
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
A Lupinus angustifolius protein hydrolysate exerts hypocholesterolemic effects in Western diet-fed ApoE−/− mice through the modulation of LDLR and PCSK9 pathways†
Guillermo
Santos-Sánchez‡
 abc,
Ivan
Cruz-Chamorro‡
abc,
Carlotta
Bollati
a,
Martina
Bartolomei
a,
Justo
Pedroche
d,
Francisco
Millán
d,
María del Carmen
Millán-Linares
bd,
Anna Laura
Capriotti
e,
Andrea
Cerrato
e,
Aldo
Laganà
e,
Anna
Arnoldi
a,
Antonio
Carrillo-Vico
*bc and
Carmen
Lammi
*a
abc,
Ivan
Cruz-Chamorro‡
abc,
Carlotta
Bollati
a,
Martina
Bartolomei
a,
Justo
Pedroche
d,
Francisco
Millán
d,
María del Carmen
Millán-Linares
bd,
Anna Laura
Capriotti
e,
Andrea
Cerrato
e,
Aldo
Laganà
e,
Anna
Arnoldi
a,
Antonio
Carrillo-Vico
*bc and
Carmen
Lammi
*a
aDepartment of Pharmaceutical Sciences, University of Milan, 20133 Milan, Italy. E-mail: carmen.lammi@unimi.it
bDepartamento de Bioquímica Médica y Biología Molecular e Inmunología, Universidad de Sevilla, 41009 Seville, Spain. E-mail: vico@us.es
cInstituto de Biomedicina de Sevilla, IBiS (Universidad de Sevilla, HUVR, Junta de Andalucía, CSIC), 41013 Seville, Spain
dDepartment of Food & Health, Instituto de la Grasa, CSIC, Ctra, Utrera Km 1, 41013 Seville, Spain
eDipartimento di Chimica, Sapienza Università di Roma, Piazzale Aldo Moro 5, 00185 Rome, Italy
First published on 2nd March 2022
Abstract
Lupin protein hydrolysates (LPHs) are gaining attention in the food and nutraceutical industries due to their several beneficial health effects. Recently, we have shown that LPH treatment reduces liver cholesterol and triglyceride levels in hypercholesterolemic mice. The aim of this study was to elucidate the effects of LPH treatment on the molecular mechanism underlying liver cholesterol metabolism in ApoE−/− mice fed the Western diet. After identifying the composition of the peptide within the LPH mixture and determining its ability to reduce HMGCoAR activity in vitro, its effect on the LDLR and PCSK9 pathways was measured in liver tissue from the same mice. Thus, the LPH reduced the protein levels of HMGCoAR and increased the phosphorylated inactive form of HMGCoAR and the pHMGCoAR/HMGCoAR ratio, which led to the deactivation of de novo cholesterol synthesis. Furthermore, the LPH decreased the protein levels of SREBP2, a key upstream transcription factor involved in the expression of HMGCoAR and LDLR. Consequently, LDLR protein levels decreased in the liver of LPH-treated animals. Interestingly, the LPH also increased the protein levels of pAMPK responsible for HMGCoAR phosphorylation. Furthermore, the LPH controlled the PSCK9 signal pathway by decreasing its transcription factor, the HNF1-α protein. Consequently, lower PSCK9 protein levels were found in the liver of LPH-treated mice. This is the first study elucidating the molecular mechanism at the basis of the hypocholesterolemic effects exerted by the LPH in an in vivo model. All these findings point out LPHs as a future lipid-lowering ingredient to develop new functional foods.
Introduction
Lupin is a leguminous plant belonging to the Fabaceae family. Among the Lupinus spp., L. albus (white lupin), L. angustifolius (narrow-leaf lupin), and L. luteus (annual-yellow lupin) are the most cultivated for either animal or human nutrition.1 Proteins derived from lupin seeds are gaining attention as a source of bioactive peptides. In fact, several pieces of evidence highlight the biological activities of lupin protein hydrolysates and peptides, including hypocholesterolemic, hypoglycemic, antimicrobial, anti-inflammatory, and immunomodulatory effects.2–10 The health-promoting effects of lupin peptides have been tested in different animal models and clinical trials. Briefly, diets containing protein from L. albus11,12 or L. angustifolius13–15 significantly reduced both total cholesterol and low-density lipoprotein (LDL) cholesterol (LDL-C) levels compared to control diets containing casein in a rat model of hypercholesterolemia. In particular, the cultivar Vitabor of lupin (L. angustifolius) administered to rats reduced triglycerides and total cholesterol through the reduction of the expression of genes SREBP-1c and HMG-CoA reductase.13 In agreement with these studies, it was also demonstrated that the lupin protein isolate from L. albus provides hepatoprotective and hypocholesterolaemic effects in a hamster model with a similar mechanism of action.16An uncontrolled clinical trial on L. albus17 and two controlled trials on L. angustifolius have confirmed the hypocholesterolemic activity in humans.18,19 Lupin protein-enriched diets including foods such as bread, biscuits, and pasta suppress the lipid profile and blood pressure in both normal and hypertensive human subjects.20–22 Similarly, lupin-based diets improved glycemic control in diabetic and normal human subjects.23 In addition, a recent study supports the pleiotropic effects of a lupin bioactive peptide-based functional food on the critical steps of atherosclerosis, including inflammation, oxidative stress, and cholesterol metabolism.24
Recently, we have demonstrated that the lupin protein hydrolysate (LPH) obtained after the hydrolysis with Alcalase® reduces hepatic steatosis and abdominal adiposity, as well as total cholesterol levels in apolipoprotein E-deficient (ApoE−/−) mice. Furthermore, the LPH increases the hepatic total antioxidant capacity and reduces the hepatic inflammatory status in these mice.25 In addition, a LPH-based beverage decreases the LDL-C/HDL-C ratio in healthy subjects (lupine-1 study), a significant predictive marker of cardiovascular disease.26
In light of these pieces of evidence, the present study was aimed at assessing the underlying molecular mechanism of cholesterol lowering by a LPH in the same animal model using a multidisciplinary strategy. Firstly, a detailed characterization of the LPH composition was performed by nano-HPLC-MS/MS and UHPLC-HRMS to identify medium- and short-sized peptides, respectively. Afterwards, in vitro experiments were carried out to evaluate the effect of a LPH on the 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCoAR) activity, the key enzyme involved in the cholesterol synthesis and the target of statins, the primary drugs used for the hypercholesterolemia treatment.27 Finally, the effects of the LPH on the cholesterol metabolism pathway in the liver tissue of ApoE−/− mice were investigated by performing protein quantification of sterol regulatory element-binding protein 2 (SREBP2), LDL receptor (LDLR), HMCCoAR, phospho-adenosine monophosphate (AMP) activated protein kinase (pAMPK), proprotein convertase subtilisin/kexin type 9 (PCSK9), and hepatocyte nuclear factor 1-α (HNF1-α).
Materials and methods
Reagents
All reagents and solvents employed were from commercial sources (see the ESI†).LPH preparation
The preparation and characterization of the LPH were carried out as previously described.28 Briefly, a lupin protein isolate was dissolved in distilled water and subsequently hydrolyzed with Alcalase® (Novozymes, Bagsvaerd, DK) for 15 min at pH 8, 50 °C. The protein hydrolysis was stopped by heating at 85 °C for 15 min. The solution was centrifuged (10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 437g for 15 min), and the supernatant was lyophilized and stored at room temperature, constituting the LPH. The absolute yield of the LPH was >50% with respect to the lupin protein isolate.
437g for 15 min), and the supernatant was lyophilized and stored at room temperature, constituting the LPH. The absolute yield of the LPH was >50% with respect to the lupin protein isolate.
Peptide purification and concentration
As previously described,29 solid-phase extraction (SPE) on C18 was used to concentrate medium-sized peptides. In this case, 1 mg of LPH was loaded on a 50 mg Bond Elut C18 EWP cartridge (Agilent, Santa Clara, USA). The cartridge was preliminary washed with 3 mL of acetonitrile (ACN) and conditioned with 3 mL of 0.1% trifluoroacetic acid (TFA); the sample was acidified with aqueous TFA to pH 2.5 and then loaded, and the cartridge was washed with 3 mL of 0.1% TFA. Peptides were eluted from the SPE column with 0.5 mL of ACN/H2O (50:50, v/v) containing 0.1% TFA, and dried in a SpeedVac SC250 Express (Thermo Savant, Holbrook, NY, USA). The residue was reconstituted in 150 μL of 0.1% formic acid in H2O for analysis.
Analysis of medium-sized peptides by nano high-performance liquid chromatography-MS/MS and peptide identification
Medium-sized peptides were analysed by nano HPLC on an Ultimate 3000 (Thermo Fisher Scientific, Bremen, Germany) coupled to an Orbitrap Elite mass spectrometer (Thermo Fisher Scientific), as previously described.30 The samples (20 μL) were on-line preconcentrated on a μ-precolumn (Thermo Fisher Scientific, 300 μm i.d. 5 mm Acclaim PepMap 100 C18, 5 μm particle size, 100 Å pore size) at 10 μL min−1 flow rate of H2O/ACN (99:1, v/v) containing 0.1% TFA (v/v). Then, they were separated on an EASY-Spray column (Thermo Fisher Scientific, 15 cm × 75 μm i.d. PepMap C18, 3 μm particles, 100 Å pore size) operated at 300 nL min−1 and 35 °C. Mobile phases were H2O (solvent A) and ACN (solvent B), both with 0.1% formic acid. The following linear gradient was used: 1% B for 5 min; 1–5% B for 2 min; 5–35% B for 90 min; 35–90% B for 3 min; finally, the column was washed with 90% B for 10 min and then equilibrated with 1% B for 29 min.
Peptide spectra were acquired in the 300–2000m/z range at 30000 resolution (full width at half maximum, FWHM, at m/z 400) for the full scan. MS/MS spectra were acquired at 15000 resolution (FWHM, at m/z 400) in top 10 data-dependent acquisition mode with the rejection of singly charged ions and unassigned charge states. Precursors were fragmented by higher-energy collisional dissociation with 35% normalized collision energy and a 2m/z isolation window. Dynamic exclusion was enabled with a repeat count of 1 and a repeat duration of 30 s with an exclusion duration of 20 s. For each sample, three technical replicates were performed. Raw data files were acquired using Xcalibur software (version 2.2, Thermo Fisher Scientific).
Raw data spectra were identified using Proteome Discoverer (version 1.3, Thermo Fisher Scientific) with the Mascot search engine (v2.3.02) using a protein sequence database downloaded from Uniprot for the taxonomy L. angustifolius (31386 sequences). Precursor ion tolerance was 10 ppm and fragment ion tolerance was 0.05 Da; no enzyme was used for digestion and methionine oxidation was selected as dynamic modification. A decoy function was used for false discovery rate calculation, which was set at 1%.
Purification of short-sized peptide sequences
SPE and cleanup of short peptides were carried out using cartridges packed with 500 mg Carbograph 4. The applied procedure has been optimized in a previous paper.31 Briefly, the cartridge was washed with 5 mL of dichloromethane (DCM)/MetOH, 80:20 (v/v) with 20 mmol L−1 TFA and 5 mL of MetOH with 20 mmol L−1 TFA. The activation was carried out by flushing with 10 mL of 0.1 mol L−1 HCl and it was finally conditioned with 10 mL of 20 mmol L−1 TFA. Then, the samples (1 mg mL−1 of proteins) were diluted in 10 mL of H2O with 20 mmol L−1 TFA and loaded into the cartridge, which was sequentially washed with 2 mL of 20 mmol L−1 TFA and 0.5 mL MetOH. Finally, the analytes were eluted by backflushing elution with 10 mL of DCM/MetOH and 80:20 (v/v) with 20 mmol L−1 TFA. The eluate was evaporated at room temperature in a Speed-Vac SC250 Express (Thermo Savant, Holbrook, NY, USA), and the residue was reconstituted in 200 μL of water for subsequent reversed-phase separation.
UHPLC-HRMS analysis and short-sized peptide identification
Short peptides were analysed with Vanquish binary pump H (Thermo Fisher Scientific) coupled to a hybrid quadrupole-Orbitrap mass spectrometer Q Exactive (Thermo Fisher Scientific) using a heated ESI source operating in positive ion mode. The mass-spectrometric strategy was set up as previously reported.32 Each sample (20 μL) was injected onto a Kinetex XB-C18 (100 × 2.1 mm, 2.6 μm particle size, Phenomenex, Torrance, USA). The chosen flow, column temperature, and gradient parameters are reported in our previous work without any modification.33 Untargeted suspect screening analysis was performed in top 5 data-dependent acquisition mode in the m/z range 150–750 with a resolution (FWHM, m/z 200) of 70000. Higher-energy collisional dissociation fragmentation was performed at 40% normalized collision energy at a resolution of 35000 (FWHM, m/z 200). An inclusion list, in which all precursor ions derived from the combination of the 20 amino acids in di-, tri-, and tetrapeptides were listed, was included in the method.32 Raw data files from three experimental replicates and a blank sample were processed by Compound Discoverer using a workflow specifically dedicated to short peptide analysis. The database of short peptide sequences with IDs, masses, and molecular formulas was implemented for the automatic matching of extracted m/z ratios. Extracted masses from the chromatograms were aligned and filtered to remove the background compounds present in the blank sample, features whose masses were not present in the database, and those not fragmented. The filtered features were manually validated by matching the experimental spectra with those generated in silico by mMass 5.5.34
Database searching, protein identification, and validation
The UniProt database (UniProt Consortium)35 was used to identify the origin proteins of medium-sized peptides, while the BIOPEP-UWM database36 was used to identify the peptides with a described hypolipidemic activity.HMGCoAR activity assay
The HMGCoAR enzyme activity was measured using the commercial HMGCoAR assay kit (Sigma-Aldrich, St Louis, MO, USA). The experiments were carried out at 37 °C following the manufacturer's instructions. Each reaction (200 μL) was performed by adding the reagents in the following order: assay buffer (162 μL); LPH (20 μL at a final concentration of 0.1, 0.5, 1.0, and 2.5 mg mL−1) or vehicle; NADPH (4 μL); substrate solution (12 μL); and finally, HMGCoAR (2 μL). Then, the samples were mixed, and the absorbance was measured at 340 nm using a Synergy H1 microplate reader (Biotek Instruments, Winooski, VT, USA) at 0 and 10 min. The inhibition caused by the LPH on the activity of HMGCoAR was calculated through the reduction of absorbance, which is directly proportional to the activity of the enzyme. 1.0 μM pravastatin (Pravastatin, Sigma-Aldrich) was used as a positive control.In vivo study design
The LPH was suspended in a physiological saline solution containing 0.25% of carboxymethylcellulose (Sigma Aldrich, St Louis, MO, USA). The experimental study design was described in detail in a previous paper.25 Briefly, 19 ApoE−/− mice were initially divided into two groups: mice that were fed with a standard diet (SD, n = 5) or Western diet (WD) (n = 14). The compositions of the SD (Envigo Teklad Global, 13% energy from fat)37 and the WD (TestDiet 58v8, 45% energy from fat) are shown in ESI Tables S1 and S2,† respectively. When the mice from the WD group turned 6 weeks old, they were randomly divided into two groups and daily intragastrically treated with 100 mg kg−1 LPH (WD + LPH, n = 7) or vehicle (WD, n = 7) for 12 weeks. The mice were housed in the animal facility of the Instituto de Biomedicina de Sevilla (IBiS) under standard conditions (12/12 light/day cycles, temperature 22 ± 2 °C and humidity < 55%) with free access to water and diet. The aim of this study was to determine whether the LPH was able to restore the altered lipid profile by the WD; for this motif, the SD group was necessary only to confirm that the WD ingestion induces an alteration of this profile, according to other study designs.25,38–43 At the end of the experiment, fasted mice were euthanized by an overdose intraperitoneal injection, set for each mouse, of sodium thiopental (50 mg kg−1, B. Braun Medical SA, Barcelona, Spain) that was effective within 1 min, and the liver was subsequently collected, rapidly frozen, and stored at −80 °C until use. Plasma and hepatic lipid profile parameters were quantified as described in the ESI.† The experimental protocol was performed under the Spanish legislation and the EU Directive 2010/63/EU for animal experiments and was approved by the Virgen Macarena and Virgen del Rocío University Hospitals ethical committee (reference 21/06/2016/105).Western blot analysis
Tissues were homogenized in lysis buffer (RIPA buffer + protease inhibitor cocktail + 1:100 PMSF + 1:100 Na–orthovanadate + 1:1000 β-mercaptoethanol). The protein concentration was determined by the Bradford method and 50 μg of total proteins were run into a pre-cast 7.5% sodium dodecyl sulfate–polyacrylamide gel (SDS–PAGE) at 130 V for 45 min and then transferred to a nitrocellulose membrane (Mini nitrocellulose Transfer Packs, BioRad, Hercules, CA, USA) using a Trans-Blot Turbo (BioRad) at 1.3 A and 25 V for 7 min, and stained with Ponceau red solution (Sigma-Aldrich). To investigate proteins of different sizes on the same membrane, the membrane was cut into horizontal stripes. The milk or BSA blocked membrane was overnight incubated at 4 °C with primary antibodies against SREBP2, HMGCoAR, LDLR, phospho AMPK (Thr172), phospho HMGCoAR (Ser872), PCSK9, HNF1-α, and β-actin. After washing, the blots were incubated with secondary antibodies conjugated with horseradish peroxidase. A chemiluminescent reagent (Euroclone, Milan, Italy) was used to visualize target proteins and densitometry analysis was performed using the software Image Lab 6.1 (BioRad). The internal control β-actin was used to normalize loading variations. ESI Fig. S1† shows that there are no differences in β-actin levels among the experimental groups.
Statistical analysis
All the datasets were checked for normal distribution by the D'Agostino and Pearson test. Since they are all normally distributed with p-values < 0.05, statistical analysis was carried out by one-way ANOVA followed by Tukey's post-hoc analysis (GraphPad Software, San Diego, CA, USA). Values were expressed as means ± SD; p-values ≤0.05 were considered to be significant.Results
Peptidomics characterization of the LPH
At the same time, short peptide identification was carried out using a previously developed method following an HRMS-based suspect screening approach.44 Notably, using a list containing the exact masses of precursors relative to all the possible combinations of the 20 natural amino acids (from two to four, resulting in 168400 unique combinations), a total of 282 unique amino acid sequences were identified in the LPH sample. However, under the operating conditions, it was not possible to discriminate the isobaric leucine and isoleucine using tandem mass spectra;45 therefore, the occurrence of the two amino acids within the identified sequence was retained as equally probable (ESI Table S4†).
Hypocholesterolemic activity of LPH sequences
The analysis of LPH sequences using BIOPEP allowed us to identify 55 sequences that contained a peptide motif associated with a hypolipidemic effect (Table 1). In detail, 28 sequences (50.9%) from conglutin proteins (α- and β-conglutin), and 27 sequences (49.1%) from non-conglutin proteins were identified. Of these, 20 sequences contained the peptide GGV or IVG, identified with IDs 9383 and 9384, respectively.| Effect | Ref. | Bioactive peptide motif | ID36 | Origin proteina | Accession numbera | N. sequences | Sequences |
|---|---|---|---|---|---|---|---|
| a According to “UniProtKB” (http://www.uniprot.org/). | |||||||
| HMGCoA reductase activity inhibitor | 51 | GGV | 9383 | α-Conglutin | F5B8V7 | 11 | ALEPDNRIESEGGVT |
| IVG | 9384 | F5B8V8 | ALEPDNRIESEGGVTE | ||||
| ALEPDNRVESEGGVTE | |||||||
| LEPDNRIESEGGVT | |||||||
| LEPDNRIESEGGVTE | |||||||
| LEPDNRVESEGGVT | |||||||
| LEPDNRVESEGGVTE | |||||||
| NALEPDNRIESEGGVT | |||||||
| NALEPDNRIESEGGVTE | |||||||
| NALEPDNRVESEGGVT | |||||||
| NALEPDNRVESEGGVTE | |||||||
| Non-conglutin proteins | 9 | ADKEGGGGGVAPNVGKF | |||||
| GRGREDKEVVEGRDKAGGVT | |||||||
| GVEGSMRPPGQNPGGVL | |||||||
| HIVGPDKKIKL | |||||||
| HIVGPDKKIKLS | |||||||
| KEGGVLPGIK | |||||||
| LVIVGDGGTGKTT | |||||||
| RYDTPTNPQGRYRSGGSGGVGGVN | |||||||
| SGREKEEEGERGRRGGVRPH | |||||||
| Hypolipidemic peptide | 52 | EF | 9580 | α-Conglutin | F5B8V7 | 3 | HQAGDEGFEF |
| F5B8V8 | HQAGDEGFEFIAF | ||||||
| F5B8V6 | SGFNDEFLEE | ||||||
| β-Conglutin | F5B8V9 | 4 | EFQSKPNTL | ||||
| F5B8W0 | EFQSKPNTLILPK | ||||||
| F5B8W1 | EFQSRPNTL | ||||||
| F5B8W2 | EFQSRPNTLILPK | ||||||
| F5B8W3 | |||||||
| F5B8W4 | |||||||
| Non-conglutin proteins | 17 | AEFPIFDKV | |||||
| DSKDEFLAIL | |||||||
| FKAEFPIFDKV | |||||||
| GGRVILHPITSPASEFE | |||||||
| GGRVILHPITSPASEFEHVE | |||||||
| KAEFPIFDKV | |||||||
| NTRGGRVILHPITSPASEF | |||||||
| NTRGGRVILHPITSPASEFE | |||||||
| NTRGGRVILHPITSPASEFEHVE | |||||||
| NVVLDEFGNPR | |||||||
| QEFPPRSNLDPETY | |||||||
| QEFPSRSKLDSK | |||||||
| RVRENPKSDVDLVEF | |||||||
| SIIEEFPRLV | |||||||
| SIIEEFPRLVTW | |||||||
| VGVPKKDAEFK | |||||||
| VLVGVPKKDAEFK | |||||||
| PCSK9/LDLR binding inhibition | 54 | GQEQSHQDEGVIVR (T9) | 9676 | β-Conglutin | F5B8V9 | 4 | DEQRHGQEQSHQDEGVIVR |
| F5B8W1 | EKQRRGQEQSHQDEGVIVR | ||||||
| F5B8W2 | EQRHGQEQSHQDEGVIVR | ||||||
| F5B8W4 | GQEQSHQDEGVIVR | ||||||
| Non-conglutin proteins | 1 | EEQSRGQEQSHQDEGVIVR | |||||
| HMGCoA reductase inhibitor | 53 and 67 | LTFPG (P7a) | β-Conglutin | F5B8V9 | 6 | GLTFPGSTEDVERL | |
| F5B8W0 | LTFPGSAQDVERL | ||||||
| F5B8W2 | LTFPGSIEDVERL | ||||||
| F5B8W5 | LTFPGSIEDVERLIK | ||||||
| LTFPGSTEDVERL | |||||||
| LTFPGSVEDVERL | |||||||
| Total | 55 | ||||||
Biological effects of the LPH
 | ||
| Fig. 1 In vitro HMGoAR activity inhibition. Inhibition ability of the LPH, tested at different concentrations (0.1–2.5 mg mL−1), in in vitro experiments on HMGoAR activity. Data were expressed as a percentage of the control group and represented as the mean ± SD. ****, p < 0.0001. Different letters represent a statistical difference (p ≤ 0.05). C, control group; LPH, lupine protein hydrolysate; PRAV, pravastatin (1.0 μM). | ||
LPH ingestion does not alter the body weight gain and improves the lipid profile
As shown in Fig. 2A and B, no differences were observed either in the baseline body weights (SD: 20.83 ± 0.67g; WD: 21.56 ± 2.14g; WD + LPH: 21.32 ± 2.26g; p = 0.838) and the final body weight (SD: 26.10 ± 2.26; WD: 31.51 ± 4.72g; WD + LPH: 30.31 ± 5.98g; p = 0.222) among the three experimental groups. However, a significant increase (p = 0.046) in the body weight gain was observed in the WD group (+9.96 ± 2.71 g) in comparison with the SD group (+5.28 ± 1.91 g) (Fig. 2C). Furthermore, no significant differences (p = 0.949) were observed in the body weight gain when the mice were fed with the WD and treated with the LPH (+9.51 ± 3.65 g) compared to the WD group (Fig. 2C). Additionally, significant differences (p < 0.0001) in the daily food intake were observed between the SD group (8.31 ± 0.98 kcal per mouse) and WD group (10.43 ± 0.82 kcal per mouse), as well as between the SD group and WD + LPH group (10.14 ± 0.77 kcal per mouse) (Fig. 2D). However, no changes in daily food intake were observed between the treated mice (p = 0.503) in comparison to the WD group (Fig. 2D). On the other hand, the lipid profile was analysed in the plasma and liver of all experimental groups. In the WD + LPH group, a reduction in the plasma levels of total cholesterol (TC) (−60 mg dL−1 = −19.7%, p = 0.020), LDL-C (−66 mg dL−1 = −21.6%, p = 0.026), and TG (−27 mg dL−1 = −27.5%, p = 0.036) was observed compared to the WD group, while no modifications were observed in the SD group (Table 2). Moreover, in the WD + LPH group a mitigation of the significant increase in TC (p = 0.008), LDL-C (p = 0.006) and TG (p = 0.005) generated by the consumption of the WD, compared to the consumption of a SD, was detected. In addition, HDL-C did not show significant differences between the three experimental groups (p = 0.373) (Table 2). In a similar way, LPH intake, in the WD + LPH group, reduced the hepatic levels of TC (−0.5 mg g−1 tissue = −19.0%, p < 0.05) and TG (−13.1 mg g−1 tissue = 26.0%, p < 0.01) in comparison to the WD group, reaching similar values to SD-fed mice (Table 2). | ||
| Fig. 2 Body weight parameters and daily food intake. Baseline body weight (A), final body weight (B), body weight gain (C), and daily food intake (D) of all experimental groups. Values are shown as the mean ± SD. *, p ≤ 0.05. Different letters represent a statistical difference (p ≤ 0.05). n.s., not significant; LPH, lupine protein hydrolysate. | ||
| Parameters | Plasma (mg dL−1) | Liver (mg g−1 tissue) | ||||||
|---|---|---|---|---|---|---|---|---|
| SD | WD | WD + LPH | p-Value | SD | WD | WD + LPH | p-Value | |
| Data are expressed as mean ± standard deviation. Total cholesterol (TC) and triglyceride (TG) concentrations were quantified in the plasma and hepatic tissue. Low-density lipoprotein cholesterol (LDL-C) and high-density lipoprotein cholesterol (HDL-C) concentrations were quantified in the plasma. WD: Western diet-fed mice group; LPH, lupine protein hydrolysate-treated mice group. The p-value indicates the statistical analysis carried out by One-way ANOVA between the three experimental groups within each tissue. Different letters represent a statistical difference (p ≤ 0.05). | ||||||||
| TC | 254.3 ± 27.21a | 337.2 ± 28.45b | 270.8 ± 39.87a | 0.006 | 1.50 ± 0.31a | 2.42 ± 0.56b | 1.96 ± 0.31a | 0.003 |
| TG | 56.17 ± 9.52a | 98.60 ± 16.99b | 71.50 ± 9.88a | 0.005 | 27.83 ± 2.90a | 50.37 ± 14.42b | 37.26 ± 4.88a | 0.003 |
| LDL-C | 199.8 ± 32.77a | 281.6 ± 25.56b | 220.8 ± 34.61a | 0.005 | ||||
| HDL-C | 41.88 ± 12.00a | 35.80 ± 4.76a | 35.67 ± 4.41a | 0.373 | ||||
LPH modulates the activation of the LDLR pathway induced by a WD
SREBP2 and LDLR proteins were quantified in the liver of the mice of the three experimental groups for investigating the effects of the LPH on the LDLR pathway. Fig. 3 shows that the LDLR pathway is activated when the mice are fed with a WD. More in detail, the WD ingestion increased the protein level of LDLR (Fig. 3A) and the SREBP2 transcription factor (Fig. 3B) by 47.1% ± 35.9% (p < 0.05) and 118.6% ± 42.7% (p < 0.01), respectively. In particular, in Fig. 3B, the qualitative differences in β-actin bands among the groups are not statistically significant from a quantitative point of view as indicated in Fig. S1,† where the β-actin levels among the groups are illustrated (see the ESI†). The treatment with the LPH for 12 weeks reduced both LDLR by 48.8% ± 21.2% (p < 0.05) and the protein levels of SREBP2 by 83.1% ± 18.7% (p < 0.01) compared with the WD group. Interestingly, the values in the WD + LPH group were similar to those for the SD-fed mice.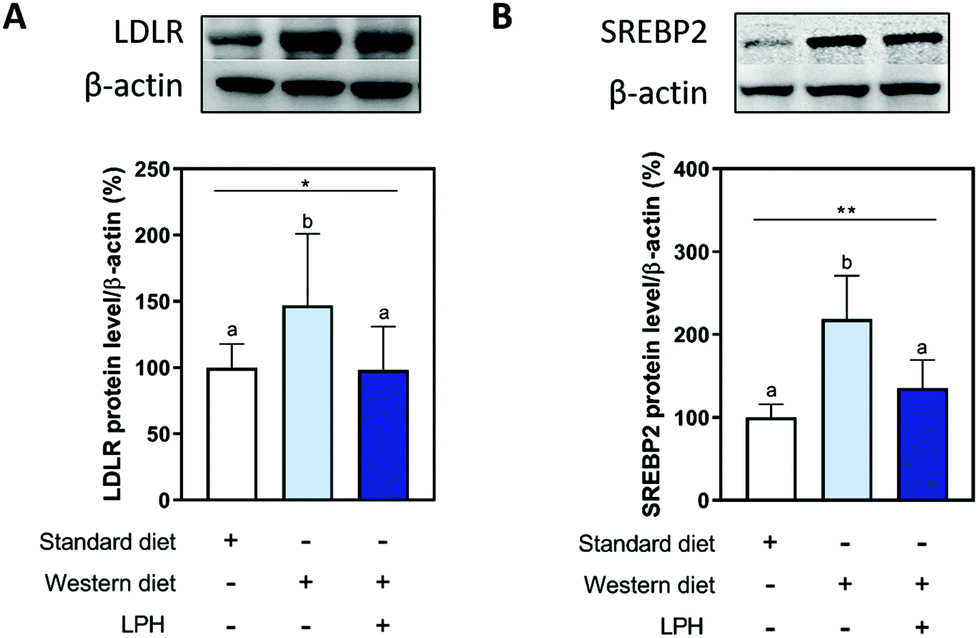 | ||
| Fig. 3 Modulation of the LDLR pathway by the LPH. The LPH modulates the increase of LDLR (A) and SREBP2 transcription factor (B) induced by WD. Data are represented as mean ± SD. *, p < 0.05; ** p < 0.01. Different letters represent a statistical difference (p ≤ 0.05). LPH, lupine protein hydrolysate; LDLR, low-density lipoprotein receptor; SREBP2, sterol regulatory element-binding protein 2. | ||
LPH reduces the WD-induced HMGCoAR activation
The effects of LPH on the HMGCoAR protein levels were evaluated by western blotting. As shown in Fig. 4, mice fed with the WD showed increased HMGCoAR protein levels by 53.0% ± 33.7% compared to the SD fed mice (p < 0.05). LPH treatment reduced 61.8% ± 12.4% of the HMGCoAR protein levels (p < 0.05) with respect to the WD-fed mice, and similar values to the SD group were obtained (p = 0.99) (Fig. 4A). Moreover, the treatment with the LPH significantly increased the phosphorylation levels of HMGCoAR (serine 872, AMPK phosphorylation site) by 22.8% ± 4.2% compared with the WD group (p < 0.05) (Fig. 4B). This result was in line with the increase observed in the pAMPK (threonine 172) level, which increased by 38.7% ± 2.5% in the LPH-treated mice in comparison to the WD group (p < 0.05) (Fig. 4C). | ||
| Fig. 4 Effects of the LPH on HMGCoAR, pHMGCoAR, and pAMPK protein levels. Representative western blots and densitometric analyses of the HMGCoAR (A), pHMGCoAR (B), and pAMPK (C) protein levels. The ratio of the inactive phosphorylated and active non-phosphorylated forms of HMGCoAR (D). Data are represented as mean ± SD. *, p < 0.05. Different letters represent a statistical difference (p ≤ 0.05). LPH, lupine protein hydrolysate; HMGCoAR, 3-hydroxy-3-methylglutaryl coenzyme A reductase; pHMGCoAR, phosphoHMGCoAR; pAMPK, phospho adenosine monophosphate activated protein kinase. | ||
Furthermore, the pHMGCoAR/HMGCoAR ratio was calculated. As shown in Fig. 3D, the ratio reduced by 36.8% ± 1.0% in the WD-fed mice compared to the SD-fed mice (p < 0.05). The treatment with the LPH overcomes this effect by increasing the pHMGCoAR/HMGCoAR ratio by 25.7% ± 8.7% compared to the WD group (p < 0.01). No differences between the WD + LPH and SD groups were observed (p = 0.581).
LPH treatment reduces the PCSK9 protein levels increased by WD ingestion
To elucidate the effects of the LPH treatment on the PCSK9 pathway, PCSK9 and HNF1-α protein levels were analysed by western blotting. As shown in Fig. 4, the ingestion of the WD increased the protein levels of PCSK9 (Fig. 4A) and HNF1-α (Fig. 4B) by 17.1% ± 8.7% (p < 0.05) and 39.3% ± 0.3% (p < 0.05), respectively, compared to the SD-fed mice. The LPH treatment for 12 weeks palliates these effects. The mice treated with the LPH showed a significant reduction of PCSK9 by 21.5% ± 6.4% (p < 0.05) and a significant reduction of HNF1-α by 43.8% ± 7.6% (p < 0.05) in comparison to the WD group, with no differences between the WD + LPH and SD-fed mice groups.Discussion
LPHs are a peptide mixture obtained after the hydrolysis of L. angustifolius protein using Alcalase®, which exerts both in vitro and in vivo anti-inflammatory and immunomodulatory effects.26,28,46,47In addition, LPH intake reduces abdominal adiposity and steatosis, and increases the hepatic total antioxidant capacity in WD-ApoE−/− mice.25 These pieces of evidence prompted us to study the molecular events behind the cholesterol-lowering properties of the LPH. To achieve this goal, a multidisciplinary study has been conducted through the combination of analytical, molecular, and biochemical techniques.
More in detail, since the biological activity of a specific protein hydrolysate is strictly correlated with its chemical composition, the identification of LPH peptide sequences was carried out using the most updated analytical techniques. Indeed, the characterization of the medium-sized peptide revealed the presence of 278 peptides. In total, 58% of these peptides derived from conglutins, the most abundant storage proteins in lupin seeds.48 Notably, 89, 71, and 1 peptide belong to α-, β-, and δ-conglutin, respectively, while no peptides from γ-conglutin were identified. It is known that the identification of short peptides is a challenging topic for several reasons.49 Standard proteomics database search engines cannot efficiently annotate short-sized peptides, since their short sequence cannot be confidently associated with single proteins.50 Moreover, the short sequences generate mostly singly charged ions, which are non-compatible with proteomics workflows and generate noisier MS/MS spectra. Finally, the fragmentation pathways of short peptide sequences are more strongly affected by the nature of the single amino acid that constitutes the sequences.32
Nevertheless, by applying a high-resolution mass spectrometry-based suspect screening approach, these particular issues were overcome and it is used to identify, for the first time, the sequence of short peptides within a lupin protein hydrolysate.
The parallel analysis of medium and short peptide sequences using the BIOPEP database allowed us to identify some sequences containing some known hypocholesterolemic motifs. More in detail, GGV and IVG motifs, identified for the first time in Amaranthus cruentus protein, show an HMGCoAR activity inhibition.51 In total, 24 sequences contained the EF motif (ID 9580), identified in Allium cepa, which attenuates lipogenesis in hepatocytes.52 Known peptides were also identified containing the P7a (LTFPG) fragmnet of the P7 (LTFPGSAED) that has been previously demonstrated to be able to reduce the in vitro HMGCoAR activity,53 whereas T9 (GQEQSHQDEGVIVR, ID 9676) possesses the capacity to directly inhibit HMGCoAR and PCSK9/LDLR protein–protein interaction.54
In light of these observations, our results suggest that the LPH decreases the in vitro HMGCoAR activity in a dose–response manner. Literature evidence suggests that the LPH exhibits a similar activity to a L. albus protein hydrolysate generated using trypsin (−37% at 0.25 mg mL−1, −57% at 0.5 mg mL−1, and −61% at 1.0 and 2.5 mg mL−1), and both are more active than a L. albus protein hydrolysate generated by digestion with pepsin, i.e. only −17% at 2.5 mg mL−1.9
In accordance with this evidence, we confirmed a decrease in lipid concentration both in plasma and liver tissue in the WD + LPH-treated mice compared to the WD-fed mice, reestablishing the normal values observed in SD-fed mice.
Based on these results, LPH effects were evaluated on the hepatic cholesterol metabolic pathway. Our findings show that mice fed with WD presented higher protein levels of LDLR, HMGCoAR, and SREBP2 than SD-fed mice, in agreement with previous studies.55–57 Specifically, the higher intake of dietary fatty acids can cause the activation of the cholesterol pathway in these mice for two reasons: (1) in situations of excess fatty acids, the generation of cholesterol activated by fatty acids gains priority over cholesterol downregulation; (2) regulation of cholesterol synthesis can be disrupted in the liver as a consequence of WD feeding.55,57 In this context, it is important to underline that even though dietary fat is known to affect the serum concentrations of total and lipoprotein cholesterol, all components of dietary triglycerides – saturated, monounsaturated, and polyunsaturated fatty acids – do not have identical effects on serum cholesterol levels.58 Notably, saturated fatty acids are mainly responsible for increasing cholesterol levels, whereas monounsaturated and mainly polyunsaturated fatty acids (i.e., omega-3) are able to lower cholesterol levels.59–61
Our results show that the LPH-treated mice had similar levels of LDLR, HMGCoAR, and SREBP2 to the SD-fed mice, supporting the LPH ability in counteracting the hepatic effects related to cholesterol metabolism caused by the WD consumption. Similar results have been previously observed with other natural compounds, i.e., polyphenols55 and spirulina lipids.56
HMGCoA reductase activity can be modulated by reversible phosphorylation–dephosphorylation, with the phosphorylated form of the enzyme being inactive (70%) and the dephosphorylated form active (30%). Thus, when HMGCoAR is phosphorylated the synthesis of de novo cholesterol is reduced. This short-term phosphorylation of HMGCoAR is induced by pAMPK that deactivates HMGCoAR by phosphorylation on Ser872. In this regard, statins can activate (phosphorylate) AMPK, inhibiting HMGCoAR. In this study, we observed an increase of the phosphorylated form of HMGCoAR and AMPK in the LPH-treated mice, compared to the WD group, which suggests that the LPH acts on HMGCoAR activity through the AMPK phosphorylation. The increase in the pHMGCoAR/HMGCoAR ratio, observed in the LPH-treated mice, supports the positive effect of the LPH on the exacerbated synthesis of cholesterol in the WD-fed mice. These results are in line with other results of the studies by our group carried out with peptides from L. albus in the human hepatic (HepG2) cell line.62,63
In addition, the LPH was able to counteract the increase of hepatic PCSK9 and HNF1-α protein levels observed in the WD-treated mice. The modulation of the PCSK9 pathway is highly significant as PCSK9 is a new important target for treating hypercholesterolemia.64 Therefore, these results confirm for the first time in an in vivo model the in vitro action mechanism previously disclosed for other L. albus peptides, through which lupin peptides synergistically modulate the cholesterol metabolism pathway.5,54,62,63,65 These results are also in line with previous studies that show a decrease of circulating PCSK9 levels in humans after lupin protein consumption.65,66 In this respect, several pieces of evidence indicate that diets containing either L. albus11,12 or L. angustifolius proteins13,14 significantly reduced both total cholesterol and LDL-C levels in vivo. In particular, specific studies have clearly underlined that the consumption of lupin proteins modulates the cholesterol pathway in both rat15 and hamster16 models, determining the hypocholesterolemic effect. In fact, with a more updated perception of the phenomenon, our study suggests that the observed health-promoting activity does not lie in the native protein, but a mixture of peptides generated from the physiological hydrolysis of proteins during digestion. Therefore, our study strongly suggests that the LPH might be directly used as a potential bioactive ingredient for the development of new dietary supplements and/or functional foods.
Conclusions
In conclusion, this is the first study to show the in vivo effects of a lupin protein hydrolysate on the hepatic cholesterol metabolic pathway (action mechanisms summarized in Fig. 5). Moreover, several sequences containing peptide motifs associated with hypolipidemic effects, HMGCoAR inhibition, and PCSK9/LDLR binding inhibition were identified within the LPH mixture. Therefore, this study confirms the pleotropic effects of the peptide mixture as a new possible ingredient for the development of supplements or functional foods with multifunctional activity including cholesterol lowering (Fig. 6). | ||
| Fig. 5 Effects of LPH treatment on the PCSK9 pathway. The LPH counteracts the increase of PCSK9 (A), and its transcription factor HNF1-α (B) caused by the ingestion of WD. Data are represented as mean ± SD. *, p < 0.05. Different letters represent a statistical difference (p ≤ 0.05). LPH, lupine protein hydrolysate; HNF1-α, hepatocyte nuclear factor 1-α; PCSK9, proprotein convertase subtilisin/kexin type 9. | ||
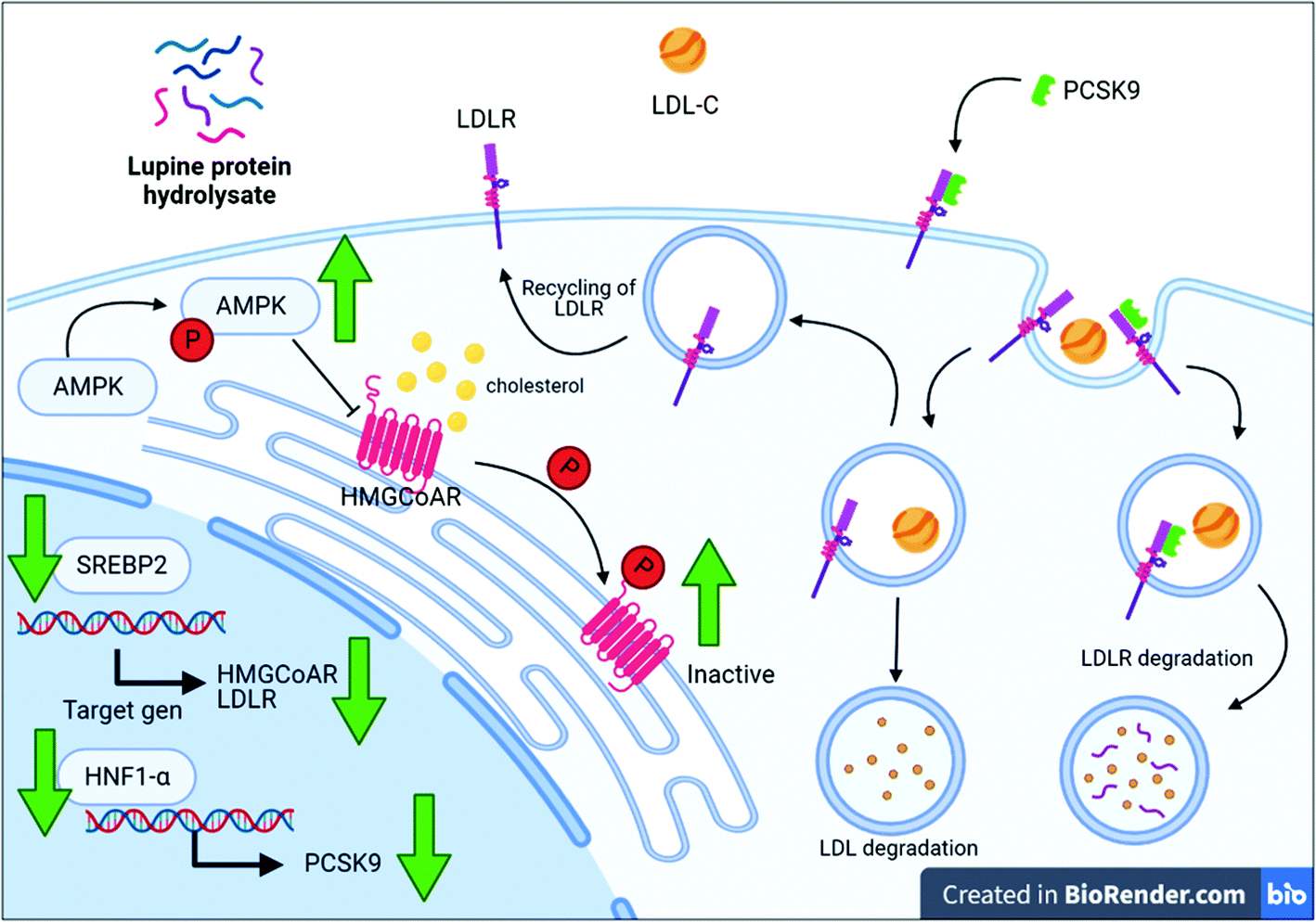 | ||
| Fig. 6 Schematic representation of the hypocholesterolemic mechanism of action mediated by the lupin protein hydrolysate in ApoE−/− mice. Green arrows show hepatic lupin protein hydrolysate (LPH) effects. AMPK, adenosine monophosphate (AMP) activated protein kinase; HMGCoAR, 3-hydroxy-3-methylglutaryl coenzyme A reductase; HNF1-α, hepatocyte nuclear factor 1-α; LDL-C, low-density lipoprotein cholesterol; LDLR, LDL-C receptor; P, inorganic phosphate; PCSK9, proprotein convertase subtilisin/kexin type 9; SREBP2, sterol regulatory element-binding protein 2. | ||
Abbreviation
| AMP | Adenosine monophosphate |
| AMPK | AMP activated protein kinase |
| C | WD-fed mice control group |
| CoA | Coenzyme-A |
| HMGCoAR | 3-Hydroxy-3-methylglutaryl CoA reductase |
| HNF1-α | Hepatocyte nuclear factor 1-α |
| LDL-C | Low-density lipoprotein cholesterol |
| LDLR | Low-density lipoprotein receptor |
| LPH | Lupine protein hydrolysate |
| PCSK9 | Proprotein convertase subtilisin/kexin type 9 |
| pHMGCoAR | Pospho-HMGCoAR |
| pAMPK | Phospho-AMP-activated protein kinase |
| SD | Standard diet-fed mice |
| SREBP2 | Sterol regulatory element-binding protein 2 |
| WD | Western diet. |
Author contributions
The following are the authors’ contributions: conceptualization: C.L. and A.C.-V.; methodology: G.S.-S., I.C.-C., C.B., M.B., A.L.C., A.C., and A.L.; data curation: G.S.-S., I.C.-C., C.B., M.B., AL.C., A.C., and A.L.; formal analysis: G.S.-S., I.C.-C., C.B., M.B., AL.C., A.C., and A.L; resources: C.L., A.C.-V., A.A., J.P., F.M., and M.C.M.-L.; writing – the original draft: G.S.-S., I.C.-C., C.L., and A.C.-V.; supervision: C.L. and A.C.-V.; funding acquisition: C.L., A.C.-V., and A.A.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This research was funded by the Andalusian Government Ministry of Health (PC-0111-2016-0111), and the PAIDI Program from the Andalusian Government (CTS160). I. C.-C. was supported by the VI Program of Inner Initiative for Research and Transfer of University of Seville (VIPPIT-2020-II.4) and a postdoctoral fellowship from the Andalusian Government Ministry of Economy, Knowledge, Business, and University (DOC_00587/2020). G. S.-S. was supported by a FPU grant from the Spanish Ministerio de Educación, Cultura y Deporte (FPU16/02339), and by an Erasmus+ Mobility Programme.The authors gratefully acknowledge the Carlo Sirtori Foundation (Milan, Italy) for having provided part of the equipment used in this experimentation. We thank all the staff from the Instituto de Biomedicina de Sevilla (IBiS) Animal Facility for their valuable assistance.
Notes and references
- A. Arnoldi, G. Boschin, C. Zanoni and C. Lammi, The health benefits of sweet lupin seed flours and isolated proteins, J. Funct. Foods, 2015, 18, 550–563 CrossRef CAS.
- M. C. Millan-Linares, A. Lemus-Conejo, M. M. Yust, J. Pedroche, A. Carrillo-Vico, F. Millan and S. Montserrat-de la Paz, GPETAFLR, a novel bioactive peptide from Lupinus angustifolius L. protein hydrolysate, reduces osteoclastogenesis, J. Funct. Foods, 2018, 47, 299–303 CrossRef CAS.
- M. C. Millan-Linares, R. Toscano, A. Lemus-Conejo, M. E. Martin, J. Pedroche, F. Millan and S. Montserrat-de la Paz, GPETAFLR, a biopeptide from Lupinus angustifolius L., protects against oxidative and inflammatory damage in retinal pigment epithelium cells, J. Food Biochem., 2019, 43, e12995 CrossRef PubMed.
- C. Lammi, G. Aiello, G. Vistoli, C. Zanoni, A. Arnoldi, Y. Sambuy, S. Ferruzza and G. Ranaldi, A multidisciplinary investigation on the bioavailability and activity of peptides from lupin protein, J. Funct. Foods, 2016, 24, 297–306 CrossRef CAS.
- C. Lammi, G. Aiello, C. Bollati, J. Li, M. Bartolomei, G. Ranaldi, S. Ferruzza, E. M. A. Fassi, G. Grazioso and Y. Sambuy, Trans-Epithelial Transport, Metabolism and Biological Activity Assessment of the Multi-Target Lupin Peptide LILPKHSDAD (P5) and Its Metabolite LPKHSDAD (P5-Met), Nutrients, 2021, 13, 863 CrossRef CAS PubMed.
- Y. Gao, X. Zhang, G. Ren, C. Wu, P. Qin and Y. Yao, Peptides from Extruded Lupin (Lupinus albus L.) Regulate Inflammatory Activity via the p38 MAPK Signal Transduction Pathway in RAW 264.7 Cells, J. Agric. Food Chem., 2020, 68, 11702–11709 CrossRef CAS PubMed.
- C. Lammi, C. Zanoni, A. Arnoldi and G. Aiello, YDFYPSSTKDQQS (P3), a peptide from lupin protein, absorbed by Caco–2 cells, modulates cholesterol metabolism in HepG2 cells via SREBP–1 activation, J. Food Biochem., 2018, 42, e12524 CrossRef.
- T. J. Guzmán, M. Düfer, M. Wiedemann, R. Olguín-Alor, G. Soldevila and C. M. Gurrola-Díaz, Lupin γ-conglutin protects against cell death induced by oxidative stress and lipotoxicity, but transiently inhibits in vitro insulin secretion by increasing KATP channel currents, Int. J. Biol. Macromol., 2021, 187, 76–90 CrossRef PubMed.
- C. Lammi, C. Zanoni, G. M. Scigliuolo, A. D'Amato and A. Arnoldi, Lupin peptides lower low-density lipoprotein (LDL) cholesterol through an up-regulation of the LDL receptor/sterol regulatory element binding protein 2 (SREBP2) pathway at HepG2 cell line, J. Agric. Food Chem., 2014, 62, 7151–7159 CrossRef CAS PubMed.
- C Lammi, C Zanoni, S Ferruzza, G Ranaldi, Y Sambuy and A Arnoldi, Hypocholesterolaemic activity of lupin peptides: Investigation on the crosstalk between human enterocytes and hepatocytes using a co-culture system including Caco-2 and HepG2 cells, Nutrients, 2018 DOI:10.3390/nu8070437.
- C. R. Sirtori, M. R. Lovati, C. Manzoni, S. Castiglioni, M. Duranti, C. Magni, S. Morandi, A. D'Agostina and A. Arnoldi, Proteins of white lupin seed, a naturally isoflavone-poor legume, reduce cholesterolemia in rats and increase LDL receptor activity in HepG2 cells, J. Nutr., 2004, 134, 18–23 CrossRef CAS PubMed.
- A. Bettzieche, C. Brandsch, K. Weiße, F. Hirche, K. Eder and G. I. Stangl, Lupin protein influences the expression of hepatic genes involved in fatty acid synthesis and triacylglycerol hydrolysis of adult rats, Br. J. Nutr., 2008, 99, 952–962 CrossRef CAS PubMed.
- A. Bettzieche, C. Brandsch, M. Schmidt, K. WEIßE, K. Eder and G. I. Stangl, Differing effect of protein isolates from different cultivars of blue lupin on plasma lipoproteins of hypercholesterolemic rats, Biosci., Biotechnol., Biochem., 2008, 0811061152–0811061152 Search PubMed.
- C. Parolini, E. Rigamonti, M. Marchesi, M. Busnelli, P. Cinquanta, S. Manzini, C. R. Sirtori and G. Chiesa, Cholesterol-lowering effect of dietary Lupinus angustifolius proteins in adult rats through regulation of genes involved in cholesterol homeostasis, Food Chem., 2012, 132, 1475–1479 CrossRef CAS PubMed.
- C. Brandsch, D. Kappis, K. Weiße and G. I. Stangl, Effects of untreated and thermally treated lupin protein on plasma and liver lipids of rats fed a hypercholesterolemic high fat or high carbohydrate diet, Plant Foods Hum. Nutr., 2010, 65, 410–416 CrossRef CAS PubMed.
- G. G. Fontanari, J. P. Batistuti, R. J. da Cruz, P. H. N. Saldiva and J. A. G. Arêas, Cholesterol-lowering effect of whole lupin (Lupinus albus) seed and its protein isolate, Food Chem., 2012, 132, 1521–1526 CrossRef CAS PubMed.
- M. Naruszewicz, G. Nowicka, L. Klosiewicz-Latoszek, A. Arnoldi and C. Sirtori, Effect of lupin protein (Lupinus albus) on cardiovascular risk factors in smokers with mild hypercholesterolemia, American Heart Association, 2006 Search PubMed.
- C. R. Sirtori, M. Triolo, R. Bosisio, A. Bondioli, L. Calabresi, V. De Vergori, M. Gomaraschi, G. Mombelli, F. Pazzucconi and C. Zacherl, Hypocholesterolaemic effects of lupin protein and pea protein/fibre combinations in moderately hypercholesterolaemic individuals, Br. J. Nutr., 2012, 107, 1176–1183 CrossRef CAS PubMed.
- M. Bähr, A. Fechner, M. Kiehntopf and G. Jahreis, Consuming a mixed diet enriched with lupin protein beneficially affects plasma lipids in hypercholesterolemic subjects: a randomized controlled trial, Clin. Nutr., 2015, 34, 7–14 CrossRef PubMed.
- F. Kamran, M. Phillips and N. Reddy, Functional properties of Australian blue lupin (Lupinus angustifolius) protein and biological activities of protein hydrolysates, Legume Sci., 2021, 3, e65 CAS.
- Y. P. Lee, T. A. Mori, I. B. Puddey, S. Sipsas, T. R. Ackland, L. J. Beilin and J. M. Hodgson, Effects of lupin kernel flour–enriched bread on blood pressure: a controlled intervention study, Am. J. Clin. Nutr., 2009, 89, 766–772 CrossRef CAS PubMed.
- R. Belski, T. A. Mori, I. B. Puddey, S. Sipsas, R. J. Woodman, T. R. Ackland, L. J. Beilin, E. R. Dove, N. Carlyon and V. Jayaseena, Effects of lupin-enriched foods on body composition and cardiovascular disease risk factors: a 12-month randomized controlled weight loss trial, Int. J. Obes., 2011, 35, 810–819 CrossRef CAS PubMed.
- R. Bouchoucha, M. Fradj, M. Bouchoucha, M. Akrout, M. Feki, N. Kaabachi, A. Raies and H. Slimane, Anti-hyperglycemic and anti-hyperlipidemic effects of Lupinus albus in type 2 diabetic patients: a randomized double-blind, placebo-controlled clinical trial, Int. J. Pharmacol., 2016, 12, 830–837 CAS.
- G. Santos-Sánchez, I. Cruz-Chamorro, A. I. Álvarez-Ríos, N. Álvarez-Sánchez, B. Rodríguez-Ortiz, A. I. Álvarez-López, M. S. Fernández-Pachón, J. Pedroche, F. Millán, M. C. Millán-Linares, P. J. Lardone, I. Bejarano, J. M. Guerrero and A. Carrillo-Vico, Bioactive peptides from Lupin (Lupinus angustifolius) prevent the early stages of atherosclerosis in Western diet-fed ApoE-/- mice, J. Agric. Food Chem., 2022 Search PubMed , Under Review.
- G. Santos-Sánchez, I. Cruz-Chamorro, A. I. Álvarez-Ríos, J. M. Fernández-Santos, M. V. Vázquez-Román, B. Rodríguez-Ortiz, N. Álvarez-Sánchez, A. I. Álvarez-López, M. d. C. Millán-Linares and F. Millán, Lupinus angustifolius Protein Hydrolysates Reduce Abdominal Adiposity and Ameliorate Metabolic Associated Fatty Liver Disease (MAFLD) in Western Diet Fed-ApoE−/− Mice, Antioxidants, 2021, 10, 1222 CrossRef PubMed.
- I. Cruz-Chamorro, N. Álvarez-Sánchez, A. I. Álvarez-Ríos, G. Santos-Sánchez, J. Pedroche, F. Millán, C. C. Sánchez, M. S. Fernández-Pachón, M. C. Millán-Linares and A. Martínez-López, Safety and Efficacy of a Beverage Containing Lupine Protein Hydrolysates on the Immune, Oxidative and Lipid Status in Healthy Subjects: An Intervention Study (the Lupine–1 Trial), Mol. Nutr. Food Res., 2021, 2100139 CrossRef CAS PubMed.
- M. Hu, B. M. Cheung and B. Tomlinson, Safety of statins: an update, Ther. Adv. Drug Saf., 2012, 3, 133–144 CrossRef PubMed.
- I. Cruz-Chamorro, N. Álvarez-Sánchez, M. del Carmen Millán-Linares, M. del Mar Yust, J. Pedroche, F. Millán, P. J. Lardone, C. Carrera-Sánchez, J. M. Guerrero and A. Carrillo-Vico, Lupine protein hydrolysates decrease the inflammatory response and improve the oxidative status in human peripheral lymphocytes, Food Res. Int., 2019, 126, 108585 CrossRef CAS PubMed.
- A. L. Capriotti, G. Caruso, C. Cavaliere, S. Piovesana, R. Samperi and A. Laganà, Proteomic characterization of human platelet-derived microparticles, Anal. Chim. Acta, 2013, 776, 57–63 CrossRef CAS PubMed.
- A. Fercha, A. L. Capriotti, G. Caruso, C. Cavaliere, R. Samperi, S. Stampachiacchiere and A. Laganà, Comparative analysis of metabolic proteome variation in ascorbate-primed and unprimed wheat seeds during germination under salt stress, J. Proteomics, 2014, 108, 238–257 CrossRef CAS PubMed.
- S. Piovesana, A. L. Capriotti, A. Cerrato, C. Crescenzi, G. La Barbera, A. Laganà, C. M. Montone and C. Cavaliere, Graphitized carbon black enrichment and UHPLC-MS/MS allow to meet the challenge of small chain peptidomics in urine, Anal. Chem., 2019, 91, 11474–11481 CrossRef CAS PubMed.
- A. Cerrato, S. E. Aita, A. L. Capriotti, C. Cavaliere, C. M. Montone, A. Laganà and S. Piovesana, A new opening for the tricky untargeted investigation of natural and modified short peptides, Talanta, 2020, 219, 121262 CrossRef CAS PubMed.
- C. M. Montone, A. L. Capriotti, A. Cerrato, M. Antonelli, G. La Barbera, S. Piovesana, A. Laganà and C. Cavaliere, Identification of bioactive short peptides in cow milk by high-performance liquid chromatography on C18 and porous graphitic carbon coupled to high-resolution mass spectrometry, Anal. Bioanal. Chem., 2019, 411, 3395–3404 CrossRef CAS PubMed.
- F. Spitaels, L. Li, A. Wieme, T. Balzarini, I. Cleenwerck, A. Van Landschoot, L. De Vuyst and P. Vandamme, Acetobacter lambici sp. nov., isolated from fermenting lambic beer, Int. J. Syst. Evol. Microbiol., 2014, 64, 1083–1089 CrossRef CAS PubMed.
- The UniProt Consortium, UniProt: The universal protein knowledgebase in 2021, Nucleic Acids Res., 2021, 49, D480–D489 CrossRef PubMed.
- P. Minkiewicz, A. Iwaniak and M. Darewicz, BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities, Int. J. Mol. Sci., 2019, 20, 5978 CrossRef CAS PubMed.
- ENVIGO, Teklad Global 14% Protein Rodent Maintenance Diet, https://insights.envigo.com/hubfs/resources/data-sheets/2014s-datasheet-0915.pdf.
- S. Li, J. You, Z. Wang, Y. Liu, B. Wang, M. Du and T. Zou, Curcumin alleviates high-fat diet-induced hepatic steatosis and obesity in association with modulation of gut microbiota in mice, Food Res. Int., 2021, 143, 110270 CrossRef CAS PubMed.
- M. C. Naranjo, B. Bermudez, I. Garcia, S. Lopez, R. Abia, F. J. Muriana and S. Montserrat-de la Paz, Dietary fatty acids on aortic root calcification in mice with metabolic syndrome, Food Funct., 2017, 8, 1468–1474 RSC.
- S. Montserrat-de la Paz, M. C. Naranjo, S. Lopez, R. Abia, F. J. Muriana and B. Bermudez, Niacin and olive oil promote skewing to the M2 phenotype in bone marrow-derived macrophages of mice with metabolic syndrome, Food Funct., 2016, 7, 2233–2238 RSC.
- M. Mahanna, M. C. Millan-Linares, E. Grao-Cruces, C. Claro, R. Toscano, N. M. Rodriguez-Martin, M. C. Naranjo and S. Montserrat-de la Paz, Resveratrol-enriched grape seed oil (Vitis vinifera L.) protects from white fat dysfunction in obese mice, J. Funct. Foods, 2019, 62, 103546 CrossRef CAS.
- M. Mijiti, R. Mori, B. Huang, K. Tsukamoto, K. Kiriyama, K. Sutoh and S. Nagaoka, Anti-obesity and hypocholesterolemic actions of protamine-derived peptide rpr (Arg-pro-arg) and protamine in high-fat diet-induced c57bl/6j mice, Nutrients, 2021, 13, 2501 CrossRef CAS PubMed.
- V. Kumar, S. Muthu Kumar and P. K. Tiku, Hypocholesterolemic Effect of Potent Peptide and Bioactive Fraction from Pigeon Pea By-Products in Wistar Rats, Int. J. Pept. Res. Ther., 2021, 27, 2403–2415 CrossRef CAS.
- A. Cerrato, S. E. Aita, C. Cavaliere, A. Laganà, C. M. Montone, S. Piovesana, R. Z. Chiozzi and A. L. Capriotti, Comprehensive identification of native medium-sized and short bioactive peptides in sea bass muscle, Food Chem., 2021, 343, 128443 CrossRef CAS PubMed.
- Y. Xiao, M. M. Vecchi and D. Wen, Distinguishing between leucine and isoleucine by integrated LC–MS analysis using an Orbitrap Fusion mass spectrometer, Anal. Chem., 2016, 88, 10757–10766 CrossRef CAS PubMed.
- M. del Carmen Millán-Linares, B. Bermúdez, M. del Mar Yust, F. Millán and J. Pedroche, Anti-inflammatory activity of lupine (Lupinus angustifolius L.) protein hydrolysates in THP-1-derived macrophages, J. Funct. Foods, 2014, 8, 224–233 CrossRef.
- M. del Carmen Millán-Linares, M. del Mar Yust, J. M. Alcaide-Hidalgo, F. Millán and J. Pedroche, Lupine protein hydrolysates inhibit enzymes involved in the inflammatory pathway, Food Chem., 2014, 151, 141–147 CrossRef PubMed.
- C. Villa, J. Costa and I. Mafra, Lupine allergens: Clinical relevance, molecular characterization, cross–reactivity, and detection strategies, Compr. Rev. Food Sci. Food Saf., 2020, 19, 3886–3915 CrossRef CAS PubMed.
- S. Piovesana, C. M. Montone, C. Cavaliere, C. Crescenzi, G. La Barbera, A. Laganà and A. L. Capriotti, Sensitive untargeted identification of short hydrophilic peptides by high performance liquid chromatography on porous graphitic carbon coupled to high resolution mass spectrometry, J. Chromatogr. A, 2019, 1590, 73–79 CrossRef CAS PubMed.
- V. R. Koskinen, P. A. Emery, D. M. Creasy and J. S. Cottrell, Hierarchical clustering of shotgun proteomics data, Mol. Cell. Proteomics, 2011, 10(6), M110.003822 CrossRef PubMed.
- R. A. M. Soares, S. Mendonça, L. Í. A. De Castro, A. C. C. C. C. Menezes and J. A. G. Arêas, Major peptides from amaranth (Amaranthus cruentus) protein inhibit HMG-CoA reductase activity, Int. J. Mol. Sci., 2015, 16, 4150–4160 CrossRef CAS PubMed.
- Y. G. Lee, J. Y. Cho, E. J. Hwang, T. I. Jeon and J. H. Moon, Glu–Phe from onion (Allium Cepa L.) attenuates lipogenesis in hepatocytes, Biosci., Biotechnol., Biochem., 2017, 81, 1409–1416 CrossRef CAS PubMed.
- C. Lammi, G. Aiello, L. Dellafiora, C. Bollati, G. Boschin, G. Ranaldi, S. Ferruzza, Y. Sambuy, G. Galaverna and A. Arnoldi, Assessment of the Multifunctional Behavior of Lupin Peptide P7 and Its Metabolite Using an Integrated Strategy, J. Agric. Food Chem., 2020, 68, 13179–13188 CrossRef CAS PubMed.
- C. Lammi, C. Bollati, D. Lecca, M. P. Abbracchio and A. Arnoldi, Lupin Peptide T9 (GQEQSHQDEGVIVR) Modulates the Mutant PCSK9D374Y Pathway: In vitro Characterization of its Dual Hypocholesterolemic Behavior, Nutrients, 2019, 11, 1665 CrossRef CAS PubMed.
- Y. Li, S. Xu, M. M. Mihaylova, B. Zheng, X. Hou, B. Jiang, O. Park, Z. Luo, E. Lefai and J. Y.-J. Shyy, AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice, Cell Metab., 2011, 13, 376–388 CrossRef CAS PubMed.
- Y. Yang, L. Du, M. Hosokawa and K. Miyashita, Effect of Spirulina lipids on high-fat and high-sucrose diet induced obesity and hepatic lipid accumulation in C57BL/6J mice, J. Funct. Foods, 2020, 65, 103741 CrossRef CAS.
- N. Wu, L. K. Sarna, S.-Y. Hwang, Q. Zhu, P. Wang, Y. L. Siow and O. Karmin, Activation of 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase during high fat diet feeding, Biochim. Biophys. Acta, Mol. Basis Dis., 2013, 1832, 1560–1568 CrossRef CAS PubMed.
- S. M. Grundy, Monounsaturated fatty acids and cholesterol metabolism: implications for dietary recommendations, J. Nutr., 1989, 119, 529–533 CrossRef CAS PubMed.
- A. C. Skulas-Ray, P. W. Wilson, W. S. Harris, E. A. Brinton, P. M. Kris-Etherton, C. K. Richter, T. A. Jacobson, M. B. Engler, M. Miller and J. G. Robinson, Omega-3 fatty acids for the management of hypertriglyceridemia: a science advisory from the American Heart Association, Circulation, 2019, 140, e673–e691 CrossRef CAS PubMed.
- A. Pizzini, L. Lunger, E. Demetz, R. Hilbe, G. Weiss, C. Ebenbichler and I. Tancevski, The role of omega-3 fatty acids in reverse cholesterol transport: A review, Nutrients, 2017, 9, 1099 CrossRef PubMed.
- P. M. Kris-Etherton, T. A. Pearson, Y. Wan, R. L. Hargrove, K. Moriarty, V. Fishell and T. D. Etherton, High–monounsaturated fatty acid diets lower both plasma cholesterol and triacylglycerol concentrations, Am. J. Clin. Nutr., 1999, 70, 1009–1015 CrossRef CAS PubMed.
- C. Zanoni, G. Aiello, A. Arnoldi and C. Lammi, Investigations on the hypocholesterolaemic activity of LILPKHSDAD and LTFPGSAED, two peptides from lupin β-conglutin: Focus on LDLR and PCSK9 pathways, J. Funct. Foods, 2017, 32, 1–8 CrossRef CAS.
- C. Lammi, C. Zanoni, S. Ferruzza, G. Ranaldi, Y. Sambuy and A. Arnoldi, Hypocholesterolaemic activity of lupin peptides: Investigation on the crosstalk between human enterocytes and hepatocytes using a co-culture system including Caco-2 and HepG2 cells, Nutrients, 2016, 8, 437 CrossRef PubMed.
- B. Chen, X. Shi, Y. Cui, A. Hou and P. Zhao, A review of PCSK9 inhibitors and their effects on cardiovascular diseases, Curr. Top. Med. Chem., 2019, 19, 1790–1817 CrossRef CAS PubMed.
- C. Lammi, C. Zanoni, L. Calabresi and A. Arnoldi, Lupin protein exerts cholesterol-lowering effects targeting PCSK9: From clinical evidences to elucidation of the in vitro molecular mechanism using HepG2 cells, J. Funct. Foods, 2016, 23, 230–240 CrossRef CAS.
- C. Pavanello, C. Lammi, M. Ruscica, R. Bosisio, G. Mombelli, C. Zanoni, L. Calabresi, C. R. Sirtori, P. Magni and A. Arnoldi, Effects of a lupin protein concentrate on lipids, blood pressure and insulin resistance in moderately dyslipidaemic patients: A randomised controlled trial, J. Funct. Foods, 2017, 37, 8–15 CrossRef CAS.
- C. Lammi, J. Sgrignani, G. Roda, A. Arnoldi and G. Grazioso, Inhibition of PCSK9D374Y/LDLR Protein–Protein Interaction by Computationally Designed T9 Lupin Peptide, ACS Med. Chem. Lett., 2018, 10, 425–430 CrossRef PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available: Table S1. Composition of the experimental Western diet; Table S2. Composition of the standard diet; Table S3. Medium-sized peptide identification; Table S4. Short-sized peptide identification. See DOI: 10.1039/d1fo03847h |
| ‡ These authors equally contributed to the work. |
| This journal is © The Royal Society of Chemistry 2022 |