
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
In vitro gastrointestinal bioaccessibility and colonic fermentation of lignans from fresh, fermented, and germinated flaxseed†
Umi Kalsum
Hussain Zaki
 ac,
Christos
Fryganas
b,
Laura
Trijsburg
a,
Edith J. M.
Feskens
a and
Edoardo
Capuano
*b
ac,
Christos
Fryganas
b,
Laura
Trijsburg
a,
Edith J. M.
Feskens
a and
Edoardo
Capuano
*b
aDivision of Human Nutrition and Health, Wageningen University & Research, Wageningen, The Netherlands
bFood Quality & Design Department, Wageningen University & Research, Wageningen, The Netherlands. E-mail: edoardo.capuano@wur.nl; Tel: +31-317485690
cFood Science and Technology Research Centre, Malaysian Agricultural Research and Development Institute (MARDI), Selangor, Malaysia
First published on 30th September 2022
Abstract
This research assessed the influence of fermentation and germination as well as of particle size on lignan bioaccessibility from flaxseed by simulated in vitro gastrointestinal digestion. In vitro simulated colonic fermentation was used to study lignan release and its conversion into enterolignans. In addition, tea was included as a representative sample to investigate the stability of lignans in the gastrointestinal tract. Only secoisolariciresinol (SECO) was detected in flaxseed samples. SECO bioaccessibility in fermented flaxseed was highest among all matrices but limited to ≈1% (P < 0.001). Lignan bioaccessibility was significantly influenced by particle size too (P < 0.001 for both). In the colon, fermented flaxseed produced the highest SECO release among all flaxseed samples (≈65%), and the highest conversion of enterolignan (≈1.0%), whereas the conversion of lignans in tea brew was relatively high (≈15%). Lignan conversion varies greatly among donors due to inter-individual differences in microbiota activity. Food fermentation could be a viable strategy for increasing lignan release and conversion to enterolignan.
Introduction
Lignans are phenolic compounds that belong to the class of non-flavonoid polyphenols that can exert both oestrogenic and anti-oestrogenic effects.1 Some lignans can be metabolized into enterolignans, i.e. enterodiol and enterolactone, by the human intestinal microbiota2 through deglycosylation, oxidation, and demethylation.3 These metabolites have been reported to have beneficial health effects. Epidemiologic studies have shown that the intake of lignan-rich foods aids in lowering the risk of several cancers4 and cardiovascular diseases,5 demonstrating their antioxidant properties as well as estrogenic activity.6 Differences in microbiota composition may lead to differences in the absorption and excretion patterns of enterolignans.2It is widely known that not all of the constituents in a food matrix are fully bioavailable. The fraction of a nutrient or non-nutrient available to the human body for physiological functions and/or storage is known as bioavailability.7 As a result, investigating the bioavailability of a food component is essential to predict its biological activity.8
Food processing affects the lignan content of foods9 and the levels and bioavailability of bioactive compounds.10 Reducing the particle size of plant tissue, for instance, increases the fraction of broken cells thus increasing the fraction of potentially bioavailable intracellular compounds.11 However, most lignans, similar to phenolic compounds in general, are covalently or physically bound to plant cell walls and thus have low bioavailbility.12 As a result, enzymatic treatments and fermentation processes targeting bran structure have been investigated to improve the bioavailability of lignans.13 In addition, germination and fermentation are also reported to affect lignan content,14,15 but how these processing methods affect the bioavailability of lignans is unknown.
Since animal and human studies are time-consuming and expensive and may raise ethical concerns, invitro approaches that predict the behaviour of phytochemicals during GI digestion have been developed. This approach enables the screening of a relatively large number of samples and/or conditions to study the separate and combined effects of each phase of digestion on the release and availability of phytochemicals, which would be impossible with in vivo approaches.8 Bioaccessibility is the fraction of a potentially available compound released from its matrix in the gastrointestinal (GI) tract and thus becomes available for intestinal absoption.16 Furthermore, it has been demonstrated that bioaccessibility as determined using in vitro models can be well associated with bioavailability determined in human studies and animal models.17
To date, there are relatively few reports on the effect of various processing techniques on the bioaccessibility of lignans from food. Thus, the present study investigated lignan bioaccessibility from fresh, fermented, and germinated flaxseed, and the effect of particle size using simulated in vitro digestion models. In addition, tea was used to study the stability of lignans under in vitro simulated gastrointestinal conditions. Moreover, the release of lignans and their conversion into enterolignans was investigated through in vitro colonic fermentation with human fecal inoculums.
Materials and methods
Chemicals and reagents
Porcine pepsin (P6887), porcine pancreatin (P1750, 4× USP), and porcine bile salt preparation (B8631) were purchased from Sigma-Aldrich (Merck KGaA, Germany). KCl, KH2PO4, NaCl, MgCl2·(H2O)6, CaCl2·(H2O)2, and pure ethanol were purchased from VWR International B.V. (Netherlands). KH2PO4, NaCl, (NH4)2CO3, NaHCO3, NaOH, HCl, and Tween 80 were purchased from Sigma-Aldrich Chemie B.V. (Netherlands), as well as yeast extract, peptone, mucine, and L-cysteine HCl.Acetonitrile, methanol, sodium acetate, sodium hydroxide, glacial acetic acid, formic acid, pure standards of pinoresinol (PINO), matairesinol (MATA), secoisolariciresinol (SECO), enterolactone (EL), enterodiol (END) and glycocholic acid (glycine-1 13C, 13C-GA) were purchased from Sigma-Aldrich Chemie NV (Zwijndrecht, Netherlands). For enzymatic hydrolysis, β-glucuronidase with an activity of ≥300![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 units per g of solid and a sulfatase activity of ≥10000 units per g of solid type H-1 from Helix pomatia were purchased from Sigma-Aldrich Chemie GmbH (Schnelldorf, Germany). The Rhizopus oryzae fungus for the fermentation of flaxseed, mung beans, and soybeans was obtained from Startercultures.eu/starterculturen.nl. All of the chemicals used were of analytical and chromatographic grade.
000 units per g of solid and a sulfatase activity of ≥10000 units per g of solid type H-1 from Helix pomatia were purchased from Sigma-Aldrich Chemie GmbH (Schnelldorf, Germany). The Rhizopus oryzae fungus for the fermentation of flaxseed, mung beans, and soybeans was obtained from Startercultures.eu/starterculturen.nl. All of the chemicals used were of analytical and chromatographic grade.
Preparation of samples
:1 to form a white layer of compact mycelium around the beans and bind them into a dense, firm cake. Mung beans were rinsed and soaked in water for 12 h, then were dehulled, split in half, and boiled in demineralized water (1:2, w/v) for 30 minutes. Subsequently, the flaxseed was rinsed, drained, and boiled for 3 minutes. Then, flaxseed and mung beans were mixed (1:1, w/w) to surround the beans with a white layer of compact mycelium. The sample was allowed to cool to below 36 °C and transferred to a container with vinegar (20 mL). Next, the samples were inoculated with the fungi Rhizopus oryzea at the level of 0.1% (w/w) of boiled samples. Finally, the samples were placed in perforated polyethylene bags and incubated for 24–48 hours at 25–30 °C, or until a compact white mycelium formed.
Simulated in vitro gastrointestinal digestion
All samples were digested using a static in vitro digestion system consisting of a simulated oral, gastric, and intestinal phase, with modifications.20,21 The compositions (%, w/w) of the simulated salivary fluid (SSF), simulated gastric fluid (SGF, pH 3.0 ± 0.05), and simulated intestinal fluid (SIF, pH 7.0 ± 0.05) were as reported.20For the oral phase, flaxseed samples (5 g) or tea brew (5 mL) were mixed with the SSF electrolyte stock solution (3.5 mL, 37 °C), CaCl2 (0.025 mL, 0.3 M), and Milli-Q water (1.475 mL) (Veolia water, Veolia Water Solutions, and Technologies Netherlands B.V.). The final volume was 10 mL.
Subsequently, an oral bolus (10 mL) was mixed with SGF (7.5 mL, 37 °C) to start the gastric phase. The pH was adjusted to 3 with HCl (0.2 mL, 1 M). Then, CaCl2 (0.005 mL, 0.3 M) was added. Finally, Milli-Q water was added to obtain the final 20 mL (approximately 0.698 mL). Porcine pepsin stock solutions (1.6 mL, 25000 units per mL, 37 °C) (2000 units per mL in final chyme) was added the SGF. Then, the chyme was shaken for 2 h (37 °C). At the end of the gastric digestion step, NaOH (0.15 mL, 1 M) was added to adjust the SGF to pH 7.0, inhibiting the gastric enzyme activity.
Thereafter, NaOH (0.15 mL, 1 M) was added to 20 mL of gastric chyme to reach pH 7.0. Subsequently, SIF (11 mL, 37 °C), fresh bile stock solution (2.5 mL, to reach a final concentration of 10 mM in the intestinal chyme), and CaCl2 (0.04 mL, 0.3 M) were added. Then, Milli-Q water (1.31 mL) was added. Finally, pancreatin solution (5 mL, 1600 U mL−1 amylase activity) was added to reach a final volume of 40 mL. At the end of the intestinal step, samples were incubated for 2 hours (37 °C) on a rotating device.
In parallel, control samples with no digestive enzymes added were tested to see if these enzymes affect lignan bioaccessibility. Milli-Q water was added to replace the enzyme solutions. All gastric and intestinal digests were centrifuged (4 °C, 20000g, 10 minutes) to stop the enzymatic reaction. Supernatants (5 mL) were collected and filtered through a 0.22 μm membrane filter (Phenomenex, Netherlands) for lignan analysis. Then, the remaining pre-digested samples were freeze-dried for further use in in vitro colonic fermentation.
The bioaccessibility of lignans was defined as the content of lignans that was detected in the supernatants after digestion, divided by the total lignans initially present in the samples before digestion:

In vitro colonic fermentation
Fresh fecal samples were donated by three healthy adults (one Malay and two Dutch, aged 25–40 years old, with a body mass index (BMI) of 18.5–23.9), who declared no smoking and no antibiotic consumption for 6 months before the beginning of the study. The fecal samples were prepared according to protocols previously reported, with modifications.22,23 Briefly, feces (20.0 g) were dissolved in anaerobic phosphate buffer (100 mL) before homogenization in a stomacher bag. The phosphate buffer contained 8.8 g L−1 K2HPO4, 6.8 g L−1 of KH2PO4, and 0.1 g L−1 of sodium thioglucolate. The resulting fecal suspension was filtered and considered to be the fecal microbiota supernatant. The colon medium consisted of 5.22 g L−1 K2HPO4, 16.32 g L−1 KH2PO4, 2.0 g L−1 NaHCO3, 2.0 g L−1 yeast extract, 2.0 g L−1 peptone, 1.0 g L−1 mucine, 0.5 g L−1L-cysteine HCl, and 2.0 mL L−1 Tween 80. Both phosphate buffer and colon medium were flushed with nitrogen (30 minutes) before autoclaving.Fecal supernatant (7 mL) was added to a penicillin bottle containing a buffered colon medium (43 mL). Then, pre-digested fresh, fermented, or germinated flaxseed (1 g) or tea sample (10 mL) was added, and sterile water was added to make up the mixture of fecal supernatant and the colon medium to 70 mL. The fermentation started with continuous shaking (60 rpm, 37 °C) under anaerobic conditions. In parallel, one control without the fecal supernatant was included to determine the release of lignans from the food matrix. All incubations were duplicated, and supernatants were collected at time intervals (2, 8, and 24 h). All supernatants were centrifuged immediately when sampled and stored at −20 °C until further use. For the in vitro colonic fermentation, only the pre-digested samples with a fine particle size were used. Two replicates were performed using two independent samples.
The release of SECO was defined as the content of SECO detected in the supernatants after fermentation, divided by the total SECO present in the pre-digested samples after in vitro gastrointestinal digestion. Then, the conversion yield was defined as the content of enterolignans that was detected in the supernatants after colonic fermentation, divided by the total lignans initially present in the pre-digested samples after in vitro gastrointestinal digestion:

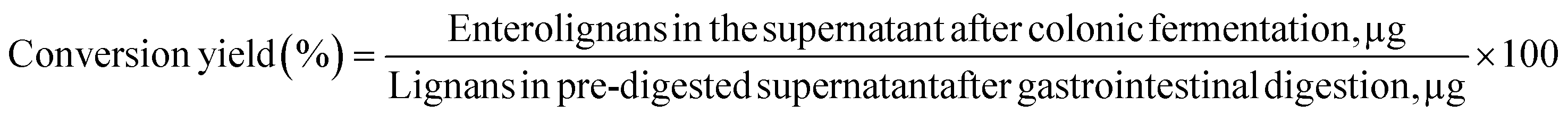
Lignan extraction
Lignans were extracted by alkaline hydrolysis as suggested by Peñalvo et al., Milder et al., and Nørskov et al. with slight modifications.24–26 In brief, 20 mg of coarse and fine fresh, fermented, and germinated flaxseed were extracted under alkaline conditions (1 mL, 0.3 M NaOH in methanol/water, 8/2, v/v) at 60 °C for 1 h. Samples were centrifuged, and the supernatants were transferred to plastic tubes and evaporated under N2 flow at 60 °C. The extracts from pre-digestion and in vitro fermentation treatments were subjected to hydrolysis by adding to them 0.6 mL of β-glucuronidase/sulfatase (2 mg mL−1 in 50 mM sodium acetate buffer, pH 5) and also to brew tea (1 mL, without alkaline extraction) using a vortex incubator (37 °C, 16 h). The hydrolysis was stopped by adding formic acid (0.5 mL, 0.4%), and the extracts were centrifuged. Subsequently, 300 μL of acetonitrile was added, and the samples were transferred to a HPLC vial for LC-MS/MS analysis.LC-MS/MS analysis of lignans
A Nexera UPLC system (Shimadzu Corporation, Kyoto, Japan) was used, coupled with a LCMS-8050 triple quadrupole mass spectrometer (Shimadzu Corporation, Kyoto, Japan). 5 μL of sample was injected into an Acquity Premier BEH C18 column, 1.7 μm, 2.1 × 100 mm BEH (Waters Chromatography B.V., 4879 AH Etten-Leur, the Netherlands). The flow rate was set at 0.3 mL min−1 and the column temperature at 40 °C. The mobile phases consisted of 0.1% formic acid (solvent A) and acetonitrile with 0.1% formic acid (solvent B) with the following elution profile (t in [min]/[%B]): (0.0/15), (9.0/75), (11.0/75), (13.0/15), (18.0/15). MS data were collected for 18 minutes.All analytes were analyzed in positive ionisation mode. The turbo ion-spray ionization voltage was 4.0 kV. The electrospray ionization probe, desolvation line, and heat block were all set to 300, 250, and 400 degrees Celsius, respectively. The collision-induced dissociation gas had a pressure of 4 kPa, and the flow rates of the drying gas, nebulizer gas, and heating gas were set at 10 mL min−1, 3 mL min−1, and 10 mL min−1, respectively. The electrode voltage of Q1 pre-bias (collision cell energy entrance potential), collision cell Q2 (collision energy), Q3 pre-bias (collision cell energy exit potential), and parent and fragment ion m/z of the multiple reaction monitoring transitions were optimized using support software (Shimadzu Corporation, Kyoto, Japan). The dwell time ranged from 11 ms to 16 ms for the analytes for single reaction monitoring (SRM). The most abundant fragment ions for SECO, 13C-GA, PINO, MATA, EL, and ENT were 363.2 → 137.05 (collision energy: −24 V), 467.20 → 413.3 (collision energy: −18 V), 359.2 → 341.15 (collision energy: −9 V), 359.2 → 137.10 (collision energy: −27 V), 299.2 → 133.0 (collision energy: −16 V), and 303.0 → 107.0 (collision energy: −25 V), respectively, and therefore were selected for quantitation. The representative chromatograms of fresh flaxseed, the intestinal phase of simulated in vitro digestion, and 2 h and 24 h of in vitro simulated colonic fermentation of fresh flaxseed are provided in Fig. S1.†
Method validation
:1, v/v) was prepared on the day of the analysis and used for the preparation of the calibration curves (12.5–2500 ng mL−1).
Statistical analysis
Data of bioaccessibility are presented as mean ± standard deviation (SD) from duplicate analyses of two independent digestions. Data for release of SECO during in vitro colonic fermentation, and conversion into enterolignans, are averages from 3 donors. Each in vitro fermentation was performed in duplicate. One-way analysis of variance (ANOVA) was used to assess the significant differences in the bioaccessibility of fresh, fermented, and germinated flaxseed and tea brew among different digestion phases. An additional ANOVA was used to test the differences in bioaccessibility values obtained with or without the inclusion of digestive enzymes in the simulated in vitro digestion. Two-way ANOVA followed by Tukey's HSD multiple comparison tests were used to test the effect of the type of processing (fresh, fermented and germinated flaxseed), particle size (fine and coarse) and their interaction on lignan bioaccessibility. Pearson's correlation was used to identify the correlation between the amount of SECO released and its conversion to EL by correlating data for each donor at all fermentation times. Then, a repeated ANOVA measurement followed by Tukey's HSD multiple comparison post-hoc tests were applied to test the difference in SECO release and conversion yield into enterolignans during in vitro colonic fermentation of fresh, germinated and fermented flaxseed and tea brew among the 3 donors. An additional repeated ANOVA measurement followed by Tukey's HSD multiple comparison post-hoc tests were used to test the differences in SECO release and conversion yield into enterolignans of fresh, germinated, and fermented flaxseed and tea brew within three time points. P-values <0.05 were regarded as statistically significant. Statistical analyses were performed using the IBM SPSS Statistics version 25 software (IBM Corp).Results and discussion
Effect of processing and particle size on lignan bioaccessibility during simulated in vitro gastro-intestinal digestion of flaxseed matrixes
In the present research, the effects of fermentation and germination as well as particle size reduction on lignan bioaccessibility were assessed using simulated in vitro gastrointestinal digestion. In addition to the flaxseed matrixes, tea brew was included as a representative sample to study the stability of lignans in the gastrointestinal environment and their interactions with digestive fluids. SECO was detected in fresh, germinated, and fermented flaxseed. Only PINO and MATA were found in tea brew (Table 1). The SECO content was the highest in germinated flaxseed, with values of 15.6 ± 1.1 mg per g dry weight (DW) and 14.6 ± 0.6 mg per g DW in fine and coarse particles, respectively. Fermented flaxseed showed the second highest content of SECO with values of 11.1 ± 2.1 mg per g DW and 9.8 ± 0.6 mg per g DW in fine and coarse particles, respectively. Fresh flaxseed has the lowest SECO content with values of 3.0 ± 0.5 mg per g DW and 2.1 ± 0.4 mg per g DW in fine and coarse particles, respectively.| Fine* | Coarse | ||||
|---|---|---|---|---|---|
| Oral | Gastric | Intestinal | Oral | Gastric | Intestinal |
| Flaxseed (SECO)a | |||||
| 0.67 ± 0.07A | 0.72 ± 0.05A | 0.87 ± 0.09A | 0.61 ± 0.12A | 0.69 ± 0.08A | 0.67 ± 0.02A |
| Germinated flaxseed (SECO)b | |||||
| 0.47 ± 0.10A | 0.52 ± 0.05A | 0.55 ± 0.04A | 0.37 ± 0.05A | 0.50 ± 0.04A | 0.53 ± 0.04A |
| Fermented flaxseed (SECO)ac | |||||
| 0.70 ± 0.06A | 0.72 ± 0.01A | 1.03 ± 0.04B | 0.63 ± 0.06A | 0.69 ± 0.08A | 1.00 ± 0.22B |
| Oral | Gastric | Intestinal |
|---|---|---|
| Different lower case letters indicate within samples (fresh, germinated, and fermented flaxseed); upper case letters indicate within the digestion phase. * Within this particle size a significant difference in bioaccessibility (P < 0.05) is indicated. SECO = secoisolariciresinol; PINO = pinoresinol; MATA = matairesinol. | ||
| Tea brew (PINO and MATA) | ||
| 88.5 ± 8.42A | 67.0 ± 3.51B | 24.4 ± 4.15C |
As a general observation, bioaccessibility of lignans from the flaxseed food matrixes was very low (1% and less). Our findings are in accordance with a study on flaxseed that showed no lignans were released during in vitro simulated gastrointestinal digestion.27
Both the type of processing and particle size significantly affected the bioaccessibility of SECO (P < 0.001 for both). No interaction between particle size and samples was observed (P = 0.142). The bioaccessibility was highest in fermented flaxseed, reaching around 1.03%, while it was 0.87% and 0.67% in fine and coarse particles of fresh flaxseed, respectively. Germinated flaxseed presented the lowest bioaccessibility of SECO (0.5%) for both particle sizes.
Fresh flaxseed showed a significant difference in SECO bioaccessibility between the digestion phases in fine and coarse particles, P = 0.047 and P = 0.002, respectively. Similarly, fermented flaxseed showed a significant difference in SECO bioaccessibility among digestion phases in fine and coarse particles, P = 0.019 and P = 0.024, respectively. However, in germinated flaxseed, coarse size showed a significant difference in SECO bioaccessibility between the digestion phases (P = 0.017) but not for fine size (P = 0.066).
In tea brew, the lignan content was 330.5 ± 29.7 ng mL−1. In tea brew, the bioaccessibility of lignans was about 88% after the oral phase and decreased to approximately 70% after the gastric phase (P = 0.04), and further decreased to around 25% after the intestinal phase.
Furthermore, the addition of digestion enzymes did not significantly affect the bioaccessibility of SECO in flaxseed samples for fine (P = 0.640) and coarse sizes (P = 0.261). Similar to flaxseed samples, digestion enzymes did not significantly affect the bioaccessibility of MATA and PINO in tea brew (P = 0.062).
Our finding is in line with previous research that showed that milling and crushing of flaxseed, i.e. decreasing the particle size, will enhance bioavailability.28 To the best of our knowledge, no data are available on the bioaccessibility of lignans during fermentation using Rhizopus oyrzae. Fungi of the genus Rhizopus have been traditionally used in food fermentation, specifically of tempeh in South-East Asia including Malaysia, for centuries. Oryzae is considered to be GRAS by the FDA but not by the EFSA, which mostly raises an issue related to mycotoxin presence.29 The chance that the fungus may survive the passage through the gastrointestinal tract, as well as colonize and thrive in the ecology of the large intestine is negligible. Nevertheless, a study on fermented raw pumpkin oil cake proved an effective release of bound phenolics did occur, which may indicate an increase in bioaccessibility.30 Likewise, a study on fermented rice bran with Rhizopus oryzae reported an increased phenolic content compared to the unfermented samples31 by releasing insoluble bound phenolic compounds via the activity of exogenous enzymes.32
To the best of our knowledge, there are no published data on the bioaccessibility of lignans in germinated food. However, one study has shown that the content and bioavailability of phenolic compounds increased in germinated compared to non-germinated grains.33 This would be due to the activity of endogenous enzymes releasing bound phenolic compounds, which increased bioaccessibility.34
The particle size of foods affects the bioaccessibility of phytochemicals. A study showed that the bioaccessibility of the antioxidant and total phenolic content of carob flour improved with a reduction of the particle size.35 Similarly, another study proved that the reduction in particle size in wheat bran increased ferulic acid bioaccessibility.36 Grinding breaks down plant cells, increasing the yield and bioaccessibility.37
All in all, our findings indicate that the SECO bioaccessibility from the food matrix may be altered according to the way foods are processed.
The results show here for the first time that lignans are unstable under simulated gastrointestinal conditions by using tea brew. Other studies have reported that polyphenols may be unstable under simulated in vitro gastrointestinal digestion, with only 20% remaining intact at the end of the intestinal phase.38 Degradation, epimerization, hydrolysis, and oxidation of polyphenols are significant transformations during digestion, particularly after the transition from the gastric to the intestinal environment.39 Furthermore, a study speculated that the presence of bile salt in the intestinal phase would reduce campesterol, β-sitosterol, and sitosterol in beverages due to reduced solubility of sterols,40 which may have impacted the bioaccessibility of lignans as well. However, the observation that lignans could be degraded during gastric and intestinal digestion is relevant when designing strategies to optimize the health effect of lignans.
We also calculated the net amount of parent lignans that would be available to our body when the simultaneous effect of fermentation and germination on content and bioaccessibility is considered. The amount of SECO available for absorption was highest in fine and coarse fermented flaxseed, with values of 113 μg per g DW and 98 μg per g DW, respectively. Germinated flaxseed showed the second highest SECO content in the intestinal phase with values of 85.8 μg per g DW and 77.4 μg per g DW in fine and coarse sizes, respectively. Then, fresh flaxseed has the lowest SECO content in the intestinal phase, with values of 26.1 μg per g DW and 14.1 μg per g DW in fine and coarse sizes, respectively.
From the findings mentioned above, it was concluded that fermentation and germination could increase the amount of circulating lignans. During fermentation, the endogenous and bacterial enzymes could modify grain constituents, thus affecting the absorption and bioavailability of bioactive compounds.41 Meanwhile, during germination, the soaking process activated the enzymes to break down the active compounds.42 Fermentation and germination are relatively simple technologies yet they are widely used to increase the nutrient density in bioactive compounds by releasing the bound fraction in the food matrix, thereby increasing the bioaccessibility.43
Lignans released from the food matrix during in vitro colonic fermentation
Next, the release of lignans in the in vitro colonic fermentation phase was further investigated. Only pre-digested fine particles were used since this particle size showed a higher bioaccessibility than the coarse size. The release of SECO from fine particles of fresh, germinated, and fermented flaxseed during in vitro colonic fermentation is shown in Fig. 1. A control was added, where the flaxseed samples were dissolved in the fermentation medium without fecal inoculum. | ||
| Fig. 1 Release of SECO (% of the pre-digested samples) in the supernatant during in vitro colon fermentation of fine particles of fresh, germinated, and fermented flaxseed. Data are from 3 donors (mean ± SD, n = 3). Different lower case letters within samples indicate a significant difference in the % SECO released during three time points (2, 8, 24 h) in the supernatant (P < 0.05). SECO = secoisolariciresinol. | ||
Notably, the SECO can be already detected in the supernatant after 2 hours of fermentation. A significant increase was found in SECO released in fermented flaxseed from 2 to 24 hours of fermentation. The SECO released was significantly increased from 8 to 24 h in flaxseed and germinated flaxseed. After 24 hours of in vitro fermentation, fermented flaxseed produced the highest SECO release among all samples, approximately 65%, showing that the metabolic activity of the fungus Rhizopus oryzae may help the release of lignans in the colon. The SECO released was comparable between fresh and germinated flaxseed (approximately 10%). However, a significantly lower percentage of SECO was released in the control (approximately 2.5%). This clearly demonstrates that microbiota activity in the large intestine aids in releasing SECO from the plant matrix.
Lignan conversion to enterolignan during in vitro colonic fermentation
In the present study, the conversion of lignans to enterolignans was also investigated. Only enterolactone (EL) was detected in the samples after colonic fermentation, probably because of the rapid dehydrogenation of END into EL.44 SECO conversion to EL during in vitro colon fermentation of fresh, germinated, and fermented flaxseed is shown in Fig. 2. The conversion of EL was significantly higher in fermented flaxseed at all times, ranging from 0.5% to 1.0%. In vitro colonic fermentation of fresh and germinated flaxseed produced a lower conversion to EL of approximately 0.1% after 24 hours of fermentation, with no significant difference between the samples.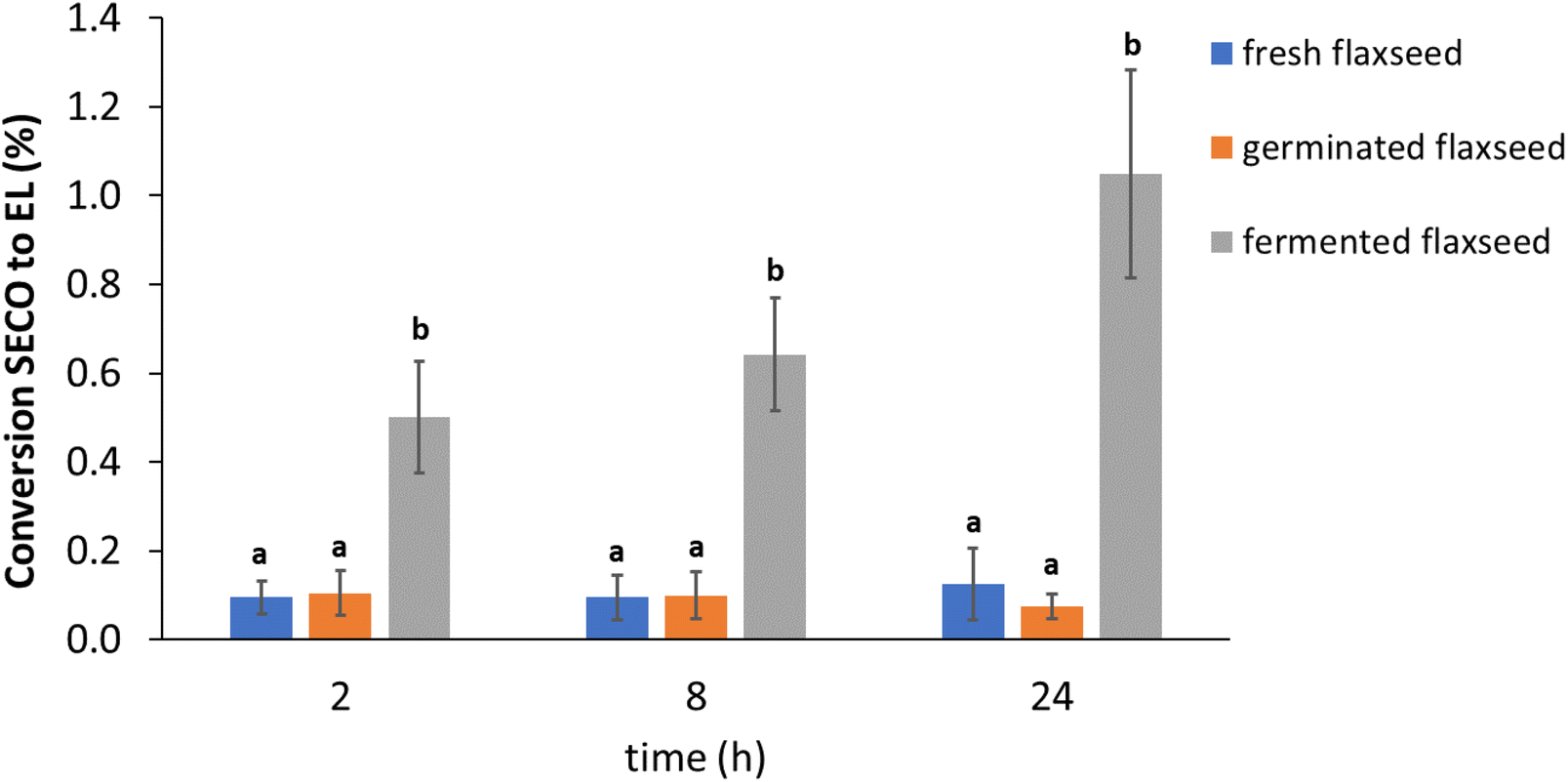 | ||
| Fig. 2 Lignan (SECO) conversion to EL during in vitro colon fermentation of fresh, germinated, and fermented flaxseed. Data are from 3 donors (mean ± SD, n = 3). Different lower case letters within samples indicate a significant difference in the SECO conversion during three time points (2, 8, 24 h) in the supernatant (P < 0.05). SECO = secoisolariciresinol; EL = enterolactone. | ||
The fermentation time did not significantly affect the conversion of SECO in EL in fresh (P = 0.881) and germinated flaxseed (P = 0.801), while the production of EL was significantly increased from 8 to 24 hours of fermentation in the fermented flaxseed (P = 0.024). As expected, no EL was detected in the control samples because microbiota are necessary for the conversion of SECO to EL.
Furthermore, the correlation between SECO release and conversion to EL in fresh, germinated and fermented flaxseed at 24 h of fermentation was evaluated. SECO release in all flaxseed samples was significantly positively correlated with EL conversion at 24 h of fermentation (r = 0.999, P < 0.029). It was concluded that the more lignans released, the higher the conversion to EL because more substrate is available.
The net amount of EL available to our body when the simultaneous effect of fermentation and germination on content and conversion of SECO is considered was calculated. The EL content produced after colonic fermentation was by far at its highest in fermented flaxseed, with a value of 116.6 μg per g DW, followed by germinated flaxseed with a value of 10.9 μg per g DW. The EL production in fresh flaxseed had its lowest value of 4.0 μg per g DW. Fermented flaxseed had an approximately 29-fold difference compared to fresh flaxseed. In addition, the germination of flaxseed would substantially increase EL production with a nearly 3-fold difference compared to fresh flaxseed. According to the aforementioned results, it was concluded that food fermentation and germination could significantly enhance EL production from flaxseed.
The inter-individual differences were looked into further and reported data for the single donors are shown in Fig. 3. Donor 3 had the highest EL conversion rate of all the samples, around 1.5% in fermented flaxseed. Nearly a 5-fold difference in EL production was observed in fresh and fermented flaxseed between the highest and lowest producers.
 | ||
| Fig. 3 Lignan (SECO) conversion to EL at 24 hours in in vitro colon fermentation of fresh, germinated, and fermented flaxseed. Data are from two replications (mean ± SD, n = 2). Different lower case letters indicate a significant difference in SECO conversion to EL among the three donors in each sample. SECO = secoisolariciresinol; EL = enterolactone. | ||
The gut microbiota pathway for the formation of enterolignans begins with demethylation and dehydroxylation of SECO, followed by oxidation of ED to EL.45 Factors such as diet, genetics,46 gender,28 and age47 affect the conversion of lignans to EL by modulating microbiota.48 In the present research, the variability observed in lignan conversion among donors is great, clearly due to differences in the individual microbiota. It was suggested that microbiota dehydrogenation of lignans to generate EL is a crucial step in the colon that could explain major variations in EL production.49 The ability of gut microbiota to metabolize lignans to EL is determined by the presence of a metabolite production gradient that gives rise to “high producers” and “low producers” of metabolites.50 To date, only a few microbiota species have been identified as enabling the production of EL. In particular, EL production has been related to the abundance of Ruminococcus,51Bacteroides, and Eggerthella species.52 Thus, microbiota differences are a factor that influences the outcome of the health effects of lignans.44 Furthermore, a study hypothesized that if high lignan products are consumed for an extended period of time, the microbiota will most likely adapt to metabolizing lignans, increasing the plasma concentration of the resulting enterolignans.27
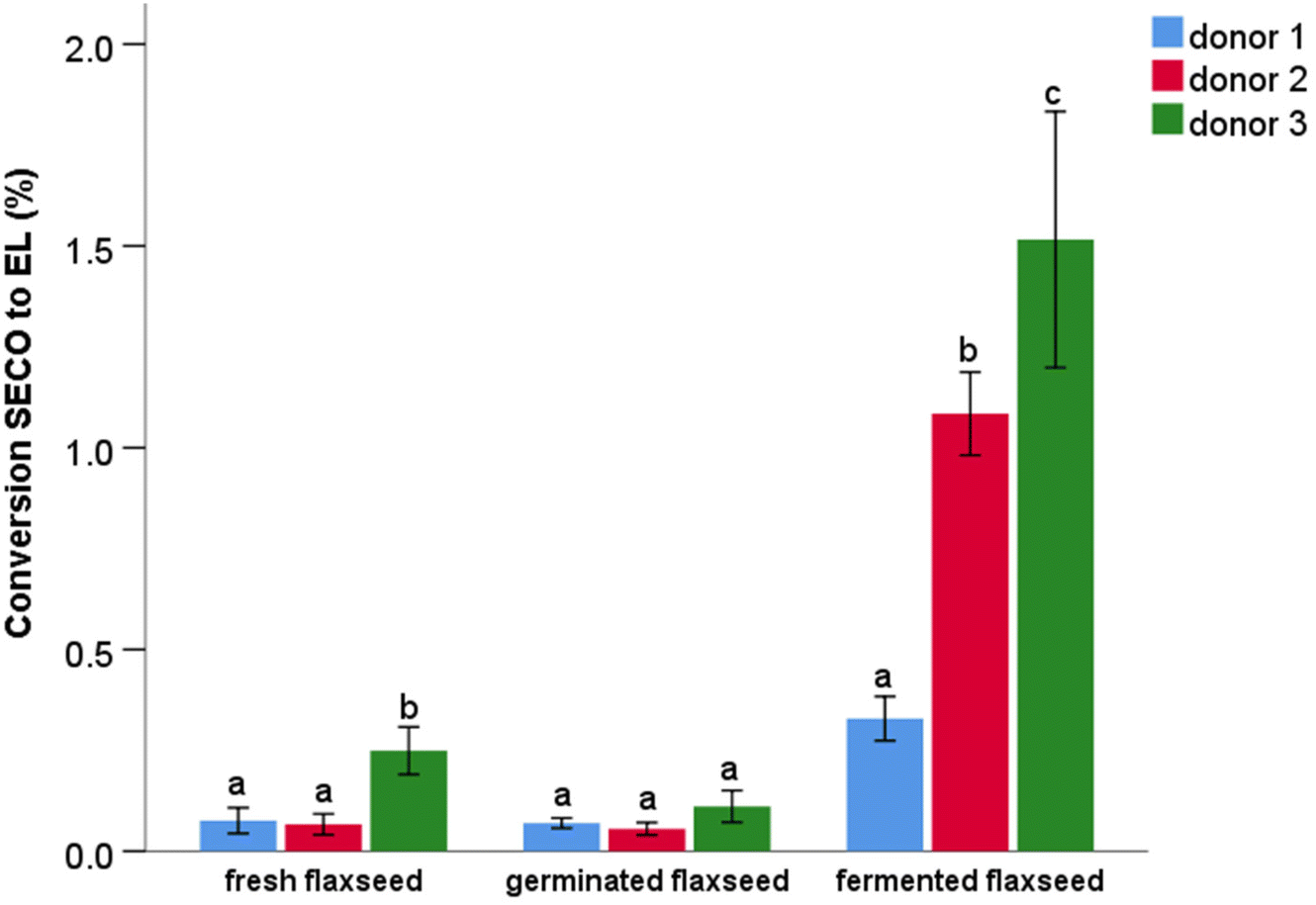
Only PINO and MATA were detected in the tea brew in colonic fermentation. The conversion of lignans (PINO and MATA) in tea brew was relatively high, approximately 15% (Fig. 4A), which was much higher than the conversion rate observed from flaxseed matrixes. The most likely explanation for this difference in conversion yield may be the different level of lignans to which microbiota have been exposed during the fermentation experiments. This amount would be highest when germinated flaxseed was fermented with a value of 15.1 mg of lignans (in 1 g of fermented material), followed by fermented flaxseed, with a value of 11.1 mg. The lowest lignan content after the colonic fermentation was in fresh flaxseed, with a value of 3.0 mg. Compared to flaxseed matrixes, the amount of lignans in 10 mL of tea brew used in the fermentation was several orders of magnitude lower, i.e. 0.003 mg. We can speculate that the high lignan concentration may have saturated the capacity of the microbiota for lignan conversion into enterolignans. Furthermore, the presence of fibre and its fermentation in the flaxseed may have delayed microbial metabolism of lignan as suggested elsewhere.27 Another possible explanation for the higher conversion of lignans in tea is that MATA can be directly converted to EL, whereas SECO needs to be converted to MATA or END prior to EL production.53
 | ||
| Fig. 4 Lignan (PINO and MATA) conversion to EL during in vitro colon fermentation of tea brew. (A) Data are from 3 donors (mean ± SD, n = 3) at different fermentation times; (B) data are from two replications of tea brew (mean ± SD, n = 2) of 3 donors. Different lower case letters indicate a significant difference (P < 0.05) in lignan conversion to EL. EL = enterolactone. | ||
The influence of inter-individual differences on the conversion of lignans to EL was further examined (Fig. 4B). Donor 3 significantly showed again the highest production of EL (approximately 25%) compared to others.
Conclusion
In the current study, for the first time, a systematic investigation of lignan bioaccessibility during gastrointestinal digestion, its release, and conversion to EL in colonic fermentation from differently processed flaxseed is reported. Our findings highlight that only a limited fraction of lignans are released in the small intestine and that processing and particle size reduction have a limited effect on the bioaccessibility of lignans. Fermentation, however, might represent a potential strategy for enhancing lignan release and conversion to EL in the large intestine. Further research into particle size reduction can be conducted to assess the impact on lignan bioaccessibility and conversion to EL.Author contributions
Umi Kalsum Hussain Zaki: conceptualization, investigation, formal analysis, writing – original draft. Christos Fryganas: conceptualization, investigation, formal analysis, writing – review and editing. Laura Trijsburg: writing – review and editing. Edith Feskens: writing – review and editing. Edoardo Capuano: supervision, conceptualization, investigation, writing – review and editing.Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.Acknowledgements
The study was supported by the Food Quality & Design Department, Wageningen University & Research, the Netherlands, and funded by the Malaysian Agricultural Research and Development Institute (MARDI), Selangor, Malaysia.References
- I. Paterni, C. Granchi, J. A. Katzenellenbogen and F. Minutolo, Estrogen receptors alpha (ERα) and beta (ERβ): subtype-selective ligands and clinical potential, Steroids, 2014, 90, 13–29 CrossRef CAS PubMed.
- C. Hu, Y. V. Yuan and D. D. Kitts, Antioxidant activities of the flaxseed lignan secoisolariciresinol diglucoside, its aglycone secoisolariciresinol and the mammalian lignans enterodiol and enterolactone in vitro, Food Chem. Toxicol., 2007, 45, 2219–2227 CrossRef CAS PubMed.
- D. Bravo, Á. Peirotén, I. Álvarez and J. M. Landete, Phytoestrogen metabolism by lactic acid bacteria: Enterolignan production by Lactobacillus salivarius and Lactobacillus gasseri strains, J. Funct. Foods, 2017, 37, 373–378 CrossRef CAS.
- N. Danbara, T. Yuri, M. Tsujita-Kyutoku, R. Tsukamoto, N. Uehara and A. Tsubura, Enterolactone induces apoptosis and inhibits growth of Colo 201 human colon cancer cells both in vitro and in vivo, Anticancer Res., 2005, 25, 2269–2276 CAS.
- M. Vanharanta, S. Voutilainen, T. H. Rissanen, H. Adlercreutz and J. T. Salonen, Risk of cardiovascular disease–related and all-cause death according to serum concentrations of enterolactone: Kuopio Ischaemic Heart Disease Risk Factor Study, Arch. Intern. Med., 2003, 163, 1099–1104 CrossRef CAS PubMed.
- Y. Zhu, K. Kawaguchi and R. Kiyama, Differential and directional estrogenic signaling pathways induced by enterolignans and their precursors, PLoS One, 2017, 12, e0171390 CrossRef PubMed.
- T. Bohn, Dietary factors affecting polyphenol bioavailability, Nutr. Rev., 2014, 72, 429–452 CrossRef PubMed.
- M. Alminger, A. M. Aura, T. Bohn, C. Dufour, S. N. El, A. Gomes, S. Karakaya, M. C. Martínez-Cuesta, G. J. McDougall, T. Requena and C. N. Santos, In Vitro Models for Studying Secondary Plant Metabolite Digestion and Bioaccessibility, Compr. Rev. Food Sci. Food Saf., 2014, 13, 413–436 CrossRef CAS PubMed.
- L. P. Meagher and G. R. Beecher, Assessment of data on the lignan content of foods, J. Food Compos. Anal., 2000, 13, 935–947 CrossRef CAS.
- K. Katina, K. H. Liukkonen, A. Kaukovirta-Norja, H. Adlercreutz, S. M. Heinonen, A. M. Lampi, J. M. Pihlava and K. Poutanen, Fermentation-induced changes in the nutritional value of native or germinated rye, J. Cereal Sci., 2007, 46, 348–355 CrossRef CAS.
- K. R. Moelants, L. Lemmens, M. Vandebroeck, S. Van Buggenhout, A. M. Van Loey and M. E. Hendrickx, Relation between particle size and carotenoid bioaccessibility in carrot-and tomato-derived suspensions, J. Agric. Food Chem., 2012, 60, 11995–12003 CrossRef CAS PubMed.
- A. Kamal-Eldin, H. N. Lærke, K.-E. B. Knudsen, A.-M. Lampi, V. Piironen, H. Adlercreutz, K. Katina, K. Poutanen and P. Åman, Physical, microscopic and chemical characterisation of industrial rye and wheat brans from the Nordic countries, Food Nutr. Res., 2009, 53, 1912 CrossRef.
- R. Coda, K. Katina and C. G. Rizzello, Bran bioprocessing for enhanced functional properties, Curr. Opin. Food Sci., 2015, 1, 50–55 CrossRef.
- A. Makowska, A. Waśkiewicz and S. Chudy, Lignans in triticale grain and triticale products, J. Cereal Sci., 2020, 102939 CrossRef CAS.
- A. Ruiz de la Bastida, Á. Peirotén, S. Langa, I. Álvarez, J. L. Arqués and J. M. Landete, Metabolism of flavonoids and lignans by lactobacilli and bifidobacteria strains improves the nutritional properties of flaxseed-enriched beverages, Food Res. Int., 2021, 147, 110488 CrossRef CAS PubMed.
- E. Fernández-García, I. Carvajal-Lérida and A. Pérez-Gálvez, In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency, Nutr. Res., 2009, 29, 751–760 CrossRef PubMed.
- E. Biehler and T. Bohn, Methods for assessing aspects of carotenoid bioavailability, Curr. Nutr. Food Sci., 2010, 6, 44–69 CrossRef CAS.
- F. N. Widaningrum, S. Ermi and P. Endang Yuli, Introductory Study on Processing of Fermented Jack Bean (Canavalia Ensiformis), Indones. J. Agric. Postharvest Res., 2015, 12, 129–136 Search PubMed.
- T. H. Emaga, N. Rabetafika, C. S. Blecker and M. Paquot, Kinetics of the hydrolysis of polysaccharide galacturonic acid and neutral sugars chains from flaxseed mucilage, Biotechnol., Agron., Soc. Environ., 2012, 16(2), 139–147 CAS.
- M. Minekus, M. Alminger, P. Alvito, S. Ballance, T. Bohn, C. Bourlieu, F. Carriere, R. Boutrou, M. Corredig and D. Dupont, A standardised static in vitro digestion method suitable for food–an international consensus, Food Funct., 2014, 5, 1113–1124 RSC.
- C. Bourlieu, O. Ménard, K. Bouzerzour, G. Mandalari, A. Macierzanka, A. R. Mackie and D. Dupont, Specificity of infant digestive conditions: some clues for developing relevant in vitro models, Crit. Rev. Food Sci. Nutr., 2014, 54, 1427–1457 CrossRef PubMed.
- S. A. Poeker, A. Geirnaert, L. Berchtold, A. Greppi, L. Krych, R. E. Steinert, T. de Wouters and C. Lacroix, Understanding the prebiotic potential of different dietary fibers using an in vitro continuous adult fermentation model (PolyFermS), Sci. Rep., 2018, 8, 1–12 CAS.
- J. I. Mosele, A. Macià, M.-P. Romero, M.-J. Motilva and L. Rubió, Application of in vitro gastrointestinal digestion and colonic fermentation models to pomegranate products (juice, pulp and peel extract) to study the stability and catabolism of phenolic compounds, J. Funct. Foods, 2015, 14, 529–540 CrossRef CAS.
- J. L. Peñalvo, K. M. Haajanen, N. Botting and H. Adlercreutz, Quantification of lignans in food using isotope dilution gas chromatography/mass spectrometry, J. Agric. Food Chem., 2005, 53, 9342–9347 CrossRef PubMed.
- I. E. Milder, I. C. Arts, D. P. Venema, J. J. Lasaroms, K. Wähälä and P. C. Hollman, Optimization of a liquid chromatography− tandem mass spectrometry method for quantification of the plant lignans secoisolariciresinol, matairesinol, lariciresinol, and pinoresinol in foods, J. Agric. Food Chem., 2004, 52, 4643–4651 CrossRef CAS.
- N. P. Nørskov and K. E. B. Knudsen, Validated LC-MS/MS Method for the Quantification of free and bound lignans in cereal-based diets and feces, J. Agric. Food Chem., 2016, 64, 8343–8351 CrossRef.
- C. Fuentealba, F. Figuerola, A. M. Estévez, J. M. Bastías and O. Muñoz, Bioaccessibility of lignans from flaxseed (Linum usitatissimum L.) determined by single–batch in vitro simulation of the digestive process, J. Sci. Food Agric., 2014, 94, 1729–1738 CrossRef CAS.
- A. Kuijsten, I. C. W. Arts, P. van't Veer and P. C. H. Hollman, The Relative Bioavailability of Enterolignans in Humans Is Enhanced by Milling and Crushing of Flaxseed, J. Nutr., 2005, 135, 2812–2816 CrossRef CAS PubMed.
- I. Cantabrana, R. Perise and I. Hernández, Uses of Rhizopus oryzae in the kitchen, Int. J. Gastron. Food Sci., 2015, 2, 103–111 CrossRef.
- A. Starzyńska-Janiszewska, R. Duliński and B. Stodolak, Fermentation with edible Rhizopus strains to enhance the bioactive potential of hull-less pumpkin oil cake, Molecules, 2020, 25, 5782 CrossRef PubMed.
- G. Janarny and K. Gunathilake, Changes in rice bran bioactives, their bioactivity, bioaccessibility and bioavailability with solid-state fermentation by Rhizopus oryzae, Biocatal. Agric. Biotechnol., 2020, 23, 101510 CrossRef.
- L. Zhang, W. Gao, X. Chen and H. Wang, The effect of bioprocessing on the phenolic acid composition and antioxidant activity of wheat bran, Cereal Chem., 2014, 91, 255–261 CrossRef CAS.
- H. Feng, B. Nemzer and J. W. Devries, Sprouted grains: Nutritional value, production, and applications, Elsevier, 2018 Search PubMed.
- F. Hübner and E. K. Arendt, Germination of cereal grains as a way to improve the nutritional value: a review, Crit. Rev. Food Sci. Nutr., 2013, 53, 853–861 CrossRef PubMed.
- A. M. Vilas-Boas, M. E. Brassesco, A. C. Quintino, M. C. Vieira, T. R. Brandão, C. L. Silva, M. Azevedo and M. Pintado, Particle Size Effect of Integral Carob Flour on Bioaccessibility of Bioactive Compounds during Simulated Gastrointestinal Digestion, Foods, 2022, 11, 1272 CrossRef CAS PubMed.
- Y. Li, M. Li, L. Wang and Z. Li, Effect of particle size on the release behavior and functional properties of wheat bran phenolic compounds during in vitro gastrointestinal digestion, Food Chem., 2022, 367, 130751 CrossRef CAS PubMed.
- A. Yeop, J. Sandanasam, S. F. Pan, S. Abdulla, M. M. Yusoff and J. Gimbun, The effect of particle size and solvent type on the gallic acid yield obtained from Labisia pumila by ultrasonic extraction, FluidsChE, 2017, 111, 02008 Search PubMed.
- R. J. Green, A. S. Murphy, B. Schulz, B. A. Watkins and M. G. Ferruzzi, Common tea formulations modulate in vitro digestive recovery of green tea catechins, Mol. Nutr. Food Res., 2007, 51, 1152–1162 CrossRef CAS.
- L. Ryan and S. L. Prescott, Stability of the antioxidant capacity of twenty–five commercially available fruit juices subjected to an in vitro digestion, Int. J. Food Sci. Technol., 2010, 45, 1191–1197 CrossRef CAS.
- L. Alemany, A. Cilla, G. Garcia-Llatas, M. T. Rodriguez-Estrada, V. Cardenia and A. Alegría, Effect of simulated gastrointestinal digestion on plant sterols and their oxides in enriched beverages, Food Res. Int., 2013, 52, 1–7 CrossRef CAS.
- A. S. Hole, I. Rud, S. Grimmer, S. Sigl, J. Narvhus and S. Sahlstrøm, Improved bioavailability of dietary phenolic acids in whole grain barley and oat groat following fermentation with probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri, J. Agric. Food Chem., 2012, 60, 6369–6375 CrossRef CAS PubMed.
- A. Vinco Pimenta, T. Agrizzi Verediano, J. C. Souza Carneiro, N. M. Brunoro Costa and A. G. Vasconcelos Costa, Bioaccessibility and bioavailability of calcium in sprouted brown and golden flaxseed, J. Sci. Food Agric., 2021, 101, 2788–2798 CrossRef CAS PubMed.
- A. Fardet, A shift toward a new holistic paradigm will help to preserve and better process grain products’ food structure for improving their health effects, Food Funct., 2015, 6, 363–382 RSC.
- A. Senizza, G. Rocchetti, J. I. Mosele, V. Patrone, M. L. Callegari, L. Morelli and L. Lucini, Lignans and gut microbiota: An interplay revealing potential health implications, Molecules, 2020, 25, 5709 CrossRef CAS PubMed.
- E. Eeckhaut, K. Struijs, S. Possemiers, J.-P. Vincken, D. D. Keukeleire and W. Verstraete, Metabolism of the lignan macromolecule into enterolignans in the gastrointestinal lumen as determined in the simulator of the human intestinal microbial ecosystem, J. Agric. Food Chem., 2008, 56, 4806–4812 CrossRef CAS PubMed.
- H. Chang, S. Yao, D. Tritchler, M. A. Hullar, J. W. Lampe, L. U. Thompson and S. E. McCann, Genetic variation in steroid and xenobiotic metabolizing pathways and enterolactone excretion before and after flaxseed intervention in African American and European American women, Cancer Epidemiol., Biomarkers Prev., 2019, 28, 265–274 CrossRef.
- G. Corona, A. Kreimes, M. Barone, S. Turroni, P. Brigidi, E. Keleszade and A. Costabile, Impact of lignans in oilseed mix on gut microbiome composition and enterolignan production in younger healthy and premenopausal women: An in vitro pilot study, Microb. Cell Fact., 2020, 19, 1–14 CrossRef PubMed.
- I. R. Rowland, H. Wiseman, T. A. Sanders, H. Adlercreutz and E. A. Bowey, Interindividual variation in metabolism of soy isoflavones and lignans: influence of habitual diet on equol production by the gut microflora, Nutr. Cancer, 2000, 36, 27–32 CrossRef CAS PubMed.
- A. F. Brito and Y. Zang, A review of lignan metabolism, milk enterolactone concentration, and antioxidant status of dairy cows fed flaxseed, Molecules, 2018, 24, 41 CrossRef.
- M. Á. Ávila-Gálvez, J. A. Giménez-Bastida, A. González-Sarrías and J. C. Espín, New insights into the metabolism of the flavanones eriocitrin and hesperidin: A comparative human pharmacokinetic study, Antioxidants, 2021, 10, 435 CrossRef PubMed.
- M. A. Hullar, S. M. Lancaster, F. Li, E. Tseng, K. Beer, C. Atkinson, K. Wähälä, W. K. Copeland, T. W. Randolph and K. M. Newton, Enterolignan-producing phenotypes are associated with increased gut microbial diversity and altered composition in premenopausal women in the United States, Cancer Epidemiol., Biomarkers Prev., 2015, 24, 546–554 CrossRef CAS PubMed.
- L. Roncaglia, A. Amaretti, S. Raimondi, A. Leonardi and M. Rossi, Role of bifidobacteria in the activation of the lignan secoisolariciresinol diglucoside, Appl. Microbiol. Biotechnol., 2011, 92, 159–168 CrossRef CAS.
- R. Consonni and G. Ottolina, NMR Characterization of Lignans, Molecules, 2022, 27, 2340 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d2fo02559k |
| This journal is © The Royal Society of Chemistry 2022 |