
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Effect of high-amylose starch branching enzyme II wheat mutants on starch digestibility in bread, product quality, postprandial satiety and glycaemic response†
Marina
Corrado
 a,
Jennifer H.
Ahn-Jarvis
a,
Brendan
Fahy
b,
George M.
Savva
a,
Cathrina H.
Edwards‡
*a and
Brittany A.
Hazard‡
ab
a,
Jennifer H.
Ahn-Jarvis
a,
Brendan
Fahy
b,
George M.
Savva
a,
Cathrina H.
Edwards‡
*a and
Brittany A.
Hazard‡
ab
aFood Innovation and Health, Quadram Institute Bioscience, Norwich Research Park, UK. E-mail: cathrina.edwards@quadram.ac.uk
bDesigning Future Wheat and Molecules from Nature, John Innes Centre, Norwich Research Park, UK
First published on 26th January 2022
Abstract
High-amylose starch branching enzyme II (sbeII) mutant wheat has potential to be low-glycaemic compared to conventional wheat; however, the effects of bread made from sbeII wheat flour on glycaemic response and product quality require investigation. We report the impact of white bread made from sbeII wheat flour on in vitro starch digestibility and product quality, and on postprandial glycaemia in vivo, compared to an isoglucidic wild-type (WT) control white bread. Starch in sbeII bread was ∼20% less susceptible to in vitro amylolysis leading to ∼15% lower glycaemic response measured in vivo, compared to the WT control bread, without major effects on bread appearance or texture, measured instrumentally. Despite the early termination of the in vivo intervention study due to the COVID-19 outbreak (n = 8 out of 19), results from this study indicate that sbeII wheat produces bread with lower starch digestibility than conventional white bread.
Introduction
Large dietary intakes of high glycaemic starchy foods can exacerbate the metabolic abnormalities of insulin resistance and over time, lead to abnormal glucose tolerance, insulin sensitivity and hyperglycaemia.1 Approximately 65% of individuals with impaired glucose tolerance eventually develop type 2 diabetes,2 which affects ∼6% of the UK population.3 Carbohydrates (including starch) account for around 45% of dietary energy intake,4 however, not all carbohydrates have the same physiological effect. For instance, the postprandial rise in blood glucose and insulin concentrations varies considerably between starch-rich foods and is strongly influenced by the rate and extent of starch digestion by α-amylase in the small intestine.5 Foods eliciting low glycaemic responses (as measured by glycaemic index) are needed for dietary prevention and management of obesity, type 2 diabetes, and cardiovascular disease.6,7 The manipulation of starch structure through non-transgenic genetic approaches provides new opportunities to develop slowly digested carbohydrate staple foods with lower glycaemic potency.Starch structure can be manipulated in planta using Targeting Induced Local Lesions in Genomes (TILLING) technology8 to induce mutations in genes involved in starch biosynthesis.9 Wheat carrying mutations in starch branching enzyme II (sbeII) genes have been generated using TILLING, yielding starch with a higher proportion of amylose and increased resistance to amylase digestion, compared to conventional wheat starch.10,11 This allows the production of high-amylose starch-based food products without the need of post-extraction processing or supplementation with starches from other botanical sources, which was shown to be a suitable approach to lower glycaemic response to high glycaemic foods, such as bread.12,13
Previously, a sbeII mutant wheat was used to produce a semolina pudding with lower starch digestibility than the control pudding made from WT wheat. When tested in humans, however, the Glycaemic Index of the sbeII pudding was not significantly different from the WT control.14 Recently, two studies have investigated glycaemic responses to pasta and bread made from sbeII mutant wheat. Sissons et al. (2020),15 reported that a serving of pasta (spaghetti) made from a sbeIIa durum wheat mutant with starch made of ∼58% amylose, corresponding to ∼7% RS, led to a lower starch susceptibility to amylase digestion in vitro and lower glycaemic index measured in vivo, compared to the WT control. Belobrajdic et al. (2019),16 reported that bread made from high-amylose wheat flour lowered the postprandial glycaemic response by 39% and the insulinemic response by 24%, however the bread provided ∼24 g less starch per 100 g and ∼7.5% less energy (kJ per 100 g) than the control bread. Therefore, it is not clear if the attenuated postprandial glycaemic response following high-amylose bread in the Belobrajdic et al. study was due to the lower amount of starch per serving (glycaemic load) or due to a change in the intrinsic starch digestibility.
Further studies are needed to better understand how and to what extent sbeII wheat can be used to attenuate the glycaemic potency of different food products. The aim of the present study was to determine the effect of sbeII wheat in bread on postprandial glycaemic response and satiety, compared to WT control bread. Unlike a Glycaemic Index study, this postprandial study was focussed on understanding the effects of sbeII starch in bread on glycaemic response curve size and shape, compared to a conventional white bread, to inform on the potential health effect of replacing conventional white bread with sbeII bread.
The bread rolls were prepared to deliver the same amount of starch per serving (75 g), such that any observed differences in the glycaemic responses to these bread rolls reflect differences in intrinsic starch amylolysis (measured in vitro). A preliminary sensory assessment (palatability) and quality analysis of sbeII bread was also conducted however, due to the small number of participants completing the study, results were not conclusive. The methodology was reported in ESI.†
An additional aspect of this study was the measurement of postprandial glucose using a continuous glucose monitoring system (CGM) to assess glycaemic responses, in parallel with fingerprick (FP) sampling for capillary glucose measurements. CGMs were fitted to the upper arm and used enzymatic-amperometric technology to measure glucose concentrations in interstitial fluid (IF) over several days. Recent studies have used CGMs to investigate the effect of dietary interventions on glucose responses in healthy individuals.17,18 Dye et al.19 and Bajka et al.20 reported a good correlation between capillary and IF glucose concentrations despite a time lag between measurements, likely due to physiological differences in the regulation of glucose exchange between the blood and the interstitial compartments. In the present study, the relationship between CGM and FP measurements was explored.
Methods
Bread formulation and process
A sbeII mutant bread wheat and a WT control bread wheat (Triticum aestivum L. ssp. aestivum) were sown in spring 2018 in a field trial using a randomised block design at the John Innes Centre Church Farm field station (Bawburgh, UK). Wheat was harvested in summer 2018, grains were transported to Campden BRI (Chipping Campden, UK). Following tempering to 16.4% humidity, grains were debranned and milled on a Bühler mill with feeding rate of 100 g min−1, then sieved through 140 μm mesh size and passed through a bran finisher fitting a 125 μm mesh to produce white flour.The sbeII and WT control wheat bread rolls were produced using a straight-dough method (AACC Method 10-10.03) at the Quadram Institute (NHS QI Clinical Research Facility, Norwich, UK). The formulation of each bread was adjusted based on the flour's starch content so that both provided the same total starch (∼75 g). Briefly, flour (1217.7 g and 1201.4 g for sbeII and WT control, respectively), yeast (35.6 g and 33.9 g for sbeII and WT control), sugar (44.5 g and 42.4 g for sbeII and WT control), salt (22.3 g and 21.2 g for sbeII and WT control), shortening (40.7 g and 38.8 for sbeII and WT control), and water (867.2 g and 781.6 g for sbeII and WT control) were mixed at low speed for 3 min and kneaded for 5 min at increasing speed, in a heavy-duty planetary mixer with hook attachment (model 5KSM7591XBSM, Kitchen Aid, Antwerp, BE). Dough was fermented 2 h at 21 °C then portioned into rolls and proofed for 15 min at 38 °C with 100% relative humidity, then baked using a combination oven (Rational Self Cooking Centre, Luton, UK) for 15 min at 185 °C (40% humidity for 10 min and 10% humidity for the last 5 min). Baked rolls reached ∼95 °C core temperature. After 2 h cooling at 21 °C, breads were packed in resealable, opaque polyethylene bags (3 mm thickness) and stored at −20 °C. Breads were produced in four batches of 12 rolls (48 rolls per type of flour), under identical conditions and paired by their baking position in the oven. Rolls were thawed for 16 h (overnight) and either served to human study participants as part of their breakfast meal or used for in vitro analyses. Breads were consumed within three months of manufacture.
Bread characteristics and in vitro amylolysis
Five pairs of each bread type were randomly selected for in vitro analysis of starch characteristics. From each roll, a minimum of three technical replicates were sampled for analysis, as specified below. Rolls were left to thaw in their packaging at room temperature (∼21 °C) for 16 h, then were blended in a food processor (Kenwood CH 180 Mini chopper) for 50 seconds and sieved to obtain a 1 mm fraction, used for moisture determination and in vitro amylolysis assay, and a 500 μm fraction, used to measure digestible and resistant starch. These fractions were weighed into dry-tared aluminium pans in triplicates (technical replicates) for moisture determination, into 2 mL safe lock tubes (STARLAB (UK), Ltd) in quadruplicates (technical replicates) for starch measurements, and into 15 mL Corning centrifuge tubes (Merck KGaA, Darmstadt, DE) in triplicates (technical replicates) for amylolysis analysis. Digestible and resistant proportions of starch were measured using the reagents from ‘Total starch kit’ and ‘Resistant starch kit’ K-RSTAR (AOAC 2002.02, Megazyme International, Wicklow, Ireland), using a small-scale version of the method, as described by Edwards et al., 2015.21 Moisture was determined using the AACC (44-15A) air oven method, one stage procedure.
Crumb texture and bread colour were measured on crust and crumb. Texture was measured instrumentally using a ‘two-bite test’ (n = 4) on a TA-XT2 Texture Analyser (Stable Micro Systems, Godalming, UK),20 equipped with a five kg load cell using a modified AACC method 74–09. The Texture Analyser was equipped with a 50 mm diameter compression plate (P50); a uniaxial compression with crosshead speed of 100 mm min−1 was applied to 25 × 25 × 25 mm samples to mimic mastication, with crumb hardness corresponding to the force (N) required for 40% compression. Exponent (version 6.0, Stable Micro Systems, Godalming, UK) software for texture profile analysis was used to assess the following texture parameters: hardness, springiness, cohesiveness, gumminess, chewiness and resilience. Four independent measurements of colour were taken for each bread roll type, using a ‘Gretag Macbeth’ twenty-four patch Colour Checker as reference.26Hue angle and Chroma were calculated using the L*, a*, b* parameters.27 Parameters were compared using independent groups t-tests, values reported throughout are means ± SEMs (n = 4) unless otherwise specified.
Acute postprandial intervention study
Enrolled participants were randomised in blocks of four stratified by sex to complete 3 visits: Visit 1, to apply the CGM sensors, followed by Visit 2 and Visit 3, where participants consumed either sbeII or WT control bread on consecutive occasions, in randomised order, with a minimum of four days washout period between visits with no dietary restriction. During intervention visits, capillary blood was obtained by fingerprick test to determine glycaemic response and in parallel, by CGM to measure glycaemic response in IF. The effect of sbeII bread consumption on satiety was determined using a visual analogue scale questionnaire (VAS)28 before and after bread consumption as well as changes in energy intake during the subsequent meal (satiety challenge).
Between interventions, participants completed a non-consecutive three-day weighted food diary, on two weekdays and one weekend day, to capture habitual diet. The study lasted approximately one month from screening to follow up.
During Visit 1, participants applied two CGM Freestyle Libre sensors (Abbott Diabetes Care, Alameda, CA) to record IF glucose continuously for 14 days. They then observed a two-day CGM calibration period during which no interventions or restrictions were applied. Then, they completed Visit 2 and Visit 3 as follows: after a standardised evening meal and ∼12 h fast, baseline measurements of capillary blood glucose and satiety were taken while CGM sensors recorded IF glucose concentrations. They were then given the intervention meal as their first meal of the day (breakfast), either sbeII or WT control bread roll, after which glycaemic, sensory (palatability) and satiety responses (VAS) were measured over 3.5 h. Four hours after breakfast, a lunch meal was provided ad libitum as satiety challenge. At the end of the satiety challenge, the visit was considered completed and participants left the Clinical Research Facility of the Quadram Institute. A follow up questionnaire was completed the day after Visit 3 to capture participants’ experience wearing and using CGM devices.
The intervention breakfast consisted of one bread roll providing ∼75 g of total starch, either sbeII or WT control, 10 g of low-fat spread (Flora Dairy Free) and approximately 250 mL of water to drink. The liquid component of the breakfast meal was standardised by adjusting the water drink to the water content of each bread. The total water content each meal was approximately 317 g, the average serving size of sbeII breads was 153.19 g ± 0.49 g (fresh weight) and the average size of WT control breads was 147.28 g ± 0.73 g (fresh weight), (mean ± SEMs, n = 8).
The lunch meal (satiety challenge) served ad libitum was made of portions of 80 g white rice, served with either ‘Option A’ 85 g of tomato and basil pasta sauce and 7 g of parmesan cheese or ‘Option B’ 90 g of beef Bolognese pasta sauce, and was served with 250 mL of water. Both sauce options had a similar nutrient composition and provided approximately 177.2 kcal ± 0.7 kcal, 27.3 g ± 0.4 g carbohydrate of which, 4.6 g ± 0.9 g sugars, 1.2 g ± 0.2 g fibre, 6.9 g ± 0.8 g protein per serving (average of two options ± SD, calculated from the nutrient declaration on food packaging). Participants were asked in advance to choose from sauce option A and B and consumed the same lunch meal during both visits.
Data and statistical analysis
Baseline fasting glucose was obtained 15, 10 and 5 min before the breakfast intervention meal by FP and at 30, 15 and 0 min before the breakfast intervention meal by CGM. The average of these three measurements was used as T0.
Intervention meals were consumed within 20 min, followed by ten pricks to measure capillary blood glucose concentrations, every 15 min for 3.5 h while glucose concentration in IF was recorded continuously every 15 min by the two sensors worn by each participant. Data up to 3.5 h was collected in order to capture the end of the postprandial response (return to baseline) to these study foods.
The glycaemic response to breads (iAUC) was calculated geometrically between 0 and 210 min (primary outcome), by applying the trapezoid rule to the area over the fasting glucose baseline and under the curve, as recommended by Brouns et al.30 Average glucose peak and glucose dip were calculated as the maximum (peak) and minimum (dip) glucose concentrations achieved after each meal; for capillary glucose this was calculated between 15 and 210 min, for IF glucose, between 15 and 240 min. Time to peak was calculated as the time to reach the maximum glucose concentration following the meal.
Datasets were curated using package reshape v. 1.4.3, graphs and plots were made using ggplot2 v. 3.3.1 and ggpmisc v. 0.3.533–35 in Rstudio.
Results
Flour characteristics
The sbell and WT control flours had similar total starch (73.9 ± 1.2 g vs. 74.94 ± 0.8 g per 100 g of flour, means ± SEMs, n = 8). Apparent amylose (means ± SEMs, n = 3) was higher in sbell starch (39 ± 1.1% of total starch) than in WT control starch (25.7 ± 0.7% of total starch), hence the proportion of resistant starch (means ± SEMs, n = 8) was much higher in sbell flour (6.4 ± 0.5% of total starch) compared to WT control (0.6 ± 0.1% of total starch). Indicators of performance in the field and extraction rate can be found in ESI Table 2.†Bread characteristics
Breads were formulated to deliver approximately 75 g of total starch per serving. Nutrient composition, indicators of in vitro amylolysis and starch characteristics of bread rolls are reported in Table 1. Starch in sbeII bread was less susceptible to amylolysis compared to the WT control (C90 mean difference = 13.03% of starch digested, 95%CI [1.91, 24.15], independent groups t-test, p-value = 0.02). First order equations fit starch digestion curves well, Table 1. The sbeII bread was characterized by a lower starch breakdown after 90 min of hydrolysis and a lower digestion rate (k), Fig. 1A.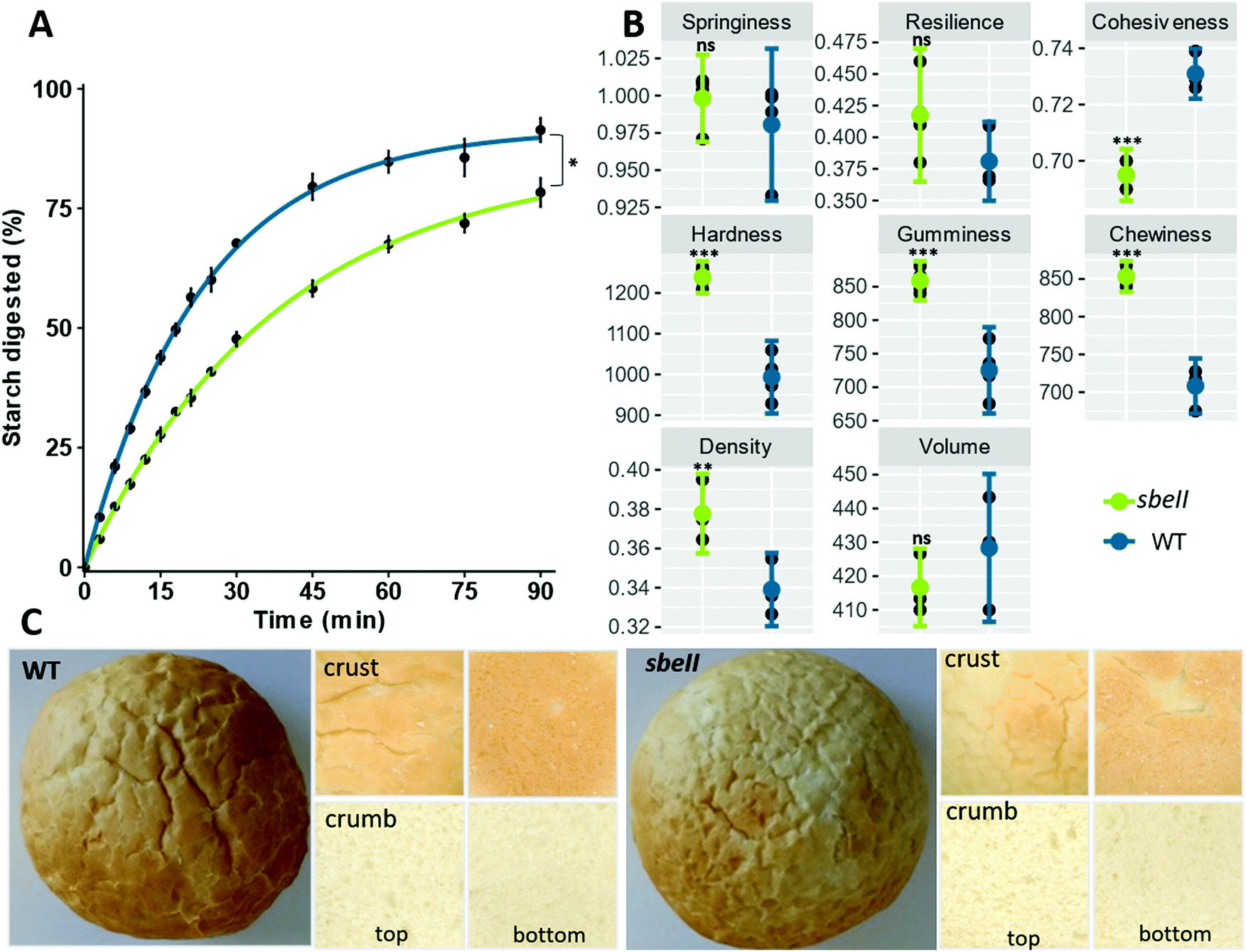 | ||
| Fig. 1 In vitro characteristics of sbeII (green) and WT (blue) bread rolls after 5 months of storage at −20 °C. (A) Starch amylolysis; each experimental data point represents the mean value from the analysis of five independent samples with error bars = ± SEMs, n = 5. Experimental data are shown by fitting a first-order equation based on the k and C∞ values obtained from a non-linear regression model. (B) Bread quality parameters, mean ± SEMs, n = 4; bread crumb texture analysis on Texture Analyser showing springiness, cohesiveness, resilience (%), and hardness (Newtons), gumminess (AU) chewiness (Newtons per s); bread volume and density (bulk density). (C) Images of bread rolls as whole, crust and crumb. | ||
| Nutrients per serving | WT control bread meal | sbeII bread meal |
|---|---|---|
| Energya (kcal) | 438.7 | 430.8 |
| Energy (kJ) | 1854.9 | 1815.7 |
| Proteinb (g) | 12.15 | 12.96 |
| Total fatc (g) | 11 | 11.54 |
| Saturated fat (g) | 2.92 | 3.07 |
| Poly-unsaturated fat (g) | 3.54 | 3.77 |
| Ash | 2.19 | 2.29 |
| Available carbohydratesd (g) | 70.9 | 65.1 |
| Total sugars (g) | 2.92 | 3.06 |
| Fibre AOACe (g) | 4.53 | 7.5 |
| Bread characteristics | WT bread | sbeII bread |
|---|---|---|
| Bread roll (g) | 146.24 ± 2.55 | 153.24 ± 0.38*** |
| Moisture (%) | 38.59 ± 0.17 | 42.13 ± 1.02*** |
| Total starch (g) | 74.9 | 74.9 |
| Digestible starch (%) | 96.5 ± 0.2 | 91.8 ± 0.8*** |
| Resistant starch (%) | 3.5 ± 0.1 | 8.2 ± 0.2*** |
| In vitro amylolysis | WT bread | sbeII bread |
|---|---|---|
| Bread roll weight is reported as the mean ± SEMs of 8 rolls measured after 16 h thaw. All other bread characteristics were measured on the same bread rolls, after 16 h thaw. Moisture and in vitro amylolysis experimental values are reported as the mean ± SEMs of 5 bread rolls; digestible and resistant proportion of starch are reported as mean ± SEMs, 4 bread rolls. Total starch content of breads is based on the flour starch content. The bread formulation was adjusted to match total starch content of sbeII and WT control breads. Parameter values that are significantly different in the sbeII bread compared to the WT control bread are indicated with an asterisk; *P < 0.05, **P < 0.01 and ***P < 0.001 (independent groups t-test).a Calculated using standard energy conversion factors (EC 2008/100 and 90/496).b Nitrogen-to-protein conversion factor = 6.25.c Determined by NMR.d Determined by ion exchange chromatography, calculated by difference.e AOAC method 985.29 (‘Prosky method’), which includes RS type 3 (retrograded starch). | ||
| C 90 (%) | 91.3 ± 2.9 | 78.3 ± 3.7* |
| C ∞ (%) | 91.6 | 85.4 |
| k (min−1) | 0.048 | 0.030 |
| r 2 | 0.99 | 0.99 |
| Sum of squares | 1706 | 767 |
Hardness, gumminess, chewiness and cohesiveness were significantly higher in sbeII bread rolls compared to WT control breads while there were no differences in springiness and in resilience between bread types (Fig. 1B.1 and B.2). The bulk density of sbeII bread was greater than the WT control but no differences in volume were detected (Fig. 1B.3 and B.4). Crust colour varied between sbeII and WT breads with a darker colour and a matte finish for the sbeII bread crust, compared to the WT control (Fig. 1C), ESI Table 3.†
Acute postprandial intervention study
Only 8 out of 19 enrolled participants completed the study due to the COVID-19 outbreak and consequent lockdown (Fig. 2). Their characteristics are reported in Table 2. All other baseline measurements are reported in ESI Table 4.† The average energy intake recorded by participants during the three-day food diary was within the UK dietary recommendations38 and their carbohydrate intake was approximately 41% of their total kcal intake (men and women), predominantly derived from starch Table 2. More details about the average dietary intake of study participants can be found in ESI Table 5.† | ||
| Fig. 2 CONSORT diagram of the REST study. Interventions were blinded to the participants and the study team using an allocation concealment. At the end of recruitment, breads were grouped as ‘A’ or ‘B’ to allow the study team to complete the data analysis. Researchers were unblinded once the data analysis was completed. Two participants lost one sensor after intervention 1 and completed intervention 2 with one sensor only. Two participants lost one sensor before intervention 1 and had it replaced. | ||
| Characteristics | Mean | SD | Min | Max |
|---|---|---|---|---|
| Age (years) | 33 | 13.6 | 23 | 58 |
| Height (m) | 1.73 | 8.6 | 1.60 | 1.90 |
| Weight (kg) | 69.8 | 10.4 | 58.1 | 91.9 |
| BMI (kg m−2) | 23.2 | 1.7 | 20.7 | 25.4 |
| Venous fasting glucose (mmoL L−1) | 4.4 | 0.2 | 4.1 | 4.7 |
| HbA1c (mmol mol−1) | 34.4 | 2.0 | 31.0 | 36.0 |
| Three-days average intake | Men | Women |
|---|---|---|
| N = 8 of which 4 males and 4 females. Diet intake is reported as mean ± SDs, n = 4 men and n = 4 women. | ||
| Energy (kcal per day) | 2780.5 ± 735.9 | 1989 ± 430.9 |
| Starch (g per day) | 187.6 ± 49 | 136.8 g ± 39.8 |
| Non-starch polysaccharide (g per day) | 24.9 g ± 3.3 | 18.7 g ± 3.1 |
| iAUC (mmoL L−1 min−1) | WT control bread | sbeII bread | Treatment effect |
|---|---|---|---|
| FP0–120 | 140 [109.1, 172] | 118 [86.2, 149] | −22.9 [−59.5, 13.6] |
| CGM0–120 | 128 [93.5, 162] | 109 [74.5, 143] | −19.2 [−67.5, 29.1] |
| FP0–210 | 182 [146, 219] | 157 [121, 194] | −25.1 [−82.3, 32.2] |
| CGM0–210 | 136 [80.3, 191] | 103 [47.5, 158] | −32.9 [−111, 45.1] |
| Bread type | Method | Fasting glucose (mmoL L−1) | Glucose peak (mmoL L−1) | Glucose dip (mmoL L−1) | Time to peak (min) | Time to baseline (min) |
|---|---|---|---|---|---|---|
| iAUC0–120 is the primary outcome measure of this study as this measure has been shown to correlate well with in vitro starch amylolysis C90 indicator. iAUC0–210 is reported here as glucose curves returned to the fasting glucose baseline approximately 210 min from the start of the test. iAUC mean ± 95% CI, n = 8. Glucose values and time to peak are reported as mean ± SEMs, n = 8. ‘peak’ and ‘dip’ are the maximum and minimum glucose concentration achieved after consuming the meal (capillary glucose 15–210 min, IF glucose 15–240 min). Time to baseline is reported as mean ± SEMs, n = 6 for FP, as the glucose response of two participants did not return to the fasting value within 210 min of test, and n = 7 participants whose glucose response returned to the fasting value within 240 min of test. | ||||||
| sbeII | FP | 5.1 ± 0.06 | 7.1 ± 0.25 | 4.9 ± 0.08 | 39.4 ± 3.9 | 185 ± 9.2 |
| CGM | 4.7 ± 0.05 | 6.4 ± 0.29 | 4.5 ± 0.16 | 53.0 ± 2.9 | 210 ± 12.6 | |
| WT control | FP | 5.03 ± 0.07 | 7.3 ± 0.24 | 4.8 ± 0.14 | 67.5 ± 19.4 | 175 ± 12 |
| CGM | 4.6 ± 0.1 | 6.7 ± 0.36 | 4.2 ± 0.25 | 49.3 ± 1.9 | 184 ± 10.8 | |
Glycaemic response (iAUC) to sbell bread was 22.9 mmoL L−1 min−1, 95% CI [−13.6, 59.5], lower than control bread (corresponding to an average 15.7% reduction following sbeII bread compared to WT control bread). Similarly, the effect size measured in IF by CGM was 19.2 mmoL L−1 min−1 (14.8% difference between sbeII and WT control breads). While these estimates from only 8 participants are not statistically significant, they are consistent with the in vitro findings and our anticipated effect size of 20% difference between groups. Participants’ fasting glucose concentrations measured in capillary blood and IF were within normal range and their fasting glucose values did not vary significantly between the two visits (FP fasting glucose p-value = 0.58, IF fasting glucose p-value = 0.62, paired t-test). The overall pattern of postprandial capillary blood glucose was similar after the consumption of sbeII bread and WT control (Fig. 3A and B). Fig. 3C shows the incremental glucose peak (iCmax) and Fig. 3D shows the average capillary glucose responses (iAUC0–120). Other postprandial capillary glucose indicators are reported in Table 3. Time to peak calculation from the start of the test showed that the IF measurements lagged 11 min compared to capillary blood glucose.
 | ||
| Fig. 3 Post-prandial glucose concentrations. Incremental glucose concentrations measured in capillary blood (A) and IF (B). Datapoints represent individual glucose values, curves are mean glucose profile (n = 8) and error bars = ± SEMs. C. Incremental maximum concentration of glucose (iCmax) in capillary and IF. (D) Individual incremental glucose over time detected in capillary blood and IF (iAUC0–120). (E) Change in appetite score from fasting until after the ad libitum lunch (satiety challenge), score calculated as [desire to eat + hunger + (100 − fullness) + prospective consumption]/4, n = 8 and error bars = ± SEMs. (F) Energy and water intake during satiety challenge following the intervention meals (n = 8). | ||
Intra-individual variation (CV%) between the two sensors used in this study by each participant ranged 0–17% during postprandial breakfast phase and 0–22.7% during the postprandial lunch phase. There was a strong correlation between the two CGM sensors worn by each participant (r = 0.91); on average, there was a 10.5 mmol L−1 min−1 difference between sensors’ measurements of glycaemic response (iAUC 0–120) within participant. We observed a 5.25 mmol L−1 min−1, 95% CI [−11.3, 21.8] difference between sensors after consuming sbeII bread (n = 8) and 17.6 mmol L−1 min−1, 95% CI [−5.73, 40.9] after WT bread (n = 6). Glucose response curves are shown in ESI Fig. 3.†
Overall, the glucose response measured by FP and by CGM was similar (iAUC and iCmax), Fig. 4.
 | ||
| Fig. 4 iAUC and iCmax measured by CGM in IF and in capillary blood by FP after consuming sbeII and WT control bread, respectively. Data points represent individual responses (black), the mean of eight subjects in grey with error bars = ± SEMs. | ||
Discussion
This study investigated the use of a sbeII mutant wheat with a higher amylose proportion for production of slowly digested and/or low glycaemic bread products, compared to conventional wheat starch. Bread rolls made from sbeII mutant wheat contained more RS and showed lower susceptibility to amylase digestion that the WT control, which is consistent with the hypothesis that the presence of mutations in sbeII genes lead to wheat foods with increased starch resistance to digestion.14Previous studies measuring glycaemic potency of high-amylose food products reported conflicting results. Often, this is derived from inconsistent carbohydrate content across test meals; Behall et al. in 200239 presented a very good overview of the different studies that compared glycaemic responses to high and low amylose food products and showed that different methods used to determine the RS and dietary fibre can contribute to the variability in glycaemic measurements. In this study, we report amylose and RS portion of starch in bread rolls, as well as starch resistance to digestion measured using a single-enzyme system that allowed us to determine the rate and extent of starch digestion, previously shown to correlate with glycaemic index measured in vivo.22
Here, we showed that the predicted difference in glycaemic response between sbeII and WT control breads from in vitro amylolysis experiments is consistent with the observed difference in glucose concentrations after consumption of sbeII and WT control breads, although our estimate of the effect in vivo is imprecise because only around one third of the planned sample could be completed.
Studies using novel wheat genotypes characterized by a high amylose content have consistently reported a lowering effect on postprandial glucose response. Previously, Belobrajdic et al.16 reported a lower glycaemic response to high amylose wheat bread measured in plasma glucose. The low glycaemic effect was likely due to the reduced amount of carbohydrate provided in the high-amylose bread compared to the control bread. In this study, we showed that sbeII bread has potential to elicit lower glycaemic response compared to a conventional white bread without changing the amount of starch provided.
Hallström et al. (2011)43 showed that bread made from a high-amylose wheat (38% of starch) induced 24% lower glycaemic responses than a control white bread (high-amylose bread iAUC 0–120 min = 185 ± 29.7 mmoL L−1 min−1, white control bread iAUC 0–120 min = 224 ± 27.3 mmoL L−1 min−1, mean ± SEM, n = 14) although all bread types in their study were baked at pumpernickel conditions (20 hours, 120 °C), which may have modulated starch resistance to enzymatic digestion. In the present study, the difference in starch susceptibility to amylase digestion between sbeII and WT control breads was observed using a shorter baking time (15 min) at higher temperature (185 °C) to produce white rolls, resembling the Chorleywood baking process.
It is noteworthy that these breads were stored at −20 °C for ∼2 months longer than those consumed by the human study participants. There is still a limited understanding of the effect of freeze–thaw treatment on starch digestibility in bread and glycaemia, however the largest effects of cold storage occur during the initial stages of freezing40 and further starch retrogradation effects thereafter would likely be minimal and negligible with regard to influencing postprandial glycaemia.41
A study by Sisson et al. in 202015 showed that pasta made from sbeIIa mutant semolina (with 58% amylose starch) produced pasta with lower glycaemic index (compared to an isoglucidic WT control pasta) and acceptable quality, although the authors did report slightly reduced firmness, increased cooking loss and inferior colour compared to the control, likely to be linked to the large increase in amylose. The sbeII bread used in the present study presented minor differences in texture compared to the WT control but was deemed overall acceptable by participants.
The results presented in this study showed that the effect of bread on glycaemic response (iAUCs) were measured similarly in IF by CGM on the upper arm and in capillary blood by FP, therefore the conclusions based on iAUCs does not differ between the metrics. Measuring glucose responses in IF by CGM was found to be an effective and minimally invasive method to carry out intervention studies: sensor measurements are reliably taken every 15 min (this may vary depending on the type of sensor), allowing studies to collect larger glucose datasets compared to capillary glucose measured by FP, which is also more painful for study participants. Previous studies have shown a good agreement between measurements taken in blood and IF42 however, the IF measurements can vary depending on the type of device and the insertion site.18
Based on the results collected from eight participants, the effect size measured in this study in IF was consistent with the expected effect size based on in vitro amylolysis. The study was underpowered due to the COVID-19 outbreak that forced us to stop all study activities ahead of time. Nevertheless, the results from the human study suggested that bread made with sbeII mutant wheat may lead to lower postprandial IF glycaemic responses.
Overall, this study showed that the small increases in amylose content in the sbeII mutant wheat raw material can be used to produce sbeII wheat-based foods with lower starch susceptibility to hydrolysis, without substantially compromising (bread) texture or appearance, measured instrumentally. Future foods made from sbeII mutant wheat may provide an alternative to high glycaemic wheat-based staple foods, including white bread. Further acute and chronic intervention studies are needed to fully explore the potential use of sbeII mutant wheat to reduce the glycaemic potency of bread and other wheat-based foods.
Abbreviations
| IF | Interstitial fluid |
| FP | Fingerprick |
| CGM | Continuous glucose monitoring |
| sbeII | Starch branching enzyme II |
| WT | Wild-type |
| RS | Resistant starch |
| iAUC | Incremental area under the curve |
Authors contribution
Marina Corrado: Conceptualization, investigation, methodology, project administration, data curation and formal analysis, writing – original draft preparation, writing – review & editing, Jennifer Ahn-Jarvis: investigation, methodology, writing – review & editing, Brendan Fahy: investigation, writing – review & editing, George M. Savva: methodology, data curation, formal analysis, Cathrina H. Edwards: conceptualization, methodology, supervision, writing – review & editing, Brittany A. Hazard: conceptualization, funding acquisition, methodology, supervision, writing – review & editing.Conflicts of interest
The authors declare no conflicts of interest.Acknowledgements
We thank Dr Natalia Perez-Moral for blinding and unblinding the intervention meals. This project was funded by the UKRI Biotechnology and Biological Sciences Research Council (BBSRC) Norwich Research Park Biosciences Doctoral Training Partnership grant number BB/M011216/1 and the BBSRC Institute Strategic Programme ‘Food Innovation and Health’ grant number BB/R012512/1 and its constituent projects BBS/E/F/000PR10343 (Theme 1, Food Innovation) and BBS/E/F/000PR10345 (Theme 2, Digestion in the Upper GI Tract) and the BBSRC Core Capability Grant BB/CCG1860/1 awarded to Quadram Institute Bioscience, and previously by the Institute Strategic Programme Grant ‘Food and Health’ Grant No. BB/J004545/1 awarded to the Institute of Food Research; the BBSRC Institute Strategic Programme Grants ‘Molecules from Nature’ – Crop Quality BBS/E/J/000PR9799 awarded to the John Innes Centre and ‘Designing Future Wheat’ BB/P016855/1 awarded to the John Innes Centre.References
- D. M. Nathan, M. B. Davidson, R. A. DeFronzo, R. J. Heine, R. R. Henry, R. Pratley and B. Zinman, Impaired Fasting Glucose and Impaired Glucose Tolerance, Implications for Care, 2007, 30, 753–759 CAS.
- F. de Vegt, J. M. Dekker, A. Jager, E. Hienkens, P. J. Kostense, C. D. A. Stehouwer, G. Nijpels, L. M. Bouter and R. J. Heine, Relation of Impaired Fasting and Postload Glucose With Incident Type 2 Diabetes in a Dutch PopulationThe Hoorn Study, J. Am. Med. Assoc., 2001, 285, 2109–2113 CrossRef CAS.
- C. A. Whicher, S. O'Neill and R. I. G. Holt, Diabetes in the UK: 2019, Diabetic Med., 2020, 37, 242–247 CrossRef CAS PubMed.
- Public Health England, National Diet and Nutrition Survey 2016/2017 to 2018/2019, 2020.
- E. E. Blaak, J.-M. Antoine, D. Benton, I. Björck, L. Bozzetto, F. Brouns, M. Diamant, L. Dye, T. Hulshof, J. J. Holst, D. J. Lamport, M. Laville, C. L. Lawton, A. Meheust, A. Nilson, S. Normand, A. A. Rivellese, S. Theis, S. S. Torekov, S. Vinoy and E. Aisbl, Impact of postprandial glycaemia on health and prevention of disease, Obes. Rev., 2012, 13, 923–984 CrossRef CAS PubMed.
- T. M. S. Wolever and C. Mehling, High-carbohydrate–low-glycaemic index dietary advice improves glucose disposition index in subjects with impaired glucose tolerance, Br. J. Nutr., 2002, 87, 477–487 CrossRef CAS.
- D. J. Jenkins, C. W. Kendall, L. S. Augustin, S. Franceschi, M. Hamidi, A. Marchie, A. L. Jenkins and M. Axelsen, Glycemic index: overview of implications in health and disease, Am. J. Clin. Nutr., 2002, 76, 266s–273s CrossRef CAS.
- C. M. McCallum, L. Comai, E. A. Greene and S. Henikoff, Targeting induced local lesions IN genomes (TILLING) for plant functional genomics, Plant Physiol., 2000, 123, 439–442 CrossRef CAS PubMed.
- K. V. Krasileva, H. A. Vasquez-Gross, T. Howell, P. Bailey, F. Paraiso, L. Clissold, J. Simmonds, R. H. Ramirez-Gonzalez, X. Wang, P. Borrill, C. Fosker, S. Ayling, A. L. Phillips, C. Uauy and J. Dubcovsky, Uncovering hidden variation in polyploid wheat, Proceedings of the National Academy of Sciences, 2017, 114, E913–E921 CrossRef CAS PubMed.
- B. Hazard, X. Zhang, P. Colasuonno, C. Uauy, D. M. Beckles and J. Dubcovsky, Induced mutations in the starch branching enzyme II (SBEII) genes increase amylose and resistant starch content in durum wheat, Crop Sci., 2012, 52, 1754–1766 CAS.
- A. Schönhofen, B. Hazard, X. Zhang and J. Dubcovsky, Registration of Common Wheat Germplasm with Mutations in SBEII Genes Conferring Increased Grain Amylose and Resistant Starch Content, J. Plant Regist., 2016, 10, 200–205 CrossRef PubMed.
- L. M. N. K. Ekström, I. M. E. Björck and E. M. Östman, An improved course of glycaemia after a bread based breakfast is associated with beneficial effects on acute and semi-acute markers of appetite, Food Funct., 2016, 7, 1040–1047 RSC.
- L. M. N. K. Ekström, I. M. E. Björck and E. M. Ostman, On the possibility to affect the course of glycaemia, insulinaemia, and perceived hunger/satiety to bread meals in healthy volunteers, Food Funct., 2013, 4, 495–660 RSC.
- M. Corrado, A. Cherta-Murillo, E. S. Chambers, A. J. Wood, A. Plummer, A. Lovegrove, C. H. Edwards, G. S. Frost and B. A. Hazard, Effect of semolina pudding prepared from starch branching enzyme IIa and b mutant wheat on glycaemic response in vitro and in vivo: a randomised controlled pilot study, Food Funct., 2020, 11, 617–627 RSC.
- M. Sissons, F. Sestili, E. Botticella, S. Masci and D. Lafiandra, Can Manipulation of Durum Wheat Amylose Content Reduce the Glycaemic Index of Spaghetti?, Foods, 2020, 9, 693 CrossRef CAS PubMed.
- D. P. Belobrajdic, A. Regina, B. Klingner, I. Zajac, S. Chapron, P. Berbezy and A. R. Bird, High-Amylose Wheat Lowers the Postprandial Glycemic Response to Bread in Healthy Adults: A Randomized Controlled Crossover Trial, J. Nutr., 2019, 149, 1335–1345 CrossRef PubMed.
- S. E. Berry, A. M. Valdes, D. A. Drew, F. Asnicar, M. Mazidi, J. Wolf, J. Capdevila, G. Hadjigeorgiou, R. Davies, H. Al Khatib, C. Bonnett, S. Ganesh, E. Bakker, D. Hart, M. Mangino, J. Merino, I. Linenberg, P. Wyatt, J. M. Ordovas, C. D. Gardner, L. M. Delahanty, A. T. Chan, N. Segata, P. W. Franks and T. D. Spector, Human postprandial responses to food and potential for precision nutrition, Nat. Med., 2020, 26(6), 964–973 CrossRef CAS PubMed.
- R. Howard, J. Guo and K. D. Hall, Imprecision nutrition? Different simultaneous continuous glucose monitors provide discordant meal rankings for incremental postprandial glucose in subjects without diabetes, Am. J. Clin. Nutr., 2020, 112, 1114–1119 CrossRef PubMed.
- L. Dye, M. Mansfield, N. Lasikiewicz, L. Mahawish, R. Schnell, D. Talbot, H. Chauhan, F. Croden and C. Lawton, Correspondence of continuous interstitial glucose measurement against arterialised and capillary glucose following an oral glucose tolerance test in healthy volunteers, Br. J. Nutr., 2010, 103, 134–140 CrossRef CAS PubMed.
- B. H. Bajka, A. M. Pinto, J. Ahn-Jarvis, P. Ryden, N. Perez-Moral, A. van der Schoot, C. Stocchi, C. Bland, S. E. Berry, P. R. Ellis and C. H. Edwards, The impact of replacing wheat flour with cellular legume powder on starch bioaccessibility, glycaemic response and bread roll quality: A double-blind randomised controlled trial in healthy participants, Food Hydrocolloids, 2021, 114, 106565 CrossRef CAS PubMed.
- C. H. Edwards, M. M. Grundy, T. Grassby, D. Vasilopoulou, G. S. Frost, P. J. Butterworth, S. E. Berry, J. Sanderson and P. R. Ellis, Manipulation of starch bioaccessibility in wheat endosperm to regulate starch digestion, postprandial glycemia, insulinemia, and gut hormone responses: a randomized controlled trial in healthy ileostomy participants, Am. J. Clin. Nutr., 2015, 102, 791–800 CrossRef CAS.
- C. H. Edwards, N. Cochetel, L. Setterfield, N. Perez-Moral and F. J. Warren, A single-enzyme system for starch digestibility screening and its relevance to understanding and predicting the glycaemic index of food products, Food Funct., 2019, 10, 4751–4760 RSC.
- C. H. Edwards, F. J. Warren, P. J. Milligan, P. J. Butterworth and P. R. Ellis, A novel method for classifying starch digestion by modelling the amylolysis of plant foods using first - order enzyme kinetic principles, Food Funct., 2014, 5, 2751–2758 RSC.
- R Development Core Team, R: A language and environment for statistical computing, R Foundation for Statistical Computing, 2008 Search PubMed.
- RStudio Team, RStudio: Integrated Development for R, R Foundation for Statistical Computing, 2019 Search PubMed.
- C. S. McCamy, H. Marcus and J. G. Davidson, A color-rendition chart, J. Appl. Photogr. Eng., 1976, 2, 95–99 Search PubMed.
- M. Maskan, Kinetics of colour change of kiwifruits during hot air and microwave drying, J. Food Eng., 2001, 48, 169–175 CrossRef.
- C. Byrne, D. Blunt, J. Burn, E. Chambers, A. Dagbasi, G. Franco Becker, G. Gibson, L. Mendoza, K. Murphy, C. Poveda, A. Ramgulam, M. Tashkova, G. Walton, C. Washirasaksiri and G. Frost, A study protocol for a randomised crossover study evaluating the effect of diets differing in carbohydrate quality on ileal content and appetite regulation in healthy humans, F1000Research, 2019, 8, 258 CAS.
- L. A. H. Rosén, L. O. B. Silva, U. K. Andersson, C. Holm, E. M. Östman and I. M. Björck, Endosperm and whole grain rye breads are characterized by low post-prandial insulin response and a beneficial blood glucose profile, Nutr. J., 2009, 8, 1–11 CrossRef PubMed.
- F. Brouns, I. Bjorck, K. N. Frayn, A. L. Gibbs, V. Lang, G. Slama and T. M. S. Wolever, Glycaemic index methodology, Nutr. Res. Rev., 2005, 18, 145–171 CrossRef CAS.
- A. Kuznetsova, P. B. Brockhoff and R. H. B. Christensen, lmerTest Package: Tests in Linear Mixed Effects Models, J. Stat. Softw., 2017, 82, 1–16 Search PubMed.
- R. Lenth, emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.4.5, 2020 Search PubMed.
- H. Wickham, Reshaping Data with the reshape Package, J. Stat. Softw., 2007, 21, 1–20 Search PubMed.
- H. Wickham, ggplot2: Elegant Graphics for Data Analysis, Springer-Verlag New York, 2016 Search PubMed.
- P. J. Aphalo, ggpmisc: Miscellaneous Extensions to ‘ggplot2′, 2021 Search PubMed.
- C. A. Schneider, W. S. Rasband and K. W. Eliceiri, NIH Image to ImageJ: 25 years of image analysis, Nat. Methods, 2012, 9, 671–675 CrossRef CAS PubMed.
- G. H. Anderson, C. E. Cho, T. Akhavan, R. C. Mollard, B. L. Luhovyy and E. T. Finocchiaro, Relation between estimates of cornstarch digestibility by the Englyst in vitro method and glycemic response, subjective appetite, and short-term food intake in young men, Am. J. Clin. Nutr., 2010, 91, 932–939 CrossRef CAS PubMed.
- Scientific Advisory Committee on Nutrition, Dietary reference values for energy, The Stationery Office, 2011 Search PubMed.
- K. M. Behall and J. Hallfrisch, Plasma glucose and insulin reduction after consumption of breads varying in amylose content, Eur. J. Clin. Nutr., 2002, 56, 913–920 CrossRef CAS PubMed.
- H. Patel, P. G. Royall, S. Gaisford, G. R. Williams, C. H. Edwards, F. J. Warren, B. M. Flanagan, P. R. Ellis and P. J. Butterworth, Structural and enzyme kinetic studies of retrograded starch: Inhibition of α-amylase and consequences for intestinal digestion of starch, Carbohydr. Polym., 2017, 164, 154–161 CrossRef CAS PubMed.
- C. H. Edwards, A. S. Veerabahu, A. J. Mason, P. J. Butterworth and P. R. Ellis, α-Amylase action on starch in chickpea flour following hydrothermal processing and different drying, cooling and storage conditions, Carbohydr. Polym., 2021, 259, 117738 CrossRef CAS PubMed.
- N. Peled, D. Wong and S. L. Gwalani, Comparison of glucose levels in capillary blood samples obtained from a variety of body sites, Diabetes Technol. Ther., 2002, 4, 35–44 CrossRef CAS PubMed ; discussion 45–37.
- E. Hallström, F. Sestili, D. Lafiandra, I. Björck and E. Östman, A novel wheat variety with elevated content of amylose increases resistant starch formation and may beneficially influence glycaemia in healthy subjects, Food Nutr. Res., 2011, 55, 1–8 CrossRef.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1fo03085j |
| ‡ These authors contributed equally. |
| This journal is © The Royal Society of Chemistry 2022 |