetolactate decarboxylase (aldc); 5, butanediol dehydrogenase (bdh). (D) Ethanol, enzymes, and genes involved: 1, pyruvate decarboxylase (pdc); 2, alcohol dehydrogenase (Adh). (E) Butanol, enzymes and genes involved: 1, acetyl-CoA synthease (acs); 2, thiolase (thl); 3, 3-hydroxybutyryl-CoA dehydrogenase (Hbd); 4, crotonase (Crt); 5, butyryl-CoA dehydrogenase (EtfAB); 6, butyraldehyde dehydrogenase (AdhE2). (F) Polyhydroxybutyrate (PHB), enzymes and genes involved: 1, acetyl-CoA synthase (acs); 2,3-ketothiolase (PhaA); 3, acetoacetyl-CoA reductase (PhaB); 4, PHB synthase (PhaC). | ||
The commercially viable yeast S. cerevisiae produces LA in very minute quantities, and hence requires either a homologous or heterologous expression to increase the titers. Ethanol is the natural and dominant product by S. cerevisiae and would be the major competitor for LA production. The competition is for the precursor/substrate, pyruvate, and cofactor NADH between pyruvate decarboxylase and alcohol dehydrogenase. In order to prevent this, a pdc deficient strain was constructed which exhibited poor growth and productivities.129 In S. cerevisiae, heterologous overexpression of Xyl1, Xyl2, and Xyl3 from S. stipitis, a cellodextrin transporter (cdt-1) and a β-glucosidase (gh1-1) from the cellulolytic fungus Neurospora crassa, and additional laboratory evolution on medium containing cellobiose, resulted in a strain that could produce 83 g L−1 LA, with 0.66 g g−1 yield when cultivated on LCB hydrolysate containing 10 g L−1 glucose, 40 g L−1 xylose and 80 g L−1 cellobiose.130
5.3 Succinic acid
Succinic acid (SA) is an aliphatic dicarboxylic acid containing four carbon atoms with potent application as a precursor in pharmaceutical, polymer, and chemical industries. Like LA, SA is a platform chemical and due to the presence of two carboxyl acid groups, SA can be converted into a variety of products such as succinic anhydride, succinic esters, 2-pyrrolidine, and polyesters for synthesizing biodegradable plastics.131 According to global market research, the market size was expected to reach $237.8 million by 2022 with a CAGR of 9.2% (Succinic Acid Market Size & Share | Industry Analysis Report, 2022 (grandviewresearch.com)). SA production from pure sugars and LCB hydrolysates has been reported using natural producers Anaerobiospirillum succiniciproducens, Actinobacillus succinogenes, Mannheimia succiniciproducens, Basfia succiniciproducens and genetically engineered strains E. coli and Y. lipolytica (Table 4).132,133 The three different biochemical pathways for SA production are the oxidative tricarboxylic acid (TCA) cycle, the reductive branch of the TCA cycle, and the non-frequent glyoxylate pathway (Fig. 6B).134Actinobacillus succinogenes and Basfia succiniciproducens are the most evaluated and predominant native SA producing strains, with the ability to utilise either pure sugars or LCB hydrolysates.135Actinobacillus succinogenes 130Z, a natural SA producer, was immobilized and continuously fed with xylose-rich hydrolysate from corn stover and generated 39.6 g L−1 SA, with yield and productivity of 0.78 g g−1 and 1.77 g L−1 h−1, respectively.136 In another study, when the same strain was cultivated using dilute acid pretreated corn stover hydrolysate, an SA titer, yield and productivity of 42.8 g L−1, 0.74 g g−1 and 1.27 g L−1 h−1 were obtained, respectively.137 Pateraki et al. (2016) cultivated A. succinogenes and B. succiniciproducens on mixed sugar feedstock (synthetic solution) containing 72% xylose, 12.2% galactose, 10.9% glucose, 4.2% mannose, and 0.1% arabinose. The SA titer, yield and productivity achieved with A. succinogenes were 26 g L−1, 0.76 g g−1 and 0.66 g L−1 h−1, respectively. Similar results were obtained with B. succiniciproducens: 27.4 g L−1, 0.69 g g−1 and 0.60 g L−1 h−1.138 In addition to single strains, microbial consortiums have been used for SA production. A microbial consortium containing Thermoanaerobacterium thermosaccharolyticum M5 and A. succinogenes 130Z was employed to utilize hemicellulosic derived sugars to produce SA. The T. thermosaccharolyticum M5 strain has the ability to saccharify the LCB components, by secretion of extracellular enzymes like xylanase where the xylose obtained was converted into SA by A. succinogenes. The consortium with the consolidated bioprocessing approach was able to generate 32.5 g L−1 SA with a yield of 0.39 g g−1.139
The E. coli KJ122 strain was previously modified to reduce the by-products by deleting the respective genes, which increased the SA yield, titers and productivity when cultivated on glucose and sucrose in mineral medium under anaerobic conditions.140 The strain was observed to be defective in growth and SA production during cultivation on xylose medium. It was speculated that the major reason behind this could be energy limitations as transport and phosphorylation of 1 mole of xylose requires 2 moles of ATP, but only 1.67 moles of ATP are generated when xylose is used for biosynthesis of SA.141 Thus, xylose as the sole carbon source cannot provide efficient energy currency for the cell growth and development. Hence Khunnonkwao and associates deleted the xylFGH (ATP dependent ABC transporter), which is an energy expensive xylose transporting transmembrane protein and the resultant mutant strain was subjected to adaptive evolution on xylose media. When the recombinant E. coli KJ12201-14T strain was cultured on a glucose and xylose mixture, it utilized both the sugars and accumulated 84.6 g L−1 SA with yield and productivity of 0.86 g g−1 and 1.01 gL−1h, respectively.142 An engineered E. coli strain, YL104H, with deleted pathways for LA, ethanol and other byproducts was evaluated for SA production using corn-based liquor containing glucose and xylose in a ratio of 2:1. Employing an alternative strategy to that followed where the ABC transporter was deleted, Zhang and associates attempted a process modification approach where the intracellular ATP concentration was maintained by co-substrate fermentation with supplementing glucose and xylose in a 2:1 ratio. The process resulted in accumulation of 61.66 g L−1 SA, with 0.95 g L−1 h−1 productivity.143
Bacterial strains are more sensitive to changes in the physiological pH and require continuous addition of neutralizing agents. The addition of neutralizing agents not only dilutes the concentration of SA in the fermented broth but also converts organic acids into the salt form which complicate the downstream processing and increase the production cost. On the other hand, yeast strains are more promising for SA production, as they have better tolerance and can withstand lower pH. In a study, Prabhu and associates (2020) engineered the Y. lipolytica PSA02004 strain to utilize xylose as the sole source of carbon and energy by overexpressing the pentose phosphate pathway comprising XR, XDH and XK under a strong constitutive promoter. The recombinant strain accumulated 22.3 g L−1 SA using xylose-rich hydrolysate from SCB hydrolysate.144
5.4 2,3-Butanediol
2,3-Butanediol (BDO) is a 4-carbon diol, with applications in food, cosmetics, fuel-additives, agrochemicals, and pharmaceuticals. One of the major applications of BDO is production of methyl ethyl ketone (MEK), an organic solvent used in production of resins and lacquers.145,146 Microbial physiology adaptation for the BDO pathway is hypothesized to prevent intracellular acidification and balance the reducing equivalents. Two moles of pyruvate undergo sequential oxidoreductive reactions to form BDO with α-acetolactate and acetoin/diacetyl as intermediates as shown in Fig. 6C.In biological BDO synthesis, the main factor influencing the economy of the process is the substrate cost which accounts for 50% of the total production cost.147 Microorganisms of different genera, Klebsiella, Lactobacillus, Enterobacter, and Bacillus (Table 4), have been reported to accumulate large amounts of BDO (50–120 g L−1) from a variety of renewable feedstocks like cane molasses, cane sugar, SCB, and fruit and vegetable waste. In our recent study, we evaluated the performance of a mutant Enterobacter ludwigii strain on pure xylose, non-detoxified and detoxified xylose-rich hydrolysates obtained from the thermochemical pretreatment of SCB. During the fed-batch cultivation, the strain produced 71.1 g L−1 BDO using pure xylose with a conversion yield and productivity of 0.40 g g−1 and 0.94 g L−1 h−1, respectively. In case of non-detoxified and detoxified hydrolysates, BDO titers of 32.7 and 63.5 g L−1 with yield of 0.33 and 0.36 g g−1, and productivity of 0.43 and 0.84 g L−1 h−1, were achieved, respectively.148 A study conducted by Wang and associates implemented a process engineering approach by optimizing the media components to improve the BDO titers and productivity using Klebsiella pneumoniae. The K. pneumoniae strain with optimal media components and physiological conditions could produce 42.7 g L−1 BDO with 95% theoretical maximum yields and 99% xylose sugar uptake efficiency.147 Although Klebsiella is a known workhorse in production of value-added chemicals, its resistance to xylose is not satisfactory; xylose concentration >70 g L−1 was observed to inhibit the growth and metabolic performance of the strain. A global transcription regulating sigma (σ) factor encoded by the rpoD gene was observed to improve the substrate consumption rate and metabolic behaviour in E. coli strains. Hence, to overcome the xylose mediated inhibition, the rpoD gene was overexpressed in K. pneumoniae which caused an increment in substrate tolerance up to 125 g L−1 xylose, and product tolerance by 200%. Xylose transport, glycerol-3-phosphate acyl transferase, and phosphate kinase genes were observed to be upregulated by 5.7, 2.2 and 3-fold, respectively.149
To modulate the commercially viable S. cerevisiae for BDO production, the biochemical pathway for BDO should be overexpressed and biosynthetic pathways leading to byproduct (ethanol, acetic acid and glycerol) formation must be eliminated. Kim et al. (2015) constructed a BDO producing S. cerevisiae strain by introducing the BDO pathway and to this end, α-acetolactate synthase (AlsS) and α-acetolactate decarboxylase (AlsD) from B. subtilis, and endogenous BDO dehydrogenase (BDH1) and NoxE from L. lactis were overexpressed. Further, production of ethanol (adh 1–5) and glycerol (gpd 1 and gpd 2) was blocked by deleting the relevant genes. The resulting engineered strain produced 72.9 g L−1 BDO with 0.41 g g−1 and 1.43 g L−1.h yield and productivity, respectively, using glucose as the carbon source.150 Extending the work, later Kim and associates constructed a xylose assimilatory S. cerevisiae strain by overexpressing the S. stipitis transaldolase (S7P + G3P → E4P + F6P) and endogenous NADH preferring XR. The recombinant strain showed a 2.1-fold increase in xylose consumption rate and 1.8-fold increase in BDO productivity. Further NOX and PDC1 genes from L. lactis and C. tropicalis were heterologously overexpressed resulting in the BD5X-TXmNP strain. The fed-batch cultivation of the resultant strain on xylose produced 96.8 g L−1 BDO with 0.58 g L−1 h−1 productivity.151
5.5 Ethanol
Ethanol/ethyl-alcohol/bioethanol is the most widely used biofuel in the transportation sector and offers several advantages such as a higher octane number, high combustion efficiency and increased heat of vaporization. Bioethanol is less toxic, readily biodegradable and produces fewer air-borne pollutants in comparison to petroleum fuel and most promising alternatives to gasoline. However, due to its hygroscopic nature, complete replacement of gasoline with ethanol is not possible as water vapour can corrode the engine.152,153 Currently, ethanol is blended with gasoline at different levels (5–20%) across the globe. It has been found that the blended fuel causes a substantial reduction in emission of hydrocarbons and greenhouse gases.154 The commercial production of ethanol from various renewable feedstocks has gained significant interest due to its increased application as a fuel component in gasoline. In 2019, the global ethanol production was 115 billion litres ($38.83 billion), and with a CAGR of 1.77% the demand has been expected to increase to $43.14 billion by 2025. (Global Ethanol Market – forecasts from 2020 to 2025 (researchandmarkets.com)). Ethanol fermentation by S. cerevisiae is one of the oldest practices in Biotechnology. S. cerevisiae is the most promising cell factory for ethanol production and is employed at the industrial level. Currently, yeast is used for generating ethanol from a variety of feedstocks.155The ethanol production from xylose follows production of X5P through the pentose phosphate pathway and further proceeds through the EMP pathway. Pyruvate, the final product of the EMP pathway, is converted to ethanol through acetaldehyde as an intermediate as shown in Fig. 6D. However, S. cerevisiae lacks pentose assimilatory pathways and can generate ethanol from xylose only after introducing the enzymes connecting xylose to the central carbon metabolism.156 Even though few strains like S. stipitis, P. fermentans, P. kudriavzevii, and Spathaspora (S. passalidarum) are well-known xylose assimilating yeast, the processes are limited due to substrate and product mediated inhibition. For example, the strains S. stipitis and S. passalidarum on xylose fermentation resulted in maximum ethanol titers of 29.9 g L−1 and 25 g L−1, with a conversion yield of 0.47 and 0.41 g g−1, and productivity of 1.5 and 1.04 g L−1 h−1, respectively.157–161
The possibility of xylose as the feedstock to produce ethanol was explored in S. cerevisiae and to this end, XR and XDH genes from S. stipitis were overexpressed in S. cerevisiae. The heterologous expression resulted in a lower ethanol titer of 10.7 g L−1 with a conversion yield of 0.19 g g−1, and xylitol titers and yield of 14.3 g L−1 and 0.26 g g−1, respectively. In Pichia sp., there is a competition between ethanol and xylitol formation for the carbon flux. The carbon flux towards xylitol synthesis can be reduced by altering the cofactor specificity. Xiong et al. (2013) expressed a mutant form of XR (K270R) in S. cerevisiae with higher specificity of XR for NADH than NADPH which resulted in a higher ethanol (0.38 g g−1) and reduced xylitol yield (0.08 g g−1).162 Along with ALE, polyploidy was also considered as an accelerative solution for adaptation of yeast. In the process, either the native or mutant haploid strains are subjected to mating to produce diploid or triploid strains. These strains were observed to have improved phenotypic and genotypic characteristics compared to the parent strains. Using this approach, S. cerevisiae XR-K270R mutant strain diploids and triploids were produced by Liu and associates. Furthermore, the comparative analysis between the haploid, diploid and triploid strains displayed better performance of the triploid on dilute acid and alkali pretreated corn cob and corn stover hydrolysates resulting in a maximum ethanol production yield of 87.3%, while the diploid strain yielded 76.2% ethanol.163 The recent discovery of non-conventional yeast S. passalidarum, with xylose fermenting ability and possessing NADH dependent XR, could enable alternative research focusing on heterologous expressions rather than protein or cofactor engineering of the known XR from S. stipitis.157 Further research towards expression of XR and XDH from Spathaspora sp. could result in increased ethanol titers and yields in commercial yeasts.
5.6 n-Butanol
n-Butanol is a four-carbon straight chain alcohol and is considered a better biofuel than ethanol due to its high octane number, higher heating value, lower volatility, ignition problems, low miscibility with water and higher viscosity.80 In a chemical approach, aldol condensation (oxo process) can produce n-butanol by hydroformylation and hydrogenation of propylene. In the biological route, n-butanol is a part of acetone–butanol–ethanol (ABE) fermentation and Clostridium spp. are well known cell factories with ABE fermentation (Fig. 6E).164,165 However, the bio-butanol production suffers from low titers, yield, and product mediated inhibition. Clostridium strains can naturally ferment xylose into n-butanol via ABE fermentation. C. beijerinckii could accumulate 26.3 g L−1 ABE with a yield of 0.44 g g−1 using Ca(OH)2 detoxified xylose-rich corn stover hydrolysate.166 Although the n-butanol yield is 20% lower than that of ethanol, the energy generated from n-butanol is 32% higher than that from ethanol.167 Currently with the available titers and yield, the cost of biobutanol production is around $1.8 L−1, but further optimization of the biocatalysts and process conditions could reduce the production cost to $0.6 L−1 which is comparable to that of gasoline and other fossil fuels.167 Jiang and associates implemented the process of consolidated bioprocessing (CBP), where a xylan degrading and n-butanol producing strain, Thermoanaerobacterium sp. M5, was evaluated. The strain was able to grow at 55 °C, with efficient expression of xylanase, β-xylosidase and alcohol dehydrogenase for the conversion of xylose to n-butanol through ABE fermentation.168 As the Thermoanaerobacterium sp. M5 strain has efficient xylan degradation efficiency, a co-cultivation strategy was investigated along with solventogenic strain C. acetobutylicum NJ4. The co-cultivation of these strains resulted in 13.3 g L−1n-butanol with a yield of 0.19 g g−1.169 Supplementing a crude hemicellulosic hydrolysate may be toxic to the microbial cells in the initial lag phases, and hence in a study, after growing C. saccharoperbutylacetonicum DSM 14923 on sugarcane molasses for 24 hours, hemicellulosic hydrolysate was added into the media resulting in 10 g L−1 butanol, with a yield and productivity of 0.31 g g−1 and 0.14 g L−1 h−1, respectively.1645.7 Polyhydroxybutyrate (polyhydroxyalkanoates)
Polyhydroxyalkanoates (PHAs) are hydroxy alkanoic polyesters which are stored as intracellular granules in various prokaryotic microorganisms and are accumulated when the carbon source is in surplus along with limitation of a key nutrient.170,171 Although the primary function of these polyesters is the storage of carbon and energy, they also play a role in protecting the microbial cell from stress. Poly-3-hydroxybutyrate (PHB) and its derivatives like PHB-co-3-hydroxyvalerate (PHB-co-HV),171 polylactate-co-3-hydroxybutyrate (PL-co-HB),172 and 4-hydroxyhexanoate are a type of PHA produced by prokaryotes (Fig. 6F). PHAs have ample applications in the field of nanotechnology, drug delivery, medical prosthetics, etc. Although PHB is well known and characterized, its brittle and crystalline structure limits its industrial relevance, but its derivatives like PHB-co-HV have impressive biomedical applications. A halophilic Bacillus sp. isolated from mangrove soil was observed to utilize a wide range of carbon sources. The strains could accumulate PHB-co-HV up to 73% of biomass weight on xylose-rich acid hydrolysates of sugarcane trash, under optimal conditions.171Burkholderia sacchari, an industrially viable strain for the production of xylitol, xylonic acid and PHB, was engineered by overexpressing the xylose transporters (XylE, and XylFGH), metabolic genes (XylA, and XylB), and the regulatory gene (XylR). The engineered B. sacchari strain showed 55, 77.3 and 71% improvements in the growth rate, polymer yield and cell dry weight, respectively.170 As explained in Section 5.3, an E. coli strain was engineered to hydrolyse the xylan fraction of hemicellulose by heterologous overexpression of β-xylosidase and an endoxylanase, and further the saccharified xylose was converted to PLA-co-HB. The resulting strain on a xylan based production medium with additional pentose sugar as a co-substrate increased the polymer yield up to 37% in comparison to the strain cultivated on pure xylose as a sole carbon source.173 In a lignocellulosic biorefinery, thermophiles are of utmost importance due to the benefit of simultaneous saccharification and fermentation as most of the enzymes utilized for hydrolysis of hemicellulosic and cellulosic residues are active at around 50 °C. The thermophilic bacterium Schelegelella thermodepolymerans DSM 15344 is a natural polymer degrading microorganism with optimal growth at 55 °C. The genome mapping revealed a conserved PHA biosynthesis pathway, with 70–76% similarity to the model PHA accumulating microorganism Cupriavidus necator N-1. The interesting feature identified in the S. thermodepolymerans strain is accumulation of more PHB on xylose (54%) in comparison to glucose (37%) as a carbon source.174 The interesting and highly investigated strain for PHA production is Ralstonia eutropha which lacks the ability to metabolize xylose. The recombinant R. eutropha strain expressing the E. coli XylAB genes was able to accumulate 33.7 g L−1 PHB which is 79% of biomass weight, and the same strain when cultured on the hydrolysate solution of sunflower stalk consisting of 16.8 g L−1 glucose and 5.9 g L−1 xylose resulted in production of 7.86 g L−1 PHB corresponding to 72.5% CDW.175 As PHB accumulation in the microorganism is growth dependent, the optimal conditions for the cell growth would favour PHB accumulation. A new isolate, B. megaterium J-65, was able to accumulate 35% CDW under optimal conditions with 2% xylose as the sole carbon source.176 Supplementing pretreated corn husk hydrolysate along with nitrogen deficient production media to B. megaterium could accumulate 57.8% PHB which is almost 3-fold higher than on glucose as a sole carbon source.177
In earlier years, researchers were more focussed on valorisation of both the carbohydrate fractions of lignocellulosic biomass to ethanol. But lately, the trend has changed, and a diverse product portfolio is preferred as it has been found to be more profitable as compared to targeting a single product. Particularly, in the last five years researchers have attempted to integrate the developed process modules with techno-economics to understand the benefits associated with holistic utilization of all biomass components. For instance, Ou et al. (2021) showed that if 1500 tonnes of miscanthus were processed for sugar production per day, the minimum sugar selling price (MSSP) would be $446 per tonne.178 However, when the xylose stream obtained after auto-hydrolysis was diverted for xylitol production, the MSSP was reduced to $347 per tonne. Similar observations were made by Giuliano et al. (2018), who found that if only cellulosic ethanol was targeted from steam exploded corn stover, the payback ethanol price was €1.62 per kg.179 But when the hydrolysed xylose stream was bio-transformed to xylitol, it reduced the overall cost of ethanol by 50.9%. In yet another study, xylitol co-production could raise the profitability of cellulosic ethanol by 2.3-fold during sugarcane biorefining when the fed-batch fermentation strategy was adopted.180 Recently, Ranganathan (2020) showed that when glucose derived from rice straw was used for ethanol production, but xylose was kept intact, the cost of ethanol was $0.627 L−1. But when xylose was converted to furfural and lignin was upgraded to biochemical its cost reduced to merely $0.25 L.181 Thus, all these recent studies give a fair indication on how xylan/xylose valorisation can augment the carbohydrate economy and increase the profitability of LCB-based biorefineries. Researchers are relentlessly working towards accelerating the biotechnological production of some bio-based and commercially important chemicals through genetic and protein engineering approaches as shown in Table 5.
| Product | Microorganism | Modification | Improvement | References |
|---|---|---|---|---|
| Xylitol | Meyerozyma guilliermondii | Cloning and overexpression of XR, and knockout of XDH | The engineered strain exhibited a 3-fold improvement in xylitol yield | 243 |
| E. coli | Expression of XR and glucose dehydrogenase | The immobilized recombinant cells could maintain the enzyme activity up to 80% after 10 repeated batches | 244 | |
| K. marxianus | Expression of XR and transporter genes | The engineered strain efficiently utilized glucose and xylose from xylose-rich hydrolysate for production of xylitol > 100 g L−1 | 245 | |
| S. cerevisiae | Expression of the constitutive GPD promoter for the ZWF1(cytoplasmic G6P dehydrogenase) gene to increase the NADPH pool, a cofactor for XR | The heterologous expression of a constitutive promoter resulted in a 12% increase in xylitol yield (0.78 vs. 0.88) | 246 | |
| Lactic acid | S. cerevisiae | Heterologous overexpression of the lactate dehydrogenase (ldhA) gene from Rhizopus oryzae under the control of the PGK1 promoter through chromosome integration | High lactate dehydrogenase activity and LA titers of 28.9 g L−1 with 0.69g g−1 yield | 247 |
| Scheffersomyces stipitis | Heterologous overexpression of ldh from L. helveticus under the control of the native ADH1 promoter | Ethanol production decreased 15 to 30% and carbon flux is shifted towards LA from xylose resulting in 58 g L−1 LA with 0.58 g g−1 yield | 248 | |
| Pediococcus acidilactici | Heterologous expression of xylose assimilating genes XylA and XylB, substitution of endogenous phosphoketolase with heterologous transketolase and transaldolase | The metabolic carbon flux is concentrated towards LA biosynthesis resulting in 97.3 g L−1 LA with 0.93 g g−1 conversion yield | 219 | |
| Lactococcus lactis | Disruption of the phosphoketolase gene, introduction of the transketolase gene | High LA titers of 50.1 g L−1 with 1.58 mol mol−1 yield and 99.6% purity after the downstream processing was quantified | 249 | |
| Candida sonorensis | Integrating lactate dehydrogenasegene from L. helveticus | The engineered strains were able to accumulate 31 g L−1 LA with 0.62 g g−1 yield under microaerobic conditions | 250 | |
| Succinic acid | Yarrowia lipolytica PSA02004 | Overexpressing the pentose pathway cassette (XR, XDH and XK genes) | The engineered strain could grow on xylose as a sole energy and carbon source producing 22.3 g L−1 SA | 144 |
| Corynebacterium glutamicum | Heterologous expression of XI, overexpression of XK, transaldolase, 6-phosphogluconate dehydrogenase and phosphoketolase | The recombinant strain enhanced the growth and xylose consumption rate resulting in 7.22 g L−1 SA with 0.18 g g−1 yield | 251 | |
| Escherichia coli KJ122 | Deletion of XylFGH and XylE genes and ALE | Improved succinate titers of 85 g L−1 with 0.85 g g−1 yield, and 0.7 g L−1 h−1 productivity | 142 | |
| E. coli K12 | Inactivation of pyruvate formate lyase (pflB) and lactate dehydrogenase (ldhA) | Significant increase in cell mass (2.5 g L−1) and succinate (11.6 g L−1) production | 252 | |
| Aspergillus niger | Overexpression of fumarate reductase, disruption of gluconic acid and oxalic acid production | The engineered strain was able to utilize xylose-rich hydrolysates derived from sugar beet and wheat straw to produce 23 and 9 g L−1 SA | 253 | |
| 2,3-Butanediol | S. cerevisiae | Heterologous expression of the BDO biosynthetic pathway, deletion of ethanol and glycerol assimilatory genes. Restoration of redox balance by overexpression of NADH oxidase | Highest yield (0.41 g g−1) and productivity (1.43 g L−1 h−1) using S. cerevisiae as the host | 85 |
| Enterobacter cloacae | Expression of BDO dehydrogenase, inactivation of glucose transporter and overexpression of galactose permease | The engineered strain could overcome CCR and was able to utilize glucose and xylose simultaneously, producing 119.4 g L−1 BDO with 2.3 g L−1 h productivity | 254 | |
| Zymomonas mobilis | Heterologous expression of BDO biosynthetic pathway consisting of acetolactate synthase, acetolactate decarboxylase, and butanediol dehydrogenase | The engineered strain was able to produce BDO (13.3 g L−1) utilizing both C5 and C6 sugars | 255 | |
| S. cerevisiae | Expression of S. stipitis transaldolase, xylose reductase, L. lactis NADH oxidase and overexpression of pdc from C. tropicalis | A 2.1-fold increase in xylose consumption and 1.8-fold increase in BDO productivity | 256 | |
| K. pneumoniae | Overexpression of transketolase, NADP transhydrogenase subunit alpha, and NADH dehydrogenase subunit F | The engineered strain increased the xylose consumption and BDO production resulting in 38.6 g L−1 BDO with 0.62 g L−1 h−1 productivity | 149 | |
| Ethanol | S. cerevisiae | Overexpressing pentose pathway genes (XR and XDH) | Assimilation of xylose towards ethanol biosynthesis with a conversion yield of 0.19 g g−1 | 162 |
| S. cerevisiae | Overexpression of mutant XR K270R | Increased specificity towards NADH rather than NADPH, resulting in increased ethanol yield (0.38 g g−1) and reduced xylitol yield (0.08 g g−1) | 162 | |
| n-Butanol | E. coli | Expression of a synthetic butanol pathway | In a defined medium 4.32 g L−1n-butanol was produced | 257 |
| Clostridium tyrobutyricum | Heterologous expression of XylT, XylA, and XylB from C. acetobutylicum and overexpression of native alcohol dehydrogenase | The engineered strain could accumulate 12 g L−1n-butanol with 0.12 g g−1 yield | 258 | |
| PHB | E. coli | Heterologous expression of XI, XK and pentose transport protein from B. subtilis in E. coli harbouring the PHB pathway from Ralstonia eutropha | Simultaneous utilization of glucose and xylose increased PHB titers 2-fold | 259 |
| S. cerevisiae | Heterologous overexpression of the PHB biosynthesis pathway from Cupriavidus necator | The engineered strain could produce 1.99 mg PHB/g xylose | 260 | |
| S. cerevisiae | Heterologous overexpression of NADH dependent acetoacetyl-CoA from Allochromatiumvinosum replacing the gene from C. necator | PHB titers increased 5-fold under aerobic and 8.4-fold under oxygen limited conditions | 261 |
6. Exploring the efficiency of multiple xylose assimilatory pathways for carbon flux towards SA and biomass production using the established genome scale models
Small scale metabolic networks were constructed by retrieving information from genome scale metabolic models. Xylose assimilation pathways were incorporated into the metabolic network of C. glutamicum, E. coli, A. succinogenes and Y. lipolytica. Elementary flux mode analysis was implemented to elucidate optimal pathways for producing biomass or succinic acid (SA) through different xylose assimilation pathways. Theoretical maximum yields are summarised in Table 6.| Yield | Glucose | Xylose-XR–XDH pathway (XR–XDH) | Xylose-isomerase pathway (XI) | Xylose-Weimburg pathway (XW) | Xylose-Dahms pathway (XD) |
|---|---|---|---|---|---|
| C. glutamicum | |||||
| Biomass yield (c-mol biomass c-mol substrate−1) | 0.84 | 0.81 | 0.84 | 0.59 | 0.54 |
| Succinic acid yield (c-mol succinate c-mol substrate−1) | 0.67 | 0.67 | 0.67 | 0.80 | 0.40 |
|
|||||
| E. coli | |||||
| Biomass yield (c-mol biomass c-mol substrate−1) | 0.90 | 0.90 | 0.90 | 0.63 | 0.58 |
| Succinic acid yield (c-mol succinate c-mol substrate−1) | 0.67 | 0.67 | 0.67 | 0.80 | 0.40 |
|
|||||
| A . succinogenes | |||||
| Biomass yield (c-mol biomass c-mol substrate−1) | 0.34 | 0.34 | 0.34 | NA | NA |
| Succinic acid yield (c-mol succinate c-mol substrate−1) | 0.67 | 0.80 | 0.80 | NA | NA |
|
|||||
| Y . lipolytica | |||||
| Biomass yield (c-mol biomass c-mol substrate−1) | 0.73 | 0.70 | 0.72 | 0.49 | 0.46 |
| Succinic acid yield (c-mol succinate c-mol substrate−1) | 0.90 | 1.0 | 1.0 | 0.80 | 0.80 |
6.1 Corynebacterium glutamicum
C. glutamicum is a well-known industrially relevant bacterium that is widely engineered to produce value-added products from a wide range of carbon sources.182 The xylose isomerase (XI) pathway was previously implemented in C. glutamicum,183 which showed a 30% theoretical maximum yield of succinic acid from xylose. As seen from Table 6, XI and XR–XDH pathways have the potential to produce yields equivalent to those on glucose on a carbon basis. Theoretical maximum yields of up to 80% can be achieved when the Weimberg (WMB) pathway is used while the lowest possible maximum yield of 40% is observed with the Dahms pathway. Optimal routes using the different xylose assimilating pathways are shown in Fig. 7. As seen from Fig. 7, XR–XDH and XI show similar optimal routes for SA production and about 2 mol mol−1 of O2 demand is observed for both the pathways. The WMB pathway seems to be the most efficient route that can reach a maximum yield of 1 mol SA per mol xylose. This is attributed to alpha-ketoglutarate that is generated in the upper xylose assimilation pathway which directly enters the TCA cycle. As seen, biomass yields are significantly lower compared to XI and XR–XDH pathways (∼20%) in both the WMB and Dahms pathways. When the Dahms pathway is used only 40% of carbon can be theoretically converted to SA under non-biomass production conditions. Under non-biomass conditions which would be normally implemented for succinic acid production (i.e. dual fermentation mode), surplus ATP must be replenished which is seen as output of ATP for maintenance purposes. A futile cycle could be generated which can replenish this surplus ATP under non-biomass production conditions. The NADPH required for xylose assimilation via the XR–XDH pathway is mainly supplied by isocitrate dehydrogenase. Optimal succinate production modes were also observed (data not shown) where the glyoxylate cycle can be active and malic enzyme could be providing the required NADPH. The XW pathway was implemented previously in C. glutamicum,184 which showed growth inhibition due to accumulation of xylose 5-phosphate. Since the advantage of using the XW pathway lies in preventing the loss of carbon via CO2 production, there is about 40% less CO2 being produced and 25% less O2 demand in comparison to the XR–XDH and XI pathways. The XD pathway loses about 80% more carbon in the form of CO2 and requires 30% more O2 compared to the XR–XDH and XI pathways. | ||
| Fig. 7 Theoretical maximum (optimal) production of SA via (A) the XR–XDH pathway, (B) XI pathway, (C) XW pathway and (D) XD pathway in C. glutamicum. Values normalised to 100% xylose uptake rate (mmol g−1 h−1). | ||
6.2 Escherichia coli
E. coli is more suitable for achieving higher biomass yields from xylose as seen in Table 6 due to its wider flexibility. The two transhydrogenases demonstrate their advantage in E. coli's added metabolic flexibility. About 8% more biomass can be achieved for all the different pathways in E. coli compared to C. glutamicum. Similar theoretical maximum yields of succinate were observed in E. coli when compared to C. glutamicum. E. coli naturally harbours the XI pathway which enables it to assimilate xylose naturally.185 Several anaerobic strategies and metabolic engineering routes have been reported for enhanced succinic acid production in E. coli on different carbon sources.186 Only aerobic related succinic acid production strategies are depicted in Fig. 8. Under anaerobic conditions, pyruvate carboxylase or PEP carboxykinase overexpression together with succinate dehydrogenase deletion were proven to be efficient targets for enhanced succinic acid production implementing mainly the reductive TCA cycle. The shown optimal strategies using xylose are under aerobic conditions. As seen from the optimal strategies, owing to the flexibility of metabolism i.e. balancing reducing equivalents in E. coli, strategies were observed for both reductive as well as oxidative TCA cycle routes. As seen in Fig. 8, both the XR–XDH and XI pathways rely on the KDPG pathway and NADPH is mainly generated via malic enzyme for the XR–XDH pathway. Excess NADPH in converted back to NADH in the XI pathway via the soluble transhydrogenase. Similar to that observed in C. glutamicum, the XW pathway requires less O2 per mol xylose assimilated and produces less CO2 with a theoretical maximum yield of 1 mol succinate per mol xylose. To date, XW and XD pathways have not been implemented in E. coli to produce succinic acid. When the XD pathway is implemented a potential overexpression target would be the glyoxylate cycle, but as shown in Fig. 8, higher O2 demand and carbon loss in the form of CO2 will make this pathway inefficient for producing succinate from xylose.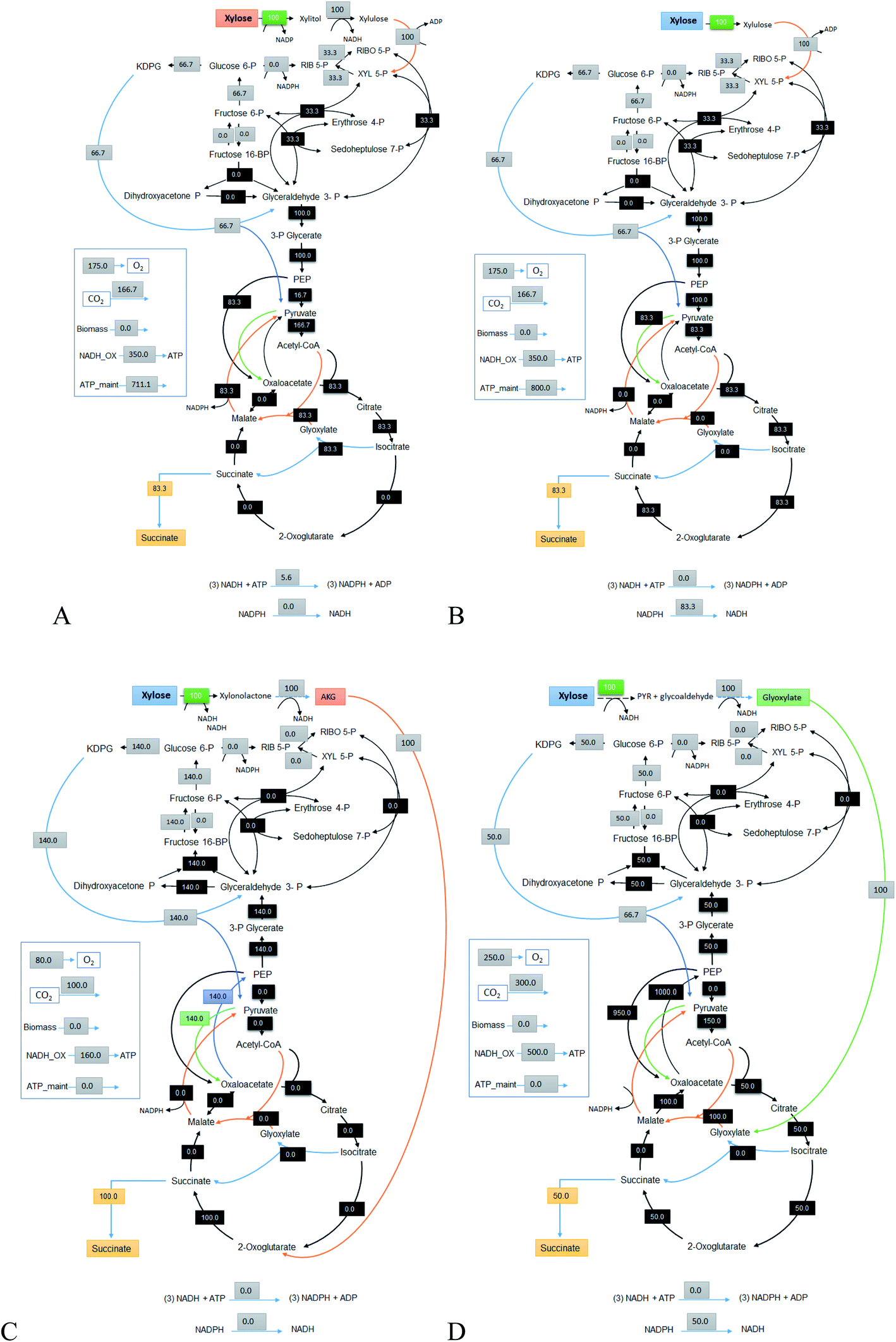 | ||
| Fig. 8 Theoretical maximum (optimal) production of SA via (A) the XR–XDH pathway, (B) XI pathway, (C) XW pathway and (D) XD pathway in E. coli. Values normalised to 100% xylose uptake rate (mmol g−1 h−1). | ||
6.3 Actinobacillus succinogenes
The acid tolerant strain A. succinogenes is known for its high production capacity of succinic acid. This facultative anaerobic bacterium can assimilate both C6 and C5 sugars. Certain studies also showed that higher biomass and succinic acid production was observed when CO2 and/or H2 are supplied additionally.187A. succinogenes does not harbour a complete TCA cycle and it is an auxotroph of glutamine, methionine and cysteine. A detailed flux analysis has been performed on A. succinogenes giving insights into its metabolism.188 It is also identified that it does not comprise the glyoxylate cycle. As shown in Fig. 9, both the XR–XDH and XI pathway yields are 1 mol succinate per mol xylose. Interestingly, most of the optimal succinic acid production pathways produced ethanol as a by-product indicating that there was excess NADH to be replenished. NADPH is mainly supplied by the PP pathway for XR–XDH based xylose consumption. On xylose, the transhydrogenase present in A. succinogenes would be a very essential reaction in balancing NADH and NADPH. To incorporate the XW pathway into A. succinogenes, alpha-ketoglutarate dehydrogenase and a succinyl-CoA synthetase have to be expressed. This could also eliminate the glutamine auxotroph. For the XD pathway, the glyoxylate cycle genes, isocitrate lyase and malate synthase must be expressed which would enable the uptake of glyoxylate produced from the XD pathway.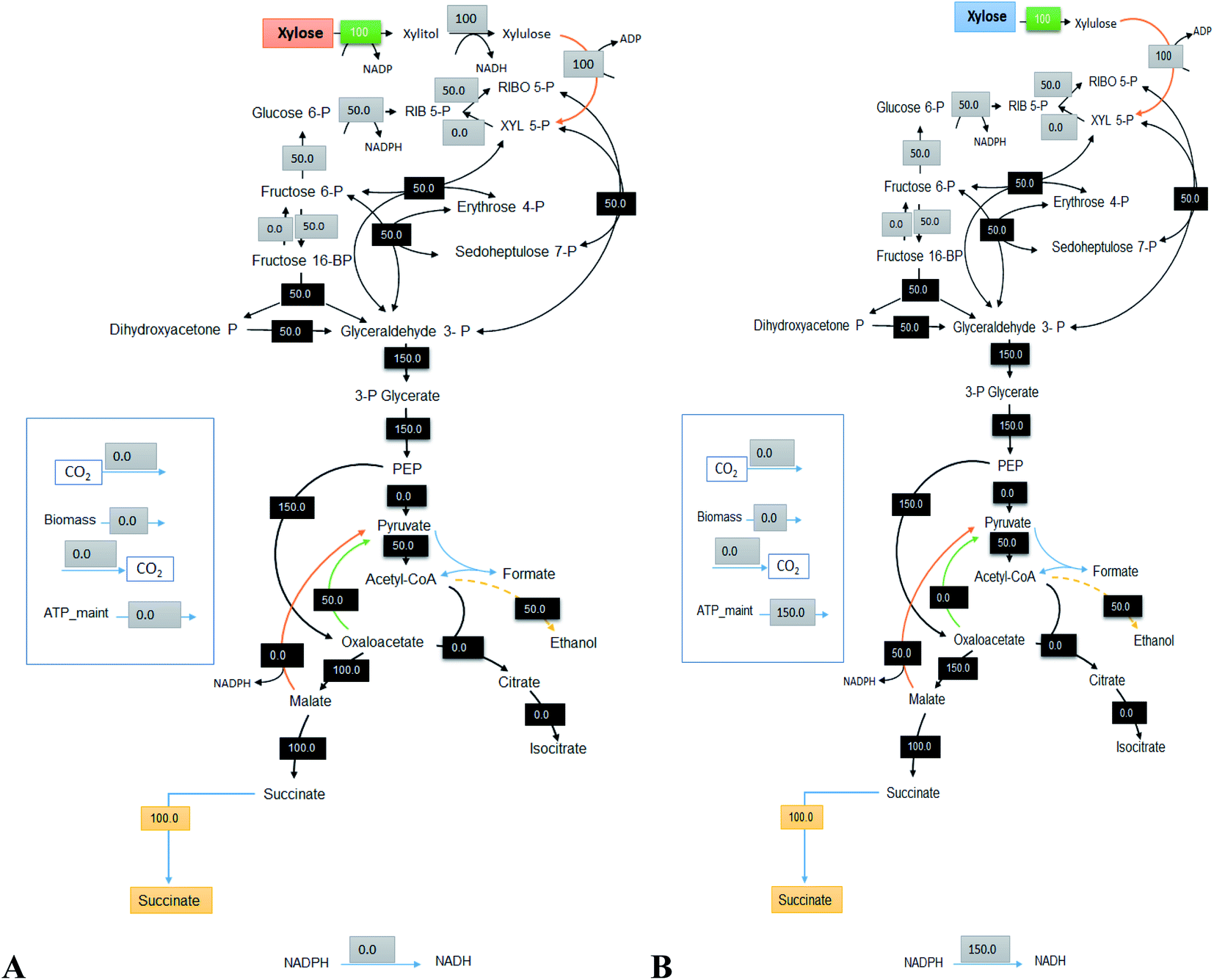 | ||
| Fig. 9 Theoretical maximum (optimal) production pathways of SA via (A) the XR–XDH pathway and (B) XI pathway in A. succinogenes. Values normalised to 100% xylose uptake rate (mmol g−1 h−1). | ||
6.4 Yarrowia lipolytica
Owing to its well-known potential in producing lipids, citric acid and other value-added products, Y. lipolytica has been demonstrated for its effectiveness in producing succinic acid from xylose.144 Availability of genetic tools and engineering for a wider substrate spectrum, tolerance at reduced pH and flexible metabolism makes this yeast a potential cell factory for succinic acid production. As depicted in Fig. 10, the optimal production of succinic acid can reach maximum yields of 1 c-mol succinic acid per c-mol xylose in XR–XDH and XI pathways. The PP pathway is shown to be the main NADPH supplying route for xylose uptake in the XR–XDH pathway which is also observed during lipid production.189 O2 demand is significantly lower per mol xylose when compared to the bacterial O2 demand for succinic acid production. Interestingly, the XW pathway is not the optimal pathway for producing succinic acid in Y. lipolytica compared to what has been observed in E. coli and C. glutamicum. Carbon loss in the form of CO2 is observed in the XW pathway in Y. lipolytica. As also observed, the bacterial XW and XD pathways require higher O2 compared to the XR–XDH and XI pathways. Addition of external CO2 to A. succinogenes and the consumption of this CO2 by PEPC or PYC will reduce the overall succinic acid yield (0.56 c-mol c-mol−1). The reductive TCA cycle is probably the best option, but optimal strategies can also be envisaged as observed in E. coli or C. glutamicum optimal pathways. As observed previously, an enhanced flux through the PP pathway especially for the XR–XDH pathway would be beneficial for enhanced xylose uptake and succinic acid production.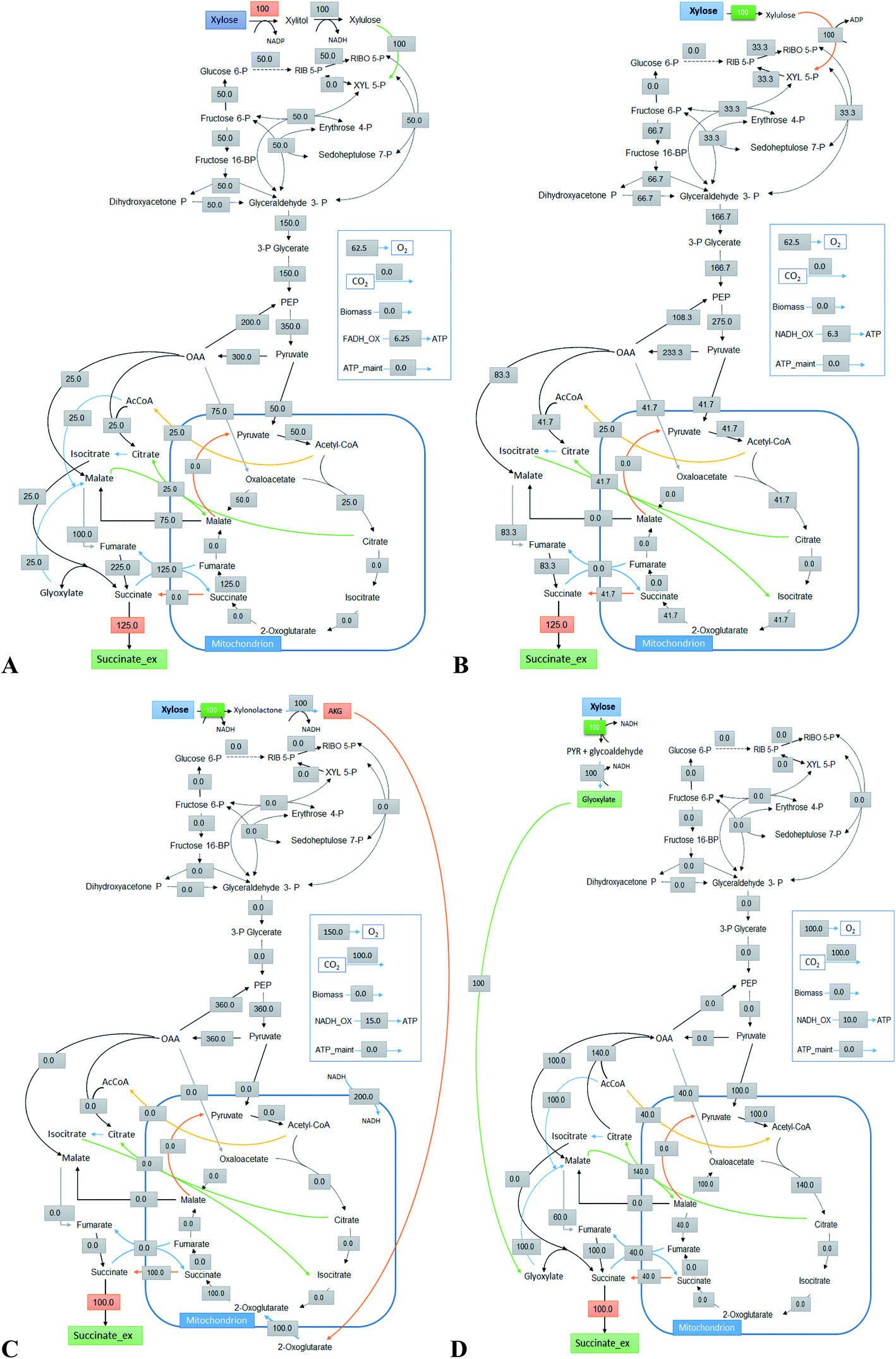 | ||
| Fig. 10 Theoretical maximum (optimal) production of SA via (A) the XR–XDH pathway, (B) XI pathway, (C) XW pathway and (D) XD pathway in Y. lipolytica. Values normalised to 100% xylose uptake rate (mmol g−1 h−1). | ||
7. Challenges for xylose based bioproduction
Xylose is a renewable sugar with great potential but has been overlooked due to low metabolic capabilities and process limitations. Although various bacteria, yeast and fungi do assimilate xylose, they utilize it in a hierarchical fashion, and these are the bottlenecks that are limiting the commercial application.7.1 Inefficient transport of pentose or absence of xylose specific transporters in the microbial cell
Xylose transport into bacterial, yeast or fungal cells through native or heterologous transporters was explained in Section 4.1. In all these microorganisms, the major limiting factor and a prerequisite objective to be addressed is the xylose uptake rate or transport efficiency of the individual cell. Most of the attempts to improve the xylose utilization efficiency of the native transporter or heterologous expression were made in E. coli and S. cerevisiae, but still the results are incomparable to glucose uptake rates. For example, the glucose uptake rate in an S. cerevisiae cell is 0.085 C-mol gCDW−1 h−1, while for xylose it is approximately three times slower (0.027 C-mol gCDW−1 h−1). In recent years, with the availability of advanced systems/synthetic biology tools and metabolic engineering techniques, there lies a scope of either engineering native promoters or investigating novel high efficiency xylose transporters through genome mining.Relative modifications in genotype and phenotype can be achieved by subjecting the microbial consortia to selective pressure.190 Radek et al. (2017) developed an automated and miniaturized ALE approach based on repetitive batch cultivations in microtiter plates. They subjected Corynebacterium glutamicum pEKEx3-xylXABCDCc bearing the Weimberg (WMB) pathway to ALE for improving xylose consumption. The evolved strain showed a 260% increase in xylose consumption efficiency.191 Overexpression of the S. stipitis Sut1 xylose transporter gene in S. cerevisiae improved the xylose uptake rate and ethanol yield by 25 and 17%, respectively, while xylose assimilation was enhanced by 25 and 40% with the introduction of Arabidopsis thaliana xylose transporter genes At5g17010 and At5g59250, respectively.192 A large improvement (75%) in xylose transport was achieved with Gxf1, a MFS transporter identified from Candida intermedia.193 This traditional xylose transporter displayed improved xylose uptake efficiency at lower xylose concentrations (∼10 g L−1) while at higher levels, the efficiency was reduced drastically. Later, comparative genome analysis of C. sojae revealed the presence of two xylose specific transporters encoded by Cs3894 and Cs4130 genes, exhibiting a substantial xylose uptake rate at concentrations up to 50 g L−1.193 The quest for novel and efficient xylose specific transporters and expression of those xylose transporters could allow rapid transport of xylose, bypass the glucose mediated repression mechanisms and enable simultaneous fermentation of mixed sugars.194
7.2 Glucose imposed carbon catabolite repression
After xylose transport, the next challenge with xylose-based cell factories is the phenomenon of carbon catabolite repression or glucose mediated inhibition which impedes the simultaneous consumption of xylose and glucose. So, the first question that comes to mind is why we need to supplement both glucose and xylose together. LCB is the most abundant material, and the abundance is so high that it can replace all the carbon coming from fossil sources. The production and process economics of LCB-biorefineries can be improved if the host strain can simultaneously utilize both glucose and xylose saccharified from the LCB. The three basic mechanisms mediating CCR have been explained in Section 3.3 (Fig. 4). As per EIIAGlc mediated catabolite repression, high levels of cAMP activate the expression of genes responsible for the metabolism of non-glucose sugars. In a study by Ammar et al. (2018) when the culture medium was supplemented with 5–10 mM cAMP, CCR was not observed in glucose–galactose co-fermentation, but 10 mM cAMP concentration was not enough to overcome CCR for simultaneous utilization of glucose and xylose.727.3 Regulation of intracellular xylose metabolism
After passing the gateway and co-substrate mediated repression, the next obstacle to address would be the slow rate of biochemical reactions using xylose and its derivatives as substrates making the overall process sluggish. That is why cell growth and metabolite production rates on xylose are slower in comparison to glucose. S. cerevisiae has been in commercial use and is known to be the best hexose utilizer, the strain lacks an active xylose utilization pathway and most of the studies were concentrated on heterologous expression of the S. stipitis xylose assimilatory pathway in S. cerevisiae, but the results observed were not satisfactory.195S. cerevisiae, for example, has a specific growth rate of 0.25 h−1 on glucose vs. 0.05 h−1 on xylose, a nearly 5-fold difference that not only limits the biomass, but also influences cell physiology and metabolism. This indicates insufficient understanding of the metabolic network and to decode this problem, it is very important to have deep understanding of the kinetics of xylose related reactions and underlying complex regulation as in vivo activity is burdened with several types of regulation. Hence, exploring the innate regulatory mechanisms of native and non-native xylose assimilatory microorganisms, and rational design and metabolic engineering leading to optimal metabolic flux and energy metabolism during the xylose assimilation is necessary.7.4 Maintaining the redox homeostasis
It is challenging for most biological processes to maintain redox homeostasis. Any deficiency of redox cofactors leads to expression of alternative metabolic pathways leading to by-product synthesis. In the xylose oxidoreductive (XR–XDH) pathway, regeneration of NAD+ is very important to direct the carbon flux into central carbon metabolism to allow smooth xylose assimilation. The imbalance between the enzymatic activities of XR and XDH results in NAD+ limitation which leads to xylitol accumulation. Under aerobic conditions, the microbial cell has the ability to regenerate NAD+, while oxygen limited or anaerobic conditions cause a shortage of NAD+ supply. Thus, maintaining optimal oxygen levels or alternative routes to generate NAD+ without interfering with the fermentation capability of the microbial strain is very important to facilitate xylose metabolism. Carlos Roseiro and associates observed the relationship between the volumetric oxygen transfer coefficient (KLa), a parameter which is a reflection of the ease of oxygen supply, and the xylitol production in a yeast, Debaryomyces hansenii. They found that increasing the KLa (h−1) from 0.2 to 1.8 caused an improvement in the xylitol accumulation, and an increase beyond 1.8 boosted the ethanol production.196 Bonan et al. (2020) attained highest ethanol titers of 28.6 g L−1 with 0.31 g g−1 yield and 1.12 g L−1 h−1 productivity with Spathaspora passalidarum at KLa (h−1) of 45.54 Hence, an ideal process engineering aspect would be to determine the optimal KLa value which maximizes the xylose flux towards the desired product with minimal or no secretion of xylitol, eventually benefiting the cell growth and product formation. Alternative approaches such as overexpression of NOX, modification of the cofactor specificity of XR, or expression of NADH specific XR over NADPH-dependent XR can alleviate the problem of redox imbalance and replenish the flux towards central carbon metabolism.197 Most of them have been attempted and only limited success has been achieved so far.8. Conclusion and future perspectives
Xylose is a readily available sugar with potential to serve as feedstock for biorefineries. For the economic viability of lignocellulose biorefineries, the efficient conversion of hemicellulosic sugars into value-added products is mandatory. Glucose-based commercially developed bioprocesses are prevalent while xylose-based ones are evolving at an industrial scale. Recent developments in biomass pretreatment technologies have led the way to extract xylose from the hemicellulosic fraction of plant cell walls with desired yields with a small amount of plant cell wall inhibitors. In nature, the xylose metabolising microorganisms are scanty compared to those metabolising glucose. Therefore, bioprospecting of novel microorganisms that could assimilate xylose separately or in combination with glucose with faster conversion rates will significantly promote efficiency of LCB-based biorefineries. However, the xylose uptake rates of the well-known xylose assimilating microorganisms are significantly lower than those assimilating glucose. Despite the exemplary developments in xylose bioconversion, there are still several challenges which need to be fixed for developing efficient microbial cell factories for high level manufacturing of biochemicals and biofuels. These challenges include efficient xylose transportation into microbial cells, faster uptake & metabolism of xylose similar to glucose, continuous availability of redox cofactors for maintaining homeostasis, glucose repression during co-fermentation, and feedback, substrate, and product mediated inhibition. Recent advancements in enzyme/metabolic/pathway engineering along with system/synthetic biology approaches have been employed to overcome these challenges but have been met with limited success. Though, xylose-based bioproduction has shown significant progress in the last few decades, many obstacles still need to be addressed to realize xylose as a feedstock at the industrial level.List of abbreviations
| ATP | Adenosine triphosphate |
| BDO | 2,3-Butanediol |
| cAMP | 3′,5′-Cyclic adenosine monophosphate |
| CBP | Consolidated bioprocessing |
| ccpA | Catabolite control protein A |
| CRE | Catabolite repressive element |
| CRP | cAMP receptor protein |
| DA | Dilute acid |
| DHA | Dihydroxyacetone |
| DHAP | Dihydroxyacetone phosphate |
| DXD | D-Xylose dehydrogenase |
| EMP pathway | Embden–Meyerhof–Parnas pathway |
| E4P | Erythrose-4-phosphate |
| F6P | Fructose-6-phosphate |
| GlpF | Glycerol facilitator protein |
| GlfZ | Glucose transporter |
| HT | Hydrothermal |
| K La | Volumetric oxygen transfer coefficient |
| K m | Michaelis–Menten constant |
| LA | Lactic acid |
| LAB | Lactic acid bacteria |
| LCB | Lignocellulosic biomass |
| LHW | Liquid hot water |
| NADH | Nicotinamide adenine dinucleotide |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| SE | Steam explosion |
| P A/PE | Promoters |
| PEP | Phosphoenol pyruvate |
| PHB | Polyhydroxybutyrate |
| PLA | Polylactic acid |
| PPP | Pentose phosphate pathway |
| SA | Succinic acid |
| TCA | Tricarboxylic acid |
| V max | Maximum velocity of an enzyme mediated reaction |
| X1P | Xylulose-1-phosphate |
| X5P | Xylulose-5-phosphate |
| XDH | Xylitol dehydrogenase |
| XI | Xylose isomerase |
| XK | Xylulose kinase |
| XR | Xylose reductase |
| Xyl1 | Xylose reductase gene |
| Xyl2 | Xylitol dehydrogenase gene |
| XylA | Xylose isomerase gene |
| XylB/Xyl3 | Xylulose kinase gene |
| XylC | Xylonolactonase |
| XylD | Xylonate dehydratase |
| XylX | 2-Keto-3-deoxy-xylonate dehydratase |
Data availability
Data sharing is not applicable to this Review article as no new data were created or analysed in this study.Author contributions
NV: conceptualization, writing-original draft, reviewing & editing; RC, RB, DA, EA, KKP and AKC: writing-original draft, reviewing & editing; SKB, DK, PB and VG: reviewing & editing; VK: conceptualization, writing-original draft, reviewing & editing and project management.Conflicts of interest
The authors declare that they have no competing interests.Acknowledgements
This study was financially supported through the vWa Project (Grant BB/S011951/1) and we acknowledge BBSRC, Innovate UK and Department of Biotechnology, India for funding this project. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.References
- V. Balan, ISRN Biotechnol., 2014, 2014, 1–31 CrossRef PubMed.
- A. J. Ragauskas, G. T. Beckham, M. J. Biddy, R. Chandra, F. Chen, M. F. Davis, B. H. Davison, R. A. Dixon, P. Gilna, M. Keller, P. Langan, A. K. Naskar, J. N. Saddler, T. J. Tschaplinski, G. A. Tuskan and C. E. Wyman, Sci. 80, 2014, 344, 1246843 CrossRef PubMed.
- B. Satari, K. Karimi and R. Kumar, Sustain. Energy Fuels, 2019, 3, 11–62 RSC.
- S. H. Hazeena, R. Sindhu, A. Pandey and P. Binod, Bioresour. Technol., 2020, 302, 122873 CrossRef CAS PubMed.
- M. Dehghanzad, M. Shafiei and K. Karimi, Renewable Energy, 2020, 158, 332–342 CrossRef CAS.
- V. Kumar, P. Binod, R. Sindhu, E. Gnansounou and V. Ahluwalia, Bioresour. Technol., 2018, 269, 443–451 CrossRef CAS PubMed.
- N. Vivek, M. Christopher, M. K. Kumar, E. Castro, P. Binod and A. Pandey, Renewable Energy, 2018, 129, 794–799 CrossRef CAS.
- J. M. Gancedo, Eur. J. Biochem., 1992, 206, 297–313 CrossRef CAS PubMed.
- F. M. Gírio, F. Carvalheiro, L. C. Duarte and R. Bogel-ŁUkasik, in D-xylitol: Fermentative Production, Application and Commercialization, Springer-Verlag Berlin Heidelberg, 2012, pp. 3–37 Search PubMed.
- S. I. Mussatto and G. M. Dragone, in Biomass Fractionation Technologies for a Lignocellulosic Feedstock Based Biorefinery, Elsevier Inc., 2016, pp. 1–22 Search PubMed.
- L. R. Lynd, C. E. Wyman and T. U. Gerngross, Biotechnol. Prog., 1999, 15, 777–793 CrossRef CAS PubMed.
- M. E. Himmel, S. Y. Ding, D. K. Johnson, W. S. Adney, M. R. Nimlos, J. W. Brady and T. D. Foust, Sci. 80, 2007, 315, 804–807 CrossRef CAS PubMed.
- R. Kumar, G. Mago, V. Balan and C. E. Wyman, Bioresour. Technol., 2009, 100, 3948–3962 CrossRef CAS PubMed.
- N. Uppugundla, L. Da Costa Sousa, S. P. S. Chundawat, X. Yu, B. Simmons, S. Singh, X. Gao, R. Kumar, C. E. Wyman, B. E. Dale and V. Balan, Biotechnol. Biofuels, 2014, 7(1), 1–14 CrossRef PubMed.
- K. Yao, Q. Wu, R. An, W. Meng, M. Ding, B. Li and Y. Yuan, AIChE J., 2018, 64, 1938–1953 CrossRef CAS.
- D. Steinbach, A. Kruse, J. Sauer and J. Storz, Processes, 2020, 8, 1–12 Search PubMed.
- J. C. Solarte-Toro, Y. Chacón-Pérez, S. Piedrahita-Rodríguez, J. A. Poveda Giraldo, J. A. Teixeira, K. Moustakas and C. A. C. Alzate, Energy, 2020, 195, 116986 CrossRef CAS.
- F. Rodríguez, A. Sanchez and L. Amaya-Delgado, Ind. Crops Prod., 2019, 134, 62–70 CrossRef.
- J. S. Kim, Y. Y. Lee and T. H. Kim, Bioresour. Technol., 2016, 199, 42–48 CrossRef CAS PubMed.
- Y. Zhang, Y. Y. Liu, J. L. Xu, Z. H. Yuan, W. Qi, X. S. Zhuang and M. C. He, Bioresources, 2012, 7, 345–353 CAS.
- S. F. Fu, K. Q. Chen, R. Zhu, W. X. Sun, H. Zou and R. B. Guo, Energy Convers. Manag., 2018, 159, 121–128 CrossRef CAS.
- Z. Gong, X. Wang, W. Yuan, Y. Wang, W. Zhou, G. Wang and Y. Liu, Biotechnol. Biofuels, 2020, 13, 1–15 CrossRef PubMed.
- Q. Ibrahim and A. Kruse, Bioresour. Technol. Rep., 2020, 11, 100506 CrossRef.
- A. Smit and W. Huijgen, Green Chem., 2017, 19, 5505–5514 RSC.
- J. Xu, Y. Fu, G. Tian, Q. Li, N. Liu, M. Qin and Z. Wang, Bioresour. Technol., 2018, 254, 353–356 CrossRef CAS PubMed.
- Z. Chen, W. D. Reznicek and C. Wan, ACS Sustainable Chem. Eng., 2018, 6, 6910–6919 CrossRef CAS.
- G. Wang, S. Qi, Y. Xia, A. M. Parvez, C. Si and Y. Ni, ACS Sustainable Chem. Eng., 2020, 8, 2772–2782 CrossRef CAS.
- Q. Zhang, Y. Deng, X. Tan, W. Wang, Q. Yu, X. Chen, C. Miao, Y. Guo, Y. Zhang, X. Zhuang and Z. Yuan, Ind. Crops Prod., 2020, 145, 112091 CrossRef CAS.
- X. Zhang, W. Zhang, F. Lei, S. Yang and J. Jiang, Bioresour. Technol., 2020, 309, 123385 CrossRef CAS PubMed.
- C. Cai, K. Hirth, R. Gleisner, H. Lou, X. Qiu and J. Y. Zhu, Green Chem., 2020, 22, 1605–1617 RSC.
- T. Y. Nguyen, C. M. Cai, O. Osman, R. Kumar and C. E. Wyman, Green Chem., 2016, 18, 1581–1589 RSC.
- C. M. Cai, T. Zhang, R. Kumar and C. E. Wyman, J. Chem. Technol. Biotechnol., 2014, 89, 2–10 CrossRef CAS.
- J. Jae, G. A. Tompsett, Y.-C. Lin, T. R. Carlson, J. Shen, T. Zhang, B. Yang, C. E. Wyman, W. C. Conner and G. W. Huber, Energy Environ. Sci., 2010, 3, 358–365 RSC.
- L. T. Mika, E. Cséfalvay and Á. Németh, Chem. Rev., 2018, 118, 505–613 CrossRef CAS PubMed.
- N. W. Dulie, B. Woldeyes, H. D. Demsash and A. S. Jabasingh, Waste Biomass Valorization, 2021, 12, 531–552 CrossRef CAS.
- J. Cui, J. Tan, X. Cui, Y. Zhu, T. Deng, G. Ding and Y. Li, ChemSusChem, 2016, 9, 1259–1262 CrossRef CAS PubMed.
- X. Hu, Y. Song, L. Wu, M. Gholizadeh and C. Z. Li, ACS Sustainable Chem. Eng., 2013, 1, 1593–1599 CrossRef CAS.
- D. Dasgupta, S. Bandhu, D. K. Adhikari and D. Ghosh, Microbiol. Res., 2017, 197, 9–21 CrossRef CAS PubMed.
- R. F. Perez, O. S. G. P. Soares, A. M. D. de Farias, M. F. R. Pereira and M. A. Fraga, Appl. Catal., B, 2018, 232, 101–107 CrossRef CAS.
- J. Wisniak, M. Hershkowitz, R. Leibowitz and S. Stein, Ind. Eng. Chem. Prod. Res. Dev., 1974, 13, 75–79 CrossRef CAS.
- S. Kwak, J. H. Jo, E. J. Yun, Y. S. Jin and J. H. Seo, Biotechnol. Adv., 2019, 37, 271–283 CrossRef CAS PubMed.
- C. Zhao, Y. Zhang and Y. Li, Biotechnol. Adv., 2019, 37, 107402 CrossRef CAS.
- P. M. Bruinenberg, P. H. M. de Bot, J. P. van Dijken and W. A. Scheffers, Eur. J. Appl. Microbiol. Biotechnol., 1983, 18, 287–292 CrossRef CAS.
- C. De Macario, D. B. A. E. Miov and T. W. Jeffries, Trends Biotechnol., 1985, 3, 208–212 CrossRef.
- Z. Zheng, X. Lin, T. Jiang, W. Ye and J. Ouyang, Biotechnol. Lett., 2016, 38, 1331–1339 CrossRef CAS PubMed.
- L. Fan, Y. Zhang, W. Qu, J. Wang and W. Shao, Biotechnol. Lett., 2011, 33, 593–598 CrossRef CAS PubMed.
- R. Y. Miyamoto, A. S. de Sousa, P. S. Vieira, R. R. de Melo, J. A. Scarpassa, C. H. I. Ramos, M. T. Murakami, R. Ruller and L. M. Zanphorlin, Biochim. Biophys. Acta, Gen. Subj., 2020, 1864, 129549 CrossRef CAS PubMed.
- D. Yang, S. Y. Park, Y. S. Park, H. Eun and S. Y. Lee, Trends Biotechnol., 2020, 1–21 Search PubMed.
- K. Karhumaa, R. G. Sanchez and B. Hahn-hägerdal, Microb. Cell Fact., 2007, 10, 1–10 Search PubMed.
- P. M. Bruinenberg, P. H. M. de Bot, J. P. van Dijken and W. A. Scheffers, Appl. Microbiol. Biotechnol., 1984, 19, 256–260 CrossRef CAS.
- H. F. Son, S. Lee and K. Kim, Sci. Rep., 2018, 1–11 Search PubMed.
- O. Bengtsson, B. Hahn-Hägerdal and M. F. Gorwa-Grauslund, Biotechnol. Biofuels, 2009, 2, 1–10 CrossRef PubMed.
- A. Mouro, A. A. Santos, D. D. Agnolo, G. F. Gubert, E. P. S. Bon, C. A. Rosa, C. Fonseca and B. U. Stambuk, Fermentation, 2020, 6, 1–14 CrossRef.
- C. I. D. G. Bonan, L. E. Biazi, S. R. Dionísio, L. B. Soares, R. Tramontina, A. S. Sousa, C. A. de Oliveira Filho, A. C. Costa and J. L. Ienczak, Bioprocess Biosyst. Eng., 2020, 43, 1509–1519 CrossRef CAS PubMed.
- A. A. Prabhu, E. Bosakornranut, Y. Amraoui, D. Agrawal, F. Coulon, V. Vivekanand, V. K. Thakur and V. Kumar, Biotechnol. Biofuels, 2020, 13, 1–15 CrossRef PubMed.
- D. K. Wilson, K. L. Kavanagh, M. Klimacek and B. Nidetzky, Chem.–Biol. Interact., 2003, 144, 515–521 CrossRef.
- K. Correia, A. Khusnutdinova, P. Y. Li, J. C. Joo, G. Brown, A. F. Yakunin and R. Mahadevan, bioRxiv, 2018, 1–28, 390401 Search PubMed.
- F. R. Rech, R. C. Fontana, C. A. Rosa, M. Camassola, M. A. Z. Ayub and A. J. P. Dillon, Bioprocess Biosyst. Eng., 2019, 42, 83–92 CrossRef CAS PubMed.
- P. Kijtter, R. Amore, C. P. Hollenberg, M. Ciriacy, I. Mikrobiologie, W. Dfisseldorf and F. Republic, Curr. Genet., 1990, 1, 493–500 Search PubMed.
- S. Watanabe, A. A. Saleh, S. P. Pack, N. Annaluru, T. Kodaki and K. Makino, Microbiology, 2007, 153, 3044–3054 CrossRef CAS PubMed.
- R. Lunzer, Y. Mamnun, D. Haltrich, K. D. Kulbe and B. Nidetzky, Biochem. J., 1998, 99, 91–99 CrossRef PubMed.
- L. B. Lockwood and G. E. N. Nelson, J. Bacteriol., 1946, 52(5), 581–586 CrossRef CAS PubMed.
- R. Weimberg, J. Biol. Chem., 1961, 236, 629–635 CrossRef CAS.
- H. Almqvist, S. J. Glaser, C. Tufvegren, L. Wasserstrom and G. Lidén, Fermentation, 2018, 4, 1–11 CrossRef.
- K. A. K. Köhler, L. M. Blank, O. Frick and A. Schmid, Environ. Microbiol., 2015, 17, 156–170 CrossRef PubMed.
- L. Shen, M. Kohlhaas, J. Enoki, R. Meier, B. Schönenberger, R. Wohlgemuth, R. Kourist, F. Niemeyer, D. van Niekerk, C. Bräsen, J. Niemeyer, J. Snoep and B. Siebers, Nat. Commun., 2020, 11, 1–13 Search PubMed.
- K. N. G. Valdehuesa, K. R. M. Ramos, G. M. Nisola, A. B. Bañares, R. B. Cabulong, W. K. Lee, H. Liu and W. J. Chung, Appl. Microbiol. Biotechnol., 2018, 102, 7703–7716 CrossRef CAS PubMed.
- I. Bator, A. Wittgens, F. Rosenau, T. Tiso and L. M. Blank, Front. Bioeng. Biotechnol., 2020, 7, 1–18 Search PubMed.
- R. B. Cabulong, W. K. Lee, A. B. Bañares, K. R. M. Ramos, G. M. Nisola, K. N. G. Valdehuesa and W. J. Chung, Appl. Microbiol. Biotechnol., 2018, 102, 2179–2189 CrossRef CAS PubMed.
- W. Wei, P. Zhang, Y. Shang, Y. Zhou and B. C. Ye, Bioresour. Technol., 2020, 314, 123726 CrossRef CAS PubMed.
- T. W. Jeffries, Curr. Opin. Biotechnol., 2006, 17, 320–326 CrossRef CAS PubMed.
- E. M. Ammar, X. Wang and C. V. Rao, Sci. Rep., 2018, 8, 1–11 CAS.
- Y. Gu, Y. Ding, C. Ren, Z. Sun, D. A. Rodionov, W. Zhang, S. Yang, C. Yang and W. Jiang, BMC Genomics, 2010, 11, 1–14 CrossRef PubMed.
- R. E. Hector and J. A. Mertens, Mol. Biotechnol., 2017, 59, 24–33 CrossRef CAS PubMed.
- C. Sievert, L. M. Nieves, L. A. Panyon, T. Loeffler, C. Morris, R. A. Cartwright, X. Wang and A. L. Demain, Proc. Natl. Acad. Sci. U. S. A., 2017, 114, 7349–7354 CrossRef CAS PubMed.
- J. G. Nijland, H. Y. Shin, R. M. De Jong, P. P. De Waal, P. Klaassen and A. J. M. Driessen, Biotechnol. Biofuels, 2014, 7, 1–11 CrossRef PubMed.
- K. A. Erlandson, J. H. Park, W. El Khal, H. H. Kao, P. Basaran, S. Brydges and C. A. Batt, Appl. Environ. Microbiol., 2000, 66, 3974–3980 CrossRef CAS PubMed.
- C. Sizemore, E. Buchner, T. Rygus, C. Witke, F. Götz and W. Hillen, MGG, Mol. Gen. Genet., 1991, 227, 377–384 CrossRef CAS PubMed.
- T. J. Verbeke, R. J. Giannone, D. M. Klingeman, N. L. Engle, T. Rydzak, A. M. Guss, T. J. Tschaplinski, S. D. Brown, R. L. Hettich and J. G. Elkins, Sci. Rep., 2017, 7, 1–11 CrossRef PubMed.
- N. Vivek, L. M. Nair, B. Mohan, S. C. Nair, R. Sindhu, A. Pandey, N. Shurpali and P. Binod, Bioresour. Technol. Rep., 2019, 7, 100224 CrossRef.
- B. Görke and J. Stülke, Nat. Rev. Microbiol., 2008, 6, 613–624 CrossRef PubMed.
- G. Aidelberg, B. D. Towbin, D. Rothschild, E. Dekel, A. Bren and U. Alon, BMC Syst. Biol., 2014, 8(1), 1–12 CrossRef PubMed.
- A. Ullmann and J. Monod, FEBS Lett., 1968, 2, 57–60 CrossRef CAS PubMed.
- J. E. Gonzalez and A. Peterkofsky, Prog. Clin. Biol. Res., 1978, 22, 325–332 Search PubMed.
- S. Kim and J. Hahn, Metab. Eng., 2015, 1–7, 94–101 CrossRef PubMed.
- T. Rygus and W. Hillen, J. Bacteriol., 1992, 174, 3049–3055 CrossRef CAS PubMed.
- M. Bruder, M. Moo-Young, D. A. Chung and C. P. Chou, Appl. Microbiol. Biotechnol., 2015, 99, 7579–7588 CrossRef CAS PubMed.
- Y. Li, K. Jin, L. Zhang, Z. Ding, Z. Gu and G. Shi, J. Agric. Food Chem., 2018, 66, 9456–9464 CrossRef CAS PubMed.
- P. Chaillou, P. H. Pouwels and P. W. Postma, J. Bacteriol., 1999, 181, 4768–4773 CrossRef PubMed.
- T. W. Jeffries, Bioenergy, 2014, 37–47 Search PubMed.
- K. L. Dunn and C. V. Rao, Appl. Microbiol. Biotechnol., 2014, 98, 6897–6905 CrossRef CAS PubMed.
- A. Farwick, S. Bruder, V. Schadeweg, M. Oreb and E. Boles, Proc. Natl. Acad. Sci. U. S. A., 2014, 111, 5159–5164 CrossRef CAS PubMed.
- M. Jeppsson, O. Bengtsson, K. Franke, H. Lee, B. Hahn-Hägerdal and M. F. Gorwa-Grauslund, Biotechnol. Bioeng., 2006, 93, 665–673 CrossRef CAS PubMed.
- S. G. Kilian and N. van Uden, Appl. Microbiol. Biotechnol., 1988, 27, 545–548 CrossRef CAS.
- C. Wang, X. Bao, Y. Li, C. Jiao, J. Hou, Q. Zhang, W. Zhang, W. Liu and Y. Shen, Metab. Eng., 2015, 30, 79–88 CrossRef CAS PubMed.
- E. M. Young, A. D. Comer, H. Huang and H. S. Alper, Metab. Eng., 2012, 14, 401–411 CrossRef CAS PubMed.
- E. M. Young, A. Tong, H. Bui, C. Spofford and H. S. Alper, Proc. Natl. Acad. Sci. U. S. A., 2014, 111, 131–136 CrossRef CAS PubMed.
- A. Singh, M. D. Lynch and R. T. Gill, Metab. Eng., 2009, 11, 347–354 CrossRef CAS PubMed.
- J. Hou, N. F. Lages, M. Oldiges and G. N. Vemuri, Metab. Eng., 2009, 11, 253–261 CrossRef CAS PubMed.
- J. Hou, G. N. Vemuri, X. Bao and L. Olsson, Appl. Microbiol. Biotechnol., 2009, 82, 909–919 CrossRef CAS PubMed.
- C. Zhao, Q. Zhao, Y. Li and Y. Zhang, Microb. Cell Fact., 2017, 16, 1–11 CrossRef PubMed.
- B. Bergdahl, D. Heer, U. Sauer, B. Hahn-Hägerdal and E. W. Van Niel, Biotechnol. Biofuels, 2012, 5, 1–19 CrossRef PubMed.
- K. Shimizu and Y. Matsuoka, Biotechnol. Adv., 2019, 37, 107441 CrossRef CAS PubMed.
- G. C. Zhang, J. J. Liu and W. T. Ding, Appl. Environ. Microbiol., 2012, 78, 1081–1086 CrossRef CAS PubMed.
- G. C. Zhang, T. L. Turner and Y. S. Jin, J. Ind. Microbiol. Biotechnol., 2017, 44, 387–395 CrossRef CAS PubMed.
- B. Petschacher and B. Nidetzky, Microb. Cell Fact., 2008, 7, 1–12 CrossRef PubMed.
- S. Watanabe, A. A. Saleh, S. P. Pack, N. Annaluru, T. Kodaki and K. Makino, Microbiology, 2007, 153, 3044–3054 CrossRef CAS PubMed.
- J. K. Lee, B. S. Koo and S. Y. Kim, Appl. Environ. Microbiol., 2003, 69, 6179–6188 CrossRef CAS PubMed.
- H. Suga, F. Matsuda, T. Hasunuma, J. Ishii and A. Kondo, Appl. Microbiol. Biotechnol., 2013, 97, 1669–1678 CrossRef CAS PubMed.
- J. Hou, G. N. Vemuri, X. Bao and L. Olsson, Appl. Microbiol. Biotechnol., 2009, 82, 909–919 CrossRef CAS PubMed.
- C. Roca, J. Nielsen and L. Olsson, Appl. Environ. Microbiol., 2003, 69, 4732–4736 CrossRef CAS PubMed.
- J. Liu, Z. Wang, V. Kandasamy, S. Y. Lee, C. Solem and P. R. Jensen, Metab. Eng., 2017, 44, 22–29 CrossRef CAS PubMed.
- C. Zhu, Q. Li, L. Pu, Z. Tan, K. Guo, H. Ying and P. Ouyang, ACS Catal., 2016, 6, 4989–4994 CrossRef CAS.
- M. Celton, I. Sanchez, A. Goelzer, V. Fromion, C. Camarasa and S. Dequin, BMC Genomics, 2012, 13(1), 1–14 CrossRef PubMed.
- S. L. Baptista, J. T. Cunha, A. Romaní and L. Domingues, Bioresour. Technol., 2018, 267, 481–491 CrossRef CAS PubMed.
- J. Trivedi, A. K. Bhonsle and N. Atray, Processing Food Waste for the Production of Platform Chemicals, Elsevier Inc., 2019 Search PubMed.
- X. Yuan, J. Wang, J. Lin, L. Yang and M. Wu, J. Ind. Microbiol. Biotechnol., 2019, 46, 1061–1069 CrossRef PubMed.
- Y. Lugani and B. S. Sooch, Lwt, 2020, 134, 109988 CrossRef CAS.
- S. Tiwari and A. Baghela, Challenges and Prospects of Xylitol Production by Conventional and Non-conventional Yeasts, Elsevier B.V., 2020 Search PubMed.
- J. Jo, S. Oh, H. Lee, Y. Park and J. Seo, Biotechnol. J., 2015, 10, 1935–1943 CrossRef CAS PubMed.
- A. A. Prabhu, D. J. Thomas, R. Ledesma-Amaro, G. A. Leeke, A. Medina, C. Verheecke-Vaessen, F. Coulon, D. Agrawal and V. Kumar, Microb. Cell Fact., 2020, 19, 1–18 CrossRef PubMed.
- J. H. Kim, K. C. Han, Y. H. Koh, Y. W. Ryu and J. H. Seo, J. Ind. Microbiol. Biotechnol., 2002, 29, 16–19 CrossRef CAS PubMed.
- C. Gao, C. Ma and P. Xu, Biotechnol. Adv., 2011, 29, 930–939 CrossRef CAS PubMed.
- Z. Qiu, Q. Gao and J. Bao, Bioresour. Technol., 2018, 249, 9–15 CrossRef CAS PubMed.
- D. Wischral, J. M. Arias, L. F. Modesto, D. de França Passos and N. Pereira, Biotechnol. Prog., 2019, 35(1), e2718 CrossRef PubMed.
- D. Parra-Ramírez, A. Martinez and C. A. Cardona, Bioresour. Technol., 2019, 273, 86–92 CrossRef PubMed.
- Z. Qiu, Q. Gao and J. Bao, Bioresour. Technol., 2017, 245, 1369–1376 CrossRef CAS PubMed.
- H. Lu, X. Zhao, Y. Wang, X. Ding, J. Wang, E. Garza, R. Manow, A. Iverson and S. Zhou, BMC Biotechnol., 2016, 16, 1–10 CrossRef PubMed.
- V. Novy, B. Brunner and B. Nidetzky, Microb. Cell Fact., 2018, 17, 1–11 CrossRef PubMed.
- T. L. Turner, G. C. Zhang, E. J. Oh, V. Subramaniam, A. Adiputra, V. Subramaniam, C. D. Skory, J. Y. Jang, B. J. Yu, I. Park and Y. S. Jin, Biotechnol. Bioeng., 2016, 113, 1075–1083 CrossRef CAS PubMed.
- Z. Dai, F. Guo, S. Zhang, W. Zhang, Q. Yang, W. Dong, M. Jiang, J. Ma and F. Xin, Biofuels, Bioprod. Biorefin., 2020, 14, 965–985 CrossRef CAS.
- Y. Chen and J. Nielsen, Curr. Opin. Biotechnol., 2016, 37, 165–172 CrossRef CAS PubMed.
- M. Jiang, J. Ma, M. Wu, R. Liu, L. Liang, F. Xin, W. Zhang, H. Jia and W. Dong, Bioresour. Technol., 2017, 245, 1710–1717 CrossRef CAS PubMed.
- J. J. Beauprez, M. De Mey and W. K. Soetaert, Process Biochem., 2010, 45, 1103–1114 CrossRef CAS.
- M. F. A. Bradfield, A. Mohagheghi, D. Salvachúa, H. Smith, B. A. Black, N. Dowe, G. T. Beckham and W. Nicol, Biotechnol. Biofuels, 2015, 8, 1–17 CrossRef PubMed.
- M. F. A. Bradfield and W. Nicol, Bioprocess Biosyst. Eng., 2016, 39, 233–244 CrossRef CAS PubMed.
- D. Salvachúa, A. Mohagheghi, H. Smith, M. F. A. Bradfield, W. Nicol, B. A. Black, M. J. Biddy, N. Dowe and G. T. Beckham, Biotechnol. Biofuels, 2016, 9, 1–15 CrossRef PubMed.
- C. Pateraki, H. Almqvist, D. Ladakis, G. Lidén, A. A. Koutinas and A. Vlysidis, Biochem. Eng. J., 2016, 114, 26–41 CrossRef CAS.
- J. Lu, Y. Lv, Y. Jiang, M. Wu, B. Xu, W. Zhang, J. Zhou, W. Dong, F. Xin and M. Jiang, ACS Sustainable Chem. Eng., 2020, 8, 9035–9045 CrossRef CAS.
- K. Jantama, X. Zhang, J. C. Moore, K. T. Shanmugam, S. A. Svoronos and L. O. Ingram, Biotechnol. Bioeng., 2008, 101, 881–893 CrossRef CAS PubMed.
- K. Jantama, M. J. Haupt, S. A. Svoronos, X. Zhang, J. C. Moore, K. T. Shanmugam and L. O. Ingram, Biotechnol. Bioeng., 2008, 99, 1140–1153 CrossRef CAS PubMed.
- P. Khunnonkwao, S. S. Jantama, S. Kanchanatawee and K. Jantama, Appl. Microbiol. Biotechnol., 2018, 102, 127–141 CrossRef CAS PubMed.
- F. Zhang, J. Li, H. Liu, Q. Liang and Q. Qi, PLoS One, 2016, 11, 1–12 CAS.
- A. A. Prabhu, R. Ledesma-Amaro, C. S. K. Lin, F. Coulon, V. K. Thakur and V. Kumar, Biotechnol. Biofuels, 2020, 13, 1–15 CrossRef PubMed.
- J. Wang, C. Li, Y. Zou and Y. Yan, Proc. Natl. Acad. Sci. U. S. A., 2020, 117, 19159–19167 CrossRef CAS PubMed.
- A. P. Zeng and W. Sabra, Curr. Opin. Biotechnol., 2011, 22, 749–757 CrossRef CAS PubMed.
- X. X. Wang, H. Y. Hu, D. H. Liu and Y. Q. Song, Nat. Biotechnol., 2016, 33, 16–22 CAS.
- Y. Amraoui, V. Narisetty, F. Coulon, D. Agrawal, A. K. Chandel, S. Maina, A. Koutinas and V. Kumar, ACS Sustainable Chem. Eng., 2021, 9, 10381–10391 CrossRef CAS.
- X. W. Guo, Y. Zhang, L. L. Li, X. Y. Guan, J. Guo, D. G. Wu, Y. F. Chen and D. G. Xiao, Biotechnol. Biofuels, 2018, 11, 1–18 CrossRef PubMed.
- J. W. Kim, S. O. Seo, G. C. Zhang, Y. S. Jin and J. H. Seo, Bioresour. Technol., 2015, 191, 512–519 CrossRef CAS PubMed.
- S. J. Kim, H. J. Sim, J. W. Kim, Y. G. Lee, Y. C. Park and J. H. Seo, Bioresour. Technol., 2017, 245, 1551–1557 CrossRef CAS PubMed.
- B. Dey, B. Roy, S. Datta and K. G. Singh, Comprehensive Overview and Proposal of Strategies for the Ethanol Sector in India, Biomass Conversion and Biorefinery, 2021 Search PubMed.
- Sujata and P. Kaushal, Biofuels, 2020, 11, 763–775 CAS.
- NITI-Aayog, Ethanol Blending in India 2020-25, 2020 Search PubMed.
- V. Menon and M. Rao, Prog. Energy Combust. Sci., 2012, 38, 522–550 CrossRef CAS.
- S. H. Lee, T. Kodaki, Y. C. Park and J. H. Seo, J. Biotechnol., 2012, 158, 184–191 CrossRef CAS PubMed.
- X. Hou, Appl. Microbiol. Biotechnol., 2012, 94, 205–214 CrossRef CAS PubMed.
- S. C. Nakanishi, L. B. Soares, L. E. Biazi, V. M. Nascimento, A. C. Costa, G. J. M. Rocha and J. L. Ienczak, Biotechnol. Bioeng., 2017, 114, 2211–2221 CrossRef CAS PubMed.
- T. F. Pacheco, B. R. C. Machado, W. G. de Morais Júnior, J. R. M. Almeida and S. B. Gonçalves, Appl. Biochem. Biotechnol., 2021, 193, 2182–2197 CrossRef CAS PubMed.
- T. Watanabe, I. Watanabe, M. Yamamoto, A. Ando and T. Nakamura, Bioresour. Technol., 2011, 102, 1844–1848 CrossRef CAS PubMed.
- U. Klinner, S. Fluthgraf, S. Freese and V. Passoth, Appl. Microbiol. Biotechnol., 2005, 67, 247–253 CrossRef CAS PubMed.
- M. Xiong, A. Woodruff, X. Tang, X. Tian, J. Zhang and L. Cao, J. Taiwan Inst. Chem. Eng., 2013, 44, 605–610 CrossRef CAS.
- L. Liu, M. Jin, M. Huang, Y. Zhu, W. Yuan, Y. Kang, M. Kong, S. Ali, Z. Jia, Z. Xu, W. Xiao and L. Cao, Front. Bioeng. Biotechnol., 2021, 9, 1–11 Search PubMed.
- S. J. Chacón, G. Matias, C. F. dos S. Vieira, T. C. Ezeji, R. Maciel Filho and A. P. Mariano, Ind. Crops Prod., 2020, 155, 112837 CrossRef.
- L. D. Gottumukkala, B. Parameswaran, S. K. Valappil, K. Mathiyazhakan, A. Pandey and R. K. Sukumaran, Bioresour. Technol., 2013, 145, 182–187 CrossRef CAS PubMed.
- N. Qureshi, B. C. Saha, R. E. Hector, B. Dien, S. Hughes, S. Liu, L. Iten, M. J. Bowman, G. Sarath and M. A. Cotta, Biomass Bioenergy, 2010, 34, 566–571 CrossRef CAS.
- N. R. Baral and A. Shah, Energy Fuels, 2016, 30, 5779–5790 CrossRef CAS.
- Y. Jiang, D. Guo, J. Lu, P. Dürre, W. Dong, W. Yan, W. Zhang, J. Ma, M. Jiang and F. Xin, Biotechnol. Biofuels, 2018, 11, 1–14 CrossRef PubMed.
- Y. Jiang, Y. Lv, R. Wu, J. Lu, W. Dong, J. Zhou, W. Zhang, F. Xin and M. Jiang, Biotechnol. Bioeng., 2020, 117, 2985–2995 CrossRef CAS PubMed.
- L. P. Guamán, C. Barba-Ostria, F. Zhang, E. R. Oliveira-Filho, J. G. C. Gomez and L. F. Silva, Microb. Cell Fact., 2018, 17, 1–11 CrossRef PubMed.
- D. Moorkoth and K. M. Nampoothiri, Bioresour. Technol., 2016, 201, 253–260 CrossRef CAS PubMed.
- J. M. Nduko, K. Matsumoto, T. Ooi and S. Taguchi, Metab. Eng., 2013, 15, 159–166 CrossRef CAS PubMed.
- L. Salamanca-Cardona, C. S. Ashe, A. J. Stipanovic and C. T. Nomura, Appl. Microbiol. Biotechnol., 2014, 98, 831–842 CrossRef CAS PubMed.
- X. Kourilova, I. Pernicova, K. Sedlar, J. Musilova, P. Sedlacek, M. Kalina, M. Koller and S. Obruca, Bioresour. Technol., 2020, 315, 123885 CrossRef CAS PubMed.
- H. S. Kim, Y. H. Oh, Y. A. Jang, K. H. Kang, Y. David, J. H. Yu, B. K. Song, J. il Choi, Y. K. Chang, J. C. Joo and S. J. Park, Microb. Cell Fact., 2016, 15, 1–13 CrossRef PubMed.
- H.-K. Jun, Y.-H. Jin, H.-N. Kim, Y.-T. Kim, S.-W. Kim and H.-S. Baik, J. Life Sci., 2008, 18, 1625–1630 CrossRef.
- L. de Souza, Y. Manasa and S. Shivakumar, Biocatal. Agric. Biotechnol., 2020, 28, 101754 CrossRef.
- L. Ou, C. Dou, J. H. Yu, H. Kim, Y. C. Park, S. Park, S. Kelley and E. Y. Lee, Biofuels, Bioprod. Biorefin., 2021, 15, 404–415 CrossRef CAS.
- A. Giuliano, D. Barletta, I. De Bari and M. Poletto, Techno-economic Assessment of a Lignocellulosic Biorefinery Co-producing Ethanol and Xylitol or Furfural, Elsevier Masson SAS, 2018, vol. 43 Search PubMed.
- P. Unrean and N. Ketsub, Ind. Crops Prod., 2018, 123, 238–246 CrossRef CAS.
- P. Ranganathan, Biomass Convers. Biorefin., 2020, 1–4 Search PubMed.
- J. Becker, C. M. Rohles and C. Wittmann, Metab. Eng., 2018, 50, 122–141 CrossRef CAS PubMed.
- S. Jo, J. Yoon, S. M. Lee, Y. Um, S. O. Han and H. M. Woo, J. Biotechnol., 2017, 258, 69–78 CrossRef CAS PubMed.
- A. Radek, K. Krumbach, J. Gätgens, V. F. Wendisch, W. Wiechert, M. Bott, S. Noack and J. Marienhagen, J. Biotechnol., 2014, 192, 156–160 CrossRef CAS PubMed.
- T. Xia, E. Altman and M. A. Eiteman, Eng. Life Sci., 2015, 15, 65–72 CrossRef CAS.
- K. K. Cheng, G. Y. Wang, J. Zeng and J. A. Zhang, BioMed Res. Int., 2013, 1, 1–12 Search PubMed.
- J. B. McKinlay and C. Vieille, Metab. Eng., 2008, 10, 55–68 CrossRef CAS PubMed.
- J. B. McKinlay, Y. Shachar-Hill, J. G. Zeikus and C. Vieille, Metab. Eng., 2007, 9, 177–192 CrossRef CAS PubMed.
- W. Sabra, R. R. Bommareddy, G. Maheshwari, S. Papanikolaou and A. P. Zeng, Microb. Cell Fact., 2017, 16, 1–14 CrossRef PubMed.
- G. L. Peabody, J. Winkler and K. C. Kao, Curr. Opin. Chem. Eng., 2014, 6, 9–17 CrossRef.
- A. Radek, N. Tenhaef, M. F. Müller, C. Brüsseler, W. Wiechert, J. Marienhagen, T. Polen and S. Noack, Bioresour. Technol., 2017, 245, 1377–1385 CrossRef CAS PubMed.
- R. E. Hector, N. Qureshi, S. R. Hughes and M. A. Cotta, Appl. Microbiol. Biotechnol., 2008, 80, 675–684 CrossRef CAS PubMed.
- J. G. R. Bueno, G. Borelli, T. L. R. Corrêa, M. B. Fiamenghi, J. José, M. De Carvalho, L. C. De Oliveira, G. A. G. Pereira and L. V. Dos Santos, Biotechnol. Biofuels, 2020, 13, 1–20 CrossRef PubMed.
- S. R. Kim, S. J. Ha, N. Wei, E. J. Oh and Y. S. Jin, Trends Biotechnol., 2012, 30, 274–282 CrossRef CAS PubMed.
- J. H. Van Vleet and T. W. Jeffries, Curr. Opin. Biotechnol., 2009, 20, 300–306 CrossRef CAS PubMed.
- J. C. Roseiro, M. A. Peito, F. M. Gírio and M. T. Amaral-Collaço, Arch. Microbiol., 1991, 156, 484–490 CrossRef CAS.
- K. O. Osiro, D. P. Brink, C. Borgström, L. Wasserstrom, M. Carlquist and M. F. Gorwa-Grauslund, FEMS Yeast Res., 2018, 18, 1–15 CrossRef PubMed.
- S. Cao, Y. Pu, M. Studer, C. Wyman and A. J. Ragauskas, RSC Adv., 2012, 2, 10925–10936 RSC.
- T. H. Kim, H. J. Ryu and K. K. Oh, Bioresour. Technol., 2016, 218, 367–372 CrossRef CAS PubMed.
- P. N. Ciesielski, W. Wang, X. Chen, T. B. Vinzant, M. P. Tucker, S. R. Decker, M. E. Himmel, D. K. Johnson and B. S. Donohoe, Biotechnol. Biofuels, 2014, 7(1), 1–12 CrossRef PubMed.
- S. M. de Vasconcelos, A. M. P. Santos, G. J. M. Rocha and A. M. Souto-Maior, Bioresour. Technol., 2013, 135, 46–52 CrossRef CAS PubMed.
- E. Liu, L. Das, B. Zhao, M. Crocker and J. Shi, BioEnergy Res., 2017, 10, 1079–1093 CrossRef CAS.
- X. Ji, H. Ma, Z. Tian, G. Lyu, G. Fang, J. Chen and H. A. M. Saeed, Bioresources, 2017, 12, 7084–7095 CAS.
- J. T. Cunha, A. Romaní, K. Inokuma, B. Johansson, T. Hasunuma, A. Kondo and L. Domingues, Biotechnol. Biofuels, 2020, 13, 1–15 Search PubMed.
- G. Batista, R. B. A. Souza, B. Pratto, M. S. R. dos Santos-Rocha and A. J. G. Cruz, Bioresour. Technol., 2019, 275, 321–327 CrossRef CAS PubMed.
- D. H. Fockink, J. H. Sánchez and L. P. Ramos, Ind. Crops Prod., 2018, 123, 563–572 CrossRef CAS.
- H. J. Zhang, X. G. Fan, X. L. Qiu, Q. X. Zhang, W. Y. Wang, S. X. Li, L. H. Deng, M. A. G. Koffas, D. S. Wei and Q. P. Yuan, Bioprocess Biosyst. Eng., 2014, 37, 2425–2436 CrossRef CAS PubMed.
- G. J. M. Rocha, A. R. Gonçalves, S. C. Nakanishi, V. M. Nascimento and V. F. N. Silva, Ind. Crops Prod., 2015, 74, 810–816 CrossRef CAS.
- R. Agrawal, A. Satlewal, R. Gaur, A. Mathur, R. Kumar, R. P. Gupta and D. K. Tuli, Biochem. Eng. J., 2015, 102, 54–61 CrossRef CAS.
- S. McIntosh, Z. Zhang, J. Palmer, H.-H. Wong, W. O. S. Doherty and T. Vancov, Biofuels, Bioprod. Biorefin., 2016, 10, 346–358 CrossRef CAS.
- M. Kapoor, S. Soam, R. Agrawal, R. P. Gupta, D. K. Tuli and R. Kumar, Bioresour. Technol., 2017, 224, 688–693 CrossRef CAS PubMed.
- D. J. Walker, J. Gallagher, A. Winters, A. Somani, S. R. Ravella and D. N. Bryant, Front. Energy Res., 2018, 6, 1–13 CrossRef.
- V. Kumar, P. P. Sandhu, V. Ahluwalia, B. B. Mishra and S. K. Yadav, Bioresour. Technol., 2019, 291, 121931 CrossRef CAS PubMed.
- S. S. Dalli, M. Patel and S. K. Rakshit, Biomass Bioenergy, 2017, 105, 402–410 CrossRef CAS.
- F. Bonfiglio, M. Cagno, C. K. Yamakawa and S. I. Mussatto, Ind. Crops Prod., 2021, 170, 113800 CrossRef CAS.
- K. S. Dhar, V. F. Wendisch and K. M. Nampoothiri, J. Biotechnol., 2016, 230, 63–71 CrossRef CAS PubMed.
- X. Yuan, Y. Mao, S. Tu, J. Lin, H. Shen, L. Yang and M. Wu, J. Agric. Food Chem., 2021, 69, 9625–9631 CrossRef CAS PubMed.
- R. Alves de Oliveira, R. Schneider, C. E. Vaz Rossell, R. Maciel Filho and J. Venus, Bioresour. Technol. Rep., 2019, 6, 26–31 CrossRef.
- Z. Qiu, Q. Gao and J. Bao, Bioresour. Technol., 2017, 245, 1369–1376 CrossRef CAS PubMed.
- Z. Zhang, Y. Li, J. Zhang, N. Peng, Y. Liang and S. Zhao, Microorganisms, 2020, 8, 1–9 Search PubMed.
- X. Kong, B. Zhang, Y. Hua, Y. Zhu, W. Li, D. Wang and J. Hong, Bioresour. Technol., 2019, 273, 220–230 CrossRef CAS PubMed.
- R. Glaser and J. Venus, Biotechnol. Rep., 2018, 18, e00245 CrossRef PubMed.
- K. L. Ong, C. Li, X. Li, Y. Zhang, J. Xu and C. S. K. Lin, Biochem. Eng. J., 2019, 148, 108–115 CrossRef CAS.
- H. Bao, R. Liu, L. Liang, Y. Jiang, M. Jiang, J. Ma, K. Chen, H. Jia, P. Wei and P. Ouyang, Enzyme Microb. Technol., 2014, 66, 10–15 CrossRef CAS PubMed.
- A. M. Olajuyin, M. Yang, T. Mu, J. Tian, A. Thygesen, O. A. Adesanoye, O. A. Adaramoye, A. Song and J. Xing, Bioprocess Biosyst. Eng., 2018, 41, 1497–1508 CrossRef CAS PubMed.
- D. Ladakis, K. Michailidi, A. Vlysidis, A. Koutinas and I. K. Kookos, Biochem. Eng. J., 2018, 137, 262–272 CrossRef CAS.
- Y. Mao, G. Li, Z. Chang, R. Tao, Z. Cui, Z. Wang, Y. J. Tang, T. Chen and X. Zhao, Biotechnol. Biofuels, 2018, 11, 1–17 CrossRef CAS PubMed.
- C. Wang, H. Zhang, H. Cai, Z. Zhou, Y. Chen, Y. Chen and P. Ouyang, Appl. Biochem. Biotechnol., 2014, 172, 340–350 CrossRef CAS PubMed.
- R. Liu, L. Liang, F. Li, M. Wu, K. Chen, J. Ma, M. Jiang, P. Wei and P. Ouyang, Bioresour. Technol., 2013, 149, 84–91 CrossRef CAS PubMed.
- A. Sharma, V. Nain, R. Tiwari, S. Singh and L. Nain, Chem. Eng. Commun., 2018, 205, 402–410 CrossRef CAS.
- P. R. D. Cortivo, J. Machado, L. R. Hickert, D. M. Rossi and M. A. Z. Ayub, Biotechnol. Prog., 2019, 35, 1–8 CrossRef PubMed.
- D. Yang, S. Y. Park, Y. S. Park, H. Eun and S. Y. Lee, Trends Biotechnol., 2020, 1–21 Search PubMed.
- C. C. Okonkwo, V. Ujor and T. C. Ezeji, Ind. Crops Prod., 2021, 159, 113047 CrossRef CAS.
- S. Rehman, M. Khairul Islam, N. Khalid Khanzada, A. Kyoungjin An, S. Chaiprapat and S. Y. Leu, Bioresour. Technol., 2021, 333, 125206 CrossRef CAS PubMed.
- J. Wu, Y. J. Zhou, W. Zhang, K. K. Cheng, H. J. Liu and J. A. Zhang, AMB Express, 2019, 9, 1–9 CrossRef PubMed.
- A. Kuenz, M. Jäger, H. Niemi, M. Kallioinen, M. Mänttäri and U. Prüße, Fermentation, 2020, 6, 86 CrossRef CAS.
- E. Tomás-Pejó, J. M. Oliva, M. Ballesteros and L. Olsson, Biotechnol. Bioeng., 2008, 100, 1122–1131 CrossRef PubMed.
- T. F. Pacheco, B. R. C. Machado, W. G. de Morais Júnior, J. R. M. Almeida and S. B. Gonçalves, Appl. Biochem. Biotechnol., 2021, 193, 2182–2197 CrossRef CAS PubMed.
- A. M. Zetty-Arenas, R. F. Alves, C. A. F. Portela, A. P. Mariano, T. O. Basso, L. P. Tovar, R. Maciel Filho and S. Freitas, Biomass Bioenergy, 2019, 126, 190–198 CrossRef CAS.
- W. Guan, G. Xu, J. Duan and S. Shi, Ind. Eng. Chem. Res., 2018, 57, 775–783 CrossRef CAS.
- Y. Jiang, Y. Lv, R. Wu, J. Lu, W. Dong, J. Zhou, W. Zhang, F. Xin and M. Jiang, Biotechnol. Bioeng., 2020, 117, 2985–2995 CrossRef CAS PubMed.
- R. Allard-Massicotte, H. Chadjaa and M. Marinova, Fermentation, 2017, 3, 31–41 CrossRef.
- D. Atzmüller, N. Ullmann and A. Zwirzitz, AMB Express, 2020, 10, 1–11 CrossRef PubMed.
- N. H. Abd Rahman, J. Md. Jahim, M. S. Abdul Munaim, R. A. Rahman, S. F. Z. Fuzi and R. Md. Illias, Enzyme Microb. Technol., 2020, 135, 109495 CrossRef CAS PubMed.
- Y. Hua, J. Wang, Y. Zhu, B. Zhang, X. Kong, W. Li, D. Wang and J. Hong, Microb. Cell Fact., 2019, 18, 1–18 CrossRef PubMed.
- S. M. S. Reshamwala and A. M. Lali, Biotechnol. Prog., 2020, 36, e2972 CrossRef CAS PubMed.
- T. L. Turner, G. C. Zhang, S. R. Kim, V. Subramaniam, D. Steffen, C. D. Skory, J. Y. Jang, B. J. Yu and Y. S. Jin, Appl. Microbiol. Biotechnol., 2015, 99, 8023–8033 CrossRef CAS PubMed.
- M. Ilmén, K. Koivuranta, L. Ruohonen, P. Suominen and M. Penttilä, Appl. Environ. Microbiol., 2007, 73, 117–123 CrossRef PubMed.
- S. Shinkawa, K. Okano, S. Yoshida, T. Tanaka, C. Ogino, H. Fukuda and A. Kondo, Appl. Microbiol. Biotechnol., 2011, 91, 1537–1544 CrossRef CAS PubMed.
- K. T. Koivuranta, M. Ilmén, M. G. Wiebe, L. Ruohonen, P. Suominen and M. Penttilä, Microb. Cell Fact., 2014, 13, 1–14 CrossRef PubMed.
- S. Jo, J. Yoon, S. M. Lee, Y. Um, S. O. Han and H. M. Woo, J. Biotechnol., 2017, 258, 69–78 CrossRef CAS PubMed.
- R. Liu, L. Liang, K. Chen, J. Ma, M. Jiang, P. Wei and P. Ouyang, Appl. Microbiol. Biotechnol., 2012, 94, 959–968 CrossRef CAS PubMed.
- L. Yang, M. M. Henriksen, R. S. Hansen, M. Lübeck, J. Vang, J. E. Andersen, S. Bille and P. S. Lübeck, Biotechnol. Biofuels, 2020, 13, 1–12 CrossRef PubMed.
- L. Li, K. Li, Y. Wang, C. Chen, Y. Xu, L. Zhang, B. Han, C. Gao, F. Tao, C. Ma and P. Xu, Metab. Eng., 2015, 28, 19–27 CrossRef CAS PubMed.
- S. Yang, A. Mohagheghi, M. A. Franden, Y. C. Chou, X. Chen, N. Dowe, M. E. Himmel and M. Zhang, Biotechnol. Biofuels, 2016, 1–15 Search PubMed.
- S. J. Kim, J. W. Kim, Y. G. Lee, Y. C. Park and J. H. Seo, Appl. Microbiol. Biotechnol., 2017, 101, 2241–2250 CrossRef CAS PubMed.
- A. S. Abdelaal, K. Jawed and S. S. Yazdani, J. Ind. Microbiol. Biotechnol., 2019, 46, 965–975 CrossRef CAS PubMed.
- L. Yu, J. Zhao, M. Xu, J. Dong, S. Varghese and M. Yu, Appl. Microbiol. Biotechnol., 2015, 99, 4917–4930 CrossRef CAS PubMed.
- G. Huo, Y. Zhu, Q. Liu, R. Tao, N. Diao and T. Chen, J. Chem. Technol. Biotechnol., 2017, 92, 2739–2745 CrossRef CAS.
- A. G. Sandström, A. Muñoz, D. Heras, D. Portugal-nunes and M. F. Gorwa-grauslund, AMB Express, 2015, 5, 1–14 CrossRef PubMed.
- A. Muñoz, D. Heras, N. Rizza, A. G. Sandström and M. F. G. Grauslund, Microb. Cell Fact., 2016, 7, 1–12 Search PubMed.
| This journal is © The Royal Society of Chemistry 2022 |